Embed Size (px)
Citation preview

Expression and function of Suppressor of zeste 12
in Drosophila melanogaster
Sa Chen
Department of Molecular Biology
Umeå University
901 87 Umeå
Sweden
2009

Copyright ○C 2009 Sa Chen
ISBN: 978-91-7264-729-9
Print by Arkitektkopia, Umeå, 2009

In memory of my father
怀念我的父亲



He who knows does not speak
He who speaks does not know
Lao Zi


TABLE OF CONTENTS
ABSTRACT……………………………………………………………………………. 9
LIST OF PAPERS .……………………………………………………………………10
1. REVIEW OF LITERATURE ………………………………………………..11
1.1 Drosophila as a model system ……………………………………………..11
1.2 Concepts of genes, genetics, epigenetics and chromatin ………………..11
1.2.1 Genes …………………………………………………………...…..12
1.2.2 Genetics ………………………………………………………...…..12
1.2.3 Epigenetics …………………………………………………………13
1.2.4 Chromatin …………………………………………………………..15
1.3 Epigenetic phenomena ……………………………………...………….…15
1.4 The regulation of gene expression, an epigenetic perspective …………..17
1.4.1 General introduction ………………………………………………..17
1.4.2 Epigenetic mechanisms in Prokaryotes …………………………….17
1.4.3 Epigenetic mechanisms in Eukaryotes ……………………………..19
1.4.3.1 DNA methylation ………………………………………………19
1.4.3.2 Histone modifications ………………………………………….22
1.5 Regulation of gene silencing by PcG proteins …………………………...42
1.5.1 Introduction ………………………………………………………...42
1.5.2 Genetics and biochemical characterization of PcG proteins ……….43
1.5.3 Mechanisms of PcG regulation of gene silencing ………………….54
1.5.3.1 The Polycomb Response Element (PRE) ………………………54
1.5.3.2 Gene silencing at specific loci ………………………………….56
1.5.3.3 Silencing at the chromatin level ………………………………..58
1.5.3.4 Three-dimensional (3D) organization of the nucleus …………..61
1.5.4 Evolution of PcG genes …………………………………………….61
1.5.5 PcG genes and cell cycle ……………………………………………63
1.6 Epigenetic inheritance ……………………………………………………..65
1.7 Epigenetics and disease …………………………………………………...69

2. AIM ……………………………………………………………………………70
3. RESULTS AND DISCUSSION ………………………………………….….71
4. CONCLUSIONS ………………………………………………………….…..75
5. ACKNOWLEDGEMENTS ………………………………………………….76
6. REFERENCES ………………………………………………………………..79
PAPERS I-IV

10
ABSTRACT
The development of animals and plants needs a higher order of regulation of gene
expression to maintain proper cell state. The mechanisms that control what, when and
where a gene should (or should not) be expressed are essential for correct organism
development. The Polycomb group (PcG) is a family of genes responsible for
maintaining gene silencing and Suppressor of zeste 12 (Su(z)12) is one of the core
components in the PcG. The gene is highly conserved in organisms ranging from plants
to humans, however, the specific function is not well known. The main tasks of this thesis
was to investigate the function of Su(z)12 and its expression at different stages of
Drosophila development.
In polytene chromosomes of larval salivary glands, Su(z)12 binds to about 90 specific
euchromatic sites. The binding along the chromosome arms is mostly in interbands,
which are the most DNA de-condensed regions. The binding sites of Su(z)12 in polytene
chromosomes correlate precisely with those of the Enhancer-of-zeste (E(z)) protein,
indicating that Su(z)12 mainly exists within the Polycomb Repressive Complex 2 (PRC2).
However, the binding pattern does not overlap well with Histone 3 lysine 27 tri-
methylations (H3K27me3), the specific chromatin mark created by PRC2. The Su(z)12
binding to chromatin is dynamically regulated during mitotic and meiotic cell division.
The two different Su(z)12 isoforms: Su(z)12-A and Su(z)12-B (resulting from alternative
RNA splicing), have very different expression patterns during development. Functional
analyses indicate that they also have different functions he Su(z)12-B form is the main
mediator of silencing. Furthermore, a neuron specific localization pattern in larval brain
and a giant larval phenotype in transgenic lines reveal a potential function of Su(z)12-A in
neuron development. In some aspects the isoforms seem to be able to substitute for each
other.
The histone methyltransferase activity of PRC2 is due to the E(z) protein. However,
Su(z)12 is also necessary for H3K27me3 methylation in vivo, and it is thus a core
component of PRC2. Clonal over-expression of Su(z)12 in imaginal wing discs results in
an increased H3K27me3 activity, indicating that Su(z)12 is a limiting factor for silencing.
When PcG function is lost, target genes normally become de-repressed. The segment
polarity gene engrailed, encoding a transcription factor, is a target for PRC2 silencing.
However, we found that it was not activated when PRC2 function was deleted. We show
that the Ultrabithorax protein, encoded by another PcG target gene, also acts as an
inhibitor of engrailed and that de-regulation of this gene causes a continued repression of
engrailed. The conclusion is that a gene can have several negative regulators working in
parallel and that secondary effects have to be taken into consideration, when analyzing
effects of mutants.
PcG silencing affects very many cellular processes and a large quantity of knowledge is
gathered on the overall mechanisms of PcG regulation. However, little is known about
how individual genes are silenced and how cells “remember” their fate through cell
generations.

11
LIST OF PAPERS:
I. In vivo analysis of Drosophila Su(z)12 function.
Chen S, Birve A, Rasmuson-Lestander Å.
Mol Genet Genomics (2008) 279: 159-70. Epub 2007 Nov 22.
II. Regulation of the Drosophila engrailed gene by Polycomb repressor complex
2.
Chen S, Rasmuson-Lestander Å.
Mechanisms of Development. 2009. In press.
III. The role of Suppressor of zeste 12 in cell cycle regulation.
Chen S, Rasmuson-Lestander Å.
Manuscript
IV. In vivo analysis of Suppressor of zeste 12 s different isoforms.
Chen S, Larsson A.L, Tegeling E, Rasmuson-Lestander Å.
Manuscript

12
1. REVIEW OF LITERATURE
1.1 Drosophila as a model system
The fruit fly, Drosophila melanogaster, was first brought to laboratory by William E.
Castle at Harvard University in 1901 (Kohler, 1994). It was his work that inspired
Thomas Hunt Morgan to use Drosophila as a model (in 1908) and led to Morgan's Nobel
Prize in 1933. He and his students established the linear organization of genes in
chromosomes. Since then, Drosophila has remained
at the forefront of genetics, developmental biology,
evolution biology and genomics research.
Why? Because fruit flies are easy to keep and breed.
During the past 100 years a whole range of mutants
have been used to combine classical genetic
techniques and modern molecular genetic tools, for
example, UAS-GAL4 and FLP-FRT, Chip (-on-
Chip), 3C (Chromatin, Conformation Capture), 4C
(Chromosome Conformation Capture–on-Chip) and 6C (Combined 3C-ChIP-cloning)
(Dekker, 2006; Tiwari et al., 2008a; Zhao et al., 2006) techniques have been fruitful.
Drosophila is an advanced representative of insects, an order that has undergone rapid
evolution in the course of the past 300 million years. To understand the tree of life, and
humans as a part of it, we need to understand genome evolution and gene regulation.
Drosophila is an indispensable model organism in these studies.
1.2 Concepts of genes, genetics, epigenetics and chromatin
Genetics has seen a spectacular progress as a science. The interest on inheritance goes
back to a time long before recorded history. Philosophers and scientists who have
contributed to the ideas on heredity and individual development include Hippocrates
(about 500 BC) and Aristotle (384-322 BC), who speculated about preformation and
epigenesis, respectively; Darwin, with his ideas of natural selection and pangenesis (ca
1859-1872); Mendel, with his theory of inheritance (1866); August Weissmann (1885)
with his germ plasm theory; Watson and Crick (1953) who discovered the structure of
Figure 1. Thomas Hunt Morgen

13
DNA. Accordingly, the meanings of “gene, genetics and epigenetics” have been subject
to an evolution that parallels our dramatically increasing knowledge of the mechanisms
underlying heredity and the regulation of gene expression.
1.2.1 Genes
The modern concept of the word “gene” originates from the work of Gregor Mendel
(1822-1884), who first showed the existence of a particulate hereditary material. In 1889,
Hugo de Vries invented the concept pangen which is derived from Darwin s word
pangenesis, where pan is a Greek prefix meaning “whole” and genesis means “birth” or
“origin”. Two decades later, Wilhelm Johannsen abbreviated this term to "gene" ("gen"
in Danish and German). People had, of course, used artificial selection ever since the
domestication of animals and plants. Nevertheless, the mechanism of heredity has been
subject to serious study only since the rediscovery of Mendel’s work in 1900. Since then,
the definition of the gene has followed our increasing knowledge about the hereditary
material. For example, the gene has been defined as a distinct locus (the 1910s), a
blueprint for a protein (the 1940s), a physical molecule (the 1950s), a transcribed code
(the 1960s) and an open reading frame (the 1970s-1980s). Recently, the Encyclopedia of
DNA Elements (ENCODE) Project defined the gene as “a union of genomic sequences
encoding a coherent set of potentially overlapping functional products” (Gerstein et al.,
2007).
1.2.2 Genetics
Genetics (from Greek “genesis”, “original”) was first coined in 1831 by a historian,
Thomas Carlyle (1795-1881) and the concept was extended to biology by Charles Darwin
in 1859 so that he used it in 1872 in the meaning of “Law of origination”. Later on,
William Bateson (1861-1926) put forward Genetics as the “study of heredity and
variation” (1905) as understood through Mendel´s work.
Unlike the definition of the word “gene”, the definition of genetics has not been changed
dramatically during the past 50 years. Some oversimplifications exist, such as stating that
it means the study of genes, which is incorrect. Therefore, the definition of genetics as the
study of heredity and variation is still widely accepted

14
1.2.3 Epigenetics
The history of epigenetics is linked to the study of genetics, evolution and development.
This concept has been changed dramatically. Epigenetics was first defined by Conrad H.
Waddington in the 1940s. The word is a combination of genetics and epigenesis. The root
term of epigenesis can be traced back to Aristotle who proposed it in opposition to
preformation (which means that individual development is just an increase of already
existing structures). Epigenesis, the concept that individual development involves a
differentiation of the originally undifferentiated, was shown to be the correct explanation
in the 18th
century, along with the advent of achromatic microscope and the discovery of
the germ layers in the chick embryo. In the 19th
century, epigenetics was used to describe
the working of epigenesis. However, from the middle of the 20th
century, the definition of
epigenetics acquired a new meaning. Waddington (1940) defined it as a branch of
biology which deals with the causal analysis of development. He illustrated his theory
with drawings (Fig. 2), where the developing system is depicted as a landscape, and the
bifurcating and deepening valleys run down from a plateau.
Figure 2. (A) Waddington's epigenetic landscape. (B) The interactions underlying the epigenetic
landscape. (Jablonka and Lamb, 2002).
In figure 2A, the plateau is the fertilized egg, and the path that the ball would take
represents the development route from the fertilized egg to a certain tissue or organ. In

15
figure 2B, the path, slopes, cross-sections of the valley are decided by genes (peg) and
their interactions (guy ropes). Canalization (Waddington s term for development) and
plasticity are two sides of the same coin. Even though they refer to the opposite view of
phenotypic change ability, they have one aspect in common: phenotypic variation is not
always coupled with genetic variation. This is the central point of Waddington s
epigenetics theory. So the difference between epigenetics and developmental genetics is
their different foci: epigenetics focuses on the complexity of developmental networks
with redundancy and compensatory mechanisms, while developmental genetics is more
concerned about how a gene (or genes) affects the phenotype.
In the 1990s the meaning of “epigenetics” changed again as the molecular mechanisms
controlling gene activity and the inheritance of cell phenotype began to be understood.
Holliday s work on cell memory, mainly his findings concerning DNA methylation,
made the main contribution. He narrowed down the definition of epigenetics as “The
study of the changes in gene expression, which occur in organisms with differentiated
cells, and the mitotic inheritance of given patterns of gene expression” (Holliday, 2006).
Today, the scope of this subject is much wider and I will regard this concept as modern
epigenetics. It includes studies of cellular regulatory networks that confer phenotypic
stability; changes of DNA regulated during development (such as those seen in the
immune system); cell memory mechanisms; self-propagating properties; studies of the
controlled responses of cells to genomic parasites and severe environmental effects,
which involve DNA methylation, RNA mediated gene silencing and enzyme-mediated
DNA rearrangements and repair. The position of modern epigenetics in biology is at the
junction of genetics, developmental biology and ecology. These sciences contribute
knowledge from different sides of life, and should be considered as being perfectly
compatible and complementary. The mechanisms and phenomena that I discuss later will
be based on this concept of epigenetics.
16
1.2.4 Chromatin
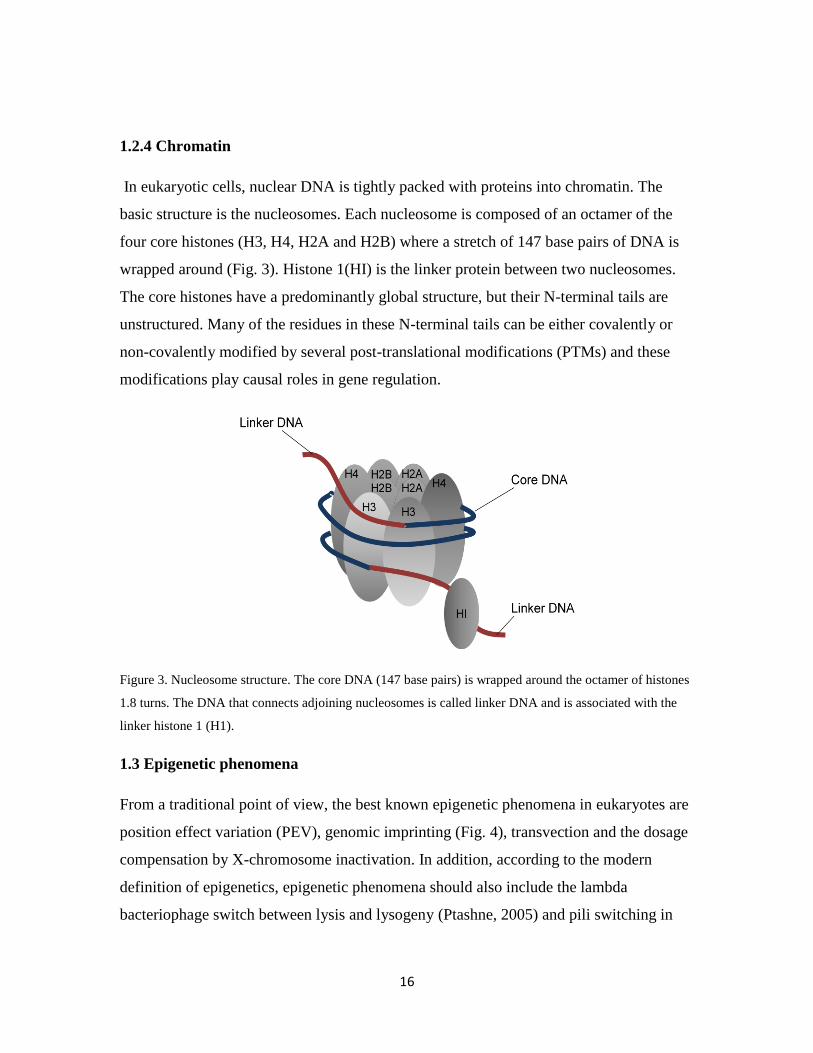
In eukaryotic cells, nuclear DNA is tightly packed with proteins into chromatin. The
basic structure is the nucleosomes. Each nucleosome is composed of an octamer of the
four core histones (H3, H4, H2A and H2B) where a stretch of 147 base pairs of DNA is
wrapped around (Fig. 3). Histone 1(HI) is the linker protein between two nucleosomes.
The core histones have a predominantly global structure, but their N-terminal tails are
unstructured. Many of the residues in these N-terminal tails can be either covalently or
non-covalently modified by several post-translational modifications (PTMs) and these
modifications play causal roles in gene regulation.
Figure 3. Nucleosome structure. The core DNA (147 base pairs) is wrapped around the octamer of histones
1.8 turns. The DNA that connects adjoining nucleosomes is called linker DNA and is associated with the
linker histone 1 (H1).
1.3 Epigenetic phenomena
From a traditional point of view, the best known epigenetic phenomena in eukaryotes are
position effect variation (PEV), genomic imprinting (Fig. 4), transvection and the dosage
compensation by X-chromosome inactivation. In addition, according to the modern
definition of epigenetics, epigenetic phenomena should also include the lambda
bacteriophage switch between lysis and lysogeny (Ptashne, 2005) and pili switching in

17
uropathogenic Escherichia coli (Hernday et al., 2002). Here, I shall only give a short
overview on PEV in Drosophila and genomic imprinting in mammals, as examples.
PEV has been observed in flies, mammals and yeast. It was first discovered in flies with a
variegated eye pigmentation phenotype by Muller in 1930 (Fig. 4A). An inversion on the
X-chromosome (In(1)wm4
) juxtaposed the white gene with centromeric heterochromatin,
which then causes repression of white gene expression in some cells. Later studies
showed that PEV can be detected in a wide range of genes, actually, all genes can show a
PEV-variegated phenotype if they are in the right rearrangement (Girton and Johansen,
2008).
Figure 4. PEV (A) and imprinting (B). Modified from (Ferguson-Smith et al., 2006; Girton and Johansen,
2008).
Imprinting is a process that causes genes to be expressed or repressed depending on their
parental origin (Fig. 4B). So far only a small number of imprinting genes has been
discovered. These genes are marked in germ cells and can “remember” their parental
origin. As a result of this their alleles are either activated or repressed. They play
important roles in prenatal growth and organ development, but also after birth, in the
regulation of metabolic pathways and in behaviors (Ferguson-Smith et al., 2006).

18
1.4. Regulation of gene expression, an epigenetic perspective
1.4.1 General principles
Some fundamental mechanisms of gene expression and regulation are conserved from
one organism to another. The central dogma of molecular biology has changed its
concept from an original simple, unidirectional pathway:
where black arrows mean the ability for DNA to replicate itself and the general procedure
for a cell to transcribe DNA to mRNA, then to translate the mRNA to protein. The gray
straight arrow indicates one special case: reverse transcriptases use a single-strand RNA
as a template to generate a double-stranded DNA copy, for example in the case of
retroviruses such as HIV, retrotransposons or telomere synthesis in eukaryotes. The gray
curved arrow over RNA shows how RNA can be self replicated into double strand RNA
via RNA-dependent RNA polymerases in the case of viruses, or in RNA silencing in
eukaryotes. The white curved arrow over protein indicates the case of prions, diseases
caused by protein conformation change.
Any particular cell type needs certain types or amounts of proteins. Most of the
specialized cells have the possibility to adjust their gene expression according to
extracellular cues. As shown in the modified central dogma above, there are many steps
from DNA to protein. That is to say, in general, gene expression can be controlled at
several levels: transcriptional control (initiation of transcription, transcription rate), RNA
processing control, RNA turnover control and translational control. However, the
transcription initiation controls appear to be the most important.
1.4.2 Epigenetic regulation in Prokaryotes
Phenotypic variation in bacteria, and in pathogenic bacteria in particular, has long been
studied. The phenomenon that the individual bacterial cell in a clonal population can have
a reversible switch of expression phase between “ON/OFF” is usually defined as phase
19
variation. Molecular mechanisms of phase variation include genetic and epigenetic
regulation. The most extensively studied epigenetic regulations are the PAP and Ag43
phase variations (van der Woude and Baumler 2004). The PAP (pyelonephritis-
associated pili) plays an important role in attachment to urinary epithelia of
uropathogenic E.coli cells. The pap operon contains a 416bp regulatory region, as well as
sites for the global regulator leucine-responsive regulator protein (Lrp), regulator proteins
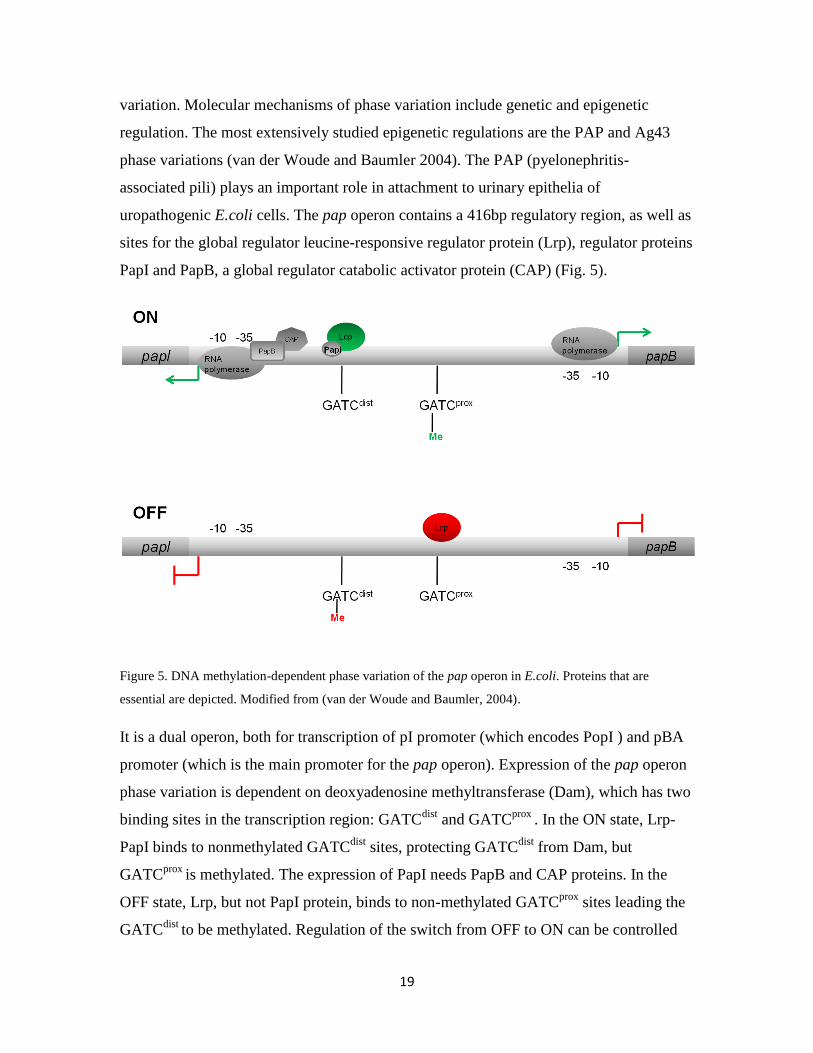
PapI and PapB, a global regulator catabolic activator protein (CAP) (Fig. 5).
Figure 5. DNA methylation-dependent phase variation of the pap operon in E.coli. Proteins that are
essential are depicted. Modified from (van der Woude and Baumler, 2004).
It is a dual operon, both for transcription of pI promoter (which encodes PopI ) and pBA
promoter (which is the main promoter for the pap operon). Expression of the pap operon
phase variation is dependent on deoxyadenosine methyltransferase (Dam), which has two
binding sites in the transcription region: GATCdist
and GATCprox
. In the ON state, Lrp-
PapI binds to nonmethylated GATCdist
sites, protecting GATCdist
from Dam, but
GATCprox
is methylated. The expression of PapI needs PapB and CAP proteins. In the
OFF state, Lrp, but not PapI protein, binds to non-methylated GATCprox
sites leading the
GATCdist
to be methylated. Regulation of the switch from OFF to ON can be controlled

