Embed Size (px)
Citation preview

B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 7 1 – 8 2
ava i l ab l e a t www.sc i enced i rec t . com
www.e l sev i e r. com/ l oca te /b ra in res
Research Report
Expression of sex-steroid receptors and steroidogenicenzymes in the carotid body of adult and newborn male rats
Vincent Joseph⁎, Van Diep Doan, Catherine-Eve Morency, Yves Lajeunesse, Aida BairamDepartment of Pediatrics, Laval University, Centre de Recherche (D0-711), Hôpital St-François d'Assise,10 rue de l'Espinay, Quebec (QC), Canada G1L 3L5
A R T I C L E I N F O
⁎ Corresponding author. Fax: +1 418 525 4195E-mail address: [email protected]
0006-8993/$ – see front matter © 2005 Elsevidoi:10.1016/j.brainres.2005.12.075
A B S T R A C T
Article history:Accepted 15 December 2005Available online 27 January 2006
This study describes the localization and pattern of expression of estradiol and progesteronereceptors as well as key enzymes for steroid synthesis (i.e. P450 side-chain-cleavage—P450scc, and P450 aromatase—P450Aro) in the carotid body (CB) and superior cervicalganglion (SCG) of adult, newborn and late fetal male rats, using immunohistochemistry,Western blot and real-time RT-PCR. Our results show a constitutive expression of the βestradiol receptor (Erβ) and the 80 kDa and 60 kDa progesterone receptors (PR-A and PR-C)isoforms in the CB, while in the SCG Erα, Erβ, PR-A and PR-C are expressed. While P450Arostaining was negative, P450scc staining was strong both in the SCG and CB. In late fetal andnewborn rats, Erα was not detected in the CB or SCG, but a slight staining appeared for P450aromatase in the CB, and to a lesser extent in SCG. P450scc was strongly expressed in CB andSCG of late fetal and newborn rats. We conclude that the carotid body shows a constitutiveexpression of Erβ and PR and may be able to synthesize steroids, including estradiol duringlate fetal life.
© 2005 Elsevier B.V. All rights reserved.
Keywords:Carotid bodyEstradiol receptorProgesterone receptorSteroidogenic enzymeAdult ratNewborn rat
Abbreviations:CB, carotid bodyErβ, estradiol receptor betaErα, estradiol receptor alphaP450scc, cytochrome P450 side-chain-cleavageP450Aro, cytochrome P450aromatasePRA, PRB and PRC, progesteronereceptor A, B and CSCG, superior cervical ganglionTH, tyrosine hydroxylase
1. Introduction
The peripheral chemoreceptors localized in the carotidbifurcation are the main oxygen sensors participating inrespiratory and sympathetic regulation (Gonzalez et al.,
.val.ca (V. Joseph).
er B.V. All rights reserved
1994). Their function is critical in various processes, includingthe maintenance of a stable respiratory rhythm (i.e. avoidingthe generation of apneas) (Nakayama et al., 2003; Smith et al.,2003; Chenuel et al., 2005) and supporting adequate respira-tory plasticity during chronic hypoxic exposure (Smith et al.,
.

72 B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 7 1 – 8 2
1986; Joseph and Pequignot, 2003). Ovarian steroids, includingestradiol and progesterone, have been recognized as majormodulators of carotid body activity, exerting an overallstimulatory effect on baseline activity and hypoxic chemo-responsiveness of the peripheral chemoreceptors (Hannhartet al., 1990; Joseph et al., 2002; Tatsumi et al., 1991, 1997). Thiseffect has significant implications for general respiratoryhomeostasis, including ventilatory adjustment during gesta-tion (Doan et al., 2004; Moore et al., 1987) or chronic hypoxicexposure (León-Velarde et al., 1997; León-Velarde et al., 2001),and is a protective factor against the occurrence of sleepapneas in adult women (Shahar et al., 2003; Saaresranta et al.,2003; Young et al., 2003).
We recently provided evidence supporting the hypothesisof a role for placental-derived steroids, such as estradiol andprogesterone, in the control of adequate carotid body devel-opment (Doan et al., 2004; Soliz and Joseph, 2005). Indeed, it islargely recognized that steroids act as multipotent develop-mental factors that exert specific effects on a variety of targettissue, including the reproductive tract (Okada et al., 2002) andcentral or peripheral nervous system (Ivanova et al., 2001,2002; Solum and Handa, 2002; Nilsen et al., 2000; Beyer et al.,2002). In the developing central or peripheral nervous system,estradiol can be provided either through maternal transfer ofplacental steroids or as the result of local synthesis in neuronsand/or glial cells that express the molecular machineryrequired for steroid synthesis (Zwain and Yen, 1999; Pezzi etal., 2003).
Despite the fact that ovarian steroids modulate theactivity of the peripheral chemoreceptors, there are nodetailed description of the type and localization of steroidreceptors in the carotid body. We thus performed a series ofstudies using immunohistochemistry, Western blot and real-time RT-PCR analysis in adult, late fetal (gestational day 21,i.e. 1 day before delivery) and newborn (postnatal days 0 and2) male rats. We assessed the expression of the mainestrogen (i.e. Erα and Erβ) and progesterone (i.e. PR-A, PR-Band PR-C) receptors and the enzymes involved in the firststep of steroid synthesis (cytochrome P450 side-chain-cleavage—P450scc) and estradiol synthesis (cytochromeP450 aromatase—P450Aro) in the carotid bodies and in thenearby superior cervical ganglion (SCG), a sympathetic relayharboring sympathetic noradrenergic cell bodies. Our resultsdemonstrate a constitutive expression of Erβ, PR-A, PR-C andP450scc in the carotid body and superior cervical ganglion ofadult male rats, clear immunostaining for Erβ, PR andP450scc in newborn and shows that the P450Aro enzyme isexpressed in the carotid body of late fetal rats, thussupporting the hypothesis of a wider role than previouslyrecognized for steroids, including local steroids synthesis, incarotid body of both newborn and adult rats.
2. Results
2.1. Ovarian steroid receptors and steroidogenic enzymesin the carotid body and superior cervical ganglion in adult rats
Fig. 1 shows immunohistochemical staining for Erβ, Erα andPR in the carotid body and superior cervical ganglion (SCG) of
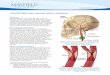
adult male rats at low (200×) and high (600×) magnification.The visual examination clearly shows that both PR and Erβ arepresent in SCG and in the carotid body, while Erα is present inthe SCG, but not in the carotid body (the staining was neverclearly distinct from background obtained after omission ofthe primary antibody). High-resolution pictures show that Erβimmunoreactivity is localized in islets of densely packed cellswith round nuclei (i.e. type I cells) but not in surroundingtissues, and the staining was apparently absent from type IIcells (flattened cells with long nuclei located at the peripheryof type I cell islets—see Fig. 1). PR was also mainly concen-trated in type I cell islets. In the SCG, Erα, Erβ and PRimmunoreactivity was localized in large neurons, and doublefluorescent labeling confirmed that Erβ is localized in THimmunoreactive cells in the carotid body and in the SCG (Fig.2A), while Erα is also localized in TH+ cells in the SCG, but notin the carotid body (Fig. 2B).
P450 aromatase staining was not detected in the carotidbody or superior cervical ganglion in adult male rats, but astrong staining appeared for the P450 side-chain-cleavageenzyme (Fig. 3).
While the antibodies used for estrogen receptors werespecifically directed against each isoform (i.e. Erα or Erβ), theantibody for progesterone receptor recognizes the three mainisoforms of the receptor (PR-A, PR-B and PR-C). To differentiatebetween these isoforms, we performed Western blot analysisof PR in protein extracted from a pool of CB and SCG of adultmale rats and from the uterus of an adult female as a positivecontrol tissue. The Western blot showed that the 80 kDa and60 kDa PR (PR-A and PR-C) are present, while the heavier PR(PR-B) isoform is not detectable in CB and SCG (Fig. 4A). Afteranalysis of the optical density of the bands and normalizationto the optical density obtained for actin, the relative expres-sion of PR was about 1.2 times higher in CB than in SCG (PR-A)and 2.8 for PR-C, while the expression of PR-A was 10 timeshigher in the CB than in the uterus (4 times higher for PR-C—Fig. 4B).
