Embed Size (px)
Citation preview

Original Article
Expression of KITENIN in human colorectal cancer and itsrelation to tumor behavior and progressionpin_2646 210..220
Seok Lee,1* Young-A Song,1* Young-Lan Park,1 Sung-Bum Cho,1 Wan-Sik Lee,1 Jae-Hyuk Lee,2 Ik-Joo Chung,1
Kyung-Keun Kim,3 Jong-Sun Rew1 and Young-Eun Joo1
Departments of 1Internal Medicine, 2Pathology and 3Pharmacology, Chonnam National University Medical School,Gwangju, Korea
KAI1 COOH-terminal interacting tetraspanin (KITENIN)contributes to tumor invasion and metastasis in variouscancers. The aim of current study was to evaluate whetherKITENIN affects tumor cell invasion and prognosis inhuman colorectal cancers. We investigated the biologicrole of KITENIN on tumor cell invasion by using smallinterfering RNA in Caco2, DLD1, and SW480. We evaluatedthe expression of KITENIN and activator protein-1 (AP-1)target genes in human colorectal cancer tissues. Thetumor cell invasion was decreased by knockdown ofKITENIN in three tested cell lines. The mRNA expression ofcyclin D1 and COX-2 was decreased in KITENIN knock-down Caco2 and the mRNA expression of MMP-3 andCOX-2 was decreased in KITENIN knockdown DLD1 andSW480. The extracellular-signal protein kinase 1/2(ERK1/2) phosphorylation was decreased in KITENINknockdown in three tested cell lines. Expression ofKITENIN and AP-1 target genes was significantly increasedin human colorectal cancer tissues. The ERK1/2, c-JunN-terminal kinase (JNK) and p38 phosphorylations wereincreased in human colorectal cancer tissues. Expressionof KITENIN was significantly associated with lymphovas-cular invasion, depth of invasion, lymph node metastasis,tumor stage and poor survival. These results indicate thatKITENIN is associated with human colorectal cancer pro-gression including invasion and metastasis.
Key words: colorectal neoplasm, KITENIN, prognosis, tumorinvasion
Metastatic cancer remains an incurable disease. Metastasisis a complex process involving the spread of malignant tumorcells from a primary tumor site to a secondary organ andcolonization of the distant organ. The progression of cancerhas been shown to correlate directly with the gene changesof oncogenes, growth factors, and tumor suppressor genesthat regulate a number of steps responsible for metastasis .1,2
KAI1/CD82 is a 40–75-kDa tetraspanin cell surface proteinwhich suppressed experimental metastasis of rat prostatetumor cells,3 and decreased motility and invasion of coloncancer cells.4 Down regulation of KAI1/CD82 is associatedwith metastatic progression and poor prognosis in manyhuman tumor types including prostate, lung, breast, pancre-atic, and colon cancers.5–9 These data show that KAI1/CD82is an important tumor suppressor gene in cancer metastasisand progression.10,11
Recently, KAI1 COOH-terminal interacting tetraspanin(KITENIN) was identified as a binding partner of the KAI1/CD82. It associates specifically with a protein homolog toVANGL1 (Drosophila) in the COOH-terminal region of KAI1/CD82. KITENIN is also a transmembrane four protein butdoes not belong to the tetraspanin superfamily.12–15 In con-trast to KAI/CD82, KITENIN increases tumorigenecity, inva-siveness, and adhesion to fibronectin of mouse colon cellsand specific knockdown of KITENIN inhibits tumor metasta-sis in a mouse model of colon cancer.12–15 Also, intratumoraladministration of anti-KITENIN short heparin RNA-loadedbiodegradable gene carrier has shown suppressed prolifera-tion and enhanced apoptosis signal in tumors, leading todelayed tumor growth in mouse colon adenocarcinoma.16
KITENIN promotes tumor growth and pulmonary metastasisin a syngeneic mouse squamous tumor models and it repre-sents a more aggressive phenotype in a murine model of oralcavity squamous carcinoma.17,18 In bladder cancer cell lines,
Correspondence: Young-Eun Joo, MD, Department of Internal Medi-cine, Chonnam National University Medical School, 8 Hak-Dong,Dong-ku, Gwangju, 501-757, Korea.Email: [email protected]
*These authors contributed equally to this workReceived 13 September 2010. Accepted for publication 14
December 2010.© 2011 The AuthorsPathology International © 2011 Japanese Society of Pathology andBlackwell Publishing Asia Pty Ltd
Pathology International 2011; 61: 210–220 doi:10.1111/j.1440-1827.2011.02646.x

KITENIN-positive cancer cells with loss of KAI1/CD82 wereassociated with increased in vitro invasive ability.19 HighKITENIN expression was significantly associated withadvanced stage, tumor extent, and lymph node metastasis inhuman laryngeal cancer.20 Collectively, these studies suggestthat KITENIN expression may be useful as a molecularmarker for the prediction of cancer progression and specificknockdown of KITENIN may be useful as the anti-metastaticcancer gene therapy.
Cellular signaling pathways are the complex signal com-munication networks which control the host cellular functionssuch as proliferation, growth, cell movement, differentiation,senescence and death. Because of the complexities of cellsignal communication, cancer cells always show the alter-ations in multiple cellular signaling pathways. Alteration ofmitogen activated protein kinase (MAPK) signaling pathwayplays a functional role in tumor promoter-induced malignanttransformation. Activator protein-1 (AP-1) transcription factoris known to be important in tumorigenesis and tumor promo-tion, and it is influenced by the MAPK signaling pathway.Therefore, these have been described as major culprits andtherapeutic targets in various human cancers including col-orectal cancer .21,22
The aims of the current study were to evaluate whetherKITENIN affects tumor cell invasion and the cell signalingpathways responsible for tumor progression, and to examinethe relationship of its expression with various clinicopatho-logical parameters and survival in human colorectal cancers.