20
by environmental factors, for example, the binding of cAMP-CAP enhances papBA
transcription, and the cAMP level is controlled by a carbon source in the environment.
Lrp may bend pap DNA between the CAP and the pap BA promoter to facilitate the
contact between cAMP-CAP (Goransson et al., 1989); Other environmental factors, such
as H-NS can modulate Pap gene expression and Pap switch rates.
1.4.3 Epigenetic mechanisms in eukaryotic gene regulation
1.4.3.1 DNA methylation
DNA methylation has been found in mammals, plants, fungi and bacteria. There is,
however, little evidence for any DNA methylation in Drosophila.
In vertebrates, the cytosine base in the dinucleotide sequence 5 CpG3 is covalently
modified by methylation through DNA methyltransferase enzymes (Dnmt). This CpG
methylation is a common phenomenon in the vertebrates examined thus far. From 60 to
90% of the mammalian CpG sites are methylated; they include a wide range of DNA
sequences such as satellite DNAs, repetitive elements and exons of genes. The non-
methylated CpG regions usually cluster together and are called CpG islands. Most CpG
islands mark the 5 regulatory and promoter regions of genes and 60% of the human
genes have CpG island promoters.
Molecular and genetic studies have showed that DNA methylation has a variety of
functions in gene regulation; it is mainly associated with mutagenesis, gene silencing
(genomic imprinting, X-chromosome inactivation). Two recent reports show that cycles
of DNA methylation are also involved in transcription activity (Kangaspeska et al., 2008;
Metivier et al., 2008). They found that the active X-chromosome has a higher DNA
methylation level in the transcribed region (gene body) than the inactive X-chromosome.
Before going into the mechanism of CpG methylation in regulation gene expression, I
would like to discuss first about how CpG is methylated. DNA methyltransferases
(Dnmts) catalyze genome-wide DNA methylation (Table 1). There are two types of CpG
methylation: initiation methylation and maintenance methylation (Fig. 6). Initiation
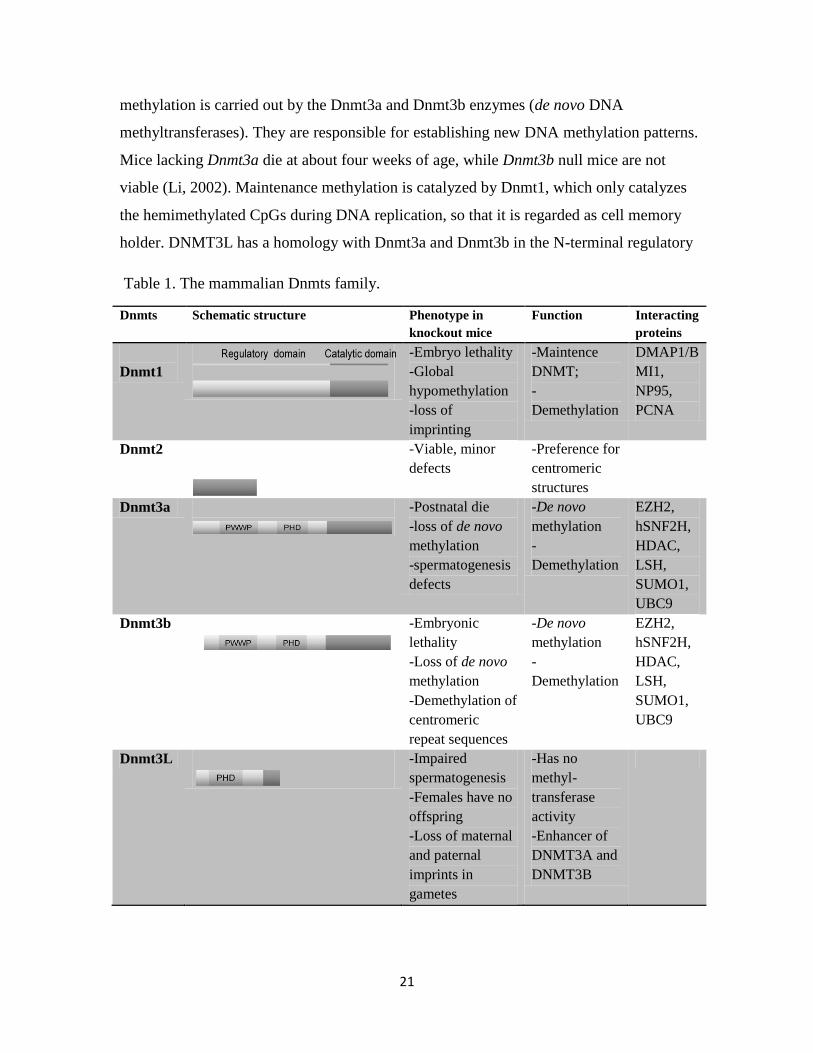
21
methylation is carried out by the Dnmt3a and Dnmt3b enzymes (de novo DNA
methyltransferases). They are responsible for establishing new DNA methylation patterns.
Mice lacking Dnmt3a die at about four weeks of age, while Dnmt3b null mice are not
viable (Li, 2002). Maintenance methylation is catalyzed by Dnmt1, which only catalyzes
the hemimethylated CpGs during DNA replication, so that it is regarded as cell memory
holder. DNMT3L has a homology with Dnmt3a and Dnmt3b in the N-terminal regulatory
Table 1. The mammalian Dnmts family.
Dnmts
Schematic structure Phenotype in
knockout mice
Function Interacting
proteins
Dnmt1
-Embryo lethality
-Global
hypomethylation
-loss of
imprinting
-Maintence
DNMT;
-
Demethylation
DMAP1/B
MI1,
NP95,
PCNA
Dnmt2
-Viable, minor
defects
-Preference for
centromeric
structures
Dnmt3a
-Postnatal die
-loss of de novo
methylation
-spermatogenesis
defects
-De novo
methylation
-
Demethylation
EZH2,
hSNF2H,
HDAC,
LSH,
SUMO1,
UBC9
Dnmt3b
-Embryonic
lethality
-Loss of de novo
methylation
-Demethylation of
centromeric
repeat sequences
-De novo
methylation
-
Demethylation
EZH2,
hSNF2H,
HDAC,
LSH,
SUMO1,
UBC9
Dnmt3L
-Impaired
spermatogenesis
-Females have no
offspring
-Loss of maternal
and paternal
imprints in
gametes
-Has no
methyl-
transferase
activity
-Enhancer of
DNMT3A and
DNMT3B

22
region. It has no methyltransferase activity on its own, but it enhances Dnmt3a and
Dnmt3b activity, and is required for establishing genomic imprints.
The DNA methylation patterns are established in the embryo and maintained through
several cell divisions, and therefore constitute a cellular memory system (Riggs, 1975)
(Fig 6).
Figure 6. Initiation methylation and maintenance methylation of DNA. Unmethylated DNA is shown as
black vertical bars, the newly synthesized DNA strand is shown as the red bars, methylated CpG pairs are
shown in red bars with pink dots. Unmethylated DNA is methylated de novo by Dnmt3a and Dnmt3b.
During cell division, the newly synthesized DNA strand is unmethylated, forming a semimethylated
double strand DNA. The methylation of semi-methylated sites to fully methylated ones is carried out by
Dnmt1.
DNA methylation is associated with transcriptional silencing, either through interference
with transcription factors binding (Fig. 7) or through recruitment of repressors (which
include histone tail modification or chromatin remodeling factors). A number of
transcription factors recognize and can bind to an unmethylated CpG sequence, but

23
cannot bind to a methylated CpG sequence. Examples include the CTCF (CCCTC
binding factor) protein in imprinting at the H19/Ifg2 locus in mice (Bell and Felsenfeld,
2000) (Fig. 7).
Figure 7. CTCF mediated maternal imprinting regulation of Igf2 through an interaction with SUZ12.
CTCF binds to the unmethylated ICR and the Igf2 promoter on the maternal allele and forms an
intrachromosomal loop through CTCF dimerization. CTCF recruits the PRC2 complex via SUZ12, which
results in histone methylation and formation of an inactive chromatin configuration around the Igf2
promoter. On the paternal allele, the CTCF cannot bind to methylated ICR, so, it cannot form a CTCF-
CTCF dimer, resulting in an active Igf2 promoter. Modified from (Li et al., 2008)
1.4.3.2 Histone modifications
Regulation of gene expression occurs at the level of individual genes (DNA sequence)
level, chromosome regions and even on entire chromosomes. The chromatin structure is
dynamic and affects gene activity, depending on the cellular signals and/or environmental
cues. ATP-dependent chromatin remodeling complexes, reversible histone modifications
and histone variants are the main causal reasons for the changes of chromatin structure.
Here I shall focus on the function of histone modifications and histone variants in gene
regulation.

24
Eight types of post-translational modification (PTM) have been found on histones (Fig.
8): acetylation/deacetylation, methylation/demethylation and phosphorylation are those
that have small chemical group modifications (covalent modifications), while
ubiquitylation, sumoylation, ADP ribosylation, deimination and proline isomerization
modifications include larger chemical modifications.
Figure 8. Some of the histone post-translational modifications (PTMs). They include
methylation/demethylation, acetylation/deacetylation, phosphorylation, ubiquitination.
Acetylation The N-terminal tails of the core histones are rich in lysines, and thus are,
accordingly, positively charged. This allows an intimate interaction with either the
negatively charged backbone of the DNA and/or with adjacent nucleosomes, which then
lead to a “tight” chromatin formation. Acetylation neutralizes this charge and weakens
the interactions, resulting in a looser chromatin structure which facilitates the binding of
transcription factor. In general, actively transcribed chromatin regions are associated with
hyperacetylation and histone acetyl-transferases (HATs) recruitment, while histone
deacetylases (HDACs) are thought to promote the return to a repressive, higher-order
25
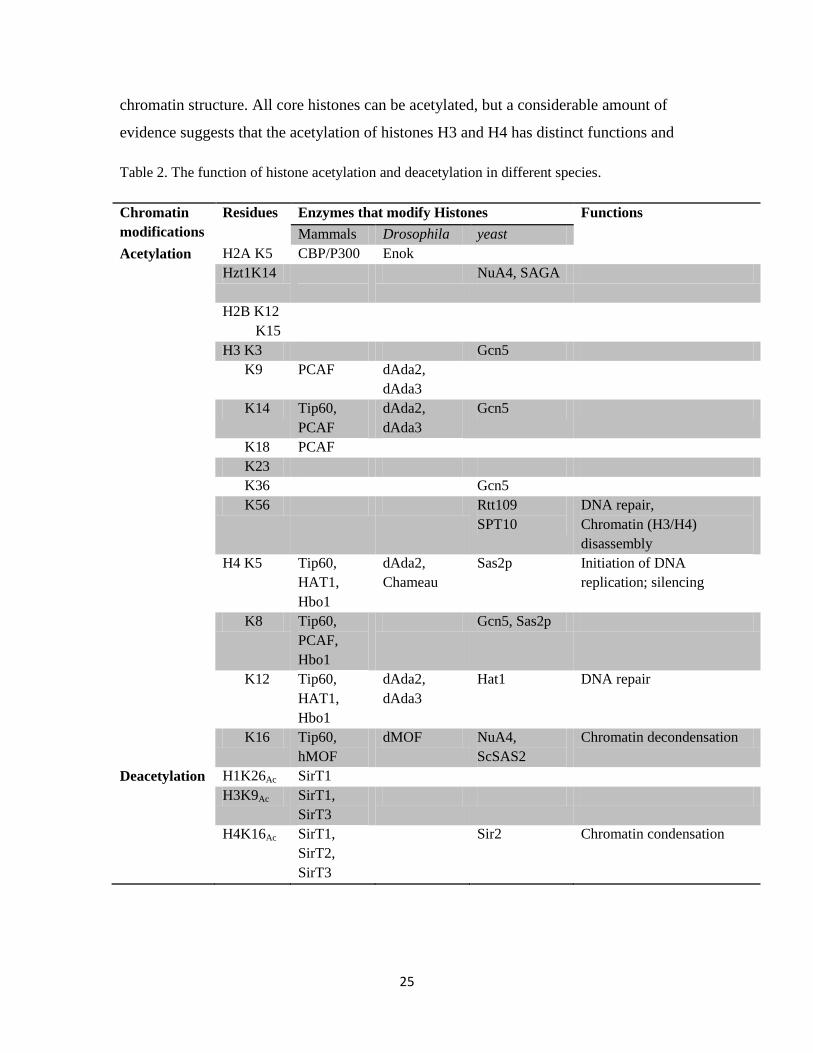
chromatin structure. All core histones can be acetylated, but a considerable amount of
evidence suggests that the acetylation of histones H3 and H4 has distinct functions and
Table 2. The function of histone acetylation and deacetylation in different species.
Chromatin
modifications
Residues Enzymes that modify Histones Functions
Mammals Drosophila yeast
Acetylation
H2A K5 CBP/P300 Enok
Hzt1K14
NuA4, SAGA
H2B K12
K15
H3 K3 Gcn5
K9 PCAF dAda2,
dAda3
K14 Tip60,
PCAF
dAda2,
dAda3
Gcn5
K18 PCAF
K23
K36 Gcn5
K56 Rtt109
SPT10
DNA repair,
Chromatin (H3/H4)
disassembly
H4 K5 Tip60,
HAT1,
Hbo1
dAda2,
Chameau
Sas2p Initiation of DNA
replication; silencing
K8 Tip60,
PCAF,
Hbo1
Gcn5, Sas2p
K12 Tip60,
HAT1,
Hbo1
dAda2,
dAda3
Hat1 DNA repair
K16 Tip60,
hMOF
dMOF NuA4,
ScSAS2
Chromatin decondensation
Deacetylation H1K26Ac SirT1
H3K9Ac SirT1,
SirT3
H4K16Ac SirT1,
SirT2,
SirT3
Sir2 Chromatin condensation

26
temporal patterns: histone H3 acetylation seem to be involved in gene expression, while
histone H4 acetylation seems to be more important in histone deposition to newly
synthesized DNA in the S phase of cell division and chromatin structure (Vaquero et al.,
2007). The acetylation/deacetylation of histones is catalyzed by HATs or HDACs. The
balance between this dynamic equilibrium is a crucial component in generating a proper
chromatin state as part of the response to environmental changes (Vaquero et al., 2007).
HAT proteins can acetylate the lysines on all four core histones, but different enzymes
have their specific substrates (Table 2). There are three HAT families: GNATs (Gcn5
related N-acetyltransferase) family, which mainly targets H3 as its substrate; MYST
proteins, which mainly target H4 and CBP/p300 (CREB-binding protein/E1A-associated
protein of 300kD), which targets both H3 and H4 (Allis, C.D, epigenetics, 2006).
The GNATs are an enormous super family (about 10,000 members) of enzymes that are
universally distributed in nature and that use acyl-CoA to acylate their cognate substrates
(Vetting et al., 2005). Histone acetyltransferases are part of this family. The first HAT
shown to be involved in transcription regulation was Hat A from Tetrahymena (Brownell
et al., 1996), a homologue of yeast Gcn5. Humans express two homologs: Gcn5 and
PCAF (p300/CBP associated factor). Gcn5/PCAF can acetylate H3K9, K14, and K18.
Gcn5 and CBP/p300 have a bromodomains, which can specifically bind to acetylated
lysines. CBP/p300 proteins are unique to metazoans.
The MYST (Moz, YBF2, Sas2p, Tip) protein family is defined by a highly conserved
histone acetyl-transferase domain, the MYST domain. Five proteins have been identified
in mammals: Querkipf (Qkf), Monocytic leukemia zinc finger protein (Moz), males
absent on the first (Mof), HIV tat-interacting protein 60 (Tip60) and histone acetyl-
transferase bound to ORC (Hbo1) (Thomas and Voss, 2007).
Qkf is required in neural stem cells, while Moz is essential for hematopoietic stem cells.
They are closely related and belong to the same group in the MYST family. Moz and Qkf
have several highly conserved domains: the PHD-type zinc fingers domain (which can
bind to H3K4me3), the MYST domain (which has the acetyl-transferase activity), an
acidic region in the central portion of the proteins, and serine-rich and methionine-rich

27
domains. The Drosophila protein Enok (enok is a mushroom) displays similarities in the
MYST domain and N-terminal compared to Moz. Enok is important in the Drosophila
mushroom body development (Scott et al., 2001)
Mof is closely related to Tip60. They share a conserved chromodomain and a MYST
domain. The chromodomain in Tip60 is found to bind to methylated histone tails, while
the Drosophila Mof chromodomain was found to mediate binding to a noncoding RNA
(Akhtar et al., 2000). In Drosophila, Mof is one of the catalytic subunits (histone
acetylation (H4K16)) of the male sex lethal (MSL) complex, which regulates the X-
chromosome gene expression. Tip60 has a broad range of functions: for example, it has
been shown to be involved in cell cycle control (by interacting with c-Myc) (Frank et al.,
2003; Patel et al., 2004), and in the regulation of apoptosis (by acting as coactivator of
p53 and NF-B) (Gaughan et al., 2002; Legube et al., 2004). Hbo1 in mammalian has
multiple functions; it is involved as an activator, in the initiation of DNA replication. On
the other hand, it can also interact with the androgen receptors where it acts as a repressor
of transcription. The Drosophila homologue of Hbo1 is Chameau. It has been shown to
interact with the origin recognition complex (Orc1), which not only binds to DNA
replication initiation sites but also interacts with the Heterochromatin protein 1 (Hp1)
(Shareef et al., 2001). In addition, a full Chameau activity is needed to maintain Hox
gene silencing mediated by Polycomb group proteins (PcG) (Grienenberger et al., 2002).
Deacetylation Histone deacetylases (HDACs) catalyze the removal of acetyl groups
from the N-terminal tails of histone proteins. There are two families: Rpd3/Hda1 family
and sirtuin family. In humans, the Rpd3/Hda1 family contains HDAC1, -2,-3, -8 (class I,
similar to yeast Rpd3 ); HDAC4, -5, -6, -7, -9, -10 (class II, similar to yeast Hda1); and
HDAC11 (class IV). The sirtuin family has seven members in humans: SIRT1-7 (class III,
similar to yeast Sir2).
Methylation Histone methylation contributes to transcriptional regulation, the
maintenance of genome integrity and epigenetic inheritance. It is also the most complex

28
covalent modification among modifications. Methylation can not only occur at different
residues (lysines, arginines) in different histone tails, but can also give rise to a different
status of methylation (mono-, di- or tri-methylation) occurs at the same residue, and these
different modifications may contribute to differential gene activity depending on the
affinity of binding proteins. (Table 3).
Table 3. The function of histone methylation and demethylation.
Residues Catalyzing enzymes Functions
Methylation
H3K4 SET1/Compass KMT2)
MLL1 (KMT2A)
MLL2(KMT2B)
MLL3 (KMT2C)
MLL4 (KMT2D)
hSet1A (KMT2E)
hSet2B (KMT2F)
ASH1 (KMT2G)
SET7/9 (KMT7)
Activation of transcription
Activation of transcription
Activation of transcription
Activation of transcription
Activation of transcription
Activation of transcription
Activation of transcription
N/D
N/D
H3K36 ySet2 (KMT3)
SET2 (KMT3A)
NSD1 (KMT3B)
SYMD2 (KMT3C)
Elongation form of Pol II
Elongation form of Pol II
N/D
Transcription activation
H3K79 yDot1 (KMT4)
DOT1L (KMT4A)
Activation of transcription
Activation of transcription
H3K9 Su(var)3-9(D)/ Clr4(fy)
(KMT1)
SUV39H1 (KMT1A)
SUV39H2 (KMT1B)
G9a (KMT1C)
EuHMTase (KMT1D)
SetDB1 (KMT1E)
CLL8 (KMT1F)
RIZ1 (KMT8)
Heterochromatin formation silencing
Heterochromatin formation silencing
Heterochromatin formation silencing
Euchromatic H3K9 methylation
Heterochromatin formation silencing
Transcriptional repression
N/D
Transcriptional repression

