Embed Size (px)
Citation preview

Expression of POU, Sox, and Pax Genes in the BrainGanglia of the Tropical Abalone Haliotis asinina
Elizabeth K. O’Brien and Bernard M. Degnan*
Department of Zoology and Entomology, University of Queensland, Brisbane, Queensland 4072, Australia
Abstract: In gastropod mollusks, neuroendocrine cells in the anterior ganglia have been shown to regulate
growth and reproduction. As a first step toward understanding the molecular mechanisms underlying the
regulation of these physiological processes in the tropical abalone Haliotis asinina, we have identified sets of
POU, Sox, and Pax transcription factor genes that are expressed in these ganglia. Using highly degenerate
oligonucleotide primers designed to anneal to conserved codons in each of these gene families, we have
amplified by reverse transcriptase polymerase chain reaction 2 POU genes (HasPOU-III and HasPOU-IV), 2 Sox
genes (HasSox-B and HasSox-C), and two Pax genes (HasPax-258 and HasPax-6). Analyses with gene-specific
primers indicated that the 6 genes are expressed in the cerebral and pleuropedal ganglia of both reproductively
active and spent adults, in a number of sensory structures, and in a subset of other adult tissues.
Key words: POU, Sox, Pax, ganglia, mollusk, lophotrochozoan.
INTRODUCTION
Regulation of molluscan growth and reproduction operates
via a first-order neurosecretory mechanism that directly af-
fects the target organ (reviewed in Geraerts et al., 1991).
Environmental regulation of these physiological processes
appears to be via the anterior ganglia, which have direct
neural connections with specialized sensory structures and
house a suite of neuroendocrine cells (Roubos and van der
Wal-Divendal, 1982; Geraerts, 1986). For example, in Lym-
naea stagnalis hormones produced by neurosecretory cells
within the cerebral ganglia directly regulate growth and re-
production and maintain an antagonism between these two
physiological states (Geraerts, 1976; reviewed in Joose,
1988).
Analysis of the neuroendocrinology of commercially
important gastropod mollusks, such as the abalone (Veti-
gastropoda: Haliotidae), has been limited. Studies on aba-
lone have identified neurosecretory cells in the cerebral gan-
glia (Yahata, 1971; Hahn 1994; Upatham et al., 1998; Je-
breen et al., in press) and correlated neurosecretory activity
in the anterior ganglia with the reproductive cycle (Yahata,
1973; Hahn, 1994; Jebreen et al., in press). Although the
abalone central nervous system (CNS) exists in the primi-
tive streptoneurous condition (Crofts, 1929) and differs
from the more complex nervous systems of pulmonates, it
has been suggested that L. stagnalis can serve as a model for
understanding the underlying mechanisms controlling
growth and reproduction in molluscan species of commer-
cial importance (Geraerts et al., 1991).
Compared with other abalone, the tropical abalone
Haliotis asinina has an extremely predictable reproductive
and spawning cycle (R.T. Counihan et al., in press), grows
Received March 21, 2000; accepted June 21, 2000.
*Corresponding author: telephone +61-7-3365 2467; fax +61-7-3365 1655; e-mail
Mar. Biotechnol. 2, 545–557, 2000DOI: 10.1007/s101260000039
© 2000 Springer-Verlag New York Inc.

rapidly, and has a short generation time (reviewed in
Counihan et al., 1998). These features make H. asinina a
suitable abalone for investigating the molecular and cellular
bases of growth and reproduction in this commercially im-
portant group (Counihan et al., 1998).
The role of neuropeptides in regulating gastropod
growth and reproduction is well known (reviewed in
Geraerts et al., 1991), but the transcription factors control-
ling the expression of these peptides and the molecular
circuitry linking environmental stimuli to physiological
changes are still relatively unexplored. As a first step toward
understanding the molecular cascade underlying environ-
mentally induced changes in the physiology of the abalone,
we sought to identify a suite of transcription factor genes
expressed in the adult ganglia. Differential expression of
these trans-factors may underlie changes in the physiologi-
cal state given that they control not only the expression of
neuropeptide genes but also genes encoding enzymes and
other factors required for the correct processing and secre-
tion of the peptides (e.g., Ojeda et al., 1999).
In this study we focused on the evolutionarily con-
served POU, Sox, and Pax gene families because of their
consistent and essential association, across phyla, with the
anterior CNS during development and their continued as-
sociation in maturity. Genes from the POU multigene fam-
ily encode proteins that include an approximately 75 amino
acid POU-specific domain and a 60 amino acid homeodo-
main (Herr et al., 1988). Members of this family regulate
the development of central and peripheral nervous systems
(reviewed in Ryan and Rosenfeld, 1997; Veenstra et al.,
1997), are involved in the activation of growth genes (Man-
agalam et al., 1989), and have been implicated in the neu-
roendocrine control of puberty and ovulation (Ryan and
Rosenfeld, 1997; Wierman et al., 1997; Ojeda et al., 1999)
and in the regulation of neurotransmitter synthesis (Treacy
et al., 1991). Sox, a high mobility group (HMG) gene family,
encodes a protein with a conserved HMG box of 79 amino
acids (Janzten et al., 1990; Wright et al., 1993). This family
includes gene products known to interact with POU pro-
teins (e.g., Ambrosetti et al., 1997; Kuhlbrodt et al., 1998;
Soriano and Russell, 1998) and to facilitate DNA binding
(e.g., Ma et al., 1998), as well as acting as transcription
factors in their own right in the CNS (e.g., Uwanogho et al.,
1995; Soriano and Russell, 1998). The Pax gene family
members encode proteins containing a paired domain and
in some cases an octapeptide or homeodomain or both
(reviewed in Mansouri et al., 1996). Pax genes are essential
in the developing CNS and sensory systems, such as eyes
(e.g., Tomarev et al., 1997; reviewed in Stoykova and Gruss,
1994; Mansouri et al., 1996). They are also expressed during
the morphogenesis of a variety of organs (reviewed in Dahl
et al., 1997)
Using a reverse transcriptase polymerase chain reaction
(RT-PCR) approach (Degnan et al., 1997), we have dem-
onstrated that at least 2 members of each of these 3 gene
families are expressed in the cerebral and pleuropedal gan-
glia of adult H. asinina. Each of these genes displays a
unique pattern of expression in the adult, suggesting that
they have a number of developmental and physiological
roles in the tropical abalone.
MATERIALS AND METHODS
Animals and Larvae
Haliotis asinina were collected on the low tide, at night,
from the coral microatolls of the outer reef flat surrounding
Heron Island (23°278S; 151°558E). Spent adult abalone were
collected between June and August; and gravid abalone,
between October and April. Animals were maintained in
aquaria in flowing ambient (18–31°C) seawater and fed ad
libitum with the macroalgae Gracillaria spp. and Laurencia
spp. They were maintained in this way for no longer than 2
weeks.
Isolation of RNA
Cerebral and pleuropedal ganglionic complexes were dis-
sected from spent and gravid adult H. asinina and trimmed
of connecting nerves and the cerebropleural connectives.
Epipodial fringe, eye, tentacle, gill, gut, gonad, muscle, and
digestive gland were dissected from gravid adult abalone.
Total RNA was isolated from these tissues as described in
Chomczynski and Sacchi (1987). Ten ganglia were homog-
enized in 3 ml of ice-cold guanidinium extraction solution.
Nucleic acids were further purified by precipitation in 4 M
LiCl as described in Sambrook et al. (1989).