MATERIALS AND METHODS
Tissue samples
Twenty colorectal cancer tissues and paired normal colontissues were collected for RNA and protein preparations bycolonoscopic biopsy at Chonnam National UniversityHwasun Hospital (Jeonnam, Korea). For immunohistochem-istry, formalin-fixed and paraffin-embedded tissue samplesfrom 158 patients who underwent surgery for pathologicallyconfirmed colorectal cancers in the Chonnam National Uni-versity Hwasun Hospital (Jeonnam, Korea) between January2004 and December 2004 were obtained. No patient hadreceived preoperative chemo- or radiotherapy. Tissue blockswere selected by viewing original pathologic slides andchoosing blocks that showed the junction between normalcolon epithelium and tumor region. Tumor staging was inaccordance with the American Joint Committee on Cancer(AJCC) staging system.23 Clinical outcomes were determinedfrom the time of surgery until follow-up at December 31,2008. This study was approved by the Institutional ReviewBoard of Chonnam National University Hwasun Hospital.
Cell culture and transfection
Human colorectal cancer cell lines (Caco2, DLD1, andSW480) were obtained from the American Type CultureCollection and cultured in Dulbecco’s modified Eagle’smedium (DMEM; Hyclone, Loan, UT, USA) supplementedwith 10% fetal bovine serum (FBS; Hyclone), 50 units/mLpenicillin, and 50 mg/mL streptomycin (Gibco, Grand Island,NY, USA). Cells were incubated in a water-saturated atmo-sphere of 5% CO2 at 37°C. For experiments, cells wereseeded into plates such that they were 40–50% confluentat the time of transfection. The negative controlsiRNA (Qiagen, Germantown, MD, USA) and synthesizedhuman KITENIN small interfering RNA (siRNA) (5′-GCUUGGACUUCAGCCUCGUAGUCAA-3′) was trans-fected with Lipofectamine 2000 (Invitrogen, Carlsbad, CA,USA). The identification of KITENIN knockdown was per-formed by reverse transcriptase polymerase chain reaction(RT-PCR) and Western blotting.
Cell invasion assay
Cell invasion assay was carried out using the Transwell filterschambers (8.0 mm pores) that were coated with 1% gelatin/RPMI1640 overnight and dried out it at room temperature.Cells transfected with siRNA were seeded at 2 ¥ 105 cells in120 mL 0.2% bovine serum albumin (BSA) medium on theupper chamber. Then, 400 mL 0.2% BSA medium containing20 mg/mL human plasma fibronectin (Calbiochem, La Jolla,CA, USA), a chemotactic factor, were loaded into lowerchamber. After 24 h of incubation, cells that invaded to thebottom surface of the transwell were fixed with 70% ethanol,strained with Diff Quik solution (Sysmex, Kobe, Japan).Stained cells were then counted in five selected fields (each0.5 mm2) using hematocytometer under a light microscope.Data are expressed as mean 1 SE of the number of cells/fieldon three individual experiments.
RNA isolation and RT-PCR
Total RNA was purified with the TRIzol reagent (Invitrogen),reverse transcribed, and amplified as previously describedusing specific primers for matrix metalloproteinase-3 (MMP-3), cyclin D1, cyclooxygenase-2 (COX-2), c-myc, CD44,KITENIN, and GAPDH (Table 1). The quantity and purity oftotal RNA were determined by measuring the optical densityat 260 and 280 nm using NanoDrop ND-1000 spectropho-tometer (BCM, Houston, TX, USA). Total RNA (1 mg in 1 mLDEPC-water) of each sample was used to prepare cDNA in areaction containing 50 ng/mL oligo-dT (Promega, Madison,WI, USA) and after incubation at 72°C for 10 min, Moloney
Expression of KITENIN in human colorectal cancer 211
© 2011 The AuthorsPathology International © 2011 Japanese Society of Pathology and Blackwell Publishing Asia Pty Ltd

Murine Leukemia Virus (MMLV) reverse transcriptase (Invit-rogen) and RNAsin (Takara, Otsu, Shiga, Japan) were addedand incubation continued at 37°C for 1 h and at 72°C for15 min. PCR amplification of cDNA was performed usinggene-specific primers and GoTaq DNA polymerase(Promega). PCR cycles were limited to 30, and a primer ofGAPDH was used as an internal control. PCR products wereseparated by electrophoresis on 1% agarose gel containingethidium bromide.
Luciferase reporter assay
Cells were plated to a 24-well plate at a density of 5 ¥ 104
cells/well and the next day, cells were transfected with 20 pMof negative control and KITENIN siRNAs. After incubation for24 h, cells were retransfected with 50 ng of the reporters(AP-1-luc), 1 ng of phRL-CMV (cytomegalovirus) by Lipo-fectamine 2000 (Invitrogen). The next day, cells were lysedand then luciferase activity in 20 mL cell extract was mea-sured in each sample using a Dual-Luciferse Reporter AssaySystem (Promega). That of Renilla luciferase activity wasassayed to control for transfection efficiency. Two separateexperiments were carried out in triplicate. Relative Luciferaseactivities were normalized to Renilla activity and induction ofreporters by effectors was calculated in comparison with thenegative control siRNA.
Protein isolation and Western blotting
Total proteins were isolated using RIPA buffer (1 M Tris-HCl, 150 mM NaCl, 1% Triton X-100, 2 mM EDTA) with1 mM PMSF, Halt phosphatase inhibitor, and Halt proteaseinhibitor cocktail (Thermo, Rockford, IL, USA). Protein con-centrations of cell lysate were quantitated by BCA proteinassay (Thermo) with BSA as a standard. Total proteins (10–20 mg) were subjected to electrophoresis on 10% SDS-polyacrylamide gel and then transferred to PVDFmembranes (Millipore, Billerica, MA, USA). The membraneswere incubated for 1 h in blocking solution (5% BSA inTBS-Tween 20 buffer) and sequentially blotted with primary
antibodies; polyclonal anti-human KITENIN, extracellular-signal protein kinase 1/2 (ERK1/2), phospo-ERK1/2, c-JunN-terminal kinase (JNK), phospho-JNK, p38, phospho-p38(1:1000 dilution, Cell Signaling Technology, Danvers, MA,USA) and polyclonal anti-human GAPDH (1:2000 dilution,Santa Cruz Biotechnology, Santa Cruz, CA, USA) antibod-ies at 4°C for overnight. After rinsing in TBST, membraneswere incubated with the horseradish peroxidase-labeledanti-rabbit or anti-mouse immunoglobulins as secondaryantibody (1:5000 dilution, Santa Cruz) at RT for 1 h. Theblot was detected by chemiluminescence (ECL) HRP sub-strate (Millipore) using image reader (Ras4000, Fujifilm,Tokyo, Japan).