29
H3K27 EZH2 (KMT6)
E(z)(D)
Polycomb silencing
H4K20
sp Set9 (KMT5)
Pr-SET7/8 (KMT5A)
SUV4-20H1 (KMT5B)
SUV4-20H2 (KMT5BC)
DNA repair
Transcriptional repression
DNA repair
N/D
H4R3 PRMT1
PRMT7
Activation of transcription
Imprinting in male germ cell
H3R2, R17, R26 PRMT4, CARM1 Activation of transcription
H3R8, H4R3 PRMT5 Repression of transcription
Demethylation
H3K4me1/2 LSD1 Repression of transcription
H3K4me2/3 Yjr119Cp(y), JARID1a-d(y)
H3K9me1/2 LSD1-AP Activation of transcription
H3K9me1/2 JHDM2A Activation of transcription
H3K9me3 JHDM3A, JMJD2 Activation of transcription
H3K36me1/2 JHDM1
H3K36me3 JHDM3A, JMJD2A
H3K27me2/3 UTX, JMJD3 Activation of transcription
Histone lysine methylation In most eukaryotes, histone methylation patterns regulate
chromatin architecture and function. H3K4 methylation demarcates euchromatin. All
histone methyltransferases, except Dot1, contain a SET domain, which has catalytic
activity. Six of the methylated substrates in histones have been well studied: H3 (K4, K9,
K27, K36, K79) and H4(K20). Among them, H3K4, H3K36, H3K79 are in general
related to the activation of transcription, while the others are related to repression.
H3K4 methylation was first discovered in trout testes chromatin (Honda et al., 1975a;
Honda et al., 1975b). It is associated with gene activation. This residue can be mono-, di-
or trimethylated. High levels of H3K4me3 are associated with the 5 regions of virtually
all active genes and that there is a strong positive correlation between H3K4me3 and

30
transcription rates, RNA pol II, and histone acetylation (Ng et al., 2003; Santos-Rosa et
al., 2002; Schubeler et al., 2004). H3K4me1 is most abundant in the 3 ends of genes (van
Dijk et al., 2005). On the other hand, the distribution patterns of H3K4me2 are quite
different from yeast to mammals: In Saccharomyces cerevisiae, H3K4me2 appears to
spread out over (among?) the genes, and is associated with a transcriptionally active gene.
In mammals, however, H3K4me2 mainly colocalizes with H3K4me3 in discrete zones
about 5-20 nucleosomes in length (Bernstein et al., 2005; Schneider et al., 2004). Only a
small subset of H3K4me2 does not overlap with H3K4me3 and these regions do not
correlate to transcription start sites, but are highly dependent on the cell types, suggesting
that they may have a role in determining lineage specific chromatin states.
In the yeast, only one enzyme so far has been found to methylate H3K4. But in mammals,
there are at least ten H3K4 methyltransferases; those that are related to the yeast Set1 and
Drosophila Trx proteins (the MLL family) or those unrelated (ASH1, SET7/9, SMYD3
and Meisetz) (Beisel, 2002, Wang 2001, Hamamoto, 2004, Hayashi, 2005). The MLL
family contains MLL1 (Mixed Lineage Leukemia 1), MLL2, MLL3, MLL4, SET1A and
SET1B. The H3K4 methylase activities in mammals are not redundant, as the deletion of
either gene causes embryonic lethality. Like most histone modifying enzymes, the MLL
family exists in multi-protein complexes. There are three common subunits (WDR5,
RbPB5 and ASH2) which are required for H3K4 methylation by MLL1 in vitro and in
vivo (Dou et al., 2006).
In S.cerevisiae, the recruitment of Set1 to H3K4 is accomplished first by the
phosphorylated form of RNA polymerase II c-terminal domain (phosphorylated Pol II
CTD). This phosphorylation recruits Set1 to the H3K4 residue close to the promoter. The
RNApol II is released from the transcription initiation complex into the elongation
complex (Promoter escape). The second factor that recruits H3K4me3 is the PAF
elongation complex, which is involved in RNA metabolism and interacts with Set1 and
H2B monoubiquitylation. This H2BK123ub1 is the third component involved in H3K4
methylation. This mechanism in yeast is regarded as a general H3K4 methylation
regulation mechanism. The existence of a larger amount of H3K4 methyltransferase in
mammals makes it reasonable to envision several possibilities: H3K4 metyltransferases

31
may be recruited 1) directly or indirectly by sequence specific DNA binding proteins; 2)
by association with basal transcriptional factors; 3) through chromatin modification
readers; or 4) by RNA from the specific RNApol II binding regions. The available
evidence suggests the existence of a combination of gene specific and general
mechanisms. Ruthenburg et al. (2007) suggest that the initial recruitment is gene specific,
mediated by specific transcription factors and/or RNAs, and is sensitive to the signaling
cues within the cell, whereas further stabilization of the complex on chromatin and
stimulation of its catalytic activity occur through general mechanisms and stabilization of
the methyltransferases on chromatin by the association with acetylated and H3K4
methylated histones .
H3K36 methylation is catalyzed by the Set2 histone methyltransferase and is necessary
for the elongation phase of transcription. It also promotes deacetylation of transcribed
chromatin. Methylated H3K36 is found within the coding regions of actively transcribed
genes. Set2 can catalyze all three states of H3K36 and negatively influence gene
expression (for example the repression of cryptic promoters within coding regions of the
gene STE11). Different states of H3K36 (H3K36me2 and H3K36me3) have different
functions: the N-terminal of Set2 (containing SET domain) is sufficient for H3K36me2,
and is correlated to whether a gene is expressed or not, with histone deacetylation and
repression of cryptic promoters at STE11(Youdell et al., 2008)), while H3K36me3 tends
to correlate to highly expressed genes via the C-terminal domain of Set2 and requires
Spt6, the H3P38 (histone proline 38), and the CTD of RNAPII (Youdell et al., 2008)).
H3K79 methylation, unlike the other histone tail modifications, is located within the
globular domain, and is thus exposed on the nucleosome surface. The yeast Dot1
(disruptor of telomeric) and its homologues in other species are the only known H3K79
methyltransferases (Feng et al., 2002). Unlike the other methyltransferases, the Dot1
family members do not have a SET domain (Feng et al., 2002). Their catalytic domain
contains conserved sequence motifs characteristic of class I methyltransferases such as
DNMTs and arginine methyltransferases (PRMT1) (Cheng et al., 2005). In the yeast,
H3K79me is localized to euchromatic regions and it is associated with the coding region
of active genes.

32
H3K9 methylation is intimately connected with heterochromatin. It is, next to
deacetylated histones, the evolutionary most stable heterochromatic marker (Krauss,
2008). However, it has also been found to be enriched in transcribed active genes (Vakoc
et al., 2005). There are three locally and functionally distinct distributions of H3K9me: 1)
heterochromatin regions, 2) some epigenetically modified silenced promoter regions of
euchromatic genes (Tachibana et al., 2005), 3) within active transcription units where it
participates in the repression of illegitimate initiations of transcription (Vakoc et al.,
2006). As in other lysine methylation residues, H3K9 can be methylated in three levels:
mono-, di-, trimethylation, and these different levels of methylation status have distinct
locations: H3K9me1 (in mouse), H3K9me2 (in maize) or H3K9me3 (in Arabidopsis,
maize and Chlamydomonas) are mainly found in euchromatin; while H3K9me3 seems
common in the heterochromatin of animals, fungi, but not or rarely in plants. Recently
Folco and colleagues found that, together with the RNAi pathway, H3K9me2 is needed
to mark the neighboring heterochromatin region to recruit CENP-ACnp1
(Centromere-
specific protein) in the central domain in fission yeast Schizosaccharomyces pombe
(Folco et al., 2008). Drosophila seems to have a similar distribution pattern of H3K9me1,
2, 3, they are all localized to heterochromatic regions (Krauss, 2008).
H3K9 is methylated by SUV39 methyltransferases family. The first HKMT (histone
lysine methyltransferase) was HEK9-SUV39H1 (Rea et al., 2000). Proteins in this family
contain a N-terminal chromo domain and a histone methyltransferase (HMTase) catalytic
domain consisting of a SET domain flanked at the N-terminus by a cysteine-rich pre-SET
domain (required for specificity toward H3K9) and a C-terminus post-SET domain.
The formation of heterochromatin involves the cooperation of two proteins: SUV39H and
its binding protein HP1. HP1 has three forms in mammals: and. HP1 and HP1
interact with methylated H3K9 within the heterochromatin and the silenced regions of
euchromatin. However, HP1 interactions with methylated H3K9 are found on the actively
transcribed gene regions (Vakoc et al., 2005). The model for these two proteins
interaction is that methylated H3K9, which is carried out by SUV39H, works as a binding
platform for HP1 through its chromodomain. Once HP1 binds, it can spread into adjacent
nucleosomes by its association with SUV39H, which then further methylates histones.

33
On the other hand, HP1 has the possibility to self-associate with its chromoshadow
domain, which in turn facilitates the spread of heterochromatin. This is like the writer
(SUV39H) and the reader (HP1) relationship. How the writer can find the specific place
to write was revealed by the discovery of siRNAs (small interfering RNAs) (Volpe et al.,
2002). These RNAs come from the bidirectional transcription of centromeric repeats via
the Dicer enzyme, and these RNAs are packed into the RITS complex, which contains the
H3K9 methylated binding protein Chp1.
H3K27 methylation is crucial in gene silencing during development (Cao and Zhang,
2004), X-inactivation (Plath et al., 2003), stem-cell pluripotency (Boyer et al., 2006),
cancer (Sparmann and van Lohuizen, 2006) and inflammation (De Santa et al., 2007).
The control of H3K27 methylation is dynamic. It is catalyzed by EZH2 ( E(z) in
Drosophila) and can be methylated at three levels as well. EZH2 is a SET domain
(Su(var)3-9, Enhancer-of-zeste, Trithorax) containing enzyme, and is a core component
of PRC2 (Polycomb repressive complex 2). It has been shown that E(z) can methylate all
three levels of H3K27 in Drosophila, while in mammals, EZH2 can only methylate di-,
and tri- H3K27. In fact, the three levels of methylation have very different distributions:
in both Drosophila and mammals, H3K27me2 is virtually ubiquitous in euchromatic
regions (more than 50%); H3K27me3 is found essentially in PcG target sites (5-10%)
(Schwartz and Pirrotta, 2008). A heterochromatic enrichment of H3K27me1, H3K27me2
and H4K20me1 has been specifically found in angiosperms (plants). In Drosophila, E(z)
or (PRC2) is recruited to chromatin via the interaction of PRE (Polycomb response
elements) binding proteins. But in mammals, the targeting of the EZH2 complex may be
mediated by a variety of transcription factors, like YY1, GAGA factor and MYC.
H4K20 mono-methylation, H4K20me1, is associated with facultative heterochromatin
(Fang et al., 2002; Martens et al., 2005; Sims et al., 2006), and the inactive X
chromosome (Kohlmaier et al., 2004). H4K20 me3 is a marker of constitutive
heterochromatin (Mikkelsen et al., 2007). Mono- methylation of H4K20 is carried out by

34
PR-Set7 HKMT. SUV4-20H1 and SUV4-20H2 methylates catalyse the formation of
H4K20 me2 and H4K20me3. There is an exception, however; H4K20 has been linked to
DNA repair via the binding of the DNA damage checkpoint protein CrB2.
Histone Arginine methylation In mammals, histone arginine methylation is found on
the H3R2, H3R8, H3R17, H3R26 and H4R3 residues. It is catalyzed by the PRMTs
(protein arginine methyltrasferases) class of histone methyltransferases and contributes to
both active and repressive effects on gene expression (Wang et al., 2001). Histone
arginine residues can be methylated in mono-methyl, symmetrical di-methyl or
asymmetrical di-methyl states (Fig. 9). So far, eleven arginine methyltransferases
(PRMT1-11) have been found in mammals and six of them have the ability to methylate
histone arginine (PRMT1,4,5,7,8,9). Drosophila has 9 homologues (DART1-9), and they
all have the histone arginine methyltransferase ability. In yeast the PRMT1 (the
Drosophila homologue is DART1,2,3,6,8,9) is recruited to actively transcribed genes
during elongation, and its activity is important for heterogenous nuclear
ribonucleoprotein (hRNP)-mediated mRNA processing and export. In mammals, PRMT1
interacts with the p160 family of nuclear receptor coactivators and facilitates
transcription driven by the androgen receptor through the methylation of histone H4R3
(Wang et al., 2001). PRMT4 can methylate both the N-terminal (R2, 17, 26) and C-
terminal (R128, 129, 131, 134) arginine residues in histone H3 and enhance transcription
activation by methylating both histone arginine and coactivators (Bauer et al., 2002).
PRMT5 symmetrically methylates histones H3R8 and H4R3 along with other cellular
proteins. This enzyme has been found in multiple complexes where it mediates diverse
functions including RNA processing, transcriptional regulation, and muscle as well as
germ line differentiation (Dacwag et al., 2007). It has been co-purified with chromatin

35
Figure 9. Histone arginine methylations and deimination by PRMTs enzymes.
remodeling complexes, hSWI/SNF and NURD, and has therefore been associated with
transcriptional repression of cell cycle regulators (by methylating the promoters of
CYCLIN E, P14ARF
, and P16INK4a
) and tumor suppressor genes (ST7 and NM23 (Bachman
et al., 2003; Le Guezennec et al., 2006). PRMT5 has also been shown to interfere with
gene expression by methylating and promoting the dissociation of the transcription
elongation factor SPT5 (Kwon et al., 2003). On the other hand, PRMT5 was found as a
component of the androgen receptor-driven transcription, which is independent of its
catalytic activity (Chie et al., 2003). Accordingly, PRMT5 affects transcription in a
methylase-dependent as well as a methylase-independent fashion. PRMT7 can methylate
H2A and H4 (Lee et al., 2005). It mediates H4R3 methylation at the H19 and GTL2 loci

36
in male germ cells that contributes to the imprinting of H19 and GTL2 genes (Jelinic et
al., 2006). PRMT8, 9, 10, 11 are recently found and their function is still unknown.
Demethylation The discovery of the first histone demethylase, lysine-specific-
demethylase-1 (LSD1) (Metzger et al., 2005; Shi et al., 2004a), and the peptidyl arginine
deiminase 4 (PAD14/PAD4) (Wang et al., 2004), open a whole new view on the dynamic
regulation of chromatin.
LSD1 is an amine oxidase that catalyzes lysine demethylation in a FAD (flavine adenine
dinucleotide) dependent manner and is involved in repression. Interestingly, the
specificity of LSD1 can be changed if it binds to a partner, for example, the androgen
receptor (AP). When LSD1 and AP form a complex, it will demethylate H3K9 instead of
H3K4, acting as an activator instead of a repressor (Metzger et al., 2005). But, LSD1 can
only demethylate mono-and dimethyl marks on H3K4me1/2 and H3K9me1/2. It can not
demethylate tri-methylated lysines (example: H3K4me3 and/or H3K9) due to the absence
of a protonated nitrogen required for oxidation (Metzger et al., 2005; Shi et al., 2004b).
Demethylation of trimethyl marks on H3K4me3 in budding yeast is carried out by
YJR119Cp (or JARID1) (Liang et al., 2007; Seward et al., 2007). Both groups show that
H3K4 methylation and demethylation can be dynamically regulated, but Liang et al.
(2007) also showed that YJR119Cp contributed to the regulation of telomeric silencing as
well. The H3K4me3 demethylase is evolutionarily conserved. The Drosophila
YJR119Cp homolog lid (little imaginal discs) is a member of trithorax group of genes
(Klose et al., 2006). Lid can demethylate H3K4me2/3 in vitro, but not in vivo for
unknown reasons.
The demethylation of H3K36 is carried out by JHDM1 for H3K36me1/2 and
JHDM3A/JMJD2A for H3K36me3 (Klose et al., 2006). There are two things here which
are interesting to note: that the different enzymes are carried out the removal of the
various methyl group in the same lysine residue. JHDM1 is only capable of removing the
H3K36me1/2 modification states (Tsukada et al., 2006; Whetstine et al., 2006), while
H3K36me3 demethylation is catalyzed by JHDM3A/JMJD2A. The second interesting

37
issue is that the enzyme JHDM3A/JMJD2A can demethylate both the repressive mark
H3K9me3 and the active mark H3K36 me3. These dual functions are probably due to
interactions with different co-factors.
H3K79 methylation might be a more static mark since so far no demethylase for this
modification has been found. In contrast to the rapid deacetylation of H3, levels of H3-
K79me2 decrease gradually through the developmental time, which indicates that
replication-mediated dilution can largely account for the loss of di-methylated H3-K79;
while the more rapid removal of H3-K4 methylation requires a replication-independent
component that is specific for the H3-K4 modification (Katan-Khaykovich and Struhl,
2005).
The demethylation of H3K27me3 and H3K27me2 to H3K27me1 is catalyzed by the
JmjC-domain proteins UTX and JMJD3 (Agger et al., 2007; Lan et al., 2007; Lee et al.,
2007; Xiang et al., 2007). So far, no enzyme has been found that catalyzes demethylation
of H3K27me1, but a recent genomic study revealed that H3K27me1, me2 and me3 are
enriched downstream from transcription start sites of active genes (Barski et al., 2007). In
pluripotent cells, Hox genes are kept in a silent state by Polycomb group complex (PcG).
The genes are marked by H3K27me3, a modification which is catalyzed by EZH2 in the
PRC2 complex (PcG function will be discussed later). During differentiation, Hox genes
are derepressed colinearly, UTX was found to be recruited to the promoters of active Hox
gene while SUZ12 and EZH2 showed decreased occupancy. In differentiated cells, UTX
controls a steady-state level of non-methylated H3K27 at transcription start sites, but not
at coding and intergenic regions.
De Santa and co-workers (De Santa et al., 2007) showed that JMJD3 is rapidly induced
in macrophages in response to an inflammatory stimulus. Recruitment of JMJD3 affected
the expression of a subset of genes induced by inflammation, Bmp-2 being one of the
genes. Upon treatment, H3K27me3 levels decrease at the Bmp-2 locus, depending on
JMJD3.
Unlike lysine demethylation, there is no true histone arginine demethylase identified, so
far, which can remove methyl groups from either symmetric or asymmetric dimethylation

38
marks. Instead, at least so far, PADI4/PAD4 is the only enzyme which has been found to
antagonize histone arginine methylation by converting either arginines or monomethyl-
arginine to citrulline in a Ca2+
and DTT dependent reaction.
Ubiquitination / Deubiquitination
Ubiquitin is an 8.5kKD (76-amino acid) protein (Goldknopf et al., 1975). Its C-terminal
four amino acids are in a random coil, whereas the N-terminal 72 amino acids form a
compact globular structure.
Ubiquitination is the process where ubiquitin covalently conjugates to substrate proteins.
This involves three separate enzymatic activities (Fig. 10): Ubiquitin is activated by a
covalent attachment to E1 enzyme in an ATP-dependent reaction and is subsequently
transferred to an E2 (ubiquitin-conjugating) enzyme. Ubiquitin is transferred from E2 to
target substrate with an E3 (ubiquitin ligase) enzyme. Substrates can be mono- or
polyubiquitinated; whereas polyubiquitination (at lease four units of linked ubiquitin) of
target proteins are recognized by proteasomes (Pickart, 2001), monoubiquitinated
proteins are stable and are usually intracellular compartments (Hicke, 2001; Hicke and
Dunn, 2003)
Figure 10: The process of ubiquitination. Modified from (Pickart, 2001)

39
The histone H2A was the first protein identified to be ubiquitinated (Goldknopf et al.,
1975). The ubiquitination site for H2A is lysine 119 (Nickel and Davie, 1989) and
ubiquitinated H2A (ubH2A) has been detected in many eukaryotic organisms except in S.
cerevisiae. The majority of ubH2A is monoubiquitinated, however, polyubiquitinated
H2A has also been detected (Nickel and Davie, 1989). In addition to H2A, H2B can also
be ubiquitinated (West and Bonner, 1980). ubH2B is less abundant (1%-2%) than ubH2A
(5%-15%) and it can be either mono- or polyubiquitinated (Geng and Tansey, 2008).
Unlike ubH2A, however, the site for H2B ubiquitination (ubH2B) is different among
organisms: for example, the ubiquitin site in S. cerevisiae is Lys-123, in S. pombe is Lys-
119, in Arabidopsis is Lys-143 and in mammals it is in Lys-120. Thus far, only
monoubiquitinated H2B has been found.
Table 4. Enzymes involved in H2A and H2B ubiquitination/deubiquitination in different
organisms
ubH2B duH2B ubH2A duH2A
E2 E3 E2 E3
S. cerevisiae Rad6 Bre1 Ubp8
Ubp10
(Dot4)
Ubp3/4
(Doa4)
- - -
S. pombe Rhp6 Brl1
Brl2
Drosophila Dhr6 Bre1 Nonstop
USP7
dRing
(Sce)
L(3)73h
Mouse mHR6A
mHR6B
LASU1
Human hHR6A/
hHR 6B
UbcH6
Mdm2
RNF20 USP22
USP3
Ring1B
2A-HUB
Ubp-M
2A-DUB
USP21
USP3
Arabidopsis HUB1 SUP32
Modified from (Weake and Workman, 2008; Zhang, 2003).