To further characterize the expression of ovarian steroidreceptors at the mRNA level, we performed real-time RT-PCRon mRNA extracted from a pool of carotid body and SCG ofadult male rats, with primers specifically designed to amplifyErα, Erβ, PR and tyrosine hydroxylase as a positive control foreach tissue. The results show a clear expression of Erα, Erβ,PR and TH mRNAs both in the carotid body and SCGconfirmed by gel analysis of the real-time RT-PCR products(Fig. 5). In the carotid body, Erβ was below the detectionthreshold for the gel analysis but was clearly detected by theSYBR green staining during real-time RT-PCR (inserts inFig. 5B).
2.2. Ovarian steroid receptors and steroidogenic enzymesin the carotid body and superior cervical ganglion in newbornand late fetal rats
Immunohistochemical analysis of the carotid artery bifurca-tion was performed in male rat pups at the age of 21gestational day, on the day of birth (P0) and 2 days followingbirth for Erα, Erβ and PR to assess the potential developmentalregulation of hormone receptors and the potential expressionof steroidogenic enzymes (P450scc and P450Aro).
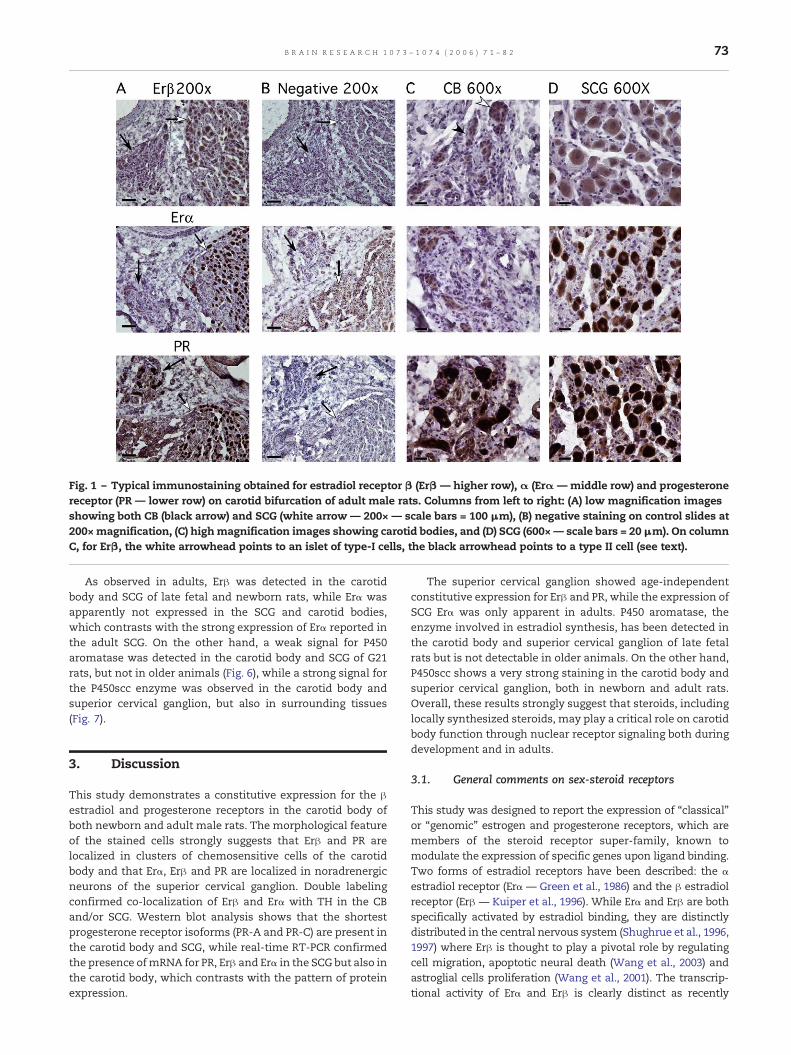
Fig. 1 – Typical immunostaining obtained for estradiol receptor β (Erβ — higher row), α (Erα — middle row) and progesteronereceptor (PR — lower row) on carotid bifurcation of adult male rats. Columns from left to right: (A) low magnification imagesshowing both CB (black arrow) and SCG (white arrow — 200× — scale bars = 100 μm), (B) negative staining on control slides at200×magnification, (C) highmagnification images showing carotid bodies, and (D) SCG (600×— scale bars = 20μm). On columnC, for Erβ, the white arrowhead points to an islet of type-I cells, the black arrowhead points to a type II cell (see text).
73B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 7 1 – 8 2
As observed in adults, Erβ was detected in the carotidbody and SCG of late fetal and newborn rats, while Erα wasapparently not expressed in the SCG and carotid bodies,which contrasts with the strong expression of Erα reported inthe adult SCG. On the other hand, a weak signal for P450aromatase was detected in the carotid body and SCG of G21rats, but not in older animals (Fig. 6), while a strong signal forthe P450scc enzyme was observed in the carotid body andsuperior cervical ganglion, but also in surrounding tissues(Fig. 7).
3. Discussion
This study demonstrates a constitutive expression for the βestradiol and progesterone receptors in the carotid body ofboth newborn and adult male rats. The morphological featureof the stained cells strongly suggests that Erβ and PR arelocalized in clusters of chemosensitive cells of the carotidbody and that Erα, Erβ and PR are localized in noradrenergicneurons of the superior cervical ganglion. Double labelingconfirmed co-localization of Erβ and Erα with TH in the CBand/or SCG. Western blot analysis shows that the shortestprogesterone receptor isoforms (PR-A and PR-C) are present inthe carotid body and SCG, while real-time RT-PCR confirmedthe presence ofmRNA for PR, Erβ and Erα in the SCG but also inthe carotid body, which contrasts with the pattern of proteinexpression.
The superior cervical ganglion showed age-independentconstitutive expression for Erβ and PR, while the expression ofSCG Erα was only apparent in adults. P450 aromatase, theenzyme involved in estradiol synthesis, has been detected inthe carotid body and superior cervical ganglion of late fetalrats but is not detectable in older animals. On the other hand,P450scc shows a very strong staining in the carotid body andsuperior cervical ganglion, both in newborn and adult rats.Overall, these results strongly suggest that steroids, includinglocally synthesized steroids, may play a critical role on carotidbody function through nuclear receptor signaling both duringdevelopment and in adults.
3.1. General comments on sex-steroid receptors
This study was designed to report the expression of “classical”or “genomic” estrogen and progesterone receptors, which aremembers of the steroid receptor super-family, known tomodulate the expression of specific genes upon ligand binding.Two forms of estradiol receptors have been described: the αestradiol receptor (Erα — Green et al., 1986) and the β estradiolreceptor (Erβ — Kuiper et al., 1996). While Erα and Erβ are bothspecifically activated by estradiol binding, they are distinctlydistributed in the central nervous system (Shughrue et al., 1996,1997) where Erβ is thought to play a pivotal role by regulatingcell migration, apoptotic neural death (Wang et al., 2003) andastroglial cells proliferation (Wang et al., 2001). The transcrip-tional activity of Erα and Erβ is clearly distinct as recently
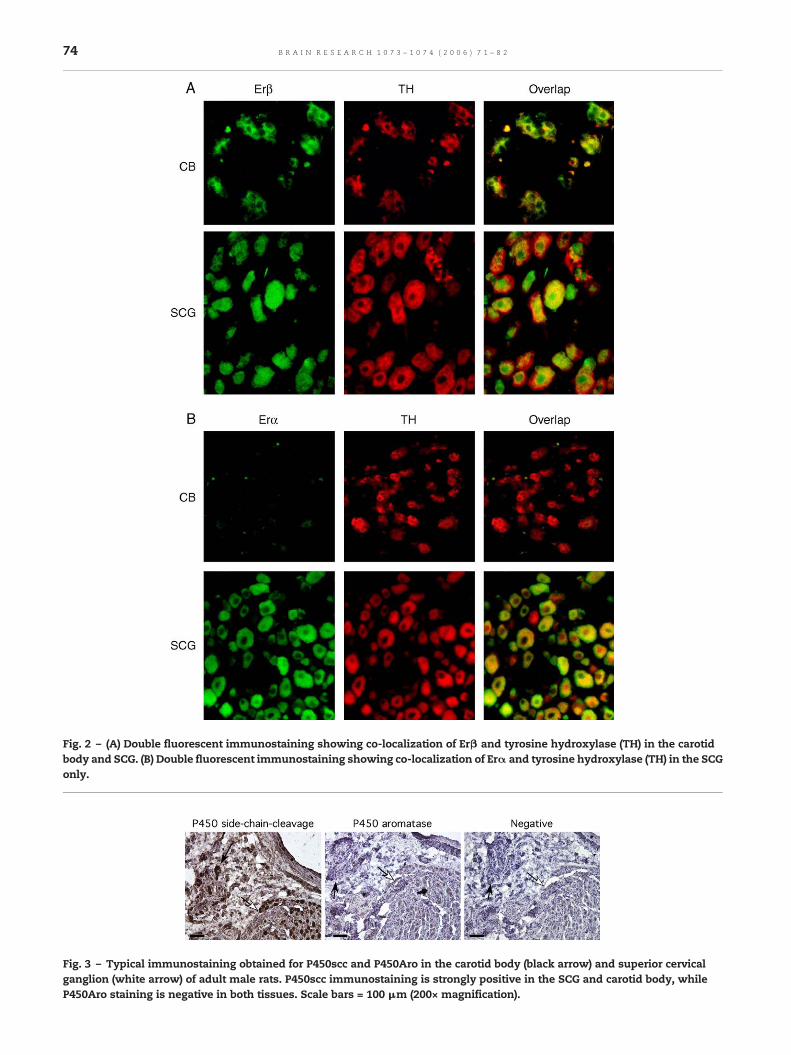
Fig. 2 – (A) Double fluorescent immunostaining showing co-localization of Erβ and tyrosine hydroxylase (TH) in the carotidbody and SCG. (B) Double fluorescent immunostaining showing co-localization of Erα and tyrosine hydroxylase (TH) in the SCGonly.