Immunohistochemistry
Immunostaining was performed on 3 mm tissue sectionsmounted on poly-L-lysine-coated slides and drived at 37°Covernight. After the slides were deparaffinized in xylene,rehydrated and retrieved with retrieval buffer. The endog-enous peroxidase activity was blocked with Peroxidase-Blocking solution. (Dako, Carpinteria, CA, USA). Sectionswere then incubated with 10% normal goat serum for 30 minat room temperature in order to reduce non-specific bindingand incubated with polyclonal rabbit anti-human KITENINantibody in primary diluent solution (Invitrogen) overnight at4°C. After washing in TBST, KITENIN on tissue weredetected using Dako Real Envision HRP/DAB detectionsystem (Dako). Stained tissues were also co-stained withhematoxylin (Sigma, St. Louis, MO, USA), dehydrated andviewed under a light microscope.
Scoring of KITENIN expression
The immunostaining was evaluated independently by twoobservers without knowledge of the clinical outcomes, anal-ysing the intensity, area and pattern of immunostaining. Ifthere was a discrepancy, a consensus was reached afterfurther evaluation. The staining intensity was graded on ascale of four grades: 0, no staining of cancer cells; 1, weak
Table 1 Specific primers for RT-PCR
Primers (forward/ reverse) Product size
MMP-3 5′-ACA GGA TTG TGA ATT ATA CAC-3′/ 5′-GTG GCC AAT TTC ATG AGC AGC-3′ 300 bpcyclin D1 5′-GAC CAT CCC CCT GAC GGC CGA G-3′/ 5′-CCG CAC GTC GGT GGG TCT GC-3′ 500 bpCOX-2 5′-GCA TTC TTT GCC CAG CAC TT-3′/ 5′-AGA CCA GGC ACC AGA CCA AAG-3′ 730 bpc-myc 5′-GAG GCC ACA GCA AAC CTC C-3′/5′-GGT CAC GCA GGG CAA AAA AGC T-3′ 250 bpCD44 5′-ATG GAC AAG TTT TGG TGC AC-3′/ 3′-CTT CTA TGA ACC CAT ACC TGC-3′ 250 bpKITENIN 5′-AAG CCT TCA TCC ACA TCC AGC-3′/ 5′-GAG AGC CAT CGA TCC TTG TCA-3′ 300 bpGAPDH 5′-ACC ACA GTC CAT GCC ATC AC-3′/ 5′-TCC ACC ACC CTG TTG CTG TA-3′ 452 bp
COX-2, cyclooxygenase-2; MMP, matrix metalloproteinase.
212 S. Lee et al.
© 2011 The AuthorsPathology International © 2011 Japanese Society of Pathology and Blackwell Publishing Asia Pty Ltd

staining; 2, moderate staining; 3, strong staining. The percent-age of staining area was also graded on a scale with fourgrades: 0, none; 1, < 10%; 2, 10–50%; 3, > 50%. An overallscore was calculated as the product of staining intensity andstaining area. Theoretically, the overall scores could rangefrom 0 to 9. The specimens with a score 3 4 were regarded aspositive expression and those with a score < 4 as negativeexpression.