40
In addition to H2A and H2B, ubiquitination of H3 and H1 has also been reported (Chen
et al., 1998; Pham and Sauer, 2000), but the sites for ubiquitination for these two proteins
are still not known. The histone variants H2A.Z and macro H2A1.2 are also reported to
be monoubiquitinated at Lys-120 or Lys-121 for H2A.Z and Lys-115 for macroH2A1.2
in mammals (Chu et al., 2006; Sarcinella et al., 2007). The monoubiquination of H2A.Z
is associated with human X-chromosome inactivation, but the biological function of
monoubiquitination of macroH2A1.2 is unclear.
Ubiquitin can be removed through hydrolysis of the peptide bond at Gly76 by
deubiquitinating enzymes (DUB). These consist of C-terminal hydrolases (UCH) and
ubiquitin-specific processing proteases (UBP) (D'Andrea and Pellman, 1998).
Enzymes involved in ubiquitination/deubiquitination in different species are listed in
Table 4, and the biological functions of the histone ubiquitination/deubiquitination
modifications are given in Table 5.
Table 5. The biological functions of histone ubiquitination/deubiquitination modifications.
Functions of ubiquitination and crosstalk with other histone modifications
Transcriptional activation Transcriptional
Silencing
DNA
damage
Cell
cycle
Initiation Early
enlongation
Late
enlongation
ubH2B
√
H3K4me2/3,
H3K79me2/3
√
√
duH2B √ √
ubH2A √
H3K4me2/3,
H3K4dem3
H1
H3K27me3
√ √
H2AXpho
ubH2AX
duH2A √ √

41
Histone ubiquitination and deubiquitination play important roles in regulating gene
transcription. They are based on the large amount of factors involved in addition to
ubiquitin-conjugating enzymes and ubiquitin ligases that regulate ubH2A/duH2A and
ubH2B/duH2B. The mechanism for transcription regulation by ubH2A/duH2A and
ubH2B/duH2B is based on a crosstalk with other histone modifications. For example, in
human HOX A7 gene silencing, H3K27me3 is a prerequisite for ubH2A and DNA
methylation, whereas ubH2A and DNA methylation are interdependent (Wu et al., 2008).
Interestingly, ubH2B can either activate transcription or silence gene activity. These dual
functions depend on gene location: for genes located in euchromatin regions, ubH2B
appears to result in gene activation, while for genes located in heterochromatic regions,
ubH2B has a negative effect. Regulation of gene transcription by ubH2B involves also a
crosstalk with other histone modifications, for instance, with H3K4me2/3 and
H3K79me2/3. However, unlike ubH2A, ubH2B is upstream of H3K4me2/3 and
H3K79me2/3 for initiation and the early stages of transcription elongation (Kao et al.,
2004; Wood et al., 2003).
Sumoylation
Even though it was discovered only a decade ago, sumoylation of proteins have got great
attention because of its vast ranges of target proteins. Similar to ubiquitination,
sumoylation is a procedure of a Small Ubiquitin-like Modifier (SUMO) protein
covalently attaching to the target protein. The SUMO proteins are about 12 kD (100-
amino acids). As their name indicates, they are 8-15% identical to ubiquitin and have
similar structure and mechanism of attachment to target proteins.
Since its identification in 1996 (Matunis et al., 1996), a large number of proteins (more
than 50 proteins) have been shown to be post-translationally modified by SUMO. For
example, the nuclear pore complex members RanGAP1 and RanBP2 (Joseph et al.,
2002); the chromatin modifiers EZH2 and SUZ12 (Riising et al., 2008); Nuclear bodies

42
members PML, HIPK2; transcription factors p53 and CTCF (MacPherson et al., 2009);
transcription co-factors HDAC1 , p300 and TIF1Mascle et al.; signal
transduction proteins IKB, FAK (Gill, 2004). So, sumoylation is involved in various
cellular processes, such as transcriptional regulation, nuclear transport, maintenance of
genome integrity, and signal transduction, targeting a protein to specific subcellular
locations (Johnson, 2004).
Here I will briefly describe some recently discoveries of the functions of sumoylation
modification in histone and histone association factors.
First, Smt3 (the Drosophila homologue of SUMO-3) is expressed throughout
development, more highly expresses during embryogenesis and in adult females (Huang
et al., 1998). It has been observed in chromocentre in polytene chromosome in
Drosophila (Lehembre et al., 2000). In humans, SUMO protein is also found restricted to
the constitutive heterochromatin regions in pachytene spermatocytes (Metzler-Guillemain
et al., 2008). Also, members involved in epigenetic regulators have been found can act
either as sumoylation E3 ligase (for example: Pc2 in mammals and Suppressor of
variegation 2-10 (Su(var)2-10) in Drosophila (Kagey et al., 2003; Mohr and Boswell,
1999)) or as target proteins (for example: SOP-2 (Zhang et al., 2004), SUZ12 and EZH2
(Riising et al., 2008)). Pc2 was first shown to act as a SUMO E3 ligase for repression of
CtBP in mammals (Kagey et al., 2003). SUMO modification of histone 4 (H4) has been
shown to relate to transcriptional repression (Shiio and Eisenman, 2003). SOP-2 (a PH
(Polyhomeotic) and SCM (Sec comb on midleg) homolog in C. elegans) is also required
to be modified by sumoylation to have the repression regulation of Hox gene. So,
sumoylation modifications of histone and histone associate factors required for their
proper repression functions.
Phosphorylation
Phosphorylation is the most well known in signaling transduction pathways. Histone 3
serine 10 (H3S10) was one of the first proteins found to be phosphorylated (Mahadevan
et al., 1991). The phosphorylation of H3S10 by JIL-1 kinase can counteract with

43
heterochromatization and gene silencing (Deng et al., 2008). Studies of linker histone H1
in mouse cells revealed that phosphorylation of this histone may affect its binding to
HP1 (Hale et al., 2006). H2A.X is another known histone which can be phosphorylated.
Phosphorylation of H2A-X is an early event in double-strand break repair.
Limited by the space in this thesis, some other aspects which should be considered in
epigenetic regulation of gene expression mechanisms have not been described. For
example, the non-coding RNA mediates gene expression mechanisms; the TrxG
regulation of gene activation mechanisms; the histone variation mechanisms……
The rapidly increasing data in the past few years strongly support the hypothesis that the
epigenetic regulations of gene expression play important roles in development.
1.5 Regulation of gene expression by PcG proteins
1.5.1 Introduction
What makes our skin cell different from our liver cells, even though they have the same
DNA sequence? The answer is simple: because they express different proteins. There are,
however, much more complicated questions: how to decide which protein should be
expressed or not; how cells maintain their determined state over many cell divisions (a
process termed “cellular memory”). We know at least some parts of the answers to these
questions. The Polycomb groups (PcG) and trithorax group (TrxG) are two gene families,
that have antagonistic functions: PcG proteins are believed to be required to maintain
repression Hox gene activity, while TrxG proteins maintain activated expression. PcG
proteins were originally identified in Drosophila as factors necessary to maintain cell-fate
decisions throughout embryogenesis by repressing Hox gene expression in a segment-
specific manner. Now PcG is known to constitute a large chromatin associated family of
proteins which is involved in many cellular memory processes: such as Hox gene
regulation (body layout), X-inactivation in female mammals and stem cell regulation.
Here, I shall first introduce PcG members and their functions, then I will describe PcG

44
mechanisms in genome programming. I shall also briefly discuss PcG genes in an
evolutionary perspective and finish this chapter by describing the inheritance of the
memory states.
1.5.2 Genetic and biochemical characterization of PcG proteins
The first PcG gene, Polycomb (Pc), was identified and analyzed by Pamela and Edward
B. Lewis (Lewis, 1978). Heterozygous Pc mutant male flies have extra sex combs on the
second and third legs. Homozygous mutants are embryonic lethals. Subsequent studies
showed that Hox genes are similarly misexpressed in several other mutants, so this set of
mutants that cause Hox gene derepression was named the Polycomb group (Duncan,
1982). More than 20 years ago, Jürgens (Jurgens, 1985) predicted that there are about 30-
40 PcG genes in the Drosophila genome based on an embryonic assay: embryos that are
double homozygous for mutations in two different PcG genes express in general strongly
enhanced homeotic transformation compared to single mutants. Until 2006, only 16 PcG
genes had been identified. Alonso and co-workers (Gaytan de Ayala Alonso et al., 2007)
tried to identify further PcG genes by a genome wide systematic genetic screen. When I
am writing this thesis in late 2008, 33 PcG genes have been identified in Drosophila.
Table 8 shows their biological functions and the mutant phenotypes.
Based on the genetic and the biochemistry studies of PcG regulation on Hox gene
repression, so far three main PcG complexes have been identified: PRC1 (Polycomb
repress complex 1), PRC2 (Polycomb repress complex 2) and PhoRC (Pho-repressive
complex). Except these three complexes, there are some genes which are loosely related
to major complexes or the ones with unknown function.

45
Table 6. Polycomb group members in Drosophila melanogaster.
Gene Name Location Protein
functional
domain
Biochemical activity Mutant
phenotype*
Polycomb
(Pc)
3L N-terminal
chromo-
domain
C-terminal Pc
box
Binds to H3K27me3
Interacts with dRing
I
Polycomb like
(Pcl)
2R PHD
Tudor
Enhances H3K27me3 I
Posterior sex
comb
(Psc)
2R Ring finger Enhances ubiquitination I
Polyhomeotic-
distal
phd
X SAM Might promote the spreading
of PcG
Oligomerization
III
Polyhomeotic-
proximal
php
X SAM Oligomerization III
Sex comb
extra
(Sce/dRing)
3R RING-type
zinc finger
H2AK119ub ubiquitination I
Pleiohomeotic
(pho)
4 Zinc finger Sequence-specific DNA
binding; can interact with
PRC1 and PRC2 components
II
Pleiohomeotic- 3L Zinc finger Sequence-specific DNA III

46
like
(phol)
binding; can interact with
PRC2 components
Scm-related
gene
containing
four mbt
domains
(dSfmbt)
2L MBT
SAM
Binds to H3K9me1/2 and
H4K20me1/2
I
Enhancer of
zeste
(E(z))
3L SET H3K27 methyltransferase
H1K26 methyltransferase
I
Extra sex
comb
(ESC)
2L WD-40 Contributes to H3K9me and
H3K27 methylation
I
Extra sex
comb like
(ESCL)
2L WD-40 Contributes to H3K9me and
H3K27 methylation
I
Suppressor of
zeste (Su(z)12)
3L VEFS box
Zinc finger
Contributes to H3K27
methylation
I
P55
(Nurf-55, or
Caf1))
3R Histone-
binding
Associates with HDAC,
H3K27me.
Sex comb on
midleg
(Scm)
3R MBT, SAM,
Zing finger
Self-association (SAM
domain), binds to mono-
methylated histone

47
super sex
comb
(sxc)
2R Unknown III
Additional sex
combs
(Asx)
2R
Su(z)2 2R
Cramped X II
Multi sex
comb
(mxc)
X II
Pipsqueak
(Psq)
2L BTB-POZ DNA binding
Dorsal Switch
Protein
(Dsp1)
X HMG Single strand DNA binding
Calypso 2R I
Siren 5 2R Unknown III
Siren 1/kto 3L Unknown III
Siren 9 /skd 3L Unknown III
Siren 2 3L Unknown III
Siren 7 3L Unknown III
Siren 8 3L Unknown III
Siren 3 3R Unknown III
Siren 4 3R Unknown III
Siren 6 3R Unknown III
Circe 3R Unknown III

48
*: Class I phenotype: Strong homeotic transformation; Homozygous lethal.
Class II phenotype: Mild homeotic transformation.
Class III phenotype: Weak homeotic transformation.
The PRC1 complex
A 2MD PRC1 (Polycomb repression complex 1) complex was purified from 0-12 hr
transgenic Drosophila embryos. It contains five core proteins: Phd, Ph
p, Psc, Pc and
dRing (Francis et al., 2001; Saurin et al., 2001; Shao et al., 1999) (Table 9).
Table 7. PcG complex paralogs in different organisms
Proteins D.
Melanogaster
M. musculus A.
thaliana
C. elegans
PRC1 complex (2Md)
Polycomb
polyhomeotic-dorsal
polyhomeotic-
proximal
Posterior Sex Combs
dRing/Sex combs
extra
Pc
phd
php
Psc
dRing/Sce
Cbx2/M33
Cbx4/MPc2
Cbx6
Cbx8/MPc3
Cbx7
Edr1/Mph1/Rae28
Edr2/Mph2
Bmi1
Mel-18
NsPc1
PcGF3
PcGF5
MBLR
Ring1/Ring1a
Rnf2/Ring1b
SOP-2

49
PRC2 complex (core complex)
Extra sex comb
Enhancer of zeste
Supressor of zeste 12
Nurf55
ESC
E(z)
Su(z)12
p55
Eed
Ezh1
Ezh2
SU(Z)12
RbAp48
RbAp46
FIE
CLF
MEA
SWN
FIS2
VRN2
EMF2
MSI1-5
MES-6
MES-2
Pho-RC complex
Pleiohomeotic
Pleiohomeotic-like
Scm-related gene
containing four MBT
domains
Pho
Phol
dSfmbt
YY1, YY2
YY1, YY2
SFMBT
Pcl-PRC2
Polycomb-like
Enhancer of zeste
Supressor of zeste 12
Extra sex comb
Nurff55
Heat-shock cognate
protein 70
Pcl
E(z)
Su(z)12
ESC
Nurff55
HSC70

50
Scm was also purified but at a lower amount. In addition to that, 25 other associated gene
products were also co-purified with these core components; for example: Zeste, six TAFs
(TATA box binding protein-associated factors) proteins: TAFII250, TAFII110, TAFII85,
TAFII62, TAFII42 and TAFII30. The large numbers of proteins associated with PRC1
implies that PRC1 may have multiple roles in gene regulation. An intriguing thing is the
co-purification of dTAFII promoter factors, which indicate a direct interaction between
PcG and promoter complexes. However, the main function of PRC1 is to maintain genes
silenced via its functional components. The N-terminus of Pc contains a chromo-domain
which preferentially binds to H3K27me3; while its C-terminus has a PC domain that can
bind to dRing. dRing has an E3 ubiquitin ligase activity, which specifically catalyzes
monoubiquitination of H2AK119. The H2AK119 monoubiquitination correlates well
with transcriptional silence and is presumed to inhibit RNA polymerase II transcriptional
elongation and DNA damage repair (Zhou et al., 2009; Zhou et al., 2008). In addition to
ubiquitination of H2AK119, a mammalian homolog of PRC1 was recently shown also to
act as E3 ubiquitin ligase for Geminin, an important factor for DNA replication (Ohtsubo
et al., 2008). The C-terminus of Ph contains a SAM (sterile alpha motif) domain. The
SAM domain is responsible for the polymerization of PcG complexes (Kim et al., 2002)
and so is thought to promote the spreading of PcG complex along the chromatin. Psc is a
ring finger-containing protein, but the function in Drosophila is still unclear except that it
can enhance the ubiquitination of H2AK119.
The PRC2 complex
The Drosophila PRC2 complexes are multiple entities that change dynamically during
development. Three distinct PRC2 complexes are found in Drosophila embryos (Table
10):
The first biochemically isolated PRC2 complex from 0-24 hr transgenic embryos has the
size of 600 kD that contains three PcG proteins Enhancer of zeste [E(z)], Suppressor of
zeste 12 (Su(z)12), Extra sex combs (ESC) and/or Extra sex combs-like (ESCL), and
Nurf55 (Muller et al., 2002). This 600 kD complex is the most common one. The main
function of this complex is the histone 3 lysine 27 methyltransferase activity carried out
by E(z) via its SET domain. However, E(z) alone cannot catalyze the H3K27 methylation,

51
it needs Nurf55 to bind to nucleosomes, while Su(z)12 and ESC are needed for the
enzyme activity. esc and escl are two homolog genes in Drosophila with a similar
function. ESC protein level is high in 0-18 hr embryos, and decreases from 18-24 hour
embryos to 2nd
instar larvae. There is no detectable ESC in 3rd
instar larvae, pupae and
adult males, but it re-appears in adult females. In a fashion different from ESC, ESCL is
expressed ubiquitously throughout development (the expression level is, however,
weaker in pupae). Genetic and biochemically studies show that ESCL can substitute for
ESC and functions as a backup capacity during development (Kurzhals et al., 2008; Ohno
et al., 2008). Su(z)12 contains a VEFS domain which is conserved from plant through
mammals. The biological function of Su(z)12 remains unclear. It is needed for all levels
of methylation in H3K27 (H3K27me1,2,3) (Chen et al., 2008; Nekrasov et al., 2007).
Table 8. The PRC2 complexes.
PRC2 Embryo
stages
Components
E(z) ESC Su(z)12 Nurf55 Rpd3 Pcl HSC70
600 kD 0-24 hour √ √ √ √
1 MDa 0-12 hour √ √ √ √ √ √
Pcl-PRC2 16-18 hour √ √ √ √ √ √
The 2nd
PRC2 complex isolated, except for the same core component as the 600 kD
complex, contains Rpd3 and Pcl. The PHD domain in Pcl can directly bind to the histone
de-acetylase Rpd3. These two complexes seem functionally distinct from each other: the
600 kD complex is present through the whole embryogenesis, while the 1 MD complex is
there until the mid embryo stage (Furuyama et al., 2003; Tie et al., 2001; Tie et al., 2003).
The 3rd
PRC2 complex, Pcl-PRC2, was purified from 16-18 hr embryo and contains Pcl
and Heat-shock cognate protein 70 (HSC70) except for the 600 kD core components
(Nekrasov et al., 2007). Pcl-PRC2 is required to generate high levels of H3-K27
trimethylation at Polycomb target genes but is not needed for the genome-wide H3-K27

52
mono- and dimethylation. Pcl mutant embryos show strong misexpression of Ubx and
Abd-B, whereas en, another target gene is only misexpressed in a few rare cells
(Nekrasov et al., 2007). However, another Pcl complex (about 1500 kD) which contains
no E(z) was purified from third stage larvae. This larval Pcl complex (PCLC) functions
upstream of E(z) and also bind to the Ubx PRE (Savla et al., 2008).
Pho-RC complex
The Pho-repressive complex (PhoRC) contains Pho (pleiohomeotic), Phol
(pleiohomeotic-like) and dSfmbt (Scm-related gene containing four mbt domains). It is
the only PcG complex that contains sequence-specific DNA binding proteins: Pho and
Phol (Klymenko et al., 2006). Pho and Phol function redundantly to maintain Hox gene
silencing (Brown et al., 2003). pho single mutant clones show slight Hox gene
misexpression, and phol single mutant clones show no Hox gene misexpression. However,
the double mutant clones of pho and phol show severe misexpression. The human
homolog of Pho and Phol is Yin Yang 1 (YY1). YY1 is a multifunctional nuclear protein
that can act as a transcriptional repressor or activator (Seto et al., 1991; Shi et al., 1991):
for example, it can repress the activity of the adeno-associated viral P5 promoter in the
absence of the oncoprotein E1A, but can also activate the promoter in the presence of
E1A. It plays roles in cell proliferation, differentiation, survival and nervous system.
Interestingly, YY1 regulated genes in the nervous system show contrasting behaviour:
some genes are repressed by YY1: (i.e. Dynamin 1, Glast), while some are activated (i.e.
Engrailed 2, Snail) (He and Casaccia-Bonnefil, 2008). Klymenko and co-workers
purified the PhoRC complex of Drosophila (Nekrasov et al., 2007). They found that Pho
actually exits in two complexes, one with dSfmbt (the Pho-RC complex which is
associated with PcG silencing). The other complex, co-purified with INO80 (a
Drosophila nucleosome-remodeling complex) is independent of Pho-RC function. While
in mammals, Cai and co-workers studied the function of Pho-INO80 in HEK293 cells
(Cai et al., 2007). They show that INO80 acts as an co-activator with YY1 to activate
transcription. Incidentally, a study on even-skipped (eve) regulation in Drosophila shows

53
that Pho maintains both active and repressed transcriptional states through Eve PRE
(Fujioka et al., 2008).
Figure 11. The biochemical activity of the PcG complex. Modified from (Schwartz and Pirrotta, 2008)
The other component in PhoRC complex is dSfmbt, which contains four copies of
“malignant bran tumor” (MBT) and a “sterile a motif” (SAM) domain. As in other PcG
lacking somatic clones, Ubx was de-repressed in dSfmbt mutant clones 72 hr after
induction in disc. dSfmbt1 homozygous embryos from heterozygours mothers showed
mild misexpression of Hox genes. However, dSfmbt1 homozygous embryos from mutant
germ line cells fail to develop. The MBT domains contribute mainly to the binding of
dSfmbt to mono- and di-methylated H3K9 and H4K20, which is in contrast to the well
documented Pc preference in binding of tri-methylated H3K27.
The three main complexes have different contributions to epigenetic marks (Table 9) and
are supposed to localize in the regulatory region of target gene (Fig. 11)

54
One may speculate that: dSfmbt “scans” PRE-flanking chromatin for nucleosomes that
are only mono- or di-methylated at H3K9 and/or H4K20 and docks onto such
nucleosomes through its MBT domain. Then the PRC2 bound to PREs interacts with
Pho/Phol, methylates H3K27me3, which then facilitates PRC1 to bind to the target gene
(Klymenko et al., 2006).
Table 9. PcG members contribute to the epigenetic marks.
PcG H3K27me H3K9me H4K20me H2A119ub
Me1 Me2 Me
3
Me1 Me2 Me3 Me1 Me2 Me3
E(z) √ √ √
Su(z)12 √ √ √
ESC √ √ √
ESCL √ √
Pcl √
dSfmbt √ √ √ √
Pc √
In addition to the themes of PcG complex, there are some variation members that have
looser relationships with the main complexes or belong to the complexes that have not
yet been purified. For example: the Additional sex combs (Asx) gene of Drosophila is
required for activation and repression of homeotic loci. It interacts specifically with
Polycomb and super sex combs.
In Drosophila, E(z) is associate with Polycomb-like (PCL) protein since PCL shares
polytene chromosome staining pattern with E(z). Indeed, a 1-MDa ESC-E(z) complex
containing PCL and the histone deacetylase RPD3 during early embryogenesis. The
different composition of the PRC2 complex may be involved in the silencing of
different
targets. PCL contains two plant homeodomain (PHD) domains, a motif present in many
proteins involved in chromatin function, such as CHD3, Mi2, TRX, ASH1, and ASH2. It
has recently been shown that the PHD domain link histone methylation to active
chromatin remodeling. Multiple PCL homologs have been identified in mammals. For

55
example, there are three genes in the mouse genome that encode PCL homologs, namely
mPcl1, mPcl2, and mPcl3 (Cao et al., 2008).
Nervous System Polycomb 1 (NSPc1), another Psc homolog in mammals, plays
important roles in promoting H2A ubiquitination and cooperates with DNA methylation
in Hox gene silencing (Wu et al., 2008). The Polycomb Group gene PCGF5 was
identified as one of the genes most strongly downregulated upon Notch inhibition. Notch
may be responsible for maintaining expression of a transcriptional repressor that
suppresses p16 and p19ARF. Whether PCGF5 acts as an analogous to Bmi-1 in this
cellular context and mediates p16/p19ARF repression is still under investigation.
MBLR (Mel18 and Bmi1-like RING finger protein) interacts with Ring1B and acts as a
transcriptional repressor in transiently transfected cells. Serine 32 of MBLR is
specifically phosphorylated during mitosis by CDK7 (Akasaka et al., 2002).
1.5.3 Mechanisms of PcG regulation of gene silencing
1.5.3.1 The Polycomb Response Element (PRE)
To understand the mechanism of how PcG complexes regulate gene expression, we have
to first understand how PcG complexes are recruited to their target genes. In Drosophila,
a functional analysis of the regulatory regions of several PcG target genes has led to the
identification of some specific elements which are necessary and sufficient for PcG-
mediated repression. All three PcG complexes bind to such regions that are in general
several hundred base pair long. These sequences are called Polycomb response elements
(PREs). So far, several studies have focused on PREs of a few genes in Drosophila: for
example: the Abd-B PRE Fab7, the Ubx PRE, bxd and the engrailed PRE (Fig. 12).