Reverse Transcriptase Polymerase Chain Reaction
Degenerate oligonucleotide primers (Table 1) were de-
signed to anneal to highly conserved codons of members of
the POU, Sox, and Pax multigene families. POU primers
were designed to anneal to codons 20 to 29 of the POU-
specific domain (F/LKQRRIKLG) and 156 to 164 of the POU
homeodomain (VVRVWFCNR). Sox primers annealed to
546 Elizabeth K. O’Brien and Bernard M. Degnan
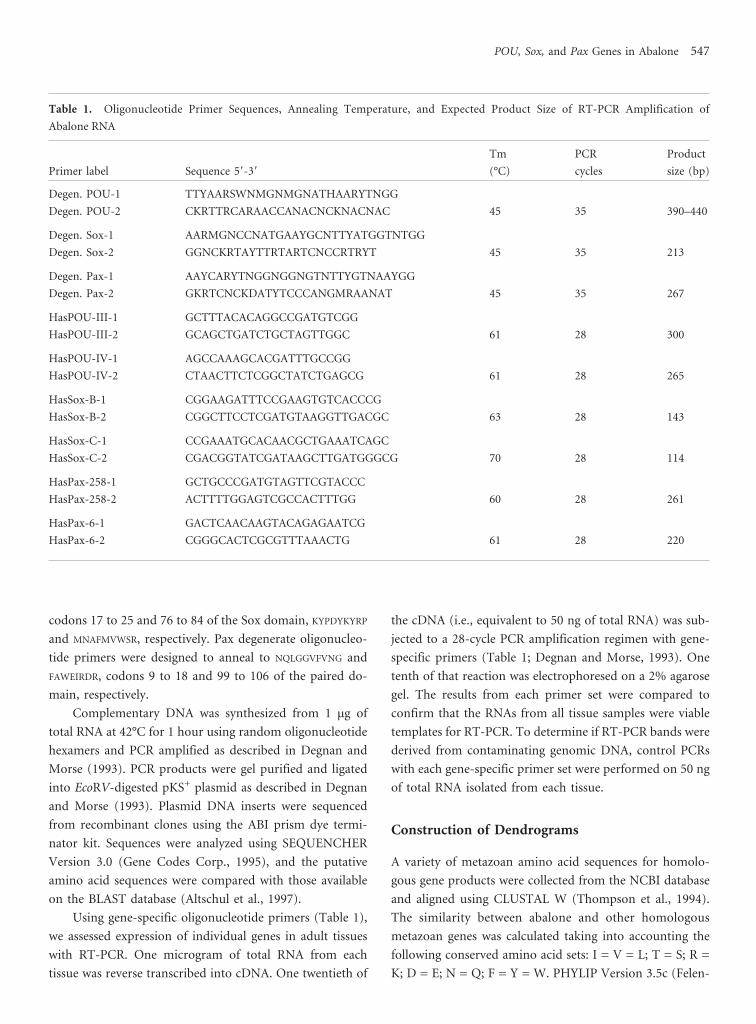
codons 17 to 25 and 76 to 84 of the Sox domain, KYPDYKYRP
and MNAFMVWSR, respectively. Pax degenerate oligonucleo-
tide primers were designed to anneal to NQLGGVFVNG and
FAWEIRDR, codons 9 to 18 and 99 to 106 of the paired do-
main, respectively.
Complementary DNA was synthesized from 1 µg of
total RNA at 42°C for 1 hour using random oligonucleotide
hexamers and PCR amplified as described in Degnan and
Morse (1993). PCR products were gel purified and ligated
into EcoRV-digested pKS+ plasmid as described in Degnan
and Morse (1993). Plasmid DNA inserts were sequenced
from recombinant clones using the ABI prism dye termi-
nator kit. Sequences were analyzed using SEQUENCHER
Version 3.0 (Gene Codes Corp., 1995), and the putative
amino acid sequences were compared with those available
on the BLAST database (Altschul et al., 1997).
Using gene-specific oligonucleotide primers (Table 1),
we assessed expression of individual genes in adult tissues
with RT-PCR. One microgram of total RNA from each
tissue was reverse transcribed into cDNA. One twentieth of
the cDNA (i.e., equivalent to 50 ng of total RNA) was sub-
jected to a 28-cycle PCR amplification regimen with gene-
specific primers (Table 1; Degnan and Morse, 1993). One
tenth of that reaction was electrophoresed on a 2% agarose
gel. The results from each primer set were compared to
confirm that the RNAs from all tissue samples were viable
templates for RT-PCR. To determine if RT-PCR bands were
derived from contaminating genomic DNA, control PCRs
with each gene-specific primer set were performed on 50 ng
of total RNA isolated from each tissue.
Construction of Dendrograms
A variety of metazoan amino acid sequences for homolo-
gous gene products were collected from the NCBI database
and aligned using CLUSTAL W (Thompson et al., 1994).
The similarity between abalone and other homologous
metazoan genes was calculated taking into accounting the
following conserved amino acid sets: I = V = L; T = S; R =
K; D = E; N = Q; F = Y = W. PHYLIP Version 3.5c (Felen-
Table 1. Oligonucleotide Primer Sequences, Annealing Temperature, and Expected Product Size of RT-PCR Amplification of
Abalone RNA
Primer label Sequence 58-38
Tm
(°C)
PCR
cycles
Product
size (bp)
Degen. POU-1 TTYAARSWNMGNMGNATHAARYTNGG
Degen. POU-2 CKRTTRCARAACCANACNCKNACNAC 45 35 390–440
Degen. Sox-1 AARMGNCCNATGAAYGCNTTYATGGTNTGG
Degen. Sox-2 GGNCKRTAYTTRTARTCNCCRTRYT 45 35 213
Degen. Pax-1 AAYCARYTNGGNGGNGTNTTYGTNAAYGG
Degen. Pax-2 GKRTCNCKDATYTCCCANGMRAANAT 45 35 267
HasPOU-III-1 GCTTTACACAGGCCGATGTCGG
HasPOU-III-2 GCAGCTGATCTGCTAGTTGGC 61 28 300
HasPOU-IV-1 AGCCAAAGCACGATTTGCCGG
HasPOU-IV-2 CTAACTTCTCGGCTATCTGAGCG 61 28 265
HasSox-B-1 CGGAAGATTTCCGAAGTGTCACCCG
HasSox-B-2 CGGCTTCCTCGATGTAAGGTTGACGC 63 28 143
HasSox-C-1 CCGAAATGCACAACGCTGAAATCAGC
HasSox-C-2 CGACGGTATCGATAAGCTTGATGGGCG 70 28 114
HasPax-258-1 GCTGCCCGATGTAGTTCGTACCC
HasPax-258-2 ACTTTTGGAGTCGCCACTTTGG 60 28 261
HasPax-6-1 GACTCAACAAGTACAGAGAATCG
HasPax-6-2 CGGGCACTCGCGTTTAAACTG 61 28 220
POU, Sox, and Pax Genes in Abalone 547
stein, 1993) was used to estimate molecular distances with
a neighbor-joining method using the Dayhoff PAM matrix
model of amino acid substitution as the distance measure.
The confidence at each node was assessed by 1000 bootstrap
pseudoreplications.