Statistical analysis
In intergroup comparisons, the student’s t-test was used todetermine statistical significance. To compare KITENINexpression with various clinicopathological parametersbetween colorectal cancer samples, statistical analyseswere performed using the c2–test and Fisher exact test.The statistical software program used was StatisticalPackage for the Social Science (Version 15.0; SPSS,Chicago, IL, USA). A P-value < 0.05 were considered sta-tistically significant.
RESULTS
Knockdown of KITENIN diminishes invasion of humancolorectal cancer cells
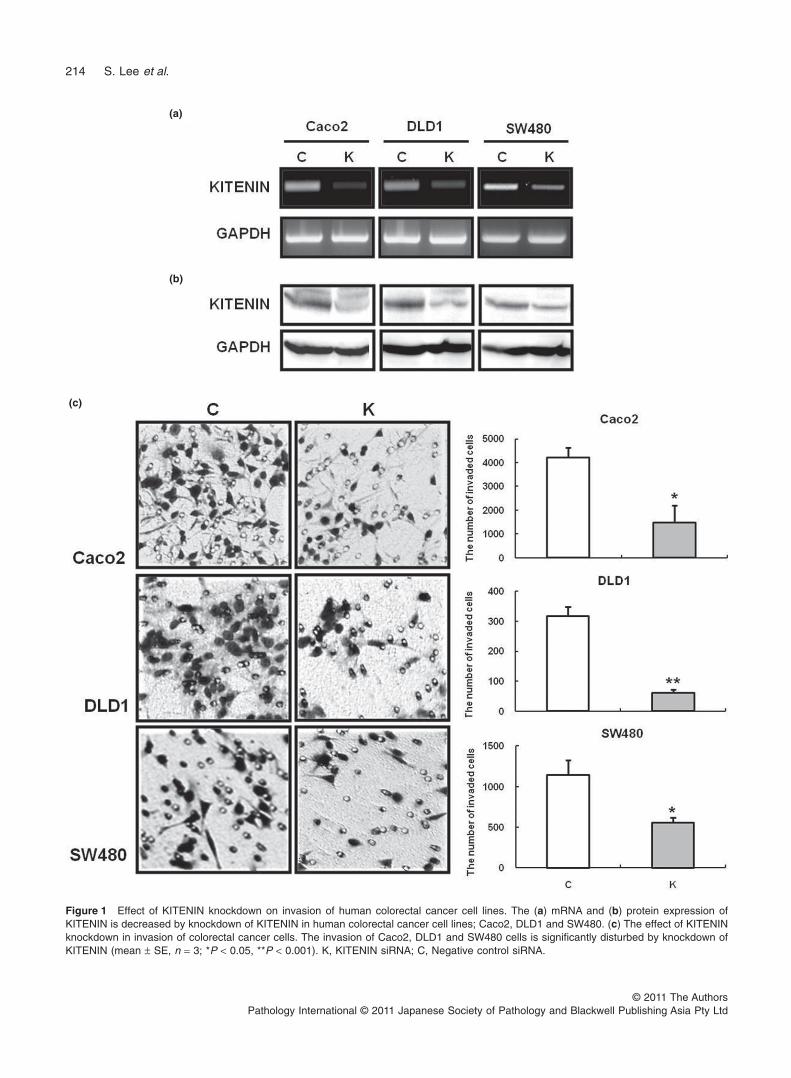
To evaluate the impact of KITENIN on invasion of humancolorectal cancer cells, we used siRNA to knockdown endog-enous KITENIN gene expression in human colorectal cancercell lines including Caco2, DLD1, and SW480. As shownin Fig. 1a and b, KITENIN gene expression in Caco2, DLD1,and SW480 cells showed a specific reduction at mRNAand protein levels by transfection of KITENIN siRNA. Thenumber of invading KITENIN siRNA-transfected Caco2,DLD1 and SW480 cells were 1490.0 1 700.3, 60.5 1 10.6, and554.7 1 63.4, whereas for the negative control cells they were4223 1 411.0, 317.5 1 30.3 and 1144.3 1 182.0 as measuredby the six random squares of the 0.5 ¥ 0.5 mm2 microscopefields under the condition of 10 mg/mL fibronectin. The differ-ence between the two was statistically significant in Caco2,DLD1 and SW480 cells (P = 0.028, <0.001, =0.037, respec-tively) (Fig. 1c).
Knockdown of KITENIN decreases activator protein-1(AP-1) transcriptional activity and expression of AP-1target genes in human colorectal cancer cells
To investigate the relationship between KITENIN and AP-1axis in human colorectal cancer cells, AP-1 target genes
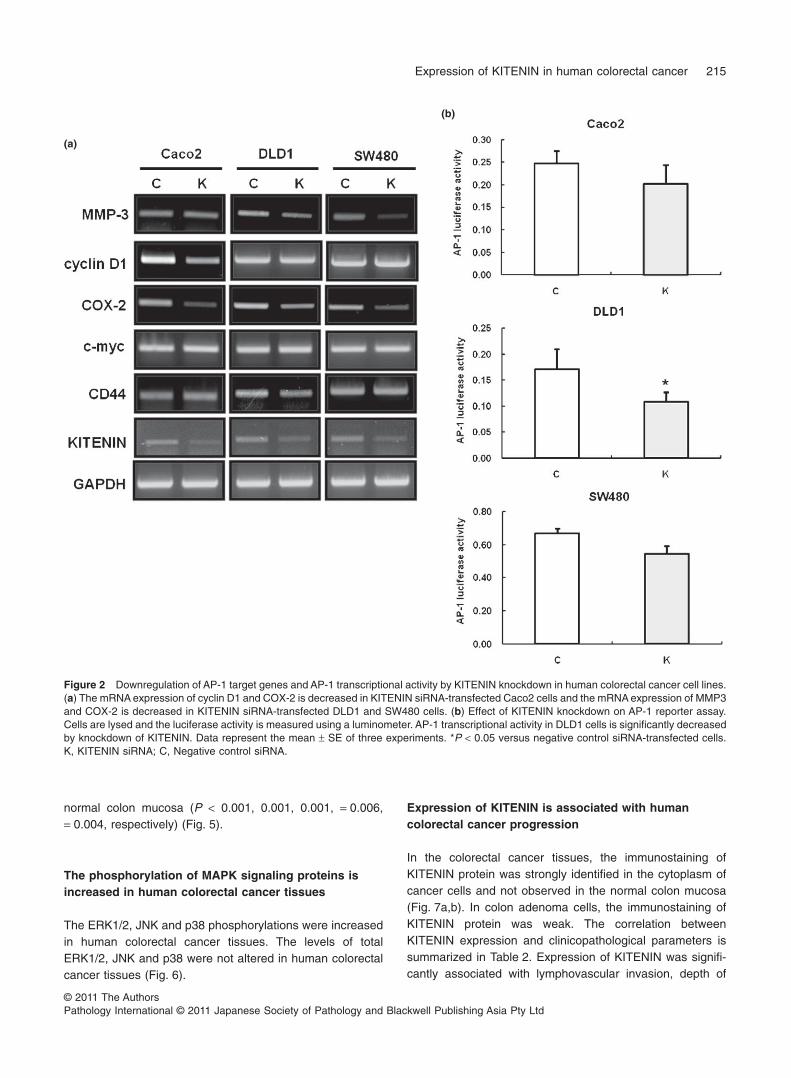
were compared between the KITENIN and negative controlsiRNA-transfected cells, such as MMP-3, cyclin D1, COX-2,c-myc and CD44. The mRNA expression of cyclin D1 andCOX-2 was decreased in KITENIN siRNA-transfected Caco2cells, and the mRNA expression of MMP-3 and COX-2 wasdecreased in KITENIN siRNA-transfected DLD1 and SW480cells (Fig. 2a). The mRNA expression of c-myc and CD44,however, was not decreased in all KITENIN siRNA-transfected colorectal cancer cells. Also, in the AP-1Luciferase reporter assay, AP-1 transcriptional activity inDLD1 cells was significantly decreased by knockdown ofKITENIN (P < 0.05) (Fig. 2b).