56
Figure 12. Some PcG target gene PREs . Modified from (Ringrose and Paro, 2007).
Even though Pho/Phol are the only DNA binding proteins among the PcG complex
components, they are not the only PRE binding proteins for PcG repression. Genetic and
biochemical studies show that in Drosophila, PREs contain a cluster of GAGAG motifs,
which are bound by the GAGA factor (GAF) and Pipsqueak (PSQ) (Horard et al., 2000;
Huang et al., 2002). Further more, SP1/KLF (promoter-specific transcription factor /
Krüppel-like factors) were found to bind to engrailed PRE (Brown et al., 2005); DSP1
(dorsal switch protein 1) is also involved in some PRE binding; the Zeste protein has long
been known to be involved in both activation and silencing at PREs (Dejardin and Cavalli,
2004); and lastly, Grh (Grainyhead) and PSQ bind to similar sequences. These proteins
are involved in homeotic gene regulation (Blastyak et al., 2006; Uv et al., 1994). Even
though these proteins often bind at or near PREs, the number and distribution of the
binding sites is quite different among different targets. None of these proteins so far has
been shown to be present at all PREs. Since PREs contain so many protein binding motif
sequences, it is not strange that, in general, PREs are not conserved sequences, but the
motif sequences are conserved (Fig 12). Interestingly, some PRE binding proteins are

57
involved both in the regulation of activation and repression of gene activity: for example:
Zeste, Pho/Phol, Dsp1, GAF/PSQ. The other two proteins, Grh and Sp1/KLF, are not
expressed uniformly but show a tissue specific expression pattern (Ringrose and Paro,
2007). The recruitment of PcG to PRE is the first step for PcG silencing to work. Already
a glimpse of what factors makes up PREs shows how complicated the system is. PREs
have, however, been only found in Drosophila, mammalian PREs (if there are any) have
not been identified.
1.5.3.2 PcG regulation of gene silencing at specific loci.
A stepwise model was proposed for PcG complex regulation of gene silencing (Fig, 13).
The first step is the binding to DNA sequence (PRE) through DNA binding protein
Pho(Phol); then PCL in PCL-PRC2 complex interacts with Pho/Phol, leading to
recruitment of PRC2 to target genes and at the same time, E(z) establishes the silence
mark by tri-methylation of H3K27me1/2/3, which, in turn, facilitates the binding of N-
terminal Chromodomain of Pc and further recruits the PRC1 complex. The Ring-type
zinc finger domain in dRing in the PRC1 complex contributes to H2AK119
monoubiquitination, thus further enhancing the silent state of the chromatin once the
silent state has been established.
PRC2 and PhoRC may be abolished from a silenced site while PRC1 may still be present
(Ohno et al., 2008). The staying of PRC1 at target gene loci while the H3K27me3 mark
has gone means that PRC1 may have a binding facility other than H3K27me3. A possible
explanation for PcG complex spreading along the chromosome and formation of a higher
order chromatin structure is the SAM domain in Ph. Ph-SAM can form visible filaments
and can enable PcG complexes to interact with weaker sites nearby (Kim et al., 2002).

58
Figure 13. Gene silencing induced by PcG binding to specific loci.
In this context, a question has to be considered: PRC2 complex has three variations that
differ in size and components: the 600kD PRC2 complex, the 1 MDa PRC2 complex and
the Pcl-PRC2 complex (Table 10). When considering the different representation of
H3K27me2 and H3K27me3 in total nucleosomes (>50% and 5-10% respectively),
Schwartz and Pirrotta (2008) proposed that the common core 600kD complex “probably
carries out the much more general function of dimethylating H3K27” (Schwartz and
Pirrotta, 2008).
Accordingly, PcG proteins inhibit transcription by changing the structure of the
chromatin fiber: PhoRC and/or other PRE binding protein find the target, the PRC2

59
complex establishes (or writes) the silencing mark H3K27me3 and the PRC1 maintains
(or reads) the silence state and decorates the chromatin with H2AK119ub (Fig 14).
Figure 14. PcG confers negative cis-regulation of target genes at the chromatin level.
1.5.3.3 PcG gene regulation at chromatin level
Genome wide analysis of PcG binding shows that PcG proteins binding to chromosomes
is not always limited to PRE domains, but many of the proteins have a wider binding
region, for example Pc, Ph and/or H3K27me3 (Negre et al., 2006; Schwartz et al., 2006;
Tolhuis et al., 2006). The regions where Pc, Ph and/or H3K27me3 bind are called PcG
domains (PcD). These domains may range from many kilobases to hundreds of kilobases
up- and downstream of PREs. PcG proteins can not only regulate silencing of a gene in a
cis configuration but can also mediate silencing of genes that are far away from PREs in
the genome (a phenomenon called long-range gene silencing) (Fig 15). The regulation of
BX-C (bithorax complex) by PcG proteins in Drosophila is one example. BX-C is a
cluster of Hox genes (Ubx, Abd-A and Abd-B) which contain several PREs. Mcp is one of
the PRE elements that controls Abdominal –B (Abd-B) expression and it is located 60 kb
far away from its target (Busturia et al., 1997; Lewis, 1978).
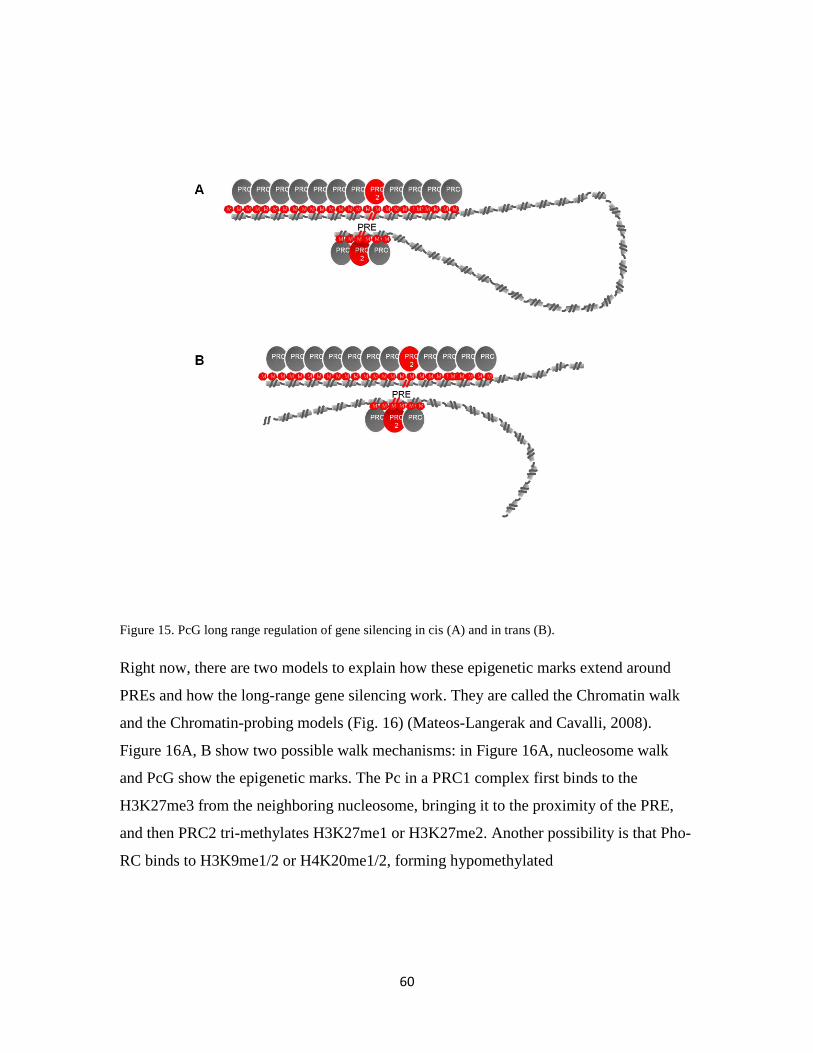
60
Figure 15. PcG long range regulation of gene silencing in cis (A) and in trans (B).
Right now, there are two models to explain how these epigenetic marks extend around
PREs and how the long-range gene silencing work. They are called the Chromatin walk
and the Chromatin-probing models (Fig. 16) (Mateos-Langerak and Cavalli, 2008).
Figure 16A, B show two possible walk mechanisms: in Figure 16A, nucleosome walk
and PcG show the epigenetic marks. The Pc in a PRC1 complex first binds to the
H3K27me3 from the neighboring nucleosome, bringing it to the proximity of the PRE,
and then PRC2 tri-methylates H3K27me1 or H3K27me2. Another possibility is that Pho-
RC binds to H3K9me1/2 or H4K20me1/2, forming hypomethylated
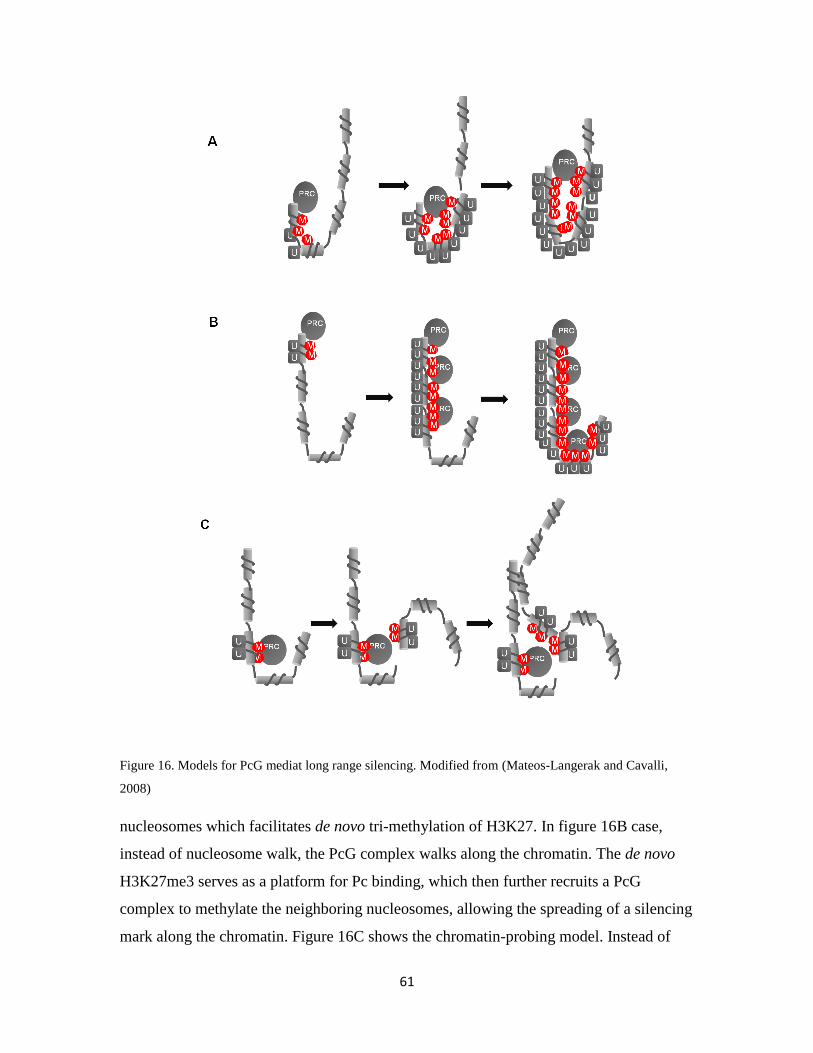
61
Figure 16. Models for PcG mediat long range silencing. Modified from (Mateos-Langerak and Cavalli,
2008)
nucleosomes which facilitates de novo tri-methylation of H3K27. In figure 16B case,
instead of nucleosome walk, the PcG complex walks along the chromatin. The de novo
H3K27me3 serves as a platform for Pc binding, which then further recruits a PcG
complex to methylate the neighboring nucleosomes, allowing the spreading of a silencing
mark along the chromatin. Figure 16C shows the chromatin-probing model. Instead of

62
spreading along the chromatin, the PRE-bound PcG can methylate the nucleosomes that
happen to interact with the enzymatic components of PcG.
1.5.3.4 PcG genes and the three-dimensional (3D) organization of the nucleus
The spatial organizations of chromatin (nuclear architecture) are important for the proper
function of the cell (Cremer et al., 2004). In Drosophila, in the repressive state, PcG
forms “PcG bodies” which are form by the PRES through long-range interactions. While
in the active state, the active gene “move out” from the PRE-promoter interaction regions
and become activated (Lanzuolo et al., 2007). In mammals, which PRE has not been
defined, similar chromatin loops are also found. These chromatin loops are PcG-occupied
and enriched of H3K27me3 (Tiwari et al., 2008b).
1.5.4 Evolution of PcG
PcG and TrxG proteins are often said to be evolutionary conserved. TrxG components are,
indeed, found in fungi, plants and animals. Table 3 shows two features: Drosophila (an
invertebrate) has in general single copies of the PcG proteins (except ph, esc and psc), but
vertebrates have multiple paralogs of most PcG members. The other feature is the lack of
PRC1 homologs in plants and C.elegans.
The components of the PRC2 complex are found in plants and animals, even though they
are missing in S. cerevisiae and S. pombe, but another fungus, Neurospora crassa,
possesses the H3K27me3 mark. Accordingly, PRC2 genes might have an ancient
function in transcriptional repression.
The evolution of the PRC1 complex must be more complex: on one hand, there are no
genes encoding core PRC1 proteins in fungi, plants (except for the Sce homolog) or C
elegans (except for the Scm homolog), which may indicate that PRC1 genes have been
added in the course of animal evolution. On the other hand, they might have been lost
from plants and fungi.
During evolution some or all genes get duplicated and subsequently their coding
sequences diverge by spontaneous mutations. For the PRC1 complex, Drosophila has
one copy of Pc gene, but vertebrates have five Pc homologs with different domain

63
structure and biochemical properties (Whitcomb et al., 2007). For the other members in
PRC1 there is also one gene copy of each in Drosophila, for example: psc and dRing,
while vertebrates have 6 and 2 homologs respectively. The key domains of these
homologs are highly conserved but regions outside the key domains diverge from their
Drosophila counterparts and from their paralogs. For example, the highly conserved
region of Pc is the chromodomain [chromobox (Cbx)] and Pc box with 75-88% amino
acid identity between a vertebrate and Drosophila. The length and other motifs within
each paralog differ, which may contribute to differential functions. In mammals, Cbx7
and Cbx8 can bypass replicative senescence due to repression of the INK4a-ARF locus.
The binding of Cbx8 to INK4a-ARF locus depends on Bmi1 (Psc homolog), whereas
Cbx7 bypasses senescence through a Bmi1-independent fashion (Bernard et al., 2005;
Dietrich et al., 2007). The five Cbx proteins in mouse (Cbx2, 4, 6, 7 and 8) bind
differentially to H3K27me3 and H3K9me3: Cbx7 and Cbx2 bind to H3K9me3 and
H3K27me3, Cbx4 prefers H3K9me3, Cbx6 and Cbx8 do not bind to either modification.
With the exception of Cbx4 all localize to the Xi during female mouse embryonic stem
cell differentiation and Cbx7 association with chromatin is developmentally regulated
(Bernstein et al., 2006).
Psc homologs in vertebrates are another example of non-redundant paralogs. There are 6
homologs in a mammalian genome: Bmi1, Mel-18, NsPc1, PcGF3, PcGF5 and MBLR.
Even though the encoded proteins have 63% amino acid identity, Mel-18 and Bmi1
mutant mice display similar but unique phenotypes. In addition, Mel-18 and Bmi1 have
opposite effects on cell-cycle regulation: Bmi1 acts as an oncogene while Mel-18
behaves like a tumor suppressor (Guo et al., 2007a; Guo et al., 2007b). Both Bmi1 and
Mel-18 can form a stable PRC1 complex, but only Bmi1 has been shown to positively
regulate the H2AK119ub formation by Ring1B. A structural analysis shows that Bmi1
contains a potential H2A-binding surface, which is absent in Mel-18. Perhaps the
different components in PRC1 contribute to its catalytic activity in vivo.
Mouse Ring1A and Ring1B are two homologs of dRing in mouse. They share 100%
identity in the Ring domain with Drosophila dRing protein, 48% and 55% similarity to
the full length of Drosophila dRing, respectively. Both Ring1A (-/-) and Ring1A (+/-)

64
mice show anterior transformations and other abnormalities of the axial skeleton (del Mar
Lorente et al., 2000). Ring1B heterozygous mice show no skeletal phenotype but
abnormal gastrulation (Voncken et al., 2003). At the molecular level, both Ring1A and
Ring1B are needed for H2A ubiquitination during X-inactivation, showing an
overlapping function of these two proteins. Ring1B is needed for H2A ubiquitination in
embryonic stem cells. A genetic interaction between YY1 and Ring1A genes has been
shown (Lorente et al., 2006).
Vertebrates have undergone a tetraploidization of the genome in the distant past and have,
therefore, retained multiple copies of many genes. Furthermore, some of their PcG
members have undergone multiple duplications, which have resulted in either related or
antagonistic functions. Some PcG genes have probably been lost in the course of
evolution, for example the plants may have lost PRC1 and the fungi and C. elegans have
lost Su(z)12.
1.5.5 PcG and cell cycle
PcG dependent silencing is not only important for correct expression of homeotic genes
during early embryogenesis, but also plays an important role in cell proliferation. PcG
complexes regulate the self-renewal of various types of embryonic and adult stem cells of
vertebrates. Many cancer types have abnormal PcG complex expression levels.
The cell cycle has been divided into four phases: mitotic (M) phase, synthesis (S) phase
and two gap phases (G1 and G2), between the M phase to S phase and S phase to M
phase, respectively (Fig. 17).

65
Figure 17. PcG regulation of cell cycle.
The progression through the cell cycle is highly regulated: at the transition from G1 phase
to S phase and from G2 phase to M phase, there are two checkpoints: the G1checkpoint
and the G2 one. In the middle of mitosis, there is one more checkpoint: the metaphase
checkpoint. These checkpoints assess cell-intrinsic signals and are governed by two key
protein families: cyclin-dependent protein kinases (Cdks) and cyclins. A total of 13 Cdks
and many cyclins have thus far been identified in mammals. Cdk1 is supposed to govern
the G2 checkpoint, while Cdk2 controls the G1 checkpoint. In addition, cell-extrinsic
signals also regulate the processes from early G1 phase to late G1 phase. The transition is
called restriction (R) point. The early G1 phase is mitogen-dependent; the late G1 phase
is mitogen-independent. In the absence of mitogen stimulation, cells can enter a quiescent
G0 phase which is characterized by small cell size and low metabolic activity. Transition
from an early G1 phase to late G1 phase is regulated by Cdk4/6 and cyclin D through
inactivation of Retinoblastoma tumor suppressor protein (RB) and activation of Cyclin E.
Cdk 2 and cyclin E regulate the mitogen-independent late G1 phase.

66
Two cyclin-dependent kinase inhibitors (CDKI) regulate the transition through the G1
phase: Ink4a family and CIP/KIP family. Ink4a family members (p15INK4b
and p16INK4a
)
directly inhibit Cdk4/6-cyclin D complexes; and members of CIP/KIP family (p21CIP
.
p27KIP1
, p57KIP2
) are direct inhibitors of the late G1 Cdk2-cyclin E complexes.
PcG members have been reported to regulate cell cycle in different ways. First, the
indirect affects of cell cycle regulation via regular cell cycle regulators, for example, Hox
genes. Several Hox genes are involved in the control of cell proliferation (Duboule, 1995;
Morgan et al., 2007; Salser and Kenyon, 1996). Second, PcG members regulate
chromatin condensation during mitosis. For example, Drosophila topoisomerase II and
Barren proteins are required for proper chromatin condensation, and they are targeted by
PcG complexes (Lupo et al., 2001). The centrosomal and chromosomal factor (ccf),
another chromosome condensation regulator in Drosophila, colocalizes with Psc binding
sites on polytene chromosomes (Kodjabachian et al., 1998). In addition, php, Pc and Psc
mutant embryos show anaphase bridges during chromatid segregation and E(z) mutants
show chromosome misalignment at metaphase (O'Dor et al., 2006). Finally, PcG
complexes seem to directly regulate the cell cycle control genes cyclin A (Martinez et al.,
2006), cyclin B (Oktaba et al., 2008) and cyclin G (Salvaing et al., 2008).
1.6 Epigenetic inheritance
I have mainly discussed the mechanisms of organization of chromatin by PcG for
establishing and maintaining cellular identity. The next key question is how the
epigenetic marks are transmitted to the daughter cells, and another, how the cells inherit
the epigenetic marks to remember their specificity. This process is referred to as mitotic
epigenetic inheritance. Many such phenomena have been studied in Drosophila. For
example, in Drosophila embryos, undifferentiated cells responsible for the formation of
adult structure can go through many cells divisions and still retain the ability to form the
appropriate structure upon hormonal induction.
In many organisms (but not in Drosophila that has very weak DNA methylation or not at
all), both DNA and histones can be modified. Both DNA and histone modifications are

67
thought to be involved in these epigenetic phenomena. I have discussed the mechanism
for DNA methylation. Here, I will first focus on describing models on chromatin
assembly pathways, the “piggy-back” and “bi-piggy-back” models (Martin and Zhang,
2007) (Fig 18), and then, from a PcG perspective, briefly describe the localization pattern
of some PcG members during cell cycle (Fig 19).
The semiconservative model of histone replication states (Fig 18A) that during DNA
replication, the nucleosome is split so that the old H3/H4 dimer go together with the old
DNA strand and the new H3/H4 dimers are assembled into the daughter DNA strand
(Tagami et al., 2004). The old histone modifications are recognized by histone
modification binding proteins and these binding proteins further recruit histone
modifying proteins, which then modify the new dimers. Positive support for this model
comes from the study on genomic regions associated with gene activity (Ahmad and
Henikoff, 2002). However, studies from the bulk chromatin indicate that most H3/H4
tetramers remain intact during DNA replication and the mono-nucleosomes isolated from
the bulk chromatin are homogenous, containing either H3.1 or H3.3. A conservative
model (Fig 18B) was proposed to explain this phenomenon: The old tetramers are intact
and found on both daughter strand DNA, the nucleosomes assembled with newly
synthesized tetramers. In semi-conservation model (Fig 18C), the histone binding
modification proteins bound at the old nucleosomes, modifing the new adjacent
nucleosomes. “Piggy-back” model (Fig 18D): The methylated copying DNA recruits
CpG-methyl binding proteins, which then recruit histone modifying proteins to methylate
histones. An example in support of this model is the recruitment of the SUV39H1 by
methyl-CpG binding protein MeCP2 to the target genes, leading to H3K9 methylation
and gene silencing (Fuks et al., 2003).