RESULTS
Sequence Analysis of POU, Sox, and Pax Genes
Using degenerate oligonucleotide primers designed to an-
neal to conserved regions of POU, Sox, and Pax genes
(Table 1), we amplified complementary DNA (cDNA) de-
rived from H. asinina cerebral and pleuropedal ganglia
RNA. Ganglia samples were separated by season thus pro-
viding 4 separate pools of RNA: cerebral and pleuropedal
ganglia from winter and summer. The inability to sex in-
dividual abalone during winter precluded the analysis of
gene expression in male and female brains. The POU, Sox,
and Pax primers successfully generated RT-PCR products,
of 390–440, 213, and 267 nucleotides, respectively, from
each of the designated pools. We purified and cloned RT-
PCR products from each RNA pool separately and se-
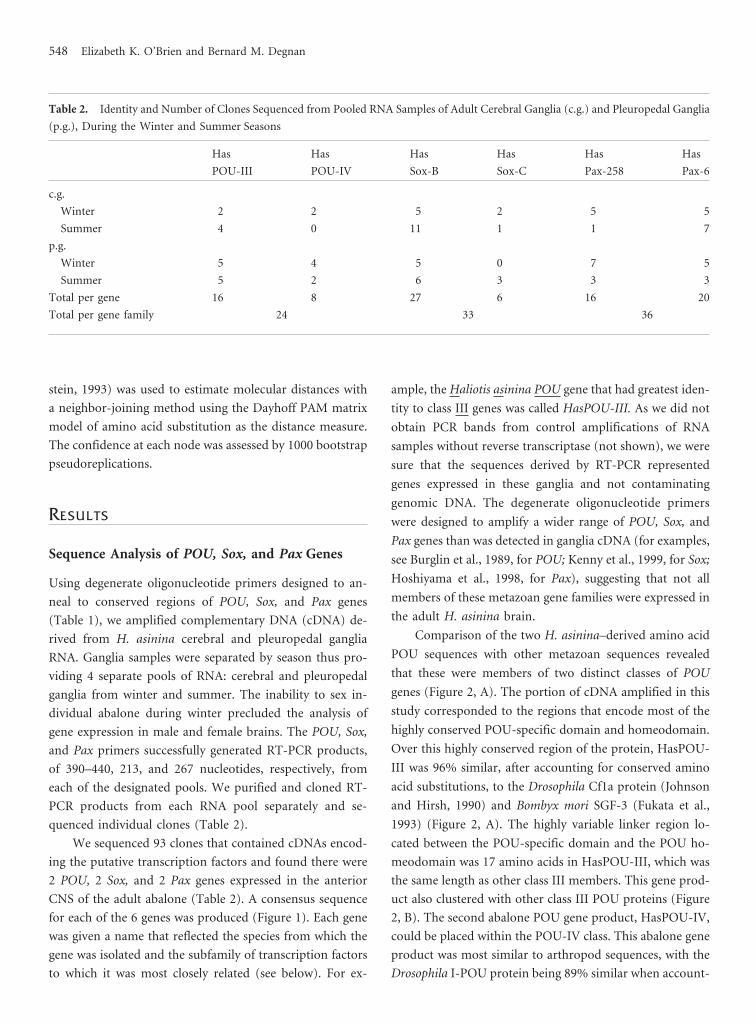
quenced individual clones (Table 2).
We sequenced 93 clones that contained cDNAs encod-
ing the putative transcription factors and found there were
2 POU, 2 Sox, and 2 Pax genes expressed in the anterior
CNS of the adult abalone (Table 2). A consensus sequence
for each of the 6 genes was produced (Figure 1). Each gene
was given a name that reflected the species from which the
gene was isolated and the subfamily of transcription factors
to which it was most closely related (see below). For ex-
ample, the Haliotis asinina POU gene that had greatest iden-
tity to class III genes was called HasPOU-III. As we did not
obtain PCR bands from control amplifications of RNA
samples without reverse transcriptase (not shown), we were
sure that the sequences derived by RT-PCR represented
genes expressed in these ganglia and not contaminating
genomic DNA. The degenerate oligonucleotide primers
were designed to amplify a wider range of POU, Sox, and
Pax genes than was detected in ganglia cDNA (for examples,
see Burglin et al., 1989, for POU; Kenny et al., 1999, for Sox;
Hoshiyama et al., 1998, for Pax), suggesting that not all
members of these metazoan gene families were expressed in
the adult H. asinina brain.
Comparison of the two H. asinina–derived amino acid
POU sequences with other metazoan sequences revealed
that these were members of two distinct classes of POU
genes (Figure 2, A). The portion of cDNA amplified in this
study corresponded to the regions that encode most of the
highly conserved POU-specific domain and homeodomain.
Over this highly conserved region of the protein, HasPOU-
III was 96% similar, after accounting for conserved amino
acid substitutions, to the Drosophila Cf1a protein (Johnson
and Hirsh, 1990) and Bombyx mori SGF-3 (Fukata et al.,
1993) (Figure 2, A). The highly variable linker region lo-
cated between the POU-specific domain and the POU ho-
meodomain was 17 amino acids in HasPOU-III, which was
the same length as other class III members. This gene prod-
uct also clustered with other class III POU proteins (Figure
2, B). The second abalone POU gene product, HasPOU-IV,
could be placed within the POU-IV class. This abalone gene
product was most similar to arthropod sequences, with the
Drosophila I-POU protein being 89% similar when account-
Table 2. Identity and Number of Clones Sequenced from Pooled RNA Samples of Adult Cerebral Ganglia (c.g.) and Pleuropedal Ganglia
(p.g.), During the Winter and Summer Seasons
Has
POU-III
Has
POU-IV
Has
Sox-B
Has
Sox-C
Has
Pax-258
Has
Pax-6
c.g.
Winter 2 2 5 2 5 5
Summer 4 0 11 1 1 7
p.g.
Winter 5 4 5 0 7 5
Summer 5 2 6 3 3 3
Total per gene 16 8 27 6 16 20
Total per gene family 24 33 36
548 Elizabeth K. O’Brien and Bernard M. Degnan

ing for conserved substitutions (Figure 2, A). The linker was
also conserved in size (16 amino acids) with others in this
class. Phylogenetic analysis grouped HasPOU-IV with other
metazoan POU class IV gene products (Figure 2, B).
Alignment of the 2 abalone partial Sox gene products
with other metazoan sequences warranted the placement of
H. asinina sequences into one of the previously established
Sox groups (groups A–F; Wright et al., 1993). The abalone
sequences encoded part of the highly conserved HMG
DNA-binding domain. HasSox-B was most similar to the
mouse Sox-2 protein (96% identity) and other group B Sox
gene products (Figure 3, A). HasSox-C was most similar
(82% similarity) to group C Sox gene products, which in-
cluded mouse Sox-4, -11, and -13 (Figure 3, A). Phyloge-
netic analysis of members of this family of transcription
factors (Figure 3, B) supported the alignment data and the
classification of the H. asinina Sox gene products in groups
B and C. A bootstrap analysis was undertaken for the Sox
gene products, but the values were very low. This was prob-
ably because of the short length of the sequence and the low
number of informative characters. Without informative
bootstrap values, the strength of the assignment of the H.
asinina Sox genes to specific groups could not be assessed.