Knockdown of KITENIN decreases phosphorylation ofmitogen-activated protein kinase (MAPK) signalingproteins in human colorectal cancer cell lines
To dissect the signaling pathways involved in AP-1 targetgene expressions in colorectal cancer cells, we determinedthe phosphorylation levels of various up-stream signalingproteins involved in AP-1 activation using Western blotting.The ERK1/2 phosphorylation was decreased in KITENINsiRNA-transfected Caco2, DLD1 and SW480 cells. The JNKphosphorylation was decreased in KITENIN siRNA-transfected SW480 cells. The levels of total ERK1/2, JNKand p38 were not altered in colorectal cancer cells by knock-down of KITENIN (Fig. 3).
Expression of KITENIN is increased in humancolorectal cancer tissues as compared with pairednormal colon mucosa
To confirm the results of the colorectal cancer cell line study,we evaluated the expression of KITENIN at RNA and proteinlevels by RT-PCR and Western blotting in 20 colorectalcancer tissues and paired normal colon mucosa of samepatients taken by colonoscopic biopsy. We confirmedup-regulation of KITENIN expression in cancer tissues com-pared with paired normal mucosa at RNA and protein levels(P = 0.009, 0.003, respectively) (Fig. 4a,b).
Expression of AP-1 target genes is increased in humancolorectal cancer tissues as compared with pairednormal colon mucosa
Expression of AP-1 target genes including MMP-3, cyclin D1,COX-2, c-myc and CD44 was significantly increased inhuman colorectal cancer tissues as compared with paired
Expression of KITENIN in human colorectal cancer 213
© 2011 The AuthorsPathology International © 2011 Japanese Society of Pathology and Blackwell Publishing Asia Pty Ltd
(a)
(b)
(c)
Figure 1 Effect of KITENIN knockdown on invasion of human colorectal cancer cell lines. The (a) mRNA and (b) protein expression ofKITENIN is decreased by knockdown of KITENIN in human colorectal cancer cell lines; Caco2, DLD1 and SW480. (c) The effect of KITENINknockdown in invasion of colorectal cancer cells. The invasion of Caco2, DLD1 and SW480 cells is significantly disturbed by knockdown ofKITENIN (mean 1 SE, n = 3; *P < 0.05, **P < 0.001). K, KITENIN siRNA; C, Negative control siRNA.
214 S. Lee et al.
© 2011 The AuthorsPathology International © 2011 Japanese Society of Pathology and Blackwell Publishing Asia Pty Ltd
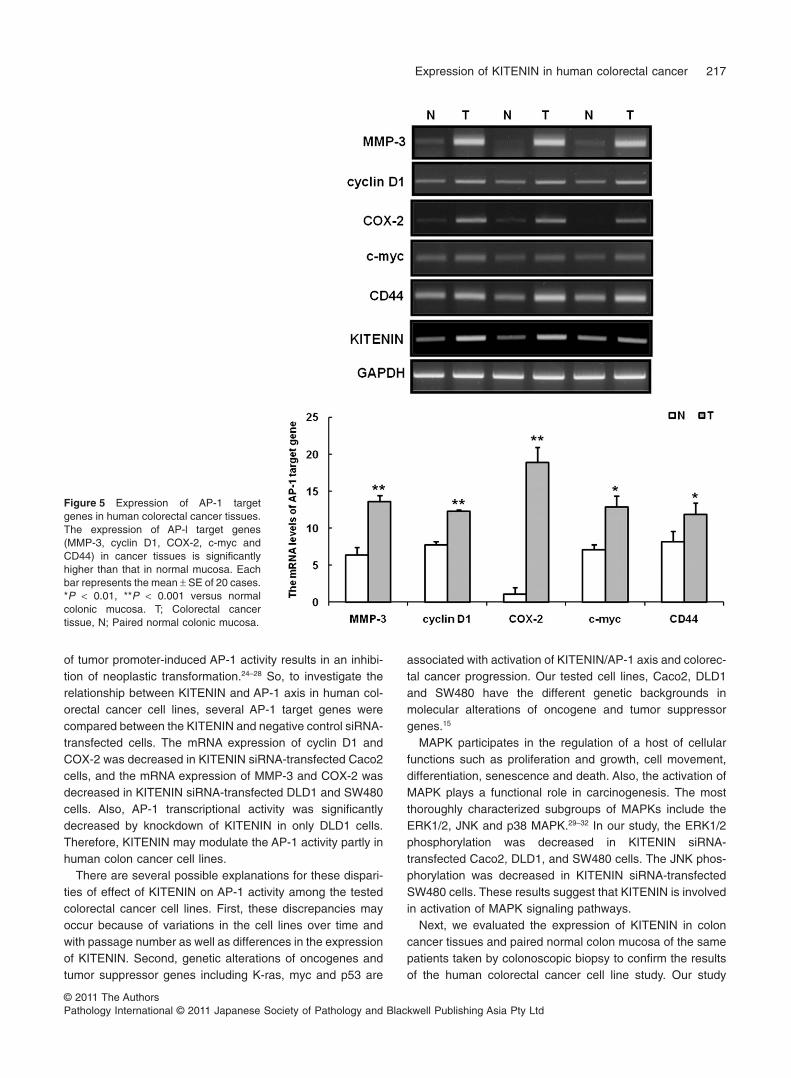
normal colon mucosa (P < 0.001, 0.001, 0.001, = 0.006,= 0.004, respectively) (Fig. 5).
The phosphorylation of MAPK signaling proteins isincreased in human colorectal cancer tissues
The ERK1/2, JNK and p38 phosphorylations were increasedin human colorectal cancer tissues. The levels of totalERK1/2, JNK and p38 were not altered in human colorectalcancer tissues (Fig. 6).