68

69
Figure 18. Chromatin replication models. (A) Semi-conservative model. (B) Conservative model. (C)
DNA semi-conservative model. (D) “Piggy-back” model. (E) “Bi-piggy-back ”model. Modified from
(Martin and Zhang, 2007).
“Bi-piggy-back” model (Fig 18E): As usual, there is always an exception for any model,
in this case for the “piggy-back” model: EZH2 interacts with DNA methyltransferases
and directs DNA methylation to target genes, and this function is necessary for gene
silencing in mammalian cells (Vire et al., 2006). In this case, DNA methylation depends
on the methylation of H3K27 by EZH2 and SUZ12 interacts directly with DNMT and
then leads to gene silencing. Vire et al. (2006) did not, however, study the dependence of
histone methylation on DNA methylation. So here I refer to this phenomenon as “bi-
piggy-back” mechanism.
In Drosophila embryos, Pc, Ph and Psc dissociate from the chromatin before mitotic
metaphase. Pc reassociates to chromatin after chromatin has decondensed at the next
interphase, while Ph and Psc rebind to chromatin already at telophase (Fig. 19)
(Buchenau et al., 1998; Dietzel et al., 1999).

70
Figure 19. The localization of PcG members during cell cycle.
1.7 Epigenetics and disease
The first evidence of a role for epigenetics in human disease came from the study of
genomic imprinting (Reik, 1989). Now more and more evidence have shown that
epigenetic mechanisms should also be considered in human diseases. Human
epidemiological studies showed that prenatal and early postnatal environmental factors
influence the adult risk of developing various diseases, such as cancer, chronic disease,
diabetes, rheumatoid arthritis (Barker, 2004; Gluckman et al., 2007; Sanchez-Pernaute et
al., 2008; Yajnik, 2004), and even schizophrenia (Malaspina et al., 2008; Quenstedt and
Parshall, 1998; Reynolds, 2000; van Os and Selten, 1998). Here I will briefly describe the
epigenetic alterations in cancers and the potential epigenetic targets for cancer therapy.
Cancer is the consequence of a combination of genetic disorder and aberrant epigenetic
states. Genetic defects usually refer to DNA sequence deletion, insertion or point
mutation. They are easy to visualize and the changes are usually permanently. Abnormal
epigenetic alterations include changes of DNA methylation, improper histone
modifications and the disruption of higher-level chromatin organization in nucleus
(Feinberg, 2007; Jirtle and Skinner, 2007). Unlike genetic changes, epigenetic changes
are reversible. Therefore epigenetic modifiers are attractive targets for epigenetic therapy.

71
2. AIM
The aims of this thesis were to investigate:
the binding pattern of Su(z)12 protein to polytene chromosomes and compare
to E(z) and H3K27me3.
whether Su(z)12 binding to the white gene could explain the dominant effect
of Su(z)12 mutants on z1 repression of white expression
the effect of Su(z)12 on H3K27me3 methylation
the expression pattern of Su(z)12 during Drosophila development
the mechanism by which Su(z)12 regulates target gene silencing
the role of Su(z)12 in cell cycle regulation.

72
3. RESULTS AND DISCUSSION
Suppressor of zeste 12 (Su(z)12) is needed for histone 3 lysine 27 tri-methylation
(H3K27me3) in vivo (paper I)
This study investigates the in vivo mechanism of Su(z)12 in gene silencing. Su(z)12
mutant flies exhibit strong homeotic transformation caused by mis-expression of
homeotic genes in embryos and larvae (Birve et al., 2001). Su(z)12 has also been
suggested to be involved in position effect variegation (PEV) and in regulation of the
white gene expression by interaction with zeste. To test the molecular mechanisms for
these two phenomena, we stained salivary gland polytene chromosomes from wildtype,
z1w
+, z
1w
+UZ and z
1w
is larvae with Su(z)12 and heterochromatin protein 1 (HP1)
antibodies. The eye pigments of the two latter phenotypes becomes completely
suppressed in heterozygous Su(z)12 mutants. Surprisingly, we did not find a Su(z)12
binding site in the white locus from any of these larvae. A weak binding of HP1 at the
white locus in polytene chromosome is found in the z1w
+UZ larvae, indicating that the
inserted p-element can recruit heterochromatic silencing proteins. We also stained
polytene chromosomes of the same fly strains with a H3K27me3 antibody, but no
binding site at the white locus was found. These data did not support the hypothesis that
PcG and HP1 are involved in z1 dependent repression of the white gene in Drosophila.
Su(z)12 co-localizes well with E(z) on polytene chromosomes, but not always with
H3K27me3. ESC protein has been shown to completely co-localize with E(z) (Tie et al.,
1998), thus our data further support that the PRC2 complex subunits are present at the
same sites on polytene chromosomes. We find that Su(z)12 binds mainly to interband
regions and these sites do not always overlap with H3K27me3 nor with HP1. Our results
confirm the chromatin immune-precipitation Chip analysis results reported by Schwartz
and the co-worker (Schwartz et al., 2006).
In humans, SUZ12 is found to regulate H3K9me and HP1distribution (de la Cruz et al.,
2007). Related to our observation that Su(z)12 mutants exhibit PEV, a characteristic
which is unique among PcG members, we speculate that Su(z)12 may have some
interaction with HP1. However, we did not observe co-staining of Su(z)12 and HP1 on

73
wildtype polytene chromosomes. So, from this point of view, mammals and Drosophila
seem to have some differences in controlling gene silencing.
This study was first to show that Su(z)12 is needed for H3K27me3 in vivo. Intriguingly,
over-expression of Su(z)12 increases H3K27me3 activity in wing discs. Since Su(z)12
does not have a methyltransferase activity of its own, we hypothesize that either Su(z)12
protein is rate limiting or that increased Su(z)12 levels induce positive feedback,
activating other PRC2 components, which lead to high H3K27me3 levels.
When analyzing the polytene chromosome staining results, we also noticed that Su(z)12
exhibits a weak staining in the nucleolus region. The nucleolus is composed of densely
packed fibrils and granules. It is the site of ribosomal RNA synthesis, which forms the
major part of the ribosomes. Recent studies show that sumoylation is important for
nucleolus organization (Heun, 2007), and that SUZ12 and EZH2 are sumoylated in vivo
and in vitro (Riising et al., 2008). We could thus speculate that Su(z)12 may have other
functions in addition to silencing.
Su(z)12 regulates the engrailed (en) gene both directly and indirectly (paper II)
The engrailed gene, is a target of PcG silencing, and has been well studied. In all those
studies the main focus have been on the regulatory role of the engrailed PRE (Devido et
al., 2008) and the PRC1 complex, i.e. Pc and Ph (Busturia and Morata, 1988; Randsholt
et al., 2000). As expected, engrailed is up-regulated in the anterior regions in wing discs
(where En is not expressed) in PRC1 somatic mutant clones due to the resulting de-
repression, but loss of PRC1 function has no effect in the posterior region (where En
usually is expressed).
In this study, we discovered a novel phenomenon. Instead of being up-regulated in the
anterior region in Su(z)12 knock-out clones in wing discs, we found down-regulation of
En in the posterior region. This could be interpreted as Su(z)12 acting as an engrailed
activator. However, we found that this “activator” role was caused by an indirect
regulatory loop. Loss of Su(z)12 leads to up-regulation of Ubx and Abd-A, which then

74
suppresses en expression. We also tested engrailed expression pattern in E(z) knock out
clones in wing discs. The result was the same as with Su(z)12. In addition to engrailed,
we also detected that other PcG target genes, like wingless and cut, behaved similarly in
Su(z)12 knock out clones.
Recently, Oktaba and co-workers found a similar phenomenon (Oktaba et al., 2008).
They tested the expression of en and other PcG target genes in different PcG gene knock-
out clones, like in dsfmbt, ph, pho, scm, Su(z)12 PcG genes, and some double knock out
clones (i.e Scm; BXC). They found that en and Dll were down-regulated in dSfmbt and
Scm knock-out clones, respectively, but in the double knock-out of Scm and BXC clones,
strong up-regulation of Dll was observed. This is in complete agreement with our results.
Our results emphasize that each gene has to be regarded as a unique entity. Their
regulation can differ between developmental stages, tissues and between compartments
within tissues.
The role of Su(z)12 in regulating cell proliferation and apoptosis (paper III)
The human homolog of Su(z)12 (SUZ12/JJAZ1/KIAA0160) was first reported to be up-
regulated in endometrial stromal tumors. The two zinc finger protein encoding genes
JAZF1 and JJAZ1 are located at the sites of the 7p15 and 17q21. Frequent fusion events
between the JAZF1 and JJAZ1 genes was found in translocation (t(7;17)(p15;q21)) in
such tumors. Since then, many reports have shown that PcG are involved in various
tumors (Ballestar and Esteller, 2008). In this study, we investigate the localization pattern
of Su(z)12 during the cell cycle and its role in cell proliferation.
Unlike the uniform localization pattern of Su(z)12 in S-phase cells, Su(z)12 has very
weak staining in metaphase chromosomes in mitotic embryonic cells. Similar patterns
have been reported for Ph, Psc and Pc in Drosophila (Buchenau et al., 1998; Dietzel et al.,
1999; Paro and Zink, 1993). This indicates that PcG proteins are dynamically regulated
during mitotic cell division in Drosophila.

75
In meiotic sperm cells the localization patterns are even more complicated: Staining
Drosophila testes for Su(z)12 protein showed that there are three types of Su(z)12
localization patterns. In spermatocytes, Su(z)12 protein is localized in nuclear foci. At the
rounded spermatid stage, Su(z)12 has a ubiquitous distribution, which is quite similar to
the distribution pattern for SUZ12 at the ovulated oocyte stage in mouse (Solter et al.,
2004). At the spermatid stage, Su(z)12 has totally left the needle shape sperm nuclei. One
could speculate that the complicated dynamic localization of Su(z)12 during Drosophila
spermatogenesis is essential to prevent the spermatids from undergoing differentiation. In
other words, Su(z)12 may be essential for keeping the germ cell lineage to retain its
pluripotency.
In this study, we also show that Su(z)12 are involved in regulation of cell proliferation
and apoptosis. The most interesting discovery here is the different phenotypes obtained
from over-expression of Su(z)12. When driven by ey-gal4 or vg-Gal4 drivers, no
overgrowth was observed, while when Su(z)12 was driven by Hem-Gal4 , the hemocyte
cell numbers increased significantly. We think that the reason for the different results
between wing discs and hemocyte stem cells are probably due to that the different tissues
have different regulatory networks.
Su(z)12 encodes different isoforms which are functionally different (paper IV)
We found that Su(z)12 encodes two isoforms and that they are developmentally and
dynamically regulated. Only Su(z)12-B and the human homolog can rescue Su(z)12
mutants. The specific neuron expression pattern indicates that Su(z)12 may involve in
regulation of neuron development.

76
4. CONCLUSIONS
Su(z)12 has 90 binding sites in polytene chromosomes. These binding sites are
overlaping well with E(z), but do not completely overlap with H3K27me3.
Su(z)12 is essential for H3K27me3 in vivo.
Su(z)12 regulate the engrailed gene both directly and indirectly via Ubx.
Su(z)12 is dynamically regulated during cell division.
Lack of Su(z)12 may decrease cell proliferation and increase apoptosis.
There are two different isoforms of Su(z)12 which are developmentally and/or
tissue specifically regulated.

77
ACKNOWLEDGEMENTS
For a long time before writing this thesis I have been thinking about what to write in this
part. Many people have encouraged and helped me on my way. I do not think for a
second that I ever could have finished this work without their support. I deeply appreciate
it. It is, therefore, my self-evident obligation to try to express thanks first to some of you:
First and foremost, my deepest gratitude to my supervisor Åsa Rasmuson-Lestander,
not only for your extensive knowledge of genetics, but also for your ability to encourage
a truth-seeking approach to science, and for criticizing my manuscripts and this thesis.
You deserve my deepest respect.
Jan Larsson, my co-supervisor, you are the kind of a person that people first ask for help
when they have a problem (or at least I have always done so). You are reliable, helpful
and wise. I am deeply indebted to you.
Anssi Saura for all your support! Without your support and encouragement I would not
have done this job. I am awed by your wide knowledge and your kindness. I respect you
as a great person.
David Robert for suggestions to this project.
Jurg Muller (EMBL, Heidelberg, Germany) for contributing with useful discussions.
I should also like to thank the folks from the Genetics of the old days:
Stefan Andersson Escher, my first genetics teacher in Sweden and a great party
organizer. Kerstin Kristiansson for showing me how to use the pipette; Per Stenberg
for the computer and sequence support; Anna Birve, for your friendship and support.
Bettan, for showing how to fish in Umeå; Astrid for our good friendship and the annual
Christmas cards! Karin E, for your kindness, Gunilla and Agneta, for always being
patient with my “late order” fly food; and other “Genetic colleagues”: Jesper, Magnus,
Markus, Tor-Mikael, Anna W, Maggan and Marianne, for creating a friendly
environment in my first few years of studying in Sweden.

78
I should also like to thank all the colleagues in the Department of Molecular Biology, in
particular the PhD students in Åsa s group: Linn and Anna L, you have a warm heart,
you are kind, honest and considerate, I have been very happy and lucky to work together
with you; Erik, for nice JC articles and discussion; John, impressed by your peaceful and
efficient work. Thank you for showing me the beautiful tradition of a Swedish wedding
party; and Anna S, I thank you all for making a friendly working atmosphere. I wish you
all good luck.
My room mates: David, for your understanding and help, people should be happy for
having a friend which can trusted, and even better if he or she is helpful. Regina, you
have been like a sister, you always want to help people, sometimes you are simply too
nice. I am so glad to have friends like you.
Jonas and Lisa for the discussion of cell cycle, Linus and Chaz, Sara, Andreas and
Sanna, for your friendship.
Victoria Shingler and Martin Gullberg, for your kindness, Roland Rosqvist, for help
in microscopy, Roland, Matt, Bernt Erik, Sun Nyunt, Iwa, Gunnar, Sven, Mikael,
Christina G, Eleonora for your support.
Edmund, for the friendly discussions and showing me the RNA stuff.
Berit, Ethel, Maria W, and in particular Helena Östbye for always help me with many
administration things and friendly smile.
I should like to thank the present or past “Fly people”:
Ruth Palmer, I am impressed by your intelligent and positive attitude; Camilla Sjögren,
for your kindness; Dan Hultmark, for giving us the driver strains; and all the colleagues
in the fly floor: Christina, Caroline, Therese, Margret, Fredrik, Jana, Gaura, Anna-
Mia, Lina, Malin, Calle, Ines, Michael, Anna-Karin, Karin B, Peter, Ingrid, Martin,
Jens-Ola, Mazen, Torbjörn, Shenhua, Lixiao. I wish you all the best.
For those people outside the work, I should also like to thank:

79
My dear friends Eleonora and Caroline, I am glad to have friends like you who can
share the happiness and also sadness. Take care!
My Chinese friends: Aihong Li, Good luck with everything. Tong Li, for always the tea
support from Beijing. Lu Li, for our discussion from the western blot to the opinion of
marriage. Kui Liu and Yan Shen, Yunshen Zhao and Yafan Mei, Yu Chen and Ou
Tang, Yu Jun and Liming Gu, Xiaoru Wang, Yan Xin and Xiaohong Lu, Zhiyun
Zhen and Chunglian Shi, Wei Jian and Weigang Gu, Jingxia Liu and Jiguo Yu,
Junfa Liu, Bo Xu and Shaojun Xiong, Ming Yuan and Jian Lu, Bo Huang,
Changchun Chen, Tianyan Song, Nu Huang (黄女), for all your friendship!
My lovely neighbors: Susan och Fredrik, Terese och John, Bigitta, Christina och
Håkan, Susan och Åke, Lotta och Jonas. Vilken tur har vi som bor granne med er.
Speciellt tack till Siv och Sigg, man säger att barn är en produkt av Gud, men att gamla
människor är en produkt av sig själva. Vi känner oss alltid välkomna hos er. Det värmer
mitt hjärta när jag ser hur ni behandlar varandra och andra. Tusen tack!
Finally, I would like to thank my family: Mama, for supporting and encouraging me all
my life, I love you, mom! 妈妈,我爱你! Brother, Weiming, for love! I wish you the
best!
I thank my husband, Wenchao, for all these years of support in general, the computer
support in particular. I am not sure if I shall ever graduate from your computer course.
Bobby and Oliver, my lovely sons, you are the sun in my life. You give me the strength
to meet each day as it comes; I love you, my sons! Thank you for taking care of your
mom!

80
REFERENCES
Agger, K., Cloos, P.A., Christensen, J., Pasini, D., Rose, S., Rappsilber, J., Issaeva, I., Canaani, E., Salcini, A.E., and Helin, K. (2007). UTX and JMJD3 are histone H3K27 demethylases involved in HOX gene regulation and development. Nature 449, 731-734. Ahmad, K., and Henikoff, S. (2002). The histone variant H3.3 marks active chromatin by replication-independent nucleosome assembly. Molecular cell 9, 1191-1200. Akasaka, T., Takahashi, N., Suzuki, M., Koseki, H., Bodmer, R., and Koga, H. (2002). MBLR, a new RING finger protein resembling mammalian Polycomb gene products, is regulated by cell cycle-dependent phosphorylation. Genes Cells 7, 835-850. Akhtar, A., Zink, D., and Becker, P.B. (2000). Chromodomains are protein-RNA interaction modules. Nature 407, 405-409. Bachman, K.E., Park, B.H., Rhee, I., Rajagopalan, H., Herman, J.G., Baylin, S.B., Kinzler, K.W., and Vogelstein, B. (2003). Histone modifications and silencing prior to DNA methylation of a tumor suppressor gene. Cancer Cell 3, 89-95. Ballestar, E., and Esteller, M. (2008). Epigenetic gene regulation in cancer. Advances in genetics 61, 247-267. Barker, D.J. (2004). The developmental origins of chronic adult disease. Acta Paediatr Suppl 93, 26-33. Barski, A., Cuddapah, S., Cui, K., Roh, T.Y., Schones, D.E., Wang, Z., Wei, G., Chepelev, I., and Zhao, K. (2007). High-resolution profiling of histone methylations in the human genome. Cell 129, 823-837. Bauer, U.M., Daujat, S., Nielsen, S.J., Nightingale, K., and Kouzarides, T. (2002). Methylation at arginine 17 of histone H3 is linked to gene activation. EMBO reports 3, 39-44. Bell, A.C., and Felsenfeld, G. (2000). Methylation of a CTCF-dependent boundary controls imprinted expression of the Igf2 gene. Nature 405, 482-485. Bernard, D., Martinez-Leal, J.F., Rizzo, S., Martinez, D., Hudson, D., Visakorpi, T., Peters, G., Carnero, A., Beach, D., and Gil, J. (2005). CBX7 controls the growth of normal and tumor-derived prostate cells by repressing the Ink4a/Arf locus. Oncogene 24, 5543-5551. Bernstein, B.E., Kamal, M., Lindblad-Toh, K., Bekiranov, S., Bailey, D.K., Huebert, D.J., McMahon, S., Karlsson, E.K., Kulbokas, E.J., 3rd, Gingeras, T.R., et al. (2005). Genomic maps and comparative analysis of histone modifications in human and mouse. Cell 120, 169-181. Bernstein, E., Duncan, E.M., Masui, O., Gil, J., Heard, E., and Allis, C.D. (2006). Mouse polycomb proteins bind differentially to methylated histone H3 and RNA and are enriched in facultative heterochromatin. Mol Cell Biol 26, 2560-2569. Birve, A., Sengupta, A.K., Beuchle, D., Larsson, J., Kennison, J.A., Rasmuson-Lestander, A., and Muller, J. (2001). Su(z)12, a novel Drosophila Polycomb group gene that is conserved in vertebrates and plants. Development (Cambridge, England) 128, 3371-3379. Blastyak, A., Mishra, R.K., Karch, F., and Gyurkovics, H. (2006). Efficient and specific targeting of Polycomb group proteins requires cooperative interaction between Grainyhead and Pleiohomeotic. Mol Cell Biol 26, 1434-1444. Boyer, L.A., Plath, K., Zeitlinger, J., Brambrink, T., Medeiros, L.A., Lee, T.I., Levine, S.S., Wernig, M., Tajonar, A., Ray, M.K., et al. (2006). Polycomb complexes repress developmental regulators in murine embryonic stem cells. Nature 441, 349-353. Brown, J.L., Fritsch, C., Mueller, J., and Kassis, J.A. (2003). The Drosophila pho-like gene encodes a YY1-related DNA binding protein that is redundant with pleiohomeotic in homeotic gene silencing. Development (Cambridge, England) 130, 285-294.