Fig. 3 – Typical immunostaining obtained for P450scc and P450Aro in the carotid body (black arrow) and superior cervicalganglion (white arrow) of adult male rats. P450scc immunostaining is strongly positive in the SCG and carotid body, whileP450Aro staining is negative in both tissues. Scale bars = 100 μm (200× magnification).
74 B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 7 1 – 8 2
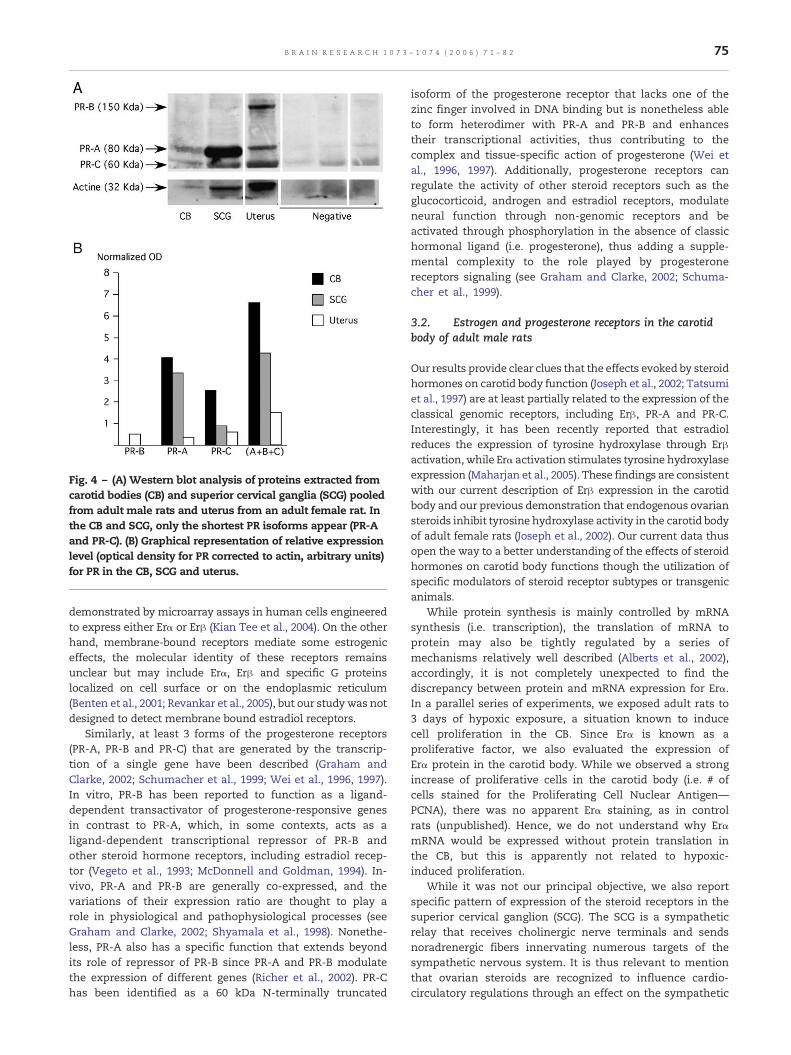
Fig. 4 – (A) Western blot analysis of proteins extracted fromcarotid bodies (CB) and superior cervical ganglia (SCG) pooledfrom adult male rats and uterus from an adult female rat. Inthe CB and SCG, only the shortest PR isoforms appear (PR-Aand PR-C). (B) Graphical representation of relative expressionlevel (optical density for PR corrected to actin, arbitrary units)for PR in the CB, SCG and uterus.
75B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 7 1 – 8 2
demonstrated by microarray assays in human cells engineeredto express either Erα or Erβ (Kian Tee et al., 2004). On the otherhand, membrane-bound receptors mediate some estrogeniceffects, the molecular identity of these receptors remainsunclear but may include Erα, Erβ and specific G proteinslocalized on cell surface or on the endoplasmic reticulum(Benten et al., 2001; Revankar et al., 2005), but our study was notdesigned to detect membrane bound estradiol receptors.
Similarly, at least 3 forms of the progesterone receptors(PR-A, PR-B and PR-C) that are generated by the transcrip-tion of a single gene have been described (Graham andClarke, 2002; Schumacher et al., 1999; Wei et al., 1996, 1997).In vitro, PR-B has been reported to function as a ligand-dependent transactivator of progesterone-responsive genesin contrast to PR-A, which, in some contexts, acts as aligand-dependent transcriptional repressor of PR-B andother steroid hormone receptors, including estradiol recep-tor (Vegeto et al., 1993; McDonnell and Goldman, 1994). In-vivo, PR-A and PR-B are generally co-expressed, and thevariations of their expression ratio are thought to play arole in physiological and pathophysiological processes (seeGraham and Clarke, 2002; Shyamala et al., 1998). Nonethe-less, PR-A also has a specific function that extends beyondits role of repressor of PR-B since PR-A and PR-B modulatethe expression of different genes (Richer et al., 2002). PR-Chas been identified as a 60 kDa N-terminally truncated
isoform of the progesterone receptor that lacks one of thezinc finger involved in DNA binding but is nonetheless ableto form heterodimer with PR-A and PR-B and enhancestheir transcriptional activities, thus contributing to thecomplex and tissue-specific action of progesterone (Wei etal., 1996, 1997). Additionally, progesterone receptors canregulate the activity of other steroid receptors such as theglucocorticoid, androgen and estradiol receptors, modulateneural function through non-genomic receptors and beactivated through phosphorylation in the absence of classichormonal ligand (i.e. progesterone), thus adding a supple-mental complexity to the role played by progesteronereceptors signaling (see Graham and Clarke, 2002; Schuma-cher et al., 1999).
3.2. Estrogen and progesterone receptors in the carotidbody of adult male rats
Our results provide clear clues that the effects evoked by steroidhormones on carotid body function (Joseph et al., 2002; Tatsumiet al., 1997) are at least partially related to the expression of theclassical genomic receptors, including Erβ, PR-A and PR-C.Interestingly, it has been recently reported that estradiolreduces the expression of tyrosine hydroxylase through Erβactivation, while Erα activation stimulates tyrosine hydroxylaseexpression (Maharjan et al., 2005). These findings are consistentwith our current description of Erβ expression in the carotidbody and our previous demonstration that endogenous ovariansteroids inhibit tyrosine hydroxylase activity in the carotid bodyof adult female rats (Joseph et al., 2002). Our current data thusopen the way to a better understanding of the effects of steroidhormones on carotid body functions though the utilization ofspecific modulators of steroid receptor subtypes or transgenicanimals.