Expression of KITENIN is associated with humancolorectal cancer progression
In the colorectal cancer tissues, the immunostaining ofKITENIN protein was strongly identified in the cytoplasm ofcancer cells and not observed in the normal colon mucosa(Fig. 7a,b). In colon adenoma cells, the immunostaining ofKITENIN protein was weak. The correlation betweenKITENIN expression and clinicopathological parameters issummarized in Table 2. Expression of KITENIN was signifi-cantly associated with lymphovascular invasion, depth of
(a)
(b)
Figure 2 Downregulation of AP-1 target genes and AP-1 transcriptional activity by KITENIN knockdown in human colorectal cancer cell lines.(a) The mRNA expression of cyclin D1 and COX-2 is decreased in KITENIN siRNA-transfected Caco2 cells and the mRNA expression of MMP3and COX-2 is decreased in KITENIN siRNA-transfected DLD1 and SW480 cells. (b) Effect of KITENIN knockdown on AP-1 reporter assay.Cells are lysed and the luciferase activity is measured using a luminometer. AP-1 transcriptional activity in DLD1 cells is significantly decreasedby knockdown of KITENIN. Data represent the mean 1 SE of three experiments. *P < 0.05 versus negative control siRNA-transfected cells.K, KITENIN siRNA; C, Negative control siRNA.
Expression of KITENIN in human colorectal cancer 215
© 2011 The AuthorsPathology International © 2011 Japanese Society of Pathology and Blackwell Publishing Asia Pty Ltd

invasion, lymph node metastasis and tumor stage (P = 0.001,0.001, <0.001, <0.001, respectively) (Table 2). The overallsurvival for patients with positive KITENIN expression wassignificantly lower than for patients with negative KITENIN(P = 0.021) (Fig. 8).
DISCUSSION
The augmentation of tumor cell invasiveness is an importantaspect of tumor progression and a principal factor of cancermorbidity and mortality.1,2 KITENIN is reported to promotetumor cell migration, invasion and metastasis in cancer celllines and mouse cancer models including colon, bladder andhead and neck cancers.12–17 Our first objective for testing thethree human colorectal cancer cell lines in cell invasion assaywas to investigate whether there were differences in invasivepotentials of the cells by knockdown of KITENIN. The tumorcell invasion was decreased by knockdown of KITENIN inCaco2, DLD1 and SW480 cells. These results suggest thatthe impact of KITENIN on tumor cell invasion is not cancercell-type specific.
AP-1 is a well characterized transcription factor composedprimarily of homodimers and/or heterodimers of Fos and Junfamily proteins and its activation has been linked to a varietyof cellular processes including cellular inflammation, prolif-eration and apoptosis. In previous studies including cellculture and animal models, AP-1 was shown to have a crucialrole in neoplastic transformation and is involved in tumorpromotion, progression and metastasis. Importantly, blocking
Figure 3 Phosphorylation of MAPK signaling proteins by KITENINknockdown in human colorectal cancer cell lines. PhosphorylatedERK1/2 is decreased by transfection of KITENIN siRNA in humancolorectal cancer cell lines, Caco2, DLD1 and SW480. The phos-phorylated JNK is decreased in KITENIN siRNA-transfected SW480cells. K, KITENIN siRNA; C, Negative control siRNA.
(a) (b)
Figure 4 Expression of KITENIN in human colorectal cancer tissues. (a) KITENIN mRNA expression. (b) KITENIN protein expression.Expression of KITENIN is significantly higher in the colorectal cancer tissue than that in normal colonic mucosa at RNA and protein levels. Eachbar represents the mean 1 SE of 20 cases. *P < 0.01 versus normal colonic mucosa. T; Colorectal cancer tissue, N; Paired normal colonicmucosa.
216 S. Lee et al.
© 2011 The AuthorsPathology International © 2011 Japanese Society of Pathology and Blackwell Publishing Asia Pty Ltd
of tumor promoter-induced AP-1 activity results in an inhibi-tion of neoplastic transformation.24–28 So, to investigate therelationship between KITENIN and AP-1 axis in human col-orectal cancer cell lines, several AP-1 target genes werecompared between the KITENIN and negative control siRNA-transfected cells. The mRNA expression of cyclin D1 andCOX-2 was decreased in KITENIN siRNA-transfected Caco2cells, and the mRNA expression of MMP-3 and COX-2 wasdecreased in KITENIN siRNA-transfected DLD1 and SW480cells. Also, AP-1 transcriptional activity was significantlydecreased by knockdown of KITENIN in only DLD1 cells.Therefore, KITENIN may modulate the AP-1 activity partly inhuman colon cancer cell lines.
There are several possible explanations for these dispari-ties of effect of KITENIN on AP-1 activity among the testedcolorectal cancer cell lines. First, these discrepancies mayoccur because of variations in the cell lines over time andwith passage number as well as differences in the expressionof KITENIN. Second, genetic alterations of oncogenes andtumor suppressor genes including K-ras, myc and p53 are
associated with activation of KITENIN/AP-1 axis and colorec-tal cancer progression. Our tested cell lines, Caco2, DLD1and SW480 have the different genetic backgrounds inmolecular alterations of oncogene and tumor suppressorgenes.15
MAPK participates in the regulation of a host of cellularfunctions such as proliferation and growth, cell movement,differentiation, senescence and death. Also, the activation ofMAPK plays a functional role in carcinogenesis. The mostthoroughly characterized subgroups of MAPKs include theERK1/2, JNK and p38 MAPK.29–32 In our study, the ERK1/2phosphorylation was decreased in KITENIN siRNA-transfected Caco2, DLD1, and SW480 cells. The JNK phos-phorylation was decreased in KITENIN siRNA-transfectedSW480 cells. These results suggest that KITENIN is involvedin activation of MAPK signaling pathways.
Next, we evaluated the expression of KITENIN in coloncancer tissues and paired normal colon mucosa of the samepatients taken by colonoscopic biopsy to confirm the resultsof the human colorectal cancer cell line study. Our study
Figure 5 Expression of AP-1 targetgenes in human colorectal cancer tissues.The expression of AP-l target genes(MMP-3, cyclin D1, COX-2, c-myc andCD44) in cancer tissues is significantlyhigher than that in normal mucosa. Eachbar represents the mean 1 SE of 20 cases.*P < 0.01, **P < 0.001 versus normalcolonic mucosa. T; Colorectal cancertissue, N; Paired normal colonic mucosa.