81
Brown, J.L., Grau, D.J., DeVido, S.K., and Kassis, J.A. (2005). An Sp1/KLF binding site is important for the activity of a Polycomb group response element from the Drosophila engrailed gene. Nucleic acids research 33, 5181-5189. Brownell, J.E., Zhou, J., Ranalli, T., Kobayashi, R., Edmondson, D.G., Roth, S.Y., and Allis, C.D. (1996). Tetrahymena histone acetyltransferase A: a homolog to yeast Gcn5p linking histone acetylation to gene activation. Cell 84, 843-851. Buchenau, P., Hodgson, J., Strutt, H., and Arndt-Jovin, D.J. (1998). The distribution of polycomb-group proteins during cell division and development in Drosophila embryos: impact on models for silencing. The Journal of cell biology 141, 469-481. Busturia, A., and Morata, G. (1988). Ectopic expression of homeotic genes caused by the elimination of the Polycomb gene in Drosophila imaginal epidermis. Development (Cambridge, England) 104, 713-720. Busturia, A., Wightman, C.D., and Sakonju, S. (1997). A silencer is required for maintenance of transcriptional repression throughout Drosophila development. Development (Cambridge, England) 124, 4343-4350. Cai, Y., Jin, J., Yao, T., Gottschalk, A.J., Swanson, S.K., Wu, S., Shi, Y., Washburn, M.P., Florens, L., Conaway, R.C., et al. (2007). YY1 functions with INO80 to activate transcription. Nature structural & molecular biology 14, 872-874. Cao, R., Wang, H., He, J., Erdjument-Bromage, H., Tempst, P., and Zhang, Y. (2008). Role of hPHF1 in H3K27 methylation and Hox gene silencing. Molecular and cellular biology 28, 1862-1872. Cao, R., and Zhang, Y. (2004). The functions of E(Z)/EZH2-mediated methylation of lysine 27 in histone H3. Current opinion in genetics & development 14, 155-164. Chen, H.Y., Sun, J.M., Zhang, Y., Davie, J.R., and Meistrich, M.L. (1998). Ubiquitination of histone H3 in elongating spermatids of rat testes. The Journal of biological chemistry 273, 13165-13169. Chen, S., Birve, A., and Rasmuson-Lestander, A. (2008). In vivo analysis of Drosophila SU(Z)12 function. Mol Genet Genomics 279, 159-170. Cheng, X., Collins, R.E., and Zhang, X. (2005). Structural and sequence motifs of protein (histone) methylation enzymes. Annual review of biophysics and biomolecular structure 34, 267-294. Chie, L., Cook, J.R., Chung, D., Hoffmann, R., Yang, Z., Kim, Y., Pestka, S., and Pincus, M.R. (2003). A protein methyl transferase, PRMT5, selectively blocks oncogenic ras-p21 mitogenic signal transduction. Ann Clin Lab Sci 33, 200-207. Chu, F., Nusinow, D.A., Chalkley, R.J., Plath, K., Panning, B., and Burlingame, A.L. (2006). Mapping post-translational modifications of the histone variant MacroH2A1 using tandem mass spectrometry. Mol Cell Proteomics 5, 194-203. Cremer, T., Kupper, K., Dietzel, S., and Fakan, S. (2004). Higher order chromatin architecture in the cell nucleus: on the way from structure to function. Biology of the cell / under the auspices of the European Cell Biology Organization 96, 555-567. D'Andrea, A., and Pellman, D. (1998). Deubiquitinating enzymes: a new class of biological regulators. Crit Rev Biochem Mol Biol 33, 337-352. Dacwag, C.S., Ohkawa, Y., Pal, S., Sif, S., and Imbalzano, A.N. (2007). The protein arginine methyltransferase Prmt5 is required for myogenesis because it facilitates ATP-dependent chromatin remodeling. Molecular and cellular biology 27, 384-394. de la Cruz, C.C., Kirmizis, A., Simon, M.D., Isono, K., Koseki, H., and Panning, B. (2007). The polycomb group protein SUZ12 regulates histone H3 lysine 9 methylation and HP1 alpha distribution. Chromosome Res 15, 299-314.

82
De Santa, F., Totaro, M.G., Prosperini, E., Notarbartolo, S., Testa, G., and Natoli, G. (2007). The histone H3 lysine-27 demethylase Jmjd3 links inflammation to inhibition of polycomb-mediated gene silencing. Cell 130, 1083-1094. Dejardin, J., and Cavalli, G. (2004). Chromatin inheritance upon Zeste-mediated Brahma recruitment at a minimal cellular memory module. The EMBO journal 23, 857-868. Dekker, J. (2006). The three 'C' s of chromosome conformation capture: controls, controls, controls. Nature methods 3, 17-21. del Mar Lorente, M., Marcos-Gutierrez, C., Perez, C., Schoorlemmer, J., Ramirez, A., Magin, T., and Vidal, M. (2000). Loss- and gain-of-function mutations show a polycomb group function for Ring1A in mice. Development (Cambridge, England) 127, 5093-5100. Deng, H., Bao, X., Cai, W., Blacketer, M.J., Belmont, A.S., Girton, J., Johansen, J., and Johansen, K.M. (2008). Ectopic histone H3S10 phosphorylation causes chromatin structure remodeling in Drosophila. Development. Devido, S.K., Kwon, D., Brown, J.L., and Kassis, J.A. (2008). The role of Polycomb-group response elements in regulation of engrailed transcription in Drosophila. Development (Cambridge, England). Dietrich, N., Bracken, A.P., Trinh, E., Schjerling, C.K., Koseki, H., Rappsilber, J., Helin, K., and Hansen, K.H. (2007). Bypass of senescence by the polycomb group protein CBX8 through direct binding to the INK4A-ARF locus. The EMBO journal 26, 1637-1648. Dietzel, S., Niemann, H., Bruckner, B., Maurange, C., and Paro, R. (1999). The nuclear distribution of Polycomb during Drosophila melanogaster development shown with a GFP fusion protein. Chromosoma 108, 83-94. Dou, Y., Milne, T.A., Ruthenburg, A.J., Lee, S., Lee, J.W., Verdine, G.L., Allis, C.D., and Roeder, R.G. (2006). Regulation of MLL1 H3K4 methyltransferase activity by its core components. Nature structural & molecular biology 13, 713-719. Duboule, D. (1995). Vertebrate Hox genes and proliferation: an alternative pathway to homeosis? Current opinion in genetics & development 5, 525-528. Duncan, I.M. (1982). Polycomblike: a gene that appears to be required for the normal expression of the bithorax and antennapedia gene complexes of Drosophila melanogaster. Genetics 102, 49-70. Fang, J., Feng, Q., Ketel, C.S., Wang, H., Cao, R., Xia, L., Erdjument-Bromage, H., Tempst, P., Simon, J.A., and Zhang, Y. (2002). Purification and functional characterization of SET8, a nucleosomal histone H4-lysine 20-specific methyltransferase. Curr Biol 12, 1086-1099. Feinberg, A.P. (2007). Phenotypic plasticity and the epigenetics of human disease. Nature 447, 433-440. Feng, Q., Wang, H., Ng, H.H., Erdjument-Bromage, H., Tempst, P., Struhl, K., and Zhang, Y. (2002). Methylation of H3-lysine 79 is mediated by a new family of HMTases without a SET domain. Curr Biol 12, 1052-1058. Ferguson-Smith, A.C., Moore, T., Detmar, J., Lewis, A., Hemberger, M., Jammes, H., Kelsey, G., Roberts, C.T., Jones, H., and Constancia, M. (2006). Epigenetics and imprinting of the trophoblast -- a workshop report. Placenta 27 Suppl A, S122-126. Folco, H.D., Pidoux, A.L., Urano, T., and Allshire, R.C. (2008). Heterochromatin and RNAi are required to establish CENP-A chromatin at centromeres. Science (New York, NY 319, 94-97. Francis, N.J., Saurin, A.J., Shao, Z., and Kingston, R.E. (2001). Reconstitution of a functional core polycomb repressive complex. Molecular cell 8, 545-556. Frank, S.R., Parisi, T., Taubert, S., Fernandez, P., Fuchs, M., Chan, H.M., Livingston, D.M., and Amati, B. (2003). MYC recruits the TIP60 histone acetyltransferase complex to chromatin. EMBO reports 4, 575-580.

83
Fujioka, M., Yusibova, G.L., Zhou, J., and Jaynes, J.B. (2008). The DNA-binding Polycomb-group protein Pleiohomeotic maintains both active and repressed transcriptional states through a single site. Development (Cambridge, England) 135, 4131-4139. Fuks, F., Hurd, P.J., Wolf, D., Nan, X., Bird, A.P., and Kouzarides, T. (2003). The methyl-CpG-binding protein MeCP2 links DNA methylation to histone methylation. The Journal of biological chemistry 278, 4035-4040. Furuyama, T., Tie, F., and Harte, P.J. (2003). Polycomb group proteins ESC and E(Z) are present in multiple distinct complexes that undergo dynamic changes during development. Genesis 35, 114-124. Gaughan, L., Logan, I.R., Cook, S., Neal, D.E., and Robson, C.N. (2002). Tip60 and histone deacetylase 1 regulate androgen receptor activity through changes to the acetylation status of the receptor. The Journal of biological chemistry 277, 25904-25913. Gaytan de Ayala Alonso, A., Gutierrez, L., Fritsch, C., Papp, B., Beuchle, D., and Muller, J. (2007). A genetic screen identifies novel polycomb group genes in Drosophila. Genetics 176, 2099-2108. Geng, F., and Tansey, W.P. (2008). Polyubiquitylation of histone H2B. Molecular biology of the cell 19, 3616-3624. Gerstein, M.B., Bruce, C., Rozowsky, J.S., Zheng, D., Du, J., Korbel, J.O., Emanuelsson, O., Zhang, Z.D., Weissman, S., and Snyder, M. (2007). What is a gene, post-ENCODE? History and updated definition. Genome research 17, 669-681. Gill, G. (2004). SUMO and ubiquitin in the nucleus: different functions, similar mechanisms? Genes & development 18, 2046-2059. Girton, J.R., and Johansen, K.M. (2008). Chromatin structure and the regulation of gene expression: the lessons of PEV in Drosophila. Advances in genetics 61, 1-43. Gluckman, P.D., Hanson, M.A., and Beedle, A.S. (2007). Early life events and their consequences for later disease: a life history and evolutionary perspective. Am J Hum Biol 19, 1-19. Goldknopf, I.L., Taylor, C.W., Baum, R.M., Yeoman, L.C., Olson, M.O., Prestayko, A.W., and Busch, H. (1975). Isolation and characterization of protein A24, a "histone-like" non-histone chromosomal protein. The Journal of biological chemistry 250, 7182-7187. Goransson, M., Forsman, K., and Uhlin, B.E. (1989). Regulatory genes in the thermoregulation of Escherichia coli pili gene transcription. Genes & development 3, 123-130. Grienenberger, A., Miotto, B., Sagnier, T., Cavalli, G., Schramke, V., Geli, V., Mariol, M.C., Berenger, H., Graba, Y., and Pradel, J. (2002). The MYST domain acetyltransferase Chameau functions in epigenetic mechanisms of transcriptional repression. Curr Biol 12, 762-766. Guo, W.J., Datta, S., Band, V., and Dimri, G.P. (2007a). Mel-18, a polycomb group protein, regulates cell proliferation and senescence via transcriptional repression of Bmi-1 and c-Myc oncoproteins. Molecular biology of the cell 18, 536-546. Guo, W.J., Zeng, M.S., Yadav, A., Song, L.B., Guo, B.H., Band, V., and Dimri, G.P. (2007b). Mel-18 acts as a tumor suppressor by repressing Bmi-1 expression and down-regulating Akt activity in breast cancer cells. Cancer research 67, 5083-5089. Hale, T.K., Contreras, A., Morrison, A.J., and Herrera, R.E. (2006). Phosphorylation of the linker histone H1 by CDK regulates its binding to HP1alpha. Molecular cell 22, 693-699. He, Y., and Casaccia-Bonnefil, P. (2008). The Yin and Yang of YY1 in the nervous system. Journal of neurochemistry 106, 1493-1502. Hernday, A., Krabbe, M., Braaten, B., and Low, D. (2002). Self-perpetuating epigenetic pili switches in bacteria. Proceedings of the National Academy of Sciences of the United States of America 99 Suppl 4, 16470-16476. Heun, P. (2007). SUMOrganization of the nucleus. Current opinion in cell biology 19, 350-355. Hicke, L. (2001). Protein regulation by monoubiquitin. Nat Rev Mol Cell Biol 2, 195-201.

84
Hicke, L., and Dunn, R. (2003). Regulation of membrane protein transport by ubiquitin and ubiquitin-binding proteins. Annu Rev Cell Dev Biol 19, 141-172. Holliday, R. (2006). Epigenetics: a historical overview. Epigenetics 1, 76-80. Honda, B.M., Candido, P.M., and Dixon, G.H. (1975a). Histone methylation. Its occurrence in different cell types and relation to histone H4 metabolism in developing trout testis. The Journal of biological chemistry 250, 8686-8689. Honda, B.M., Dixon, G.H., and Candido, E.P. (1975b). Sites of in vivo histone methylation in developing trout testis. The Journal of biological chemistry 250, 8681-8685. Horard, B., Tatout, C., Poux, S., and Pirrotta, V. (2000). Structure of a polycomb response element and in vitro binding of polycomb group complexes containing GAGA factor. Mol Cell Biol 20, 3187-3197. Huang, D.H., Chang, Y.L., Yang, C.C., Pan, I.C., and King, B. (2002). pipsqueak encodes a factor essential for sequence-specific targeting of a polycomb group protein complex. Mol Cell Biol 22, 6261-6271. Huang, H.W., Tsoi, S.C., Sun, Y.H., and Li, S.S. (1998). Identification and characterization of the SMT3 cDNA and gene encoding ubiquitin-like protein from Drosophila melanogaster. Biochem Mol Biol Int 46, 775-785. Jablonka, E., and Lamb, M.J. (2002). The changing concept of epigenetics. Annals of the New York Academy of Sciences 981, 82-96. Jelinic, P., Stehle, J.C., and Shaw, P. (2006). The testis-specific factor CTCFL cooperates with the protein methyltransferase PRMT7 in H19 imprinting control region methylation. PLoS biology 4, e355. Jirtle, R.L., and Skinner, M.K. (2007). Environmental epigenomics and disease susceptibility. Nature reviews 8, 253-262. Johnson, E.S. (2004). Protein modification by SUMO. Annual review of biochemistry 73, 355-382. Joseph, J., Tan, S.H., Karpova, T.S., McNally, J.G., and Dasso, M. (2002). SUMO-1 targets RanGAP1 to kinetochores and mitotic spindles. The Journal of cell biology 156, 595-602. Jurgens, G. (1985). A group of genes controlling the spatial expression of the bithorax complex in Drosophila. Nature 316, 153-155. Kagey, M.H., Melhuish, T.A., and Wotton, D. (2003). The polycomb protein Pc2 is a SUMO E3. Cell 113, 127-137. Kangaspeska, S., Stride, B., Metivier, R., Polycarpou-Schwarz, M., Ibberson, D., Carmouche, R.P., Benes, V., Gannon, F., and Reid, G. (2008). Transient cyclical methylation of promoter DNA. Nature 452, 112-115. Kao, C.F., Hillyer, C., Tsukuda, T., Henry, K., Berger, S., and Osley, M.A. (2004). Rad6 plays a role in transcriptional activation through ubiquitylation of histone H2B. Genes & development 18, 184-195. Katan-Khaykovich, Y., and Struhl, K. (2005). Heterochromatin formation involves changes in histone modifications over multiple cell generations. The EMBO journal 24, 2138-2149. Kim, C.A., Gingery, M., Pilpa, R.M., and Bowie, J.U. (2002). The SAM domain of polyhomeotic forms a helical polymer. Nature structural biology 9, 453-457. Klose, R.J., Kallin, E.M., and Zhang, Y. (2006). JmjC-domain-containing proteins and histone demethylation. Nature reviews 7, 715-727. Klymenko, T., Papp, B., Fischle, W., Kocher, T., Schelder, M., Fritsch, C., Wild, B., Wilm, M., and Muller, J. (2006). A Polycomb group protein complex with sequence-specific DNA-binding and selective methyl-lysine-binding activities. Genes & development 20, 1110-1122.

85
Kodjabachian, L., Delaage, M., Maurel, C., Miassod, R., Jacq, B., and Rosset, R. (1998). Mutations in ccf, a novel Drosophila gene encoding a chromosomal factor, affect progression through mitosis and interact with Pc-G mutations. The EMBO journal 17, 1063-1075. Kohlmaier, A., Savarese, F., Lachner, M., Martens, J., Jenuwein, T., and Wutz, A. (2004). A chromosomal memory triggered by Xist regulates histone methylation in X inactivation. PLoS biology 2, E171. Krauss, V. (2008). Glimpses of evolution: heterochromatic histone H3K9 methyltransferases left its marks behind. Genetica 133, 93-106. Kurzhals, R.L., Tie, F., Stratton, C.A., and Harte, P.J. (2008). Drosophila ESC-like can substitute for ESC and becomes required for Polycomb silencing if ESC is absent. Developmental biology 313, 293-306. Kwon, T., Chang, J.H., Kwak, E., Lee, C.W., Joachimiak, A., Kim, Y.C., Lee, J., and Cho, Y. (2003). Mechanism of histone lysine methyl transfer revealed by the structure of SET7/9-AdoMet. The EMBO journal 22, 292-303. Lan, F., Bayliss, P.E., Rinn, J.L., Whetstine, J.R., Wang, J.K., Chen, S., Iwase, S., Alpatov, R., Issaeva, I., Canaani, E., et al. (2007). A histone H3 lysine 27 demethylase regulates animal posterior development. Nature 449, 689-694. Lanzuolo, C., Roure, V., Dekker, J., Bantignies, F., and Orlando, V. (2007). Polycomb response elements mediate the formation of chromosome higher-order structures in the bithorax complex. Nature cell biology 9, 1167-1174. Le Guezennec, X., Vermeulen, M., Brinkman, A.B., Hoeijmakers, W.A., Cohen, A., Lasonder, E., and Stunnenberg, H.G. (2006). MBD2/NuRD and MBD3/NuRD, two distinct complexes with different biochemical and functional properties. Molecular and cellular biology 26, 843-851. Lee, J.H., Cook, J.R., Yang, Z.H., Mirochnitchenko, O., Gunderson, S.I., Felix, A.M., Herth, N., Hoffmann, R., and Pestka, S. (2005). PRMT7, a new protein arginine methyltransferase that synthesizes symmetric dimethylarginine. The Journal of biological chemistry 280, 3656-3664. Lee, M.G., Villa, R., Trojer, P., Norman, J., Yan, K.P., Reinberg, D., Di Croce, L., and Shiekhattar, R. (2007). Demethylation of H3K27 regulates polycomb recruitment and H2A ubiquitination. Science (New York, NY 318, 447-450. Legube, G., Linares, L.K., Tyteca, S., Caron, C., Scheffner, M., Chevillard-Briet, M., and Trouche, D. (2004). Role of the histone acetyl transferase Tip60 in the p53 pathway. The Journal of biological chemistry 279, 44825-44833. Lehembre, F., Badenhorst, P., Muller, S., Travers, A., Schweisguth, F., and Dejean, A. (2000). Covalent modification of the transcriptional repressor tramtrack by the ubiquitin-related protein Smt3 in Drosophila flies. Molecular and cellular biology 20, 1072-1082. Lewis, E.B. (1978). A gene complex controlling segmentation in Drosophila. Nature 276, 565-570. Li, E. (2002). Chromatin modification and epigenetic reprogramming in mammalian development. Nat Rev Genet 3, 662-673. Li, T., Hu, J.F., Qiu, X., Ling, J., Chen, H., Wang, S., Hou, A., Vu, T.H., and Hoffman, A.R. (2008). CTCF regulates allelic expression of Igf2 by orchestrating a promoter-polycomb repressive complex 2 intrachromosomal loop. Molecular and cellular biology 28, 6473-6482. Liang, G., Klose, R.J., Gardner, K.E., and Zhang, Y. (2007). Yeast Jhd2p is a histone H3 Lys4 trimethyl demethylase. Nature structural & molecular biology 14, 243-245. Lorente, M., Perez, C., Sanchez, C., Donohoe, M., Shi, Y., and Vidal, M. (2006). Homeotic transformations of the axial skeleton of YY1 mutant mice and genetic interaction with the Polycomb group gene Ring1/Ring1A. Mechanisms of development 123, 312-320.