While protein synthesis is mainly controlled by mRNAsynthesis (i.e. transcription), the translation of mRNA toprotein may also be tightly regulated by a series ofmechanisms relatively well described (Alberts et al., 2002),accordingly, it is not completely unexpected to find thediscrepancy between protein and mRNA expression for Erα.In a parallel series of experiments, we exposed adult rats to3 days of hypoxic exposure, a situation known to inducecell proliferation in the CB. Since Erα is known as aproliferative factor, we also evaluated the expression ofErα protein in the carotid body. While we observed a strongincrease of proliferative cells in the carotid body (i.e. # ofcells stained for the Proliferating Cell Nuclear Antigen—PCNA), there was no apparent Erα staining, as in controlrats (unpublished). Hence, we do not understand why ErαmRNA would be expressed without protein translation inthe CB, but this is apparently not related to hypoxic-induced proliferation.
While it was not our principal objective, we also reportspecific pattern of expression of the steroid receptors in thesuperior cervical ganglion (SCG). The SCG is a sympatheticrelay that receives cholinergic nerve terminals and sendsnoradrenergic fibers innervating numerous targets of thesympathetic nervous system. It is thus relevant to mentionthat ovarian steroids are recognized to influence cardio-circulatory regulations through an effect on the sympathetic
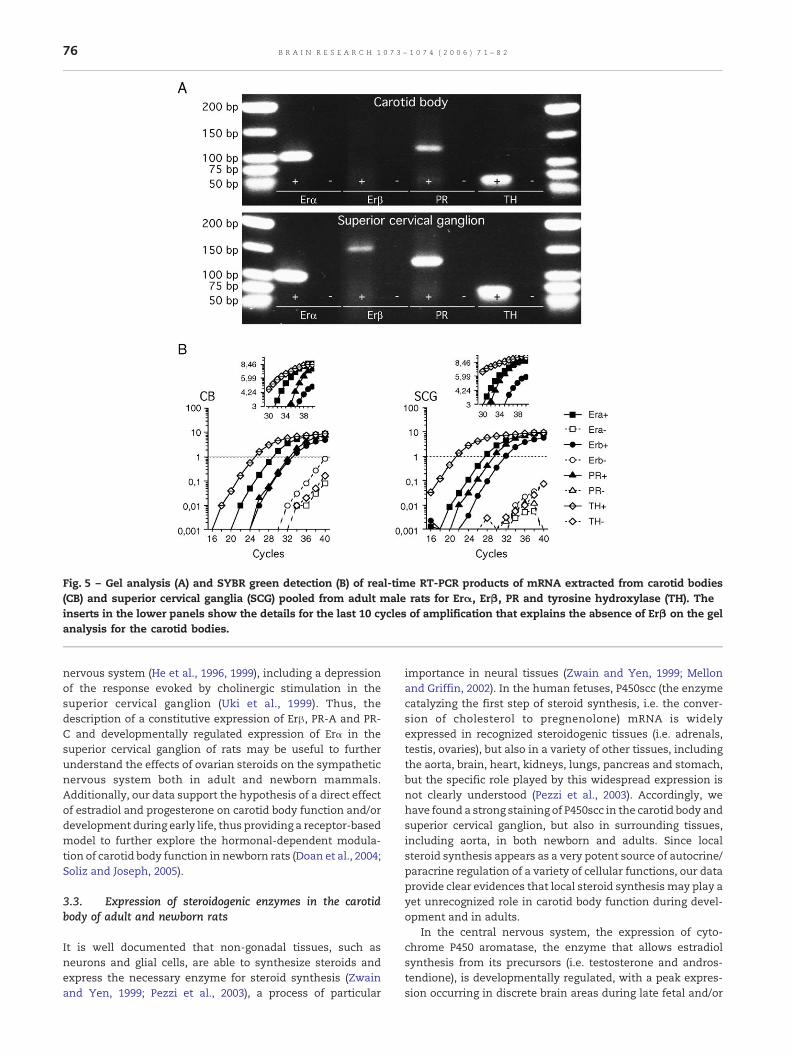
Fig. 5 – Gel analysis (A) and SYBR green detection (B) of real-time RT-PCR products of mRNA extracted from carotid bodies(CB) and superior cervical ganglia (SCG) pooled from adult male rats for Erα, Erβ, PR and tyrosine hydroxylase (TH). Theinserts in the lower panels show the details for the last 10 cycles of amplification that explains the absence of Erβ on the gelanalysis for the carotid bodies.
76 B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 7 1 – 8 2
nervous system (He et al., 1996, 1999), including a depressionof the response evoked by cholinergic stimulation in thesuperior cervical ganglion (Uki et al., 1999). Thus, thedescription of a constitutive expression of Erβ, PR-A and PR-C and developmentally regulated expression of Erα in thesuperior cervical ganglion of rats may be useful to furtherunderstand the effects of ovarian steroids on the sympatheticnervous system both in adult and newborn mammals.Additionally, our data support the hypothesis of a direct effectof estradiol and progesterone on carotid body function and/ordevelopment during early life, thus providing a receptor-basedmodel to further explore the hormonal-dependent modula-tion of carotid body function in newborn rats (Doan et al., 2004;Soliz and Joseph, 2005).
3.3. Expression of steroidogenic enzymes in the carotidbody of adult and newborn rats
It is well documented that non-gonadal tissues, such asneurons and glial cells, are able to synthesize steroids andexpress the necessary enzyme for steroid synthesis (Zwainand Yen, 1999; Pezzi et al., 2003), a process of particular
importance in neural tissues (Zwain and Yen, 1999; Mellonand Griffin, 2002). In the human fetuses, P450scc (the enzymecatalyzing the first step of steroid synthesis, i.e. the conver-sion of cholesterol to pregnenolone) mRNA is widelyexpressed in recognized steroidogenic tissues (i.e. adrenals,testis, ovaries), but also in a variety of other tissues, includingthe aorta, brain, heart, kidneys, lungs, pancreas and stomach,but the specific role played by this widespread expression isnot clearly understood (Pezzi et al., 2003). Accordingly, wehave found a strong staining of P450scc in the carotid body andsuperior cervical ganglion, but also in surrounding tissues,including aorta, in both newborn and adults. Since localsteroid synthesis appears as a very potent source of autocrine/paracrine regulation of a variety of cellular functions, our dataprovide clear evidences that local steroid synthesis may play ayet unrecognized role in carotid body function during devel-opment and in adults.
In the central nervous system, the expression of cyto-chrome P450 aromatase, the enzyme that allows estradiolsynthesis from its precursors (i.e. testosterone and andros-tendione), is developmentally regulated, with a peak expres-sion occurring in discrete brain areas during late fetal and/or
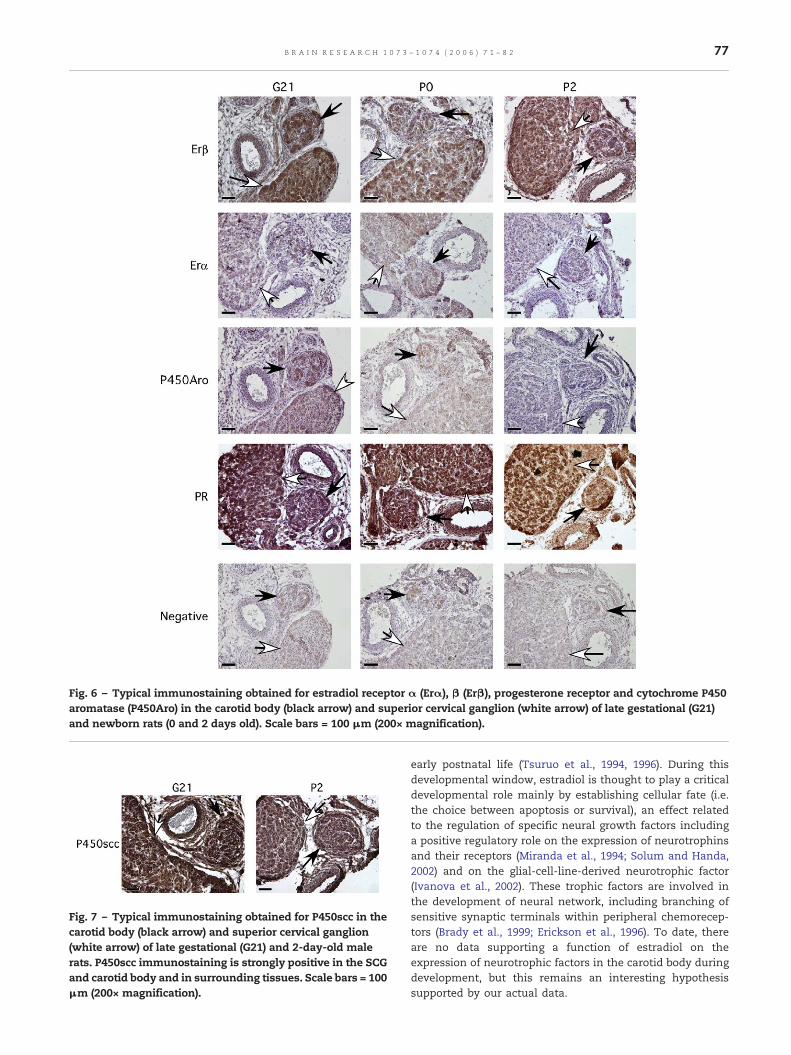
Fig. 6 – Typical immunostaining obtained for estradiol receptor α (Erα), β (Erβ), progesterone receptor and cytochrome P450aromatase (P450Aro) in the carotid body (black arrow) and superior cervical ganglion (white arrow) of late gestational (G21)and newborn rats (0 and 2 days old). Scale bars = 100 μm (200× magnification).