Expression of KITENIN in human colorectal cancer 217
© 2011 The AuthorsPathology International © 2011 Japanese Society of Pathology and Blackwell Publishing Asia Pty Ltd

showed the increase of KITENIN expression in cancertissues compared with normal paired mucosa. Also, expres-sion of AP-1 target genes including MMP-3, cyclin D1,COX-2, c-myc and CD44 was significantly increased inhuman colon cancer tissues as compared with normal colonmucosa. The ERK1/2, JNK and p38 phosphorylations wereincreased in human colon cancer tissues as comparednormal colon mucosa, and AP-1 activity is induced by acti-vated MAPK signaling pathways, leading to transcription ofFos and Jun.24–28 These results suggest that KITENIN maybe associated with activation of AP-1 target genes via MAPKsignaling pathways in vivo.
Recently, high KITENIN was significantly associated withadvanced stage, tumor extent, and lymph node metastasis in
Figure 6 Phosphorylation of MAPK signaling proteins in humancolorectal cancer tissues. The phosphorylations of ERK, JNK andp38 are significantly increased in colorectal cancer tissues. T; Col-orectal cancer tissue, N; Paired normal colonic mucosa.
Figure 7 Immunohistochemical detec-tion of KITENIN expression in colorectalcancer tissues and paired normal colonmucosa. (a) Paired normal colon mucosa.(b) Colorectal cancer tissue. KITENINimmunoreactivity is strongly identified inthe cytoplasm of the tumor cells (¥200).
(a) (b)
Table 2 Correlation between the KITENIN expression and the clini-copathological parameters of colon cancer
Total(n = 158)
KITENIN
PNegative Positive(n = 64) (n = 94)
Age (years) 0.188<67 62 21 41367 96 43 53
Sex 1.000Male 94 38 56Female 64 26 38
Tumor size (cm) 0.410<5.0 94 41 5335.0 64 23 41
Location 0.425Cecum 7 4 3Ascending 22 8 14Transverse 8 3 5Descending 10 5 5Sigmoid 34 9 25Rectum 77 35 42
Stage <0.001I 18 17 1II 60 34 26III 70 11 59IV 10 2 8
Lymphovascular invasion 0.001Negative 100 50 50Positive 58 14 44
Histologic type 0.236WD 93 39 54MD 45 15 30PD 7 2 5Mucinous 12 8 4Signet 1 0 1
Depth of invasion (T) 0.001T1 6 5 1T2 20 14 6T3 124 44 80T4 8 1 7
Lymph node metastasis (N) (metastasis(N) <0.001N0 80 51 29N1-3 78 13 65
Distant metastasis (M) 0.204M0 148 62 86M1 10 2 8
MD, moderately differentiated, PD, poorly differentiated; WD, welldifferentiated.
218 S. Lee et al.
© 2011 The AuthorsPathology International © 2011 Japanese Society of Pathology and Blackwell Publishing Asia Pty Ltd

human laryngeal cancer. However, there was no difference inthe overall survival and disease-free survival between thelow- and high-KITENIN expression groups among patientswith laryngeal cancer.20 Finally, we examined the relationshipbetween expression of KITENIN and various clinicopatho-logical parameters and survival in human colorectal cancerpatients. In our study, expression of KITENIN was signifi-cantly associated with lymphovascular invasion, depth ofinvasion, lymph node metastasis and tumor stage. Moreover,overall survival for patients with positive KITENIN expressionwere significantly lower than for patients with negativeKITENIN. These results suggest that expression of KITENINis associated with human colorectal cancer progression andpoor prognosis.
In summary, although a disparity of the effect of KITENINamong the tested cell lines, the tumor cell invasion, AP-1activity and phosphorylation of MAPK signaling proteins weredecreased by knockdown of KITENIN in human colorectalcancer cell lines. The expression of KITENIN and AP-1 targetgenes was significantly increased in human colorectal cancertissues as compared with normal colon mucosa. Also, thephosphorylation of MAPK signaling proteins was increased inhuman colorectal cancer tissues as compared with normalcolon mucosa. Expression of KITENIN was significantlyassociated with lymphovascular invasion, depth of invasion,lymph node metastasis, tumor stage and poor survival. Takentogether, these results indicate that KITENIN is associatedwith human colorectal cancer progression including invasionand metastasis.
ACKNOWLEDGMENTS
This work was supported by a grant (0720570) from theNational R&D Program for Cancer Control, Ministry of Health& Welfare, Republic of Korea, and partly by a grant from theKorea Science & Engineering Foundation through the Medi-cal Research Center for Gene Regulation (R13-2002-013-04001-0) at Chonnam National University, Republic of Korea.
REFERENCES
1 Chambers AF, Groom AC, MacDonald IC. Dissemination andgrowth of cancer cells in metastatic sites. Nat Rev Cancer 2002;2: 563–72.
2 Townson JL, Chambers AF. Dormancy of solitary metastaticcells. Cell Cycle 2006; 5: 1744–50.
3 Dong JT, Lamb PW, Rinker-Schaeffer CW et al. KAI1, ametastasis suppressor gene for prostate cancer on human chro-mosome 11p11.2. Science 1995; 268: 884–6.
4 Takaoka A, Hinoda Y, Satoh S et al. Suppression of invasiveproperties of colon cancer cells by a metastasis suppressorKAI1 gene. Oncogene 1998; 16: 1443–53.
5 Dong JT, Suzuki H, Pin SS et al. Down-regulation of the KAI1metastasis gene during the progression of human prostaticcancer infrequently involves gene mutation or allelic loss.Cancer Res 1996; 56: 4387–90.
6 Adachi M, Taki T, Leki Y, Huang CI, Higashiyama M, Miyake M.Correlation of KAI1/CD82 gene expression with good prognosisin patients with non-small cell lung cancer. Cancer Res 1996;56: 1751–5.
7 Huang CI, Kohno N, Ogawa E, Adachi M, Taki T, Miyake M.Correlation of reduction in MRP/CD9 and KAI1/CD82 expres-sion with recurrences in breast cancer patients. Am J Pathol1998; 153: 973–83.