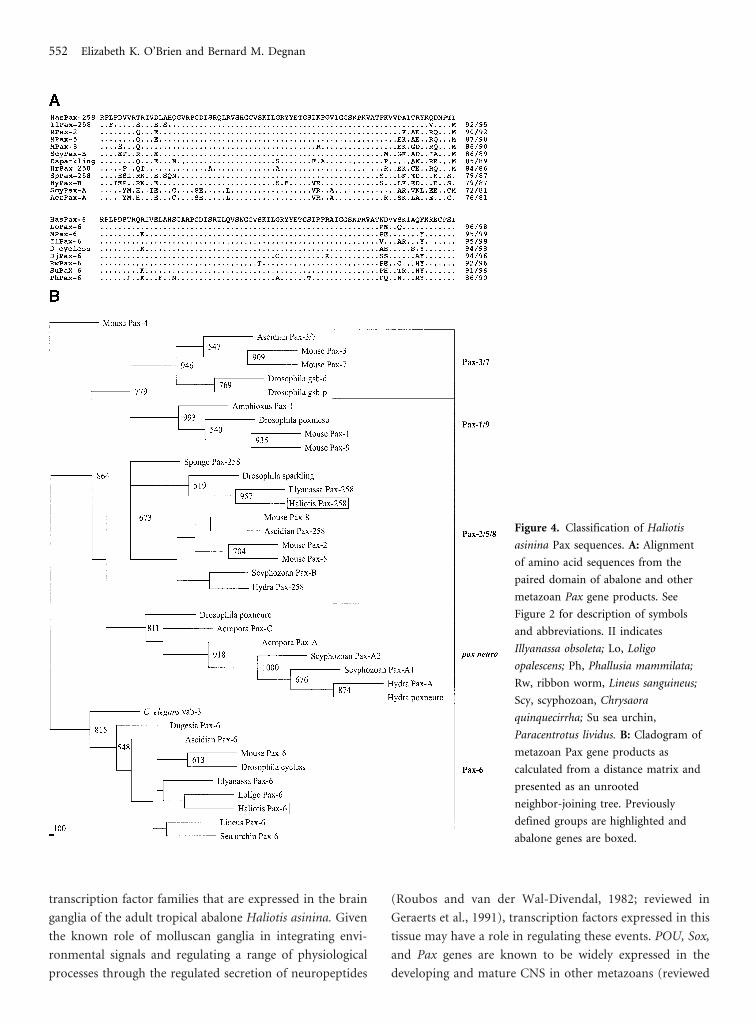
The abalone Pax-derived amino acid sequences
matched most closely to those of other mollusks. The
HasPax-258 amino acid sequence was 95% similar to the
Pax258 gene product from another gastropod, Illyanassa
obsoleta (Figure 4, A), and highly similar to mouse gene
products from this group (90%–92%). The other abalone
Pax gene product, HasPax-6, was 99% and 98% similar to
squid and mouse Pax6 amino acid sequences, respectively
(Figure 4, B). The consensus neighbor-joining tree (Figure
4, C) indicated an inherent difficulty in confidently dem-
onstrating relatedness within the Pax-6 grouping as deter-
mined by the low bootstrap values. Compared with other Pax
gene products, HasPax-6 grouped with 82% certitude with
other metazoan Pax-6 products (Figure 4, C). The Pax-258
product grouped with other Pax-258 members and was sepa-
rate from the other Pax gene classes (Balczarek et al., 1997).
Expression of POU, Sox, and Pax Genes
Using gene-specific oligonucleotide primers (Table 1), we
determined by RT-PCR amplification whether HasPOU-III,
POU-IV, Sox-B, Sox-C, Pax-258, and Pax-6 transcripts were
present in a variety of adult tissues. This approach allowed
us to determine gross levels of transcript abundance only
(i.e., presence or absence of messenger RNAs and expres-
sion at relatively high or low levels). RNAs isolated from all
adult tissues appeared to act as viable RT-PCR templates, as
each RNA sample was amplified by at least one gene-
specific oligonucleotide primer set (Figure 5). Direct PCR
amplification of equivalent amounts of RNA, not reverse
transcribed into cDNA, did not yield detectable bands with
any of the oligonucleotide primer sets (data not shown),
indicating that the bands produced in the RT-PCR ampli-
fication were derived from RNA and not contaminating
genomic DNA.
Both Has-POU genes were expressed in the cerebral
Figure 1. Consensus nucleotide sequences and derived amino
acid sequences of putative transcription factor cDNA fragments
from the adult abalone ganglia. Nucleotide and amino acid num-
bering are to the right. NCBI accession numbers: HasPOU-III,
AF231942; HasPOU-IV, AF231943; HasSox-B, AF231944; HasSox-
C, AF231945; HasPax-258, AF231946; HasPax-6, AF231947
POU, Sox, and Pax Genes in Abalone 549

and pleuropedal ganglia from adults collected from
winter and summer (Figure 5). Although not quanti-
tative, there appeared to be an increase in the levels of
HasPOU-III transcripts in both ganglia during the winter.
The other abalone POU gene, HasPOU-IV, appeared to be
expressed more consistently in the ganglia throughout the
year. HasPOU-III transcripts were also detected in the epi-
podial fringe, tentacle, eye, gill, and muscle (Figure 5), al-
though transcripts appeared to be less abundant in the gill.
HasPOU-IV was also expressed in the fringe, eye, tentacle,
and gill but was undetectable in the gut, gonad, muscle, or
digestive gland.
The 2 abalone Sox genes were expressed in the adult
ganglia but exhibited different patterns of expression in the
other adult tissues (Figure 5). Except for the gut and diges-
tive gland HasSox-B expression was detected in all other
tissues examined. HasSox-C transcripts were detected in all
tissues, although transcript abundance in the muscle ap-
peared to be lower (Figure 5).
Pax gene expression also appeared to be mainly restricted
Figure 2. Classification of Haliotis
asinina POU sequences. A:
Alignment of derived amino acid
sequence of abalone POU genes
with metazoan POU sequences.
Dots indicate identity and dashes
are gaps. The first value to the
right of the sequence is percentage
of amino acid sequence identity to
the H. asinina sequence, and the
second value is percentage of
amino acid similarity, taking into
account the following conservative
amino acid substitutions: I = V =
L, W = F = Y, T = S, D = E, R =
K, N = Q. Abbreviations are as
follows: Has, Haliotis asinina; D,
Drosophila; Bo, Bombyx mori; A,
Artemia franciscana; M, mouse,
Mus musculus; Dj, planaria,
Dugesia japonica; Hc, Herdmania
curvata; Ce, Caenorhabditis elegans.
B: Unrooted consensus tree
generated by the neighbor-joining
method from a distance matrix
bootstrapped 1000 times, using
metazoan POU amino acid
sequences. H. asinina POU gene
products are boxed. Classes
determined from the literature are
listed to the right of the tree.
550 Elizabeth K. O’Brien and Bernard M. Degnan

to adult tissues associated with the CNS including cerebral and
pleuropedal ganglia and to sensory structures such as the epi-
podial fringe, eyes, tentacles and gills, as well as in the muscle
(Figure 5). HasPax-258 mirrored HasSox-B expression in that
it was expressed in the gonad as well (Figure 5).
DISCUSSION
Using degenerate oligonucleotide primers designed to am-
plify conserved regions of members of POU, Sox, and Pax
multigene families, we identified 2 members of each of these
Figure 3. Classification of Haliotis
asinina Sox sequences. A: Alignment
of conceptual translation of H. asinina
Sox partial cDNAs with metazoan Sox
domains. See Figure 2 for description
of symbols and abbreviations. B:
Unrooted neighbor-joining tree
calculated from a distance matrix of
representative metazoan and abalone
Sox gene products; abalone genes are
boxed.