Fig. 7 – Typical immunostaining obtained for P450scc in thecarotid body (black arrow) and superior cervical ganglion(white arrow) of late gestational (G21) and 2-day-old malerats. P450scc immunostaining is strongly positive in the SCGand carotid body and in surrounding tissues. Scale bars = 100μm (200× magnification).
77B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 7 1 – 8 2
early postnatal life (Tsuruo et al., 1994, 1996). During thisdevelopmental window, estradiol is thought to play a criticaldevelopmental role mainly by establishing cellular fate (i.e.the choice between apoptosis or survival), an effect relatedto the regulation of specific neural growth factors includinga positive regulatory role on the expression of neurotrophinsand their receptors (Miranda et al., 1994; Solum and Handa,2002) and on the glial-cell-line-derived neurotrophic factor(Ivanova et al., 2002). These trophic factors are involved inthe development of neural network, including branching ofsensitive synaptic terminals within peripheral chemorecep-tors (Brady et al., 1999; Erickson et al., 1996). To date, thereare no data supporting a function of estradiol on theexpression of neurotrophic factors in the carotid body duringdevelopment, but this remains an interesting hypothesissupported by our actual data.

Table 1 – List of antibodies (and their respective antigens)used to perform immunohistochemistry of Erα, Erβ, PR,P450Aro and P450scc analysis
Antigen Primary antibody Secondaryantibody
Erα—prokaryoticrecombinantprotein,full-length Erα(Bevitt et al., 1997)
Mouse monoclonal1/100 (clone 6F11)—NovoCastraLaboratories,Newcastle uponTyne, UK
Horse anti-mouse(1/200—1 h)—VectorLaboratories
Erβ—18 aminoacid syntheticpeptides derivedfrom theC-terminus ofmouse Erβ(Hrabovszky et al.,2001; ShughrueandMerchenthaler,2001)
Rabbit polyclonal(clone Z8P)1 μg/ml—(ZymedLaboratories,South SanFrancisco, CA,USA)
Goat anti-rabbit(1/200—1 h)—VectorLaboratories
PR-syntheticpeptide against
Polyclonal (clone sc539)1/200—Santa Cruz
Goat anti-rabbit(1/200—1 h)—
78 B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 7 1 – 8 2
3.4. Conclusion
The detailed significance of the specific pattern of expressionfor progesterone and estradiol receptors aswell as P450scc andP450Aro expression in the carotid body of fetal or newborn ratsmay be put in perspective by considering recent results in thefield. In this context, our current data and actual experimentalmodel (Doan et al., 2004) are critical to further explore the roleplayed by steroids during respiratory control development, aparticularly important topic regarding the particular respira-tory physiology of premature newborn, which are criticallydependent on carotid body function to maintain adequaterespiratory drive (Al-Matary et al., 2004) and are chronicallydeprived from circulating steroids normally furnished by theplacenta (Trotter et al., 1999). It is also particularly relevant tounderlie the fact thatmaternal treatmentwith progesterone toavoid recurrent preterm delivery reduced the need for supple-mental oxygen in the newborn, an effect that could be, at leastpartially, related to an effect on neural respiratory control(Meis et al., 2003). Accordingly, future studies should aim atfurther elucidating the role of steroids, including but notlimited to estradiol andprogesterone for adequate carotidbodyfunctions both in newborn and adults.
an internaldomain of thehumanprogesteronereceptor
Biotechnology,Santa Cruz, CA, USA
VectorLaboratories
P450Aro—syntheticpeptide, aminoacids 376–390of humanaromatase (Turneret al., 2002)
Mouse monoclonal(clone H4) 1/20—Serotec, Raleigh,NC, USA
Horse anti-mouse(1/200—1 h)—VectorLaboratories
P450scc—aminoacids 421–441 of ratP450scc protein(Roby et al., 1991)
Rabbit polyclonal1/200—US Biological(Swampscott,MA, USA)
Goat anti-rabbit(1/200—1 h)—VectorLaboratories
4. Experimental procedures
4.1. Tissue preparation for immunohistochemistry
All experimental procedures have been approved by the localethic committee (Université Laval) on animal use and care. Twoweeks following arrival in the laboratory, adult Sprague–Dawleymale rats (n = 10, 2–4 months old, ordered from Charles River,St Constant, Canada) were deeply anesthetized (ketamine/xylazine), the chest was rapidly opened and the animals wereperfused with 500 ml ice-cold PBS solution through the heart.Following perfusion, the neck was opened, and the carotidbifurcation was exposed, cleaned from surrounding tissues anddissected en-bloc with the superior cervical ganglion andcarotid body. The dissected bifurcation was post-fixed in 4%PFA for 24 h at 4 °C, cryoprotected in 30% sucrose at 4 °C for 24 h,embedded in tissue-tek and frozen on dry ice. Tissues slices(10 μm thickness) were cut on a cryostat, mounted on glass slidesand kept frozen (−80 °C).
Newborn male rats (P0, i.e. 6–12 h following birth, P2, i.e. 48–60h following birth, n = 4 for each age, born from a total of 2 litters)were anesthetized inmelting ice, the heart was rapidly removed toavoid bleeding during dissection, the neck was opened and thecarotid bifurcation removed en-bloc. For late fetal rats, timepregnant rats (n = 2) were obtained in our animal facility on thebasis of daily vaginal smears to determine estrous cycle andoptimal mating day. On the 21st gestational day (i.e. 1 day beforedelivery), the pregnant damwas deeply anesthetized (as describedfor adults) and the fetuses removed from the uterine horns fordissection of the carotid bifurcation as described above. Tissueswere then fixed, embedded, cut and mounted on glass slides asdescribed for adult rats.
4.2. Immunohistochemistry
We used antibodies that have been firmly recognized for bindingspecificity (see Table 1 and included references), and preliminary
studies showed staining at adequatemolecular weight inWesternblots on protein extracted from control tissues (not shown).
4.2.1. With DAB revelationTissues slices were dried 2 h (37 °C) and washed 3 × 5 min in PBS.Antigen retrieval was performed by placing the slides in amicrowave oven at maximum power (700 W) and boiled for 20min in sodium citrate (10 mM, pH 6.0). Slides have been allowed tocool to room temperature for 30minwithin the citrate solution andwashed 3 × 5min in PBS before blockade of endogenous peroxidaseactivity with H2O2 (3%) and washed 3 × 5 min in PBS. Non-specificbinding sites have been blocked for 30–60minwith blocking serum(goat serum 2%, Triton 0.2% in PBS) before incubation with theselected primary antibody (see Table 1) for 16 h at 4 °C.
Slides are then washed 3 × 5 min in PBS before incubation withsecondary biotinylated antibody (see Table 1). Slides are thenwashed 3 × 5 min in PBS, incubated in the Vectastain Elite avidin-biotinylated enzyme complex method and the antigen–antibodycomplex is visualized with diaminiobenzidine peroxidase (DAB-Vector Laboratories). Following DAB reaction, counterstaining wasperformed with hematoxylin for 30–60 s. Negative control slideswere treated under strictly similar conditions without primaryantibodies. All slices have been observed under a Nikon Eclipse

79B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 7 1 – 8 2
E600 microscope. For each antibody tested, at least 2 animals(from different litters for newborn) have been tested.