Figure 8 Kaplan-Meier survival curvecorrelating overall survival with positiveexpression (solid line) or negative expres-sion (dotted line) of KITENIN (P = 0.021).
Expression of KITENIN in human colorectal cancer 219
© 2011 The AuthorsPathology International © 2011 Japanese Society of Pathology and Blackwell Publishing Asia Pty Ltd

8 Sho M, Adachi M, Taki T et al. Transmembrane 4 superfamily asa prognostic factor in pancreatic cancer. Int J Cancer 1998; 79:509–16.
9 Hashida H, Takabayashi A, Tokuhara T et al. Clinical signifi-cance of transmembrane 4 superfamily in colon cancer. Br JCancer 2003; 89: 158–67.
10 Liu WM, Zhang XA. KAI1/CD82, a tumor metastasis suppres-sor. Cancer Lett 2006; 240: 183–94.
11 Jackson P, Marreiros A, Russell PJ. KAI1 tetraspanin andmetastasis suppressor. Int J Biochem Cell Biol 2005; 37: 530–34.
12 Lee JH, Seo YW, Park SR, Kim YJ, Kim KK. Expression of asplice variant of KAI1, a tumor metastasis suppressor gene,influences tumor invasion and progression. Cancer Res 2003;63: 7247–55.
13 Lee JH, Park SR, Chay KO et al. KAI1 COOH-terminal interact-ing tetraspanin (KITENIN), a member of the tetraspanin family,interacts with KAI1, a tumor metastasis suppressor, andenhances metastasis of cancer. Cancer Res 2004; 64: 4235–43.
14 Lee JH, Cho ES, Kim MY et al. Suppression of progression andmetastasis of established colon tumors in mice by travenousdelivery of short interfering RNA targeting KITENIN, ametastasis-enhancing protein. Cancer Res 2005; 65: 8993–9003.
15 Kho DH, Bae JA, Lee JH et al. KITENIN recruits Dishevelled/PKC delta to form a functional complex and controls the migra-tion and invasiveness of colorectal cancer cells. Gut 2009; 58:509–19.
16 Park IK, Kim KK, Cho SH et al. Intratumoral administration ofanti-KITENIN shRNA-loaded PEI-alt-PEG nanoparticles sup-pressed colon carcinoma established subcutaneously in mice.J Nanosci Nanotechnol 2010; 10: 3280–83.
17 Lee JK, Bae JA, Sun EG et al. KITENIN increases invasion andmigration of mouse squamous cancer cells and promotes pul-monary metastasis in a mouse squamous tumor model. FEBSLett 2009; 583: 711–17.
18 Lee JK, Lim SC, Kim HD et al. KITENIN represents a moreaggressive phenotype in a murine model of oral cavity squamouscarcinoma. Otolaryngol Head Neck Surg 2010; 142: 747–52.
19 Rowe A, Jackson P. Expression of KITENIN, a KAI1/CD82binding protein and metastasis enhancer, in bladder cancer celllines: Relationship to KAI1/CD82 levels and invasive behaviour.Oncol Rep 2006; 16: 1267–72.
20 Lee JK, Yoon TM, Seo DJ et al. KAI1 COOH-terminal interactingtetraspanin (KITENIN) expression in early and advanced laryn-geal cancer. Laryngoscope 2010; 120: 953–8.
21 Bode AM, Dong Z. Targeting signal transduction pathways bychemopreventive agents. Mutat Res 2004; 555: 33–51.
22 WS W. The signaling mechanism of ROS in tumor progression.Cancer Metastasis Rev 2006; 25: 695–705.
23 American Joint Committee on Cancer Classification (AJCC).Cancer Staging Manual. 6th edn, revised, Philadelphia:Lippincott-Raven, 2002; 113–23.
24 Shaulian E, Karin M. AP-1 in cell proliferation and survival.Oncogene 2001; 20: 2390–400.
25 Hasselblatt P, Gresh L, Kudo H, Guinea-Viniegra J, Wagner EF.The role of the transcription factor AP-1 in colitis-associated andbeta-catenin-dependent intestinal tumorigenesis in mice. Onco-gene 2008; 27: 6102–9.
26 Matthews CP, Colburn NH, Young MR. AP-1 a target for cancerprevention. Curr Cancer Drug Targets 2007; 7: 317–24.
27 Shaulian E. AP-1-The Jun proteins: Oncogenes or tumor sup-pressors in disguise? Cell Signal 2010; 22: 894–9.
28 Ozanne BW, Spence HJ, McGarry LC, Hennigan RF. Transcrip-tion factors control invasion: AP-1 the first among equals. Onco-gene 2007; 26: 1–10.
29 Keshet Y, Seger R. The MAP kinase signaling cascades: Asystem of hundreds of components regulates a diverse array ofphysiological functions. Methods Mol Biol 2010; 661: 3–38.
30 Pratilas CA, Solit DB. Targeting the mitogen-activated proteinkinase pathway: Physiological feedback and drug response.Clin Cancer Res 2010; 16: 3329–34.
31 Bermudez O, Pagès G, Gimond C. The dual-specificity MAPkinase phosphatases: Critical roles in development and cancer.Am J Physiol Cell Physiol 2010; 299: C189–202.
32 Haagenson KK, Wu GS. Mitogen activated protein kinase phos-phatases and cancer. Cancer Biol Ther 2010; 9: 337–40.
220 S. Lee et al.
© 2011 The AuthorsPathology International © 2011 Japanese Society of Pathology and Blackwell Publishing Asia Pty Ltd



![RNA-binding proteins in tumor progression...Human colorectal and endometrial cancer cell lines PKR pathway Promotes or inhibits cell proliferation and invasion [18– 20] hnRNP E1](https://img.pdfslide.us/doc/110x75/60f8c3a4e3d6c424af6d4c05/rna-binding-proteins-in-tumor-progression-human-colorectal-and-endometrial-cancer.jpg)
