POU, Sox, and Pax Genes in Abalone 551
transcription factor families that are expressed in the brain
ganglia of the adult tropical abalone Haliotis asinina. Given
the known role of molluscan ganglia in integrating envi-
ronmental signals and regulating a range of physiological
processes through the regulated secretion of neuropeptides
(Roubos and van der Wal-Divendal, 1982; reviewed in
Geraerts et al., 1991), transcription factors expressed in this
tissue may have a role in regulating these events. POU, Sox,
and Pax genes are known to be widely expressed in the
developing and mature CNS in other metazoans (reviewed
Figure 4. Classification of Haliotis
asinina Pax sequences. A: Alignment
of amino acid sequences from the
paired domain of abalone and other
metazoan Pax gene products. See
Figure 2 for description of symbols
and abbreviations. II indicates
Illyanassa obsoleta; Lo, Loligo
opalescens; Ph, Phallusia mammilata;
Rw, ribbon worm, Lineus sanguineus;
Scy, scyphozoan, Chrysaora
quinquecirrha; Su sea urchin,
Paracentrotus lividus. B: Cladogram of
metazoan Pax gene products as
calculated from a distance matrix and
presented as an unrooted
neighbor-joining tree. Previously
defined groups are highlighted and
abalone genes are boxed.
552 Elizabeth K. O’Brien and Bernard M. Degnan

in Stoykova and Gruss, 1994; Scotting and Rex, 1996) and
have been implicated in a number of physiological changes
in mammals (e.g., Wiermann et al., 1997; Hunt and Clarke,
1999).
Molecular Evolution of POU, Sox, and Pax Genes
In order to understand the function of genes in nonmodel
animals, such as the abalone, it is important to examine
their conserved roles in other, better studied animals and
the evolutionary relationship of the genes themselves. Meta-
zoan phylogenies based chiefly on small subunit ribosomal
RNA data suggest that bilaterians can be divided into 3 great
evolutionary lineages: the deuterostomes and 2 protostome
groups, the lophotrochozoans and ecdysozoans (reviewed
in Adoutte et al., 1999; Valentine et al., 1999). The Lopho-
trochozoa include mollusks, annelids, platyhelminths, and
lophophorates and a range of minor phyla. Although there
is extensive knowledge about the role of a range of POU,
Sox, and Pax genes in ecdysozoan (i.e., Drosophila and Cae-
norhabditis elegans) and deuterostome (i.e., vertebrate) rep-
resentatives, there have been only a few reports of these
genes and their expression in mollusks and their lophotro-
chozoan relatives. These include djPOU-1, a class III POU
gene isolated from a planaria (Munoz et al., 1998), and
Pax-6 homologues in squid (Tomarev et al., 1997), planaria
(Callaerts et al., 1999), and ribbonworm (Loosli et al.,
1996).
This report identifies 6 putative transcription factors, in-
cluding the first molluscan POU genes and the first lopho-
trochozoan Sox genes. These are important additions to the
lophotrochozoan molecular database. Molecular phyloge-
netic analyses classify the H. asinina genes into known
clades within the 3 gene families, indicating the antiquity of
these gene classes within the larger multigene families. That
each of these genes is expressed in the CNS and sensory
systems of representatives of the proposed superphyla, ar-
thropods (ecdysozoan), vertebrates (deuterostone), and H.
asinina (lophotrochozoan), suggests a conserved role for
these genes in the development of the bilaterian nervous
system. These data, combined with recent reports of con-
served expression for Hox genes in the CNS of abalone and
other lophotrochozoans (e.g., Kourakis et al., 1997; Irvine
and Martindale, 2000; Giusti et al., 2000), suggest that the
genetic regulatory circuitry underlying the specification,
patterning, differentiation, and maintenance of bilaterian
nervous systems is highly conserved.
In this study we focused solely on POU, Sox, and Pax
genes expressed in the anterior ganglia of H. asinina. This
approach identified only a subset of the known members of
these gene families. In previous studies similar primer sets
have successfully amplified a wider range of gene classes
from invertebrate taxa (e.g., POU, Burglin et al., 1989; Sox,
Kenny et al., 1999; Pax, Hoshiyama et al., 1998). Combined,
these data suggest that only a subset of the total number of
POU, Sox, and Pax genes in the H. asinina genome are
expressed in adult brain ganglia.
POU Gene Expression and Evolution
Of the 6 classes of POU genes present in the bilaterians,
only 3—classes II, III, and IV—appear to be represented
in invertebrates (see Figure 2, B). Molecular phylogenetic
analyses predict that in addition to the HasPOU-III and
HasPOU-IV genes, the H. asinina genome should possess
a class II representative. However, POU-II transcripts in
invertebrates appear to be expressed only in the larval
stages (reviewed in Veenstra et al., 1997). HasPOU-III and
HasPOU-IV, as is the case with other POU-III and POU-IV
genes, are expressed in the adult CNS and several sensory
tissues (epipodial fringe, eye, cephalic tentacle, and gills).
Vertebrate, insect, and nematode homologues of
HasPOU-III play a variety of roles in the adult, including
Figure 5. RT-PCR analysis of POU, Sox, and Pax gene expression
in Haliotis asinina adult brain ganglia and tissues. Total RNA (50
ng) was RT-PCR amplified with gene-specific primers (Table 1).
One tenth of PCR amplification product was resolved on a 2%
agarose gel. PCR amplification of an equivalent amount of RNA,
not reverse transcribed into cDNA, did not produce a detectable
band in any of the tissue-primer combinations. At least one set of
primers amplified every RNA sample, indicating that RNAs from
all tissues were viable RT-PCR templates.
POU, Sox, and Pax Genes in Abalone 553

the regulation of expression of mammalian hormones
(Wierman et al., 1997; Murphy et al., 1998; Ramkumar and
Adler, 1999) and Ddc (dopa decarboxylase) in Drosophila,
which catalyzes the formation of the neurotransmitters do-
pamine and serotonin (Johnson and Hirsh, 1990).
HasPOU-IV homologues can function both as tran-
scriptional activators and as repressors in the CNS. For
example, mouse Brn-3b, a POU-IV gene, activates the ex-
pression of the estrogen receptor gene (Budhram-Mahadeo
et al., 1998) and represses a-internexin and SNAP-25 ex-
pression (Budhram-Mahadeo et al., 1995; Lakin et al., 1995;
Morris et al., 1997; Gay et al., 1998). The POU-IV Dro-
sophila homologue, I-POU, can function as an activator or
a repressor, depending on the isoform (Treacy et al., 1992).
I-POU forms a stable heterodimer with Cfla (POU class III)
preventing it from transactivating the Ddc gene (Treacy et
al., 1991). Given that class III and IV POU gene products
have been shown to interact in both mammals and insects,
there is a possibility that HasPOU-III and HasPOU-IV in
the H. asinina ganglia are operating in a similar fashion. The
titer and ratio of these factors in the nuclei of brain cells
may dictate the profile of genes that are expressed, for ex-
ample, during reproductive and nonreproductive seasons.
Changes in transcription factor concentrations as small as
2-fold to 3-fold have been shown to result in differential
regulation of target genes (Struhl et al., 1989; Kolman et al.,
1992). In the present study we did not determine relative
levels of HasPOU-III and HasPOU-IV transcripts or pro-
teins.