86
Lupo, R., Breiling, A., Bianchi, M.E., and Orlando, V. (2001). Drosophila chromosome condensation proteins Topoisomerase II and Barren colocalize with Polycomb and maintain Fab-7 PRE silencing. Molecular cell 7, 127-136. MacPherson, M.J., Beatty, L.G., Zhou, W., Du, M., and Sadowski, P.D. (2009). The CTCF insulator protein is posttranslationally modified by SUMO. Molecular and cellular biology 29, 714-725. Mahadevan, L.C., Willis, A.C., and Barratt, M.J. (1991). Rapid histone H3 phosphorylation in response to growth factors, phorbol esters, okadaic acid, and protein synthesis inhibitors. Cell 65, 775-783. Malaspina, D., Corcoran, C., Kleinhaus, K.R., Perrin, M.C., Fennig, S., Nahon, D., Friedlander, Y., and Harlap, S. (2008). Acute maternal stress in pregnancy and schizophrenia in offspring: a cohort prospective study. BMC Psychiatry 8, 71. Martens, J.H., O'Sullivan, R.J., Braunschweig, U., Opravil, S., Radolf, M., Steinlein, P., and Jenuwein, T. (2005). The profile of repeat-associated histone lysine methylation states in the mouse epigenome. The EMBO journal 24, 800-812. Martin, C., and Zhang, Y. (2007). Mechanisms of epigenetic inheritance. Current opinion in cell biology 19, 266-272. Martinez, A.M., Colomb, S., Dejardin, J., Bantignies, F., and Cavalli, G. (2006). Polycomb group-dependent Cyclin A repression in Drosophila. Genes & development 20, 501-513. Mascle, X.H., Germain-Desprez, D., Huynh, P., Estephan, P., and Aubry, M. (2007). Sumoylation of the transcriptional intermediary factor 1beta (TIF1beta), the Co-repressor of the KRAB Multifinger proteins, is required for its transcriptional activity and is modulated by the KRAB domain. The Journal of biological chemistry 282, 10190-10202. Mateos-Langerak, J., and Cavalli, G. (2008). Polycomb group proteins and long-range gene regulation. Advances in genetics 61, 45-66. Matunis, M.J., Coutavas, E., and Blobel, G. (1996). A novel ubiquitin-like modification modulates the partitioning of the Ran-GTPase-activating protein RanGAP1 between the cytosol and the nuclear pore complex. The Journal of cell biology 135, 1457-1470. Metivier, R., Gallais, R., Tiffoche, C., Le Peron, C., Jurkowska, R.Z., Carmouche, R.P., Ibberson, D., Barath, P., Demay, F., Reid, G., et al. (2008). Cyclical DNA methylation of a transcriptionally active promoter. Nature 452, 45-50. Metzger, E., Wissmann, M., Yin, N., Muller, J.M., Schneider, R., Peters, A.H., Gunther, T., Buettner, R., and Schule, R. (2005). LSD1 demethylates repressive histone marks to promote androgen-receptor-dependent transcription. Nature 437, 436-439. Metzler-Guillemain, C., Depetris, D., Luciani, J.J., Mignon-Ravix, C., Mitchell, M.J., and Mattei, M.G. (2008). In human pachytene spermatocytes, SUMO protein is restricted to the constitutive heterochromatin. Chromosome Res 16, 761-782. Mikkelsen, T.S., Ku, M., Jaffe, D.B., Issac, B., Lieberman, E., Giannoukos, G., Alvarez, P., Brockman, W., Kim, T.K., Koche, R.P., et al. (2007). Genome-wide maps of chromatin state in pluripotent and lineage-committed cells. Nature 448, 553-560. Mohr, S.E., and Boswell, R.E. (1999). Zimp encodes a homologue of mouse Miz1 and PIAS3 and is an essential gene in Drosophila melanogaster. Gene 229, 109-116. Morgan, R., Pirard, P.M., Shears, L., Sohal, J., Pettengell, R., and Pandha, H.S. (2007). Antagonism of HOX/PBX dimer formation blocks the in vivo proliferation of melanoma. Cancer research 67, 5806-5813. Muller, J., Hart, C.M., Francis, N.J., Vargas, M.L., Sengupta, A., Wild, B., Miller, E.L., O'Connor, M.B., Kingston, R.E., and Simon, J.A. (2002). Histone methyltransferase activity of a Drosophila Polycomb group repressor complex. Cell 111, 197-208.

87
Negre, N., Hennetin, J., Sun, L.V., Lavrov, S., Bellis, M., White, K.P., and Cavalli, G. (2006). Chromosomal distribution of PcG proteins during Drosophila development. PLoS biology 4, e170. Nekrasov, M., Klymenko, T., Fraterman, S., Papp, B., Oktaba, K., Kocher, T., Cohen, A., Stunnenberg, H.G., Wilm, M., and Muller, J. (2007). Pcl-PRC2 is needed to generate high levels of H3-K27 trimethylation at Polycomb target genes. The EMBO journal 26, 4078-4088. Ng, H.H., Robert, F., Young, R.A., and Struhl, K. (2003). Targeted recruitment of Set1 histone methylase by elongating Pol II provides a localized mark and memory of recent transcriptional activity. Molecular cell 11, 709-719. Nickel, B.E., and Davie, J.R. (1989). Structure of polyubiquitinated histone H2A. Biochemistry 28, 964-968. O'Dor, E., Beck, S.A., and Brock, H.W. (2006). Polycomb group mutants exhibit mitotic defects in syncytial cell cycles of Drosophila embryos. Developmental biology 290, 312-322. Ohno, K., McCabe, D., Czermin, B., Imhof, A., and Pirrotta, V. (2008). ESC, ESCL and their roles in Polycomb Group mechanisms. Mechanisms of development 125, 527-541. Ohtsubo, M., Yasunaga, S., Ohno, Y., Tsumura, M., Okada, S., Ishikawa, N., Shirao, K., Kikuchi, A., Nishitani, H., Kobayashi, M., et al. (2008). Polycomb-group complex 1 acts as an E3 ubiquitin ligase for Geminin to sustain hematopoietic stem cell activity. Proceedings of the National Academy of Sciences of the United States of America 105, 10396-10401. Oktaba, K., Gutierrez, L., Gagneur, J., Girardot, C., Sengupta, A.K., Furlong, E.E., and Muller, J. (2008). Dynamic Regulation by Polycomb Group Protein Complexes Controls Pattern Formation and the Cell Cycle in Drosophila. Developmental cell. Paro, R., and Zink, B. (1993). The Polycomb gene is differentially regulated during oogenesis and embryogenesis of Drosophila melanogaster. Mechanisms of development 40, 37-46. Patel, J.H., Du, Y., Ard, P.G., Phillips, C., Carella, B., Chen, C.J., Rakowski, C., Chatterjee, C., Lieberman, P.M., Lane, W.S., et al. (2004). The c-MYC oncoprotein is a substrate of the acetyltransferases hGCN5/PCAF and TIP60. Molecular and cellular biology 24, 10826-10834. Pham, A.D., and Sauer, F. (2000). Ubiquitin-activating/conjugating activity of TAFII250, a mediator of activation of gene expression in Drosophila. Science (New York, NY 289, 2357-2360. Pickart, C.M. (2001). Mechanisms underlying ubiquitination. Annual review of biochemistry 70, 503-533. Plath, K., Fang, J., Mlynarczyk-Evans, S.K., Cao, R., Worringer, K.A., Wang, H., de la Cruz, C.C., Otte, A.P., Panning, B., and Zhang, Y. (2003). Role of histone H3 lysine 27 methylation in X inactivation. Science (New York, NY 300, 131-135. Ptashne, M. (2005). Regulation of transcription: from lambda to eukaryotes. Trends in biochemical sciences 30, 275-279. Quenstedt, M., and Parshall, A. (1998). Prenatal exposure to maternal stress and subsequent schizophrenia. Br J Psychiatry 173, 183. Randsholt, N.B., Maschat, F., and Santamaria, P. (2000). polyhomeotic controls engrailed expression and the hedgehog signaling pathway in imaginal discs. Mechanisms of development 95, 89-99. Rea, S., Eisenhaber, F., O'Carroll, D., Strahl, B.D., Sun, Z.W., Schmid, M., Opravil, S., Mechtler, K., Ponting, C.P., Allis, C.D., et al. (2000). Regulation of chromatin structure by site-specific histone H3 methyltransferases. Nature 406, 593-599. Reik, W. (1989). Genomic imprinting and genetic disorders in man. Trends Genet 5, 331-336. Reynolds, G. (2000). Symposia: maternal stress, motor neuron disease, antidepressants, and schizophrenia. IDrugs 3, 1144-1145. Riggs, A.D. (1975). X inactivation, differentiation, and DNA methylation. Cytogenet Cell Genet 14, 9-25.

88
Riising, E.M., Boggio, R., Chiocca, S., Helin, K., and Pasini, D. (2008). The polycomb repressive complex 2 is a potential target of SUMO modifications. PLoS ONE 3, e2704. Ringrose, L., and Paro, R. (2007). Polycomb/Trithorax response elements and epigenetic memory of cell identity. Development (Cambridge, England) 134, 223-232. Salser, S.J., and Kenyon, C. (1996). A C. elegans Hox gene switches on, off, on and off again to regulate proliferation, differentiation and morphogenesis. Development (Cambridge, England) 122, 1651-1661. Salvaing, J., Nagel, A.C., Mouchel-Vielh, E., Bloyer, S., Maier, D., Preiss, A., and Peronnet, F. (2008). The enhancer of trithorax and polycomb corto interacts with cyclin G in Drosophila. PLoS ONE 3, e1658. Sanchez-Pernaute, O., Ospelt, C., Neidhart, M., and Gay, S. (2008). Epigenetic clues to rheumatoid arthritis. Journal of autoimmunity 30, 12-20. Santos-Rosa, H., Schneider, R., Bannister, A.J., Sherriff, J., Bernstein, B.E., Emre, N.C., Schreiber, S.L., Mellor, J., and Kouzarides, T. (2002). Active genes are tri-methylated at K4 of histone H3. Nature 419, 407-411. Sarcinella, E., Zuzarte, P.C., Lau, P.N., Draker, R., and Cheung, P. (2007). Monoubiquitylation of H2A.Z distinguishes its association with euchromatin or facultative heterochromatin. Molecular and cellular biology 27, 6457-6468. Saurin, A.J., Shao, Z., Erdjument-Bromage, H., Tempst, P., and Kingston, R.E. (2001). A Drosophila Polycomb group complex includes Zeste and dTAFII proteins. Nature 412, 655-660. Savla, U., Benes, J., Zhang, J., and Jones, R.S. (2008). Recruitment of Drosophila Polycomb-group proteins by Polycomblike, a component of a novel protein complex in larvae. Development (Cambridge, England) 135, 813-817. Schneider, R., Bannister, A.J., Myers, F.A., Thorne, A.W., Crane-Robinson, C., and Kouzarides, T. (2004). Histone H3 lysine 4 methylation patterns in higher eukaryotic genes. Nature cell biology 6, 73-77. Schubeler, D., MacAlpine, D.M., Scalzo, D., Wirbelauer, C., Kooperberg, C., van Leeuwen, F., Gottschling, D.E., O'Neill, L.P., Turner, B.M., Delrow, J., et al. (2004). The histone modification pattern of active genes revealed through genome-wide chromatin analysis of a higher eukaryote. Genes & development 18, 1263-1271. Schwartz, Y.B., Kahn, T.G., Nix, D.A., Li, X.Y., Bourgon, R., Biggin, M., and Pirrotta, V. (2006). Genome-wide analysis of Polycomb targets in Drosophila melanogaster. Nature genetics 38, 700-705. Schwartz, Y.B., and Pirrotta, V. (2008). Polycomb complexes and epigenetic states. Current opinion in cell biology 20, 266-273. Scott, E.K., Lee, T., and Luo, L. (2001). enok encodes a Drosophila putative histone acetyltransferase required for mushroom body neuroblast proliferation. Curr Biol 11, 99-104. Seto, E., Shi, Y., and Shenk, T. (1991). YY1 is an initiator sequence-binding protein that directs and activates transcription in vitro. Nature 354, 241-245. Seward, D.J., Cubberley, G., Kim, S., Schonewald, M., Zhang, L., Tripet, B., and Bentley, D.L. (2007). Demethylation of trimethylated histone H3 Lys4 in vivo by JARID1 JmjC proteins. Nature structural & molecular biology 14, 240-242. Shao, Z., Raible, F., Mollaaghababa, R., Guyon, J.R., Wu, C.T., Bender, W., and Kingston, R.E. (1999). Stabilization of chromatin structure by PRC1, a Polycomb complex. Cell 98, 37-46. Shareef, M.M., King, C., Damaj, M., Badagu, R., Huang, D.W., and Kellum, R. (2001). Drosophila heterochromatin protein 1 (HP1)/origin recognition complex (ORC) protein is associated with HP1 and ORC and functions in heterochromatin-induced silencing. Molecular biology of the cell 12, 1671-1685.

89
Shi, Y., Lan, F., Matson, C., Mulligan, P., Whetstine, J.R., Cole, P.A., and Casero, R.A. (2004a). Histone demethylation mediated by the nuclear amine oxidase homolog LSD1. Cell 119, 941-953. Shi, Y., Lan, F., Matson, C., Mulligan, P., Whetstine, J.R., Cole, P.A., Casero, R.A., and Shi, Y. (2004b). Histone demethylation mediated by the nuclear amine oxidase homolog LSD1. Cell 119, 941-953. Shi, Y., Seto, E., Chang, L.S., and Shenk, T. (1991). Transcriptional repression by YY1, a human GLI-Kruppel-related protein, and relief of repression by adenovirus E1A protein. Cell 67, 377-388. Shiio, Y., and Eisenman, R.N. (2003). Histone sumoylation is associated with transcriptional repression. Proceedings of the National Academy of Sciences of the United States of America 100, 13225-13230. Sims, J.K., Houston, S.I., Magazinnik, T., and Rice, J.C. (2006). A trans-tail histone code defined by monomethylated H4 Lys-20 and H3 Lys-9 demarcates distinct regions of silent chromatin. The Journal of biological chemistry 281, 12760-12766. Solter, D., Hiiragi, T., Evsikov, A.V., Moyer, J., De Vries, W.N., Peaston, A.E., and Knowles, B.B. (2004). Epigenetic mechanisms in early mammalian development. Cold Spring Harbor symposia on quantitative biology 69, 11-17. Sparmann, A., and van Lohuizen, M. (2006). Polycomb silencers control cell fate, development and cancer. Nat Rev Cancer 6, 846-856. Tachibana, M., Ueda, J., Fukuda, M., Takeda, N., Ohta, T., Iwanari, H., Sakihama, T., Kodama, T., Hamakubo, T., and Shinkai, Y. (2005). Histone methyltransferases G9a and GLP form heteromeric complexes and are both crucial for methylation of euchromatin at H3-K9. Genes & development 19, 815-826. Tagami, H., Ray-Gallet, D., Almouzni, G., and Nakatani, Y. (2004). Histone H3.1 and H3.3 complexes mediate nucleosome assembly pathways dependent or independent of DNA synthesis. Cell 116, 51-61. Thomas, T., and Voss, A.K. (2007). The diverse biological roles of MYST histone acetyltransferase family proteins. Cell cycle (Georgetown, Tex 6, 696-704. Tie, F., Furuyama, T., and Harte, P.J. (1998). The Drosophila Polycomb Group proteins ESC and E(Z) bind directly to each other and co-localize at multiple chromosomal sites. Development (Cambridge, England) 125, 3483-3496. Tie, F., Furuyama, T., Prasad-Sinha, J., Jane, E., and Harte, P.J. (2001). The Drosophila Polycomb Group proteins ESC and E(Z) are present in a complex containing the histone-binding protein p55 and the histone deacetylase RPD3. Development (Cambridge, England) 128, 275-286. Tie, F., Prasad-Sinha, J., Birve, A., Rasmuson-Lestander, A., and Harte, P.J. (2003). A 1-megadalton ESC/E(Z) complex from Drosophila that contains polycomblike and RPD3. Mol Cell Biol 23, 3352-3362. Tiwari, V.K., Cope, L., McGarvey, K.M., Ohm, J.E., and Baylin, S.B. (2008a). A novel 6C assay uncovers Polycomb-mediated higher order chromatin conformations. Genome research 18, 1171-1179. Tiwari, V.K., McGarvey, K.M., Licchesi, J.D., Ohm, J.E., Herman, J.G., Schubeler, D., and Baylin, S.B. (2008b). PcG proteins, DNA methylation, and gene repression by chromatin looping. PLoS biology 6, 2911-2927. Tolhuis, B., de Wit, E., Muijrers, I., Teunissen, H., Talhout, W., van Steensel, B., and van Lohuizen, M. (2006). Genome-wide profiling of PRC1 and PRC2 Polycomb chromatin binding in Drosophila melanogaster. Nature genetics 38, 694-699. Tsukada, Y., Fang, J., Erdjument-Bromage, H., Warren, M.E., Borchers, C.H., Tempst, P., and Zhang, Y. (2006). Histone demethylation by a family of JmjC domain-containing proteins. Nature 439, 811-816.

90
Uv, A.E., Thompson, C.R., and Bray, S.J. (1994). The Drosophila tissue-specific factor Grainyhead contains novel DNA-binding and dimerization domains which are conserved in the human protein CP2. Mol Cell Biol 14, 4020-4031. Vakoc, C.R., Mandat, S.A., Olenchock, B.A., and Blobel, G.A. (2005). Histone H3 lysine 9 methylation and HP1gamma are associated with transcription elongation through mammalian chromatin. Molecular cell 19, 381-391. Vakoc, C.R., Sachdeva, M.M., Wang, H., and Blobel, G.A. (2006). Profile of histone lysine methylation across transcribed mammalian chromatin. Molecular and cellular biology 26, 9185-9195. van der Woude, M.W., and Baumler, A.J. (2004). Phase and antigenic variation in bacteria. Clinical microbiology reviews 17, 581-611, table of contents. van Dijk, K., Marley, K.E., Jeong, B.R., Xu, J., Hesson, J., Cerny, R.L., Waterborg, J.H., and Cerutti, H. (2005). Monomethyl histone H3 lysine 4 as an epigenetic mark for silenced euchromatin in Chlamydomonas. The Plant cell 17, 2439-2453. van Os, J., and Selten, J.P. (1998). Prenatal exposure to maternal stress and subsequent schizophrenia. The May 1940 invasion of The Netherlands. Br J Psychiatry 172, 324-326. Vaquero, A., Sternglanz, R., and Reinberg, D. (2007). NAD+-dependent deacetylation of H4 lysine 16 by class III HDACs. Oncogene 26, 5505-5520. Vetting, M.W., LP, S.d.C., Yu, M., Hegde, S.S., Magnet, S., Roderick, S.L., and Blanchard, J.S. (2005). Structure and functions of the GNAT superfamily of acetyltransferases. Archives of biochemistry and biophysics 433, 212-226. Vire, E., Brenner, C., Deplus, R., Blanchon, L., Fraga, M., Didelot, C., Morey, L., Van Eynde, A., Bernard, D., Vanderwinden, J.M., et al. (2006). The Polycomb group protein EZH2 directly controls DNA methylation. Nature 439, 871-874. Volpe, T.A., Kidner, C., Hall, I.M., Teng, G., Grewal, S.I., and Martienssen, R.A. (2002). Regulation of heterochromatic silencing and histone H3 lysine-9 methylation by RNAi. Science (New York, NY 297, 1833-1837. Voncken, J.W., Roelen, B.A., Roefs, M., de Vries, S., Verhoeven, E., Marino, S., Deschamps, J., and van Lohuizen, M. (2003). Rnf2 (Ring1b) deficiency causes gastrulation arrest and cell cycle inhibition. Proceedings of the National Academy of Sciences of the United States of America 100, 2468-2473. Wang, H., Huang, Z.Q., Xia, L., Feng, Q., Erdjument-Bromage, H., Strahl, B.D., Briggs, S.D., Allis, C.D., Wong, J., Tempst, P., et al. (2001). Methylation of histone H4 at arginine 3 facilitating transcriptional activation by nuclear hormone receptor. Science (New York, NY 293, 853-857. Wang, Y., Wysocka, J., Sayegh, J., Lee, Y.H., Perlin, J.R., Leonelli, L., Sonbuchner, L.S., McDonald, C.H., Cook, R.G., Dou, Y., et al. (2004). Human PAD4 regulates histone arginine methylation levels via demethylimination. Science (New York, NY 306, 279-283. Weake, V.M., and Workman, J.L. (2008). Histone ubiquitination: triggering gene activity. Molecular cell 29, 653-663. West, M.H., and Bonner, W.M. (1980). Histone 2B can be modified by the attachment of ubiquitin. Nucleic acids research 8, 4671-4680. Whetstine, J.R., Nottke, A., Lan, F., Huarte, M., Smolikov, S., Chen, Z., Spooner, E., Li, E., Zhang, G., Colaiacovo, M., et al. (2006). Reversal of histone lysine trimethylation by the JMJD2 family of histone demethylases. Cell 125, 467-481. Whitcomb, S.J., Basu, A., Allis, C.D., and Bernstein, E. (2007). Polycomb Group proteins: an evolutionary perspective. Trends Genet 23, 494-502.

91
Wood, A., Krogan, N.J., Dover, J., Schneider, J., Heidt, J., Boateng, M.A., Dean, K., Golshani, A., Zhang, Y., Greenblatt, J.F., et al. (2003). Bre1, an E3 ubiquitin ligase required for recruitment and substrate selection of Rad6 at a promoter. Molecular cell 11, 267-274. Wu, X., Gong, Y., Yue, J., Qiang, B., Yuan, J., and Peng, X. (2008). Cooperation between EZH2, NSPc1-mediated histone H2A ubiquitination and Dnmt1 in HOX gene silencing. Nucleic acids research 36, 3590-3599. Xiang, Y., Zhu, Z., Han, G., Lin, H., Xu, L., and Chen, C.D. (2007). JMJD3 is a histone H3K27 demethylase. Cell research 17, 850-857. Yajnik, C.S. (2004). Early life origins of insulin resistance and type 2 diabetes in India and other Asian countries. The Journal of nutrition 134, 205-210. Youdell, M.L., Kizer, K.O., Kisseleva-Romanova, E., Fuchs, S.M., Duro, E., Strahl, B.D., and Mellor, J. (2008). Roles for Ctk1 and Spt6 in regulating the different methylation states of histone H3 lysine 36. Molecular and cellular biology 28, 4915-4926. Zhang, H., Smolen, G.A., Palmer, R., Christoforou, A., van den Heuvel, S., and Haber, D.A. (2004). SUMO modification is required for in vivo Hox gene regulation by the Caenorhabditis elegans Polycomb group protein SOP-2. Nature genetics 36, 507-511. Zhang, Y. (2003). Transcriptional regulation by histone ubiquitination and deubiquitination. Genes & development 17, 2733-2740. Zhao, Z., Tavoosidana, G., Sjolinder, M., Gondor, A., Mariano, P., Wang, S., Kanduri, C., Lezcano, M., Sandhu, K.S., Singh, U., et al. (2006). Circular chromosome conformation capture (4C) uncovers extensive networks of epigenetically regulated intra- and interchromosomal interactions. Nature genetics 38, 1341-1347. Zhou, W., Wang, X., and Rosenfeld, M.G. (2009). Histone H2A ubiquitination in transcriptional regulation and DNA damage repair. Int J Biochem Cell Biol 41, 12-15. Zhou, W., Zhu, P., Wang, J., Pascual, G., Ohgi, K.A., Lozach, J., Glass, C.K., and Rosenfeld, M.G. (2008). Histone H2A monoubiquitination represses transcription by inhibiting RNA polymerase II transcriptional elongation. Molecular cell 29, 69-80.














![NEUROSCIENCE Copyright © 2020 Persistent epigenetic … · (H3K27) methyltransferase enhancer of zeste [E(z)] and the obligate accessory factors suppressor of zeste 12 [Su(z)12]](https://img.pdfslide.us/doc/110x75/60b801fa57ab57285d32d107/neuroscience-copyright-2020-persistent-epigenetic-h3k27-methyltransferase-enhancer.jpg)