4.2.2. Double immunofluorescence stainingTissues slices have been treated as described above with Erβ or Erαprimary and secondary antibodies washed 3 × 5 min in PBS thenincubated with fluorescent avidin-D (2 μg/ml—30 min at roomtemperature, Vector Laboratories, Burlingame, CA, USA) for FITC(i.e. green) fluorescence. Slides were washed in PBS (3 × 5 min),incubated with blocking serum 15 min, washed (2 × 5 min) andincubated with the avidin/biotin blocking kit (Vector Laboratories)for 15 min and washed 2 × 5 min then incubated with normalblocking serum 30–60 min at room temperature. Slides were thenincubated with tyrosine hydroxylase antibody 90 min at 37 °C(mouse TH monoclonal antibody, 1/500—Chemicon, Temecula,CA, USA), washed 3 × 5 min in PBS before incubation withsecondary biotinylated antibody (horse anti-mouse, 1/200—1 h).Slideswerewashed 3 × 5min in PBS then incubatedwith Texas RedAvidin D for 30 min at room temperature (2 μg/ml—VectorLaboratories, Burlingame, CA, USA) for Texas Red fluorescence.Slides were washed 3 × 5 min in PBS, covered with Vectashieldmounting media (Vector Laboratories) and observed under afluorescent microscope (Nikon Eclipse E600). Negative controlslides were treated under strictly similar conditions withoutprimary antibodies. TH staining appeared with a red fluorescence(excitation wavelength: 540–580 nm, filtered emission: 600–660nm) and Erβ or Erα with a green fluorescence (excitationwavelength: 465–495 nm, filtered emission: 515–555 nm). All theimages presented have been processed using the followingprocedure: for each fluorochrome, the acquisition software deter-mined the adequate exposure time, and the selected images werestored on a computer. Negative control slides were then recordedunder strictly similar conditions for FITC and Texas Red fluores-cence. The images were analyzed with Adobe Photoshop using astandard procedure for background subtraction: the histogram inthe adequate color channel was read from the negative control andadjusted until elimination of background and auto-fluorescencesignals. The same correction was then applied to the corres-ponding positive image to obtain the final corrected image.
4.3. Tissue dissection for Western blot and real-time RT-PCR
The animal preparation, surgery and organ collection were similarto that described previously (Kinkead et al., 2005). Briefly, eachanimal (n = 17, 5–6 months old) was anesthetized with an i.p.injection of ketamine (10 mg/100g) and xylazine (1.5 mg/100 g).Then, the rat was rapidly tracheotomized and artificially venti-lated. The carotid body and superior cervical ganglion werecleaned from the surrounding tissues, quickly removed, frozenimmediately on dry ice and stored at −80 °C. Two pools containing
Table 2 – Sequence of primers pairs used to perform real-time R
Gene Sequence
Erα Sense 5′ GGTGGACCCATTGCCTTTC 3′Antisense 5′ GATGAGACACCAGGGCGTTAA
Erβ Sense 5′ GTCTCCTTTAGCGACCCATTG 3Antisense 5′ CATGTAACAGGGCTGGCACAAC
PR Sense 5′ CGACACTTCCAGCTCTTTGCT 3Antisense 5′ GAATCAGGGTTATCTGGTCATC
TH Sense 5′ CTCAGAGCAGGATGCCAAGC 3′Antisense 5′ TGTCGCCGTCCGATGAA 3′
respectively 12 and 5 pairs of carotid body and superior cervicalganglion were constituted for Western blot and real-time RT-PCRanalysis respectively.
4.4. Western blot
Carotid bodies (n = 24, weight = 2.2mg) and SCG (n = 24, weight = 25mg) pools were placed in 300 μl (carotid body) or 500 μl (superiorcervical ganglion) RIPA extraction buffer (20 mM Tris, 300 mMNaCl, 0.5% SDS, 0.5% NP-40, 0.5% sodium deoxycholate, pH 7.2)containing complete protease inhibitor following the descriptionof manufacturer (Roche Diagnostic, Laval, QC, Canada) and 1 μg/ml pepstatin A, 0.1 mg/ml benzamidine HCL and 1 mM sodiumortho-vanadate (Sigma-Aldrich, Oakville, ON, Canada). Then, thehomogenization wasmade with a Polytron homogenizer for 30–90s at 4 °C (pta-7, Brinkmann, Mississauga, ON, Canada).
The homogenized materials were kept on ice for 30 minbefore centrifugation at 13,000×g for 30 min. The supernatantfluid was collected, and total protein concentrations weredetermined by spectrophotometry using a BCA™ protein assaykit (Pierce, Rockford, IL, USA). For carotid bodies, the proteinassay was below detection threshold. The protein extract (200 μl)was thus concentrated to 35 μl using a microcon YM-10(Millipore corporation, Bedford, MA, USA) filter and preparedfor loading in 2 immunoblotting wells (1 positive and 1 negative,see below). For superior cervical ganglion and uterus, 75 μg ofproteins was used. Proteins were mixed with loading buffer (75mM Tris–HCL, pH 6.8, 10% glycerol, 720 mM beta-mercaptoetha-nol, 2% SDS, 0.1% bromophenol blue) and denatured for 5 min at95 °C, electrophoresed by SDS-PAGE on 7.5% gel and electro-transferred overnight onto nitrocellulose membranes (GE Osmo-nics, Minnetonka, Minnesota, USA). Membranes were thenrinsed in PBS with 0.1% Tween-20 (PBS-T), blocked for 1 h in5% fat-free milk, secondary antibody (1/10,000) and 0.1% sodiumazide in PBS-T at room temperature, washed three times inblocking solution without secondary antibody. Membranes werecut around the 48 kDa weight maker. They were then incubatedin blocking solution without secondary antibody for 3 h at roomtemperature with progesterone receptor antibody (upper part ofthe membrane—see Table 1) or with mouse anti-actin mono-clonal primary antibody (lower part of the membrane, 1/200,clone JLA20, DSHB, University of Iowa, IA, USA). After threewashes in PBS-T, the membranes were incubated for 1 h at roomtemperature in horseradish-peroxidase-conjugated secondaryantibody of either 1/20,000 goat anti-rabbit (Jackson Immuno-research, West Grove, PA, USA) or 1/10,000 goat anti-mouse(Jackson Immunoresearch, West Grove, PA, USA) in PBS-T. Afterwashing with PBS-T three times, specific bands were visualizedby chemiluminescence reagents (Amersham Pharmacia Biotech,Montreal, QC, Canada).
T-PCR analysis of Erα, Erβ, PR and TH
Expected product size Accession no.
101 bp NM_0010085113′′ 159 bp NM_012754.13′
′ 131 bp NM_022847.1G 3′
63 bp NM_012740.2

80 B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 7 1 – 8 2
Membranes were exposed to X-ray film (FUJI) for 45 min(progesterone) or 30min (actin), and the intensity of the signal wasquantified by densitometry using the NIH ImageJ software. Thespecificity of antibody was tested in samples run and incubatedwithout primary antibody.
4.5. Real-time RT-PCR
Total RNA extraction and the RT phase (with 1 μg RNA for superiorcervical ganglion and 100 ng for carotid body) using randomdecamer primers for transcripts amplifications were performed aspreviously described (Joseph and Bairam, 2004; Kinkead et al.,2005). Aliquots of 2 μl from the resulting single-stranded cDNAproducts were used along with the appropriate primers (see Table2) for Erα, Erβ and PR, and 1 μl was used for tyrosine hydroxylase(TH) mRNA amplification. Each amplifications was run in separatewells with 2× SYBRGreenUniversal PCRMasterMix containing 400nMofeachof specific forwardand reverse geneprimers (total 25μl).Primers used have been designed to overlap over the boundaries oftwo exons to avoid amplification of genomicDNA, using the PrimerExpress software program (Applied Biosystems, version 2.0, Fostercity, CA). PCR amplification was performed on an ABI Prism 7000Sequence Detector System (Applied Biosystems, Foster city, CA)according to manufacturer's instruction. Amplification conditionswere the same for all assays: one cycle at 95 °C for 10min, 40 cyclesat 95 °C for 15 s and a final step at 60 °C for 60 s. Control samplesamplified without RTwere run and did not yield any amplificationsignal. PCR products were loaded on a 2% agarose gel stained withethidiumbromide (2μl/100ml) formigrationduring 3hat 200Vandvisualized under UV trans-illumination.