Expression and Possible Function of Sox Genes
The H. asinina Sox genes expressed in the brain and a wide
range of adult tissues appear to be members of the Sox B
and C groups. It is likely that the abalone genome encodes
more representatives from the Sox family, as Sox transcripts
from groups D and F are expressed in other invertebrates.
However, on the basis of the RT-PCR approach used in this
study, they do not appear to be expressed in the adult
ganglia of abalone. Although vertebrate and abalone Sox B
and C genes are expressed in a wide range of tissues (e.g.,
van der Wetering et al., 1993; Collingnon et al., 1996;
Czerny et al., 1997; Jay et al., 1997; Hunt and Clarke, 1999),
on the basis of the expression patterns of Sox homologues in
other invertebrates (Ma et al., 1998; Soriano and Russell,
1998), it appears that their conserved role may be in CNS
development and maintenance.
There are also a number of studies reporting the inter-
action of POU and Sox proteins (e.g., Yuan et al., 1995;
Ambrosetti et al., 1997; Kuhlbrodt et al., 1998; Ma et al.,
1998). For example, mouse Sox 11 (group C) interacts with
Brn 1 (POU class III) during oligodendrocyte development
(Kuhlbrodt et al., 1998). Sox2 (group B) and Oct-3 (POU
class IV) act in synergy to activate fibroblast growth factor
(FGF-4) gene expression (Yuan et al., 1995; Ambrosetti et
al., 1997). It has been also been suggested that the Dro-
sophila Sox 2 homologue Fish-hook can interact with Cfla
(POU class III) (Soriano and Russell, 1998).
Conservation of Pax Gene Expression
Pax genes are an ancient group of metazoan transcription
factors that have been detected in all taxa surveyed, includ-
ing sponges (Hoshiyama et al., 1998). There are 5 major
classes of Pax genes. In this survey, we have detected only 2
abalone Pax genes, HasPax-258 and HasPax-6, expressed in
the adult abalone brain ganglia. Homologues of these genes
play central roles in the development of metazoan nervous
and sensory systems (e.g., Czerny et al., 1997; Kozmik et al.,
1997; Pfeffer et al., 1998), and a number of these genes have
previously been isolated from mollusks; Pax-6 is expressed
during eye development in squid (Tomarev et al., 1997).
Pax genes are also expressed in the brain and reproductive
system of bilaterian adults (reviewed in Strachan and Read,
1994). As we initially isolated HasPax-258 and HasPax-6
from ganglia, we infer that these genes are playing a role in
the abalone CNS beyond its initial development. Both
HasPax-258 and HasPax-6 also are expressed in a range of
abalone tissues known to have photosensory and chemo-
sensory capabilities: epipodial fringe, eye, cephalic tentacle,
and gill. In addition, HasPax-258 is expressed in the gonad.
Mouse Pax2 and Pax5 genes are also expressed in adult
reproductive organs (Strachan and Read, 1994), suggesting
a conserved role for this gene orthologue group in animal
reproduction.
Conclusions
A number of POU, Sox, and Pax genes that are known to
play important roles in regulating gene transcription during
nervous system development of other bilaterians are ex-
pressed in the brain ganglia and sensory tissues of the tropi-
cal abalone Haliotis asinina. To date, there is a paucity of
information about the expression of these genes in mollusks
and other lophotrochozoans; we have identified the first
molluscan POU genes (HasPOU-III and HasPOU-IV) and
554 Elizabeth K. O’Brien and Bernard M. Degnan

the first lophotrochozoan Sox genes (HasSox-B and HasSox-
C). The expression of these genes in the adult brain suggests
that they may have a role in regulating the expression of
genes directly responsible for controlling growth and repro-
duction in the abalone.
ACKNOWLEDGMENTS
We thank the staff at the Heron Island Research Station for
their assistance in maintenance of abalone. This work was
supported by Australian Research council grants to B.M.D.
REFERENCES
Adoutte, A., Balavoine, G., Latillot, N., and de Rosa, R. (1999).
Animal evolution: the end of intermediate taxa? Trends Genet 15:
104–108.
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z.,
Miller, W., and Lipman, D.J. (1997). Gapped BLAST and PSI-
BLAST: a new generation of protein database search programs.
Nucleic Acids Res 25:2289–3402.
Ambrosetti, D.C., Basilico, C., and Dailey, L. (1997). Synergistic
activation of the fibroblast growth factor 4 enhancer by Sox2 and
Oct-3 depends on protein-protein interactions facilitated by a spe-
cific spatial arrangement of factor binding sites. Mol Cell Biol
17:6321–6329.
Balczarek, K.A., Lai, Z.C., and Kumar, S. (1997). Evolution and
functional diversification of the paired box (Pax) DNA-binding
domains. Mol Biol Evol 14:829–842.
Budhram-Mahadeo, V., Morris, P.J., Lakin, N.D., Theil, T., Ching,
G.Y., Lillycrop, K.A., Moroy, T., Liem, R.K.H., and Latchman, D.S.
(1995). Activation of the a-internexin promoter by the Brn-3a
transcription factor is dependent on the N-terminal region of the
protein. J Biol Chem 270:5853–5858.
Budhram-Mahadeo, V., Parker, M., and Latchman, D.S. (1998).
POU transcription factors Brn-3a and Brn-3b interact with the
estrogen receptor and differentially regulate transcriptional activ-
ity via an estrogen response element. Mol Cell Biol 18:1029–1041.
Burglin, T.R., Finney, M., Coulson, A., and Ruvkun, G. (1989)
Caenorhabolitis has scores of homeobox-containing genes. Nature
341:239–243.
Callaerts, P., Munoz Marmol, A.M., Glardon, S., Castillo, E., Sun,
H., Li, W.H., Gehring, W.J., and Salo, E. (1999). Isolation and
expression of a Pax-6 gene in the regenerating and intact planar-
ian, Dugesia(G) tigrina. Proc Natl Acad Sci USA 96:558–563.
Chomczynski, P., and Sacchi, N. (1987). Single-step method of
RNA isolation by acid-guanidinium-thiocyanate-phenol-
chloroform extraction. Anal Biochem 162:156–159.
Collignon, J., Sockananthan, S., Hacker, A., Cohen Tannoudji, M.,
Norris, D., Rastan, D., Stevanovic, M., Godfellow, P.N., and Lov-
ell-Badge, R. (1996). A comparison of the properties of Sox-3 with
Sry and two related genes, Sox-1 and Sox-2. Development 122:509–
520.
Counihan, R.T., McNamora, D.C., Souter, D.C., Jebreen, E.J., Pre-
ston, N.P., Johnson, C.R., and Degnan, B.M. (2001) Pattern, Syn-
chrony and predictability of spawning of the tropical abalone
Haliotis asinina from Heron Reef, Australia. Mar. Ecol. Prog. Ser.
(in press).
Counihan, R.T., Preston, N.P., and Degnan, B.M. (1998). The
tropical abalone, Haliotis asinina, as a model species to investigate
the molecular and cellular mechanisms controlling growth in aba-
lone. In: New Developments in Marine Biotechnology, Le Gal, Y.,
and Halvorson, H.O. (eds.). New York: Plenum Publishing, 135–
140.
Crofts, D.R. (1929). Haliotis. Liverpool Mar Biol Comm Memoirs
Typical Br Mar Plants Animals 29:1–174.
Czerny, C., Bouchard, T., Kozmik, M., and Busslinger, Z. (1997).