Acknowledgments
The authors are thankful to Evelyne Vachon and Sylvie Vigerfor animal care. Jorge Soliz for careful reading of themanuscript and helpful comments. Catherine-Eve Morencyreceived a fellowship from Centre de Recherche de l'Hôpital StFrançois d'Assise.
Grants: supported by Fond de la Recherche en Santé duQuébec, The Hospital for Sick Children Foundation (Grant #XG04-032), Canadian Institute for Health Research and Fondationde la Recherche des Maladies Infantiles.
R E F E R E N C E S
Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K., Walter, P.,2002. Postranscriptional Controls. Molecular Biology of TheCell, Fourth ed. Garland Science, Taylo and Francis Group, NewYork.
Al-Matary, A., Kutbi, I., Qurashi, M., Khalil, M., Alvaro, R.,Kwiatkowski, K., Cates, D., Rigatto, H., 2004. Increasedperipheral chemoreceptor activity may be critical indestabilizing breathing in neonates. Semin. Perinatol. 28,264–272.
Benten, W.P., Stephan, C., Lieberherr, M., Wunderlich, F., 2001.Estradiol signaling via sequestrable surface receptorsEndocrinology 142, 1669–1677.
Bevitt, D.J., Milton, I.D., Piggot, N., Henry, L., Carter, M.J., Toms,G.L., Lennard, T.W., Westley, B., Angus, B., Horne, C.H., 1997.New monoclonal antibodies to oestrogen and progesterone
receptors effective for paraffin sectionimmunohistochemistry. J. Pathol. 183, 228–232.
Beyer, C., Damm, N., Brito, V., Kuppers, E., 2002. Developmentalexpression of progesterone receptor isoforms in the mousemidbrain. NeuroReport 13, 877–880.
Brady, R., Zaidi, S.I., Mayer, C., Katz, D.M., 1999. BDNF is atarget-derived survival factor for arterial baroreceptor andchemoafferent primary sensory neurons. J. Neurosci. 19,2131–2142.
Chenuel, B.J., Smith, C.A., Henderson, K.S., Dempsey, J.A., 2005.Increased propensity for apnea via dopamine-induced carotidbody inhibition in sleeping dogs. J. Appl. Physiol. 98,1732–1739.
Doan, V.D., Gagnon, S., Joseph, V., 2004. Prenatal blockade ofestradiol synthesis impairs respiratory and metabolicresponses to hypoxia in newborn and adult rats. Am. J. Physiol.:Regul., Integr. Comp. Physiol. 287, R612–R618.
Erickson, J.T., Conover, J.C., Borday, V., Champagnat, J., Barbacid,M., Yancopoulos, G., Katz, D.M., 1996. Mice lackingbrain-derived neurotrophic factor exhibit visceral sensoryneuron losses distinct from mice lacking NT4 and display asevere developmental deficit in control of breathing.J. Neurosci. 16, 5361–5371.
Gonzalez, C., Almaraz, L., Obeso, A., Rigual, R., 1994. Carotid bodychemoreceptors: from natural stimuli to sensory discharges.Physiol. Rev. 74, 829–898.
Graham, J.D., Clarke, C.L., 2002. Expression and transcriptionalactivity of progesterone receptor A and progesterone receptor Bin mammalian cells. Breast Cancer Res. 4, 187–190.
Green, S., Walter, P., Kumar, V., Krust, A., Bornert, J.M., Argos, P.,Chambon, P., 1986. Human oestrogen receptor cDNA:sequence, expression and homology to v-erb-A. Nature 320,134–139.
Hannhart, B., Pickett, C.K., Moore, L.G., 1990. Effects of estrogenand progesterone on carotid body neural outputresponsiveness to hypoxia. J. Appl. Physiol. 68, 1909–1916.
He, L., Chen, J., Dinger, B., Stensaas, L., Fidone, S., 1996. Endothelinmodulates chemoreceptor cell function in mammalian carotidbody. Adv. Exp. Med. Biol. 410, 305–311.
He, X.R., Wang, W., Crofton, J.T., Share, L., 1999. Effects of17beta-estradiol on the baroreflex control of sympatheticactivity in conscious ovariectomized rats. Am. J. Physiol. 277,R493–R498.
Hrabovszky, E., Steinhauser, A., Barabas, K., Shughrue, P.J.,Petersen, S.L., Merchenthaler, I., Liposits, Z., 2001. Estrogenreceptor-beta immunoreactivity in luteinizing hormone-releasing hormone neurons of the rat brain. Endocrinology 142,3261–3264.
Ivanova, T., Kuppers, E., Engele, J., Beyer, C., 2001. Estrogenstimulates brain-derived neurotrophic factor expression inembryonic mouse midbrain neurons through amembrane-mediated and calcium-dependent mechanism.J. Neurosci. Res. 66, 221–230.
Ivanova, T., Karolczak, M., Beyer, C., 2002. Estradiol stimulatesGDNF expression in developing hypothalamic neurons.Endocrinology 143, 3175–3178.
Joseph, V., Bairam, A., 2004. Differential regulation of short andlong dopamine D2 receptor mRNA levels by hypoxia in theadrenals of 1-day-old and adult rabbits. Brain Res. Mol. BrainRes. 130, 115–123.
Joseph, V., Pequignot, J.M., 2003. Neurochemical processesinvolved in acclimatization to long term hypoxia. In: Lahiri, S.,Semenza, G.L., Prabhakar, N.R. (Eds.), Oxygen Sensing:Responses and Adaptation to Hypoxia. Marcel Dekker Inc.,New York.
Joseph, V., Soliz, J., Soria, R., Pequignot, J., Favier, R., Spielvogel, H.,Pequignot, J.M., 2002. Dopaminergic metabolism in carotidbodies and high altitude acclimatization in female rats. Am. J.Physiol.: Regul., Integr. Comp. Physiol. 282, R765–R773.

81B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 7 1 – 8 2
Kian Tee, M., Rogatsky, I., Tzagarakis-Foster, C., Cvoro, A., An, J.,Christy, R.J., Yamamoto, K.R., Leitman, D.C., 2004. Estradiol andselective estrogen receptor modulators differentially regulatetarget genes with estrogen receptors alpha and beta. Mol. Biol.Cell 15, 1262–1272.
Kinkead, R., Joseph, V., Lajeunesse, Y., Bairam, A., 2005. Neonatalmaternal separation enhances dopamine D2-receptor andtyrosine hydroxylase mRNA expression levels in carotid bodyof rats. Can. J. Physiol. Pharmacol. 83, 76–84.
Kuiper, G.G., Enmark, E., Pelto-Huikko, M., Nilsson, S., Gustafsson,J.A., 1996. Cloning of a novel receptor expressed in rat prostateand ovary. Proc. Natl. Acad. Sci. U. S. A. 93, 5925–5930.
León-Velarde, F., Ramos, M.A., Hernández, J.A., De Idiáquez, D.,Muñoz, L.S., Gaffo, A., Córdova, S., Durand, D., Monge-C, C.,1997. The role of menopause in the development of chronicmountain sickness. Am. J. Physiol. 272, R90–R94.
León-Velarde, F., Rivera-Chira, M., Tapia, R., Huicho, L.,Monge-C, C., 2001. Relationship of ovarian hormones tohypoxemia in women residents of 4,300 m. Am. J. Physiol.280, R488–R493.
Maharjan, S., Serova, L., Sabban, E.L., 2005. Transcriptionalregulation of tyrosine hydroxylase by estrogen: oppositeeffects with estrogen receptors alpha and beta and interactionswith cyclic AMP. J. Neurochem. 93, 1502–1514.
McDonnell, D.P., Goldman, M.E., 1994. RU486 exertsantiestrogenic activities through a novel progesteronereceptor A form-mediated mechanism. J. Biol. Chem. 269,11945–11949.
Meis, P.J., Klebanoff, M., Thom, E., Dombrowski, M.P., Sibai, B.,Moawad, A.H., Spong, C.Y., Hauth, J.C., Miodovnik, M., Varner,M.W., Leveno, K.J., Caritis, S.N., Iams, J.D., Wapner, R.J.,Conway, D., O'Sullivan, M.J., Carpenter, M., Mercer, B., Ramin, S.M., Thorp, J.M., Peaceman, A.M., Gabbe, S., 2003. Prevention ofrecurrent preterm delivery by 17 alpha-hydroxyprogesteronecaproate. N. Engl. J. Med. 348, 2379–2385.
Mellon, S.H., Griffin, L.D., 2002. Neurosteroids: biochemistry andclinical significance. Trends Endocrinol. Metab. 13, 35–43.
Miranda, R.C., Sohrabji, F., Toran-Allerand, D., 1994. Interactions ofestrogen with the neurotrophins and their receptors duringneural development. Horm. Behav. 28, 367–375.
Moore, L.G., McCullough, R.E., Weil, J.V., 1987. Increased HVR inpregnancy: relationship to hormonal and metabolic changes.J. Appl. Physiol. 62, 158–163.
Nakayama, H., Smith, C.A., Rodman, J.R., Skatrud, J.B., Dempsey,J.A., 2003. Carotid body denervation eliminates apnea inresponse to transient hypocapnia. J. Appl. Physiol. 94,155–164.
Nilsen, J., Mor, G., Naftolin, F., 2000. Estrogen-regulateddevelopmental neuronal apoptosis is determined by estrogenreceptor subtype and the Fas/Fas ligand system. J. Neurobiol.43, 64–78.
Okada, A., Ohta, Y., Buchanan, D., Sato, T., Iguchi, T., 2002. Effect ofestrogens on ontogenetic expression of progesterone receptorin the fetal female rat reproductive tract. Mol. Cell. Endocrinol.195, 55–64.
Pezzi, V., Mathis, J.M., Rainey, W.E., Carr, B.R., 2003. Profilingtranscript levels for steroidogenic enzymes in fetal tissues.J. Steroid Biochem. Mol. Biol. 87, 181–189.
Revankar, C.M., Cimino, D.F., Sklar, L.A., Arterburn, J.B., Prossnitz,E.R., 2005. A transmembrane intracellular estrogen receptormediates rapid cell signaling. Science 307, 1625–1630.
Richer, J.K., Jacobsen, B.M., Manning, N.G., Abel, M.G., Wolf, D.M.,Horwitz, K.B., 2002. Differential gene regulation by the twoprogesterone receptor isoforms in human breast cancer cells.J. Biol. Chem. 277, 5209–5218.
Roby, K.F., Larsen, D., Deb, S., Soares, M.J., 1991. Generation andcharacterization of antipeptide antibodies to rat cytochromeP-450 side-chain cleavage enzyme. Mol. Cell. Endocrinol. 79,13–20.
Saaresranta, T., Aittokallio, T., Polo-Kantola, P., Helenius, H., Polo,O., 2003. Effect of medroxyprogesterone on inspiratory flowshapes during sleep in postmenopausal women. Respir.Physiol. Neurobiol. 134, 131–143.
Schumacher, M., Coirini, H., Robert, F., Guennoun, R., El-Etr, M.,1999. Genomic and membrane actions of progesterone:implications for reproductive physiology and behavior. Behav.Brain Res. 105, 37–52.
Shahar, E., Redline, S., Young, T., Boland, L.L., Baldwin, C.M., Nieto,F.J., O'Connor, G.T., Rapoport, D.M., Robbins, J.A., 2003.Hormone replacement therapy and sleep-disorderedbreathing. Am. J. Respir. Crit. Care Med. 167, 1186–1192.
Shughrue, P.J., Merchenthaler, I., 2001. Distribution of estrogenreceptor beta immunoreactivity in the rat central nervoussystem. J. Comp. Neurol. 436, 64–81.
Shughrue, P.J., Komm, B., Merchenthaler, I., 1996. The distributionof estrogen receptor-beta mRNA in the rat hypothalamus.Steroids 61, 678–681.
Shughrue, P.J., Lane, M.V., Merchenthaler, I., 1997. Comparativedistribution of estrogen receptor-alpha and -beta mRNA inthe rat central nervous system. J. Comp. Neurol. 388,507–525.
Shyamala, G., Yang, X., Silberstein, G., Barcellos-Hoff, M.H., Dale,E., 1998. Transgenic mice carrying an imbalance in the nativeratio of A to B forms of progesterone receptor exhibitdevelopmental abnormalities in mammary glands. Proc. Natl.Acad. Sci. U. S. A. 95, 696–701.
Smith, C., Bisgard, G., Nielsen, A., Daristotle, L., Kressin, N., Forster,H., Dempsey, J., 1986. Carotid bodies are required forventilatory acclimatization to chronic hypoxia. J. Appl. Physiol.60, 1003–1010.
Smith, C.A., Nakayama, H., Dempsey, J.A., 2003. The essential roleof carotid body chemoreceptors in sleep apnea. Can. J. Physiol.Pharmacol. 81, 774–779.
Soliz, J., Joseph, V., 2005. Perinatal steroid exposure and respiratorycontrol during early postnatal life. Respir. Physiol. Neurobiol.149, 111–122.
Solum, D.T., Handa, R.J., 2002. Estrogen regulates the developmentof brain-derived neurotrophic factor mRNA and protein in therat hippocampus. J. Neurosci. 22, 2650–2659.
Tatsumi, K., Hannhart, B., Pickett, C.K., Weil, J.V., Moore, L.G., 1991.Influences of gender and sex hormones on hypoxic ventilatoryresponse in cats. J. Appl. Physiol. 71, 1746–1751.
Tatsumi, K., Pickett, C.K., Jacoby, C.R., Weil, J.V., Moore, L.G., 1997.Role of endogenous female hormones in hypoxicchemosensitivity. J. Appl. Physiol. 83, 1706–1710.
Trotter, A., Maier, L., Grill, H.J., Wudy, S.A., Pohlandt, F., 1999.17Beta-estradiol and progesterone supplementation inextremely low-birth-weight infants. Pediatr. Res. 45,489–493.
Tsuruo, Y., Ishimura, K., Fujita, H., Osawa, Y., 1994.Immunocytochemical localization of aromatase-containingneurons in the rat brain during pre- and postnataldevelopment. Cell Tissue Res. 278, 29–39.
Tsuruo, Y., Ishimura, K., Hayashi, S., Osawa, Y., 1996.Immunohistochemical localization of estrogen receptorswithin aromatase-immunoreactive neurons in the fetal andneonatal rat brain. Anat. Embryol. (Berl.) 193, 115–121.
Turner, K.J., Macpherson, S., Millar, M.R., McNeilly, A.S., Williams,K., Cranfield, M., Groome, N.P., Sharpe, R.M., Fraser, H.M.,Saunders, P.T., 2002. Development and validation of a newmonoclonal antibody to mammalian aromatase. J. Endocrinol.172, 21–30.
Uki, M., Nabekura, J., Akaike, N., 1999. Suppression of the nicotinicacetylcholine response in rat superior cervical ganglionicneurons by steroids. J. Neurochem. 72, 808–814.
Vegeto, E., Shahbaz, M.M., Wen, D.X., Goldman, M.E., O'Malley, B.W., McDonnell, D.P., 1993. Human progesterone receptor Aform is a cell- and promoter-specific repressor of human

82 B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 7 1 – 8 2
progesterone receptor B function. Mol. Endocrinol. 7,1244–1255.
Wang, L., Andersson, S., Warner, M., Gustafsson, J.A., 2001.Morphological abnormalities in the brains of estrogenreceptor beta knockout mice. Proc. Natl. Acad. Sci. U. S. A. 98,2792–2796.
Wang, L., Andersson, S., Warner, M., Gustafsson, J.A., 2003.Estrogen receptor (ER)beta knockout mice reveal a role forERbeta inmigration of cortical neurons in the developing brain.Proc. Natl. Acad. Sci. U. S. A. 100, 703–708.
Wei, L.L., Hawkins, P., Baker, C., Norris, B., Sheridan, P.L.,Quinn, P.G., 1996. An amino-terminal truncatedprogesterone receptor isoform, PRc, enhances
progestin-induced transcriptional activity. Mol. Endocrinol.10, 1379–1387.
Wei, L.L., Norris, B.M., Baker, C.J., 1997. An N-terminally truncatedthird progesterone receptor protein, PR(C), forms heterodimerswith PR(B) but interferes in PR(B)-DNA binding. J. SteroidBiochem. Mol. Biol. 62, 287–297.
Young, T., Finn, L., Austin, D., Peterson, A., 2003. Menopausalstatus and sleep-disordered breathing in the WisconsinSleep Cohort Study. Am. J. Respir. Crit. Care Med. 167,1181–1185.
Zwain, I.H., Yen, S.S., 1999. Neurosteroidogenesis in astrocytes,oligodendrocytes, and neurons of cerebral cortex of rat brain.Endocrinology 140, 3843–3852.