The characteristics of novel Pax genes of the sea urchin and Dro-
sophila reveal an ancient evolutionary origin of Pax2/5/8 subfam-
ily. Mech Dev 67:179–192.
Dahl, E., Koseki, H., and Balling, R. (1997). Pax genes and or-
ganogenesis. Bioessays 19:755–765.
Degnan, B.M., and Morse, D.E. (1993). Identification of eight
homeobox-containing transcripts expressed during larval develop-
ment and at metamorphosis in the gastropod mollusc Haliotis
rufescens. Mol Mar Biol Biotechnol 2:1–9.
Degnan, B.M., Degnan, S.M., and Morse, D.E. (1997). Muscle-
specific regulation of tropomyosin gene expression and myofibril-
logenesis differs among muscle systems examined at metamorpho-
sis of the gastropod Haliotis rufescens. Dev Genes Evol 206:464–471.
Felenstein, J. (1993). PHYLIP: Phylogenetic Inference Package. Ver-
sion 3.5C. University of Washington, Seattle.
Fukata, M., Matsuno, K., Hui, C.C., Nagata, T., Takiya, S., Xu,
P.X., Ueno, K., and Suzuki, Y. (1993). Molecular cloning of a POU
domain-containing factor involved in the regulation of the Bom-
byx sericin-1 gene J Biol Chem 268:19471–19475.
Gay, R.D., Dawson, S.J., Murphy, W.J., Russell, S.W., and Latch-
man, D.S. (1998). Activation of the iNOS gene promoter by Brn-3
POU family transcription factors dependent upon the octamer
motif in the promoter. Biochim Biophys Acta 1443:315–322.
POU, Sox, and Pax Genes in Abalone 555

Geraerts, W.P.M. (1976). The role of the lateral lobes in the con-
trol of growth and reproduction. Gen Comp Endocrinol 29:97–108.
Geraerts, W.P.M. (1986). Environmental endocrinology and the
control of reproductive season in molluscs. Adv Invert Reprod
4:151–162.
Geraerts, W.P.M., Smit, A.B., Li, K.W., Hordijk, P.L., and Joosse,
J. (1991). Molecular biology of hormones involved in the regula-
tion of reproduction and growth in molluscs. Bull Inst Zool Acad
Sinica Monogr 16:387–440.
Giusti, A.F., Hinman, V.F., Degnan, S.M., Degnan, B.M., Morse,
D.E. (2000). Expression of a Hox5/Scr gene in the larval central
nervo is system of the gastropod Haliotis, a non-segmented spira-
lian lophotrocozoan. Evol Dev 2:294–302.
Hahn, K.O. (1994). The neurosecretory staining in the cerebral
ganglia of the Japanese abalone (ezowabi), Haliotis discus hannai,
and its relationship to reproduction. Gen Comp Endocrinol 93:
295–303.
Herr, W., Sturm, R.A., Clerc, R.G., Corcoran, L.M., Baltimore, D.,
Sharp, P.A., Ingraham, H.A., Rosenfeld, M.G., Finney, M., Ruv-
kun, G., and Horvitz, H.R. (1988). The POU domain: a large
conserved region in the mammalian pit-1, oct-1, oct-2 and Cae-
norhabditis elegans unc-86 gene products. Genes Dev 2:1513–1516.
Hoshiyama, D., Suga, H., Iwabe, N., Koyanagi, M., Nikoh, N.,
Kuma Ke’i, I., Matsuda, F., Honjo, T., and Miyata, T. (1998).
Sponge Pax cDNA related to Pax-2/5/8 and ancient gene duplica-
tions in the Pax family. J Mol Evol 47:640–648.
Hunt, S.M.N., and Clarke, C.L. (1999). Expression and hormonal
regulation of the Sox4 gene in mouse female reproductive tissues.
Biol Reprod 61:476–481.
Hussain, M.A., and Habener, J.F. (1999). Glucagen gene transcrip-
tion activation mediated by synergistic interactions of Pax-6 and
Cdx-2 with the p300 co-activator. J Biol Chem 274:28950–28957.
Irvine, S.Q., and Martindale, M.Q. (2000). Expression patterns of
anterior Hox genes in the polychaete Chaetopterus: correlation
with morphological boundaries. Dev Biol 217:333–351.
Jantzen, H.M., Admon, A., Bell, S.P., and Tijan, R. (1990). Nucleo-
lar transcription factor hUBF contains a DNA-binding motif with
homology to the HMG proteins. Nature 344:830–836.
Jay, P., Sahly, I., Goze, C., Taviaux, S., Poulat, F., Couly, F., Abit-
bol, M., and Berta, P. (1997). SOX22 is a new member of the Sox
gene family mainly expressed in the human nervous tissue. Hum
Mol Genet 6:1069–1077.
Johnson, W.A., and Hirsh, J. (1990). Binding of a Drosophila
POU-domain protein to a sequence element regulating gene ex-
pression in specific dopaminergic neurons. Nature 343:467–470.
Joose, J. (1988). The hormones of molluscs. In: Endocrinology of
Selected Invertebrate Types, Laufer, H., and Downer, G.H. (eds.).
New York: Alan R. Liss, 89–140.
Kenny, A.P., Kozlowski, D., Oleksyn, D.W., Angerer, L.M., and
Angerer, R.C. (1999) Sp Sox Bl, a maternally encoded transcrip-
tion factor asymmetrically distributed amoung early sea urchin
blastomeres. Development 126:5473–5483.
Kourakis, M.J., Master, V.A., Lokhorst, D.K., Nardelli-Haefliger,
D., Wedeen, C.J., Martindale, M.Q., and Shankland, M. (1997).
Conserved anterior boundaries of Hox gene expression in the
central nervous system of the leech Helobdella. Dev Biol 190:284–
300.
Kolman, A., Bohusova, T., Lambert, B., and Simons, J.W.I.M.
(1992). Induction of 6-thioguanine-resistant mutants in human
diploid fibroblasts in vitro with ethylene oxide. Environ Mol Mu-
tagen 19:93–97.
Kozmik, Z., Czerny, T., and Busslinger, M. (1997). Alternatively
spliced insertions in the paired domain restrict the DNA sequence
specificity of Pax-6 and Pax-8. EMBO J 16:6793–6803.
Kuhlbrodt, K., Herbarth, B., Sock, E., Enderich, J., HermansBorg-
meyer, I., and Wegner, M. (1998). Cooperative function of POU
proteins and Sox proteins in glial cells. J Biol Chem 273:16050–
16057.
Lakin, N., Morris, P., Theil, T., Sato, T., Moroy, T., Wilson, M.,
and Latchman, D. (1995). Regulation of neurite outgrowth and
SNAP-25 gene expression by the Brn-3a transcription factor. J Biol
Chem 270:15858–15863.
Loosli, F., KmitaCunisse, M., and Gehring, W.J. (1996). Isolation
of a Pax-6 homolog from the ribbonworm Lineus sanguineus. Proc
Natl Acad Sci USA 93:2658–2663.
Ma, Y., Niemitz E.L., Nambu P.A., Shan, X., Sackerson, C., Fu-
jioka, M., Goto, T., and Nambu J.R. (1998). Gene regulatory func-
tions of Drosophila fish-hook, a high mobility group domain Sox
protein. Mech Dev 73:169–182.
Mangalam, H.J., Albert, V.R., Ingraham, H.A., Kapiloff, M., Wil-
son, L., Nelson, C., Elsholtz, H., and Rosenfeld, M.G. (1989). A
pituitary POU domain protein, Pit-1, activates both growth hor-
mone and prolactin promoters transcriptionally. Genes Dev 3:946–
958.
Mansouri, A., Hallonet, M., and Gruss, P. (1996). Pax genes and
their roles in cell differentiation and development. Curr Opin Cell
Biol 8:851–857.
Morris, P.J., Theil, T., Ring, C.I.A., Lillycrop, K.A., Moroy, T., and
Latchman, D.S. (1994). The opposite and antagonistic effects of
the closely related POU family transcription factors Brn-3a and
Brn-3b on the activity of a target promoter are dependent on
differences in the POU domain. Mol Cell Biol 14:6907–6914.
Morris, P.J., Dawson, S.J., Wilson, M.C., and Latchman, D.S.
(1997). A single residue within the homeodomain of the Brn-3
POU family transcription factors determines whether they activate
or repress the SNAP-25 promoter. NeuroReport 8:2041–2045.
556 Elizabeth K. O’Brien and Bernard M. Degnan

Munoz, M.M., Casali, A., Miralles, A., Bueno, D., Bayascas, J.R.,
Romero, R.L., and Salo, E. (1998). Characterization of platyhel-
minth POU domain genes: ubiquitous and specific anterior nerve
cell expression of different epitopes of GtPOU-1. Mech Dev 76:
127–140.
Murphy, D., Si-Hoe, S.L., Brenner, S., and Venkatesh, B. (1998).
Something fishy in the rat brain: molecular genetics of the hypo-
thalamo-neurohypophysial system. Bioassays 20:741–749.
Ojeda, S., Hill, J., Hill, D., Costa, M.E., Tapia, V., Cornea, A., and
Ma, Y. (1999). The Oct-2 POU domain gene in the neuroendo-
crine brain: atranscriptional regulator of mammalian puberty. En-
docrinology 140:3774–3789.
Pfeffer, P.L., Gerster, T., Lun, K., Brand, M., and Busslinger, Z.
(1998). Characterisation of three novel members of the zebrafish
Pax2/5/8 family: dependency of Pax5 and its expression on the
Pax2.1 (noi) function. Development 125:3063–3074.
Ramkumar, T., and Adler, S. (1999). A requirement for the POU
transcription factor, Brn-2, in corticotropin-releasing hormone
expression in a neuronal cell line. Mol Endocrinol 13:1237–1248.
Ryan, A.K., and Rosenfeld, M.G. (1997). POU domain family val-
ues: flexibility, partnerships, and developmental codes. Genes Dev
11:1207–1225.
Roubos, E.W., and van der Wal-Divendal, R.M. (1982). Sensory
input to growth stimulating neuroendocrine cells of Lymnaea stag-
nalis. Cell Tissue Res 227:371–386.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989). Molecular
Cloning: A Laboratory Manual. 2nd ed. Cold Spring Harbor, N.Y.:
Cold Spring Harbor Laboratory Press.
Scotting, P.J., and Rex, M. (1996). Transcription factors in early
development of the central nervous system. Neuropathol Appl
Neurobiol 22:469–481.
Soriano, N.S., and Russell, S. (1998). The Drosophila SOX-domain
protein Dichaete is required for the development of the central
nervous system midline. Development 125:3989–3996.
Stoykova, A., and Gruss, P. (1994). Roles of Pax genes in devel-
oping and adult brain as suggested by expression patterns. J Neu-
rosci 14:1395–1412.
Strachan, T., and Read, A.P. (1994). Pax genes. Curr Opin Genet
Dev 4:427–438.
Struhl, G., Struhl, K. and Macdonald, P.M. (1989). The gradient
morphogen bicoid is a concentration-dependent transcriptional
activator. Cell 57:1259–1274.
Thompson, J.D., Higgins, D.G., and Gibson, T.J. (1994). Clustal
W: improving the sensitivity of progressive multiple sequence
alignment through sequence weighting, positions-specific gap
penalties and weight matrix choice. Nucleic Acids Res 22:4673–
4680.
Tomarev, S.I., Callaerts, P., Kos, L., Zinovieva, R., Halder, G.,
Gehring, W., and Piatigorsky, J. (1997). Squid Pax-6 and eye de-
velopment. Proc Natl Acad Sci USA 94:2421–2426.
Treacy, M.N., He, X., and Rosenfeld, M.G. (1991). I-POU: a POU-
domain protein that inhibits neuron-specific gene activation. Na-
ture 350:577–584.
Treacy, M.N., Neilson, L.I., Turner, E.E., He, X., and Rosenfeld,
M.G. (1992). Twin of I-POU: a two amino acid difference in the
I-POU homeodomain distinguishes an activator from an inhibitor
of transcription. Cell 68:491–506.
Upatham, E.S., Thongkukiatkul, A., Kruatrachue, M., Wanicha-
non, C., Chitramvong, Y.P., Sahavacharin, S., and Sobhon, P.
(1998). Classification of neurosecretory cells, neurons, neuroglia
in the cerebral ganglia of Haliotis asinina, Linnaeus by light mi-
croscopy. J Shellfish Res 17:737–742.
Uwanogho, D., Rex, M., Cartwright, E.J., Pearl, G., Healy, C.,
Scotting, P.J., and Sharpe, P.T. (1995). Embryonic expression of
the chicken Sox2, Sox3 and Sox11 genes suggests an interactive role
in neuronal development. Mech Dev 49:23–36.
Valentine, J.W., Jablonski, D., and Erwin, D.H. (1999). Fossils,
molecules and embryos: new perspectives on the Cambrian explo-
sion. Development 126:851–859.
Van der Wetering, M., Oosterwegel, M., van Norren, K., and Clev-
ers, H. (1998). Sox-4, an Sry-like HMG box protein is a transcrip-
tional activator in lymphocytes. EMBO J 12:3847–3854.
Veenstra, G.J.C., Van Der Vliet, P.C., and Destree, O.H.J. (1997).
POU domain transcription factors in embryonic development.
Mol Biol Rep 24:139–155.
Wierman, M.E., Xiong, X., Kepa, J.K., Spaudling, A.J., Jacobsen,
B.M., Fang, Z., Nilaver, G., and Ojeda, S.R. (1997). Repression of
gonadotropin-releasing hormone promoter activity by the POU
homeodomain transcription factor SCIP/Oct-6/Tst-1: a regulatory
mechanism of phenotype expression. Mol Cell Biol 17:1652–1665.
Wright, E.M., Snopek, B., and Koopman, P. (1993). Seven new
members of the Sox gene family expressed during mouse devel-
opment. Nucleic Acids Res 21:744.
Yahata, T. (1971). Demonstration of neurosecretory cells in the
cerebral ganglion of the abalone, Nordotid discus. Bull Fac Fish
Hokkaido Univ 22:207–214.
Yahata, T. (1973). Induced spawning of abalone (Nordotis discus)
injected with ganglional suspensions. Bull Jpn Soc Sci Fish 39:1117–
1122.
Yuan, H.B., Corbi, N., Basilico, C., and Dailey, L. (1995). Devel-
opmental-specific activity of the FGF-4 enhancer requires the syn-
ergistic action of Sox2 and Oct-3. Genes Dev 9:2635–2645.
POU, Sox, and Pax Genes in Abalone 557





























