Embed Size (px)
Citation preview

www.elsevier.com/locate/ygeno
Genomics 86 (2
Expression analysis of SIX3 and SIX6 in human tissues reveals differences
in expression and a novel correlation between the expression of SIX3 and
the genes encoding isocitrate dehydrogenase and cadherin 18
Saima Aijaza, Jennifer Allenb, Robert Tregidgob, Veronica van Heyningenc,
Isabel Hansond, Brian J. Clarka,b,e,TaInstitute of Ophthalmology, University College London, Bath Street, London EC1V 9EL, UK
bPharmagene Laboratories Ltd., 2 Orchard Road, Royston, Hertfordshire SG8 5HD, UKcMRC Human Genetics Unit, Western General Hospital, Edinburgh EH4 2XU, UK
dDepartment of Medical Genetics, University of Edinburgh, Western General Hospital, Edinburgh EH4 2XU, UKeMoorfields Eye Hospital, City Road, London, UK
Received 19 November 2004; accepted 3 March 2005
Available online 18 April 2005
Abstract
SIX3 and SIX6 are transcription factors expressed during early stages of eye development. Limited expression data for SIX3 and
SIX6 are available in the literature but, to date, there are no reports of the relative levels of expression of these genes throughout the
human body and in adult tissues in particular. In this paper, we report extensive real-time quantitative PCR analyses of SIX3 and SIX6
expression in many different tissues of the adult human body, including ocular tissues, and a comparison of expression data with that of
many other genes to identify similarity in expression. Using this powerful technique, we have detected a novel statistical correlation
between the spatial distribution and the quantitative expression of SIX3 and 5 other transcripts including IDH1, the gene encoding the
NADP+-dependent enzyme isocitrate dehydrogenase, and cadherin 18, type 2 (CDH14). Our data demonstrate that this novel technique
can be used to generate hypotheses by comparison of gene expression profiles to identify possible interactions between genes or gene
products.
D 2005 Elsevier Inc. All rights reserved.
Keywords: SIX3; SIX6; Expression analysis; Real-time quantitative PCR
Introduction
Eye development is a multistep process that is precisely
regulated by genes that are highly conserved throughout
evolution. Many of these genes were identified as a result
of their role in eye formation in Drosophila melanogaster
[1–3]. In Drosophila eye morphogenesis, an interactive
network of the regulatory genes comprising eyeless (ey),
eyes absent (eya), sine oculis (so), and dachshund (dac) is
0888-7543/$ - see front matter D 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.ygeno.2005.03.002
* Corresponding author. National Cancer Tissue Resource, Room G15,
Cancer Research UK, 44 Lincoln’s Inn Fields, London WC2A 3PX, UK.
Fax: +44 (0) 20 7269 3111.
E-mail address: [email protected] (B.J. Clark).
required for normal eye formation and targeted ectopic
expression of some of these genes induces the formation of
ectopic eyes [4–11]. Vertebrate homologues of all these
genes have been identified, which include Pax6 (ey), the
Dach gene family (dac), the Eya gene family (eya), and the
Six gene family (so) [12–17].
To date, 6 members of the Six gene family have been
identified in mice (Six1-6) [18–23] and humans (SIX1-6)
[24] and these code for transcription factors characterized
by the presence of a homeodomain and a six domain [25].
Of these, only Six3 and Six6 are expressed during early
stages of eye development [17,26–28]. The mouse Six3
gene was originally considered to be the functional
homologue of sine oculis [17]. However, phylogenetic
005) 86 – 99

S. Aijaz et al. / Genomics 86 (2005) 86–99 87
analysis of two additional Six genes in Drosophila, Optix,
and Dsix4 has shown that Six3/Six6 are more closely
related to Optix than to sine oculis, which is more closely
related to Six1/Six2 [29,30]. Unlike sine oculis, which
requires interaction with the protein product of the eya
gene to regulate multiple steps in Drosophila eye develop-
ment, neither optix nor Six3 interact with eya proteins but
each is capable of inducing ectopic eyes upon over-
expression through a mechanism independent of eya
[10,31–35]. These data suggest that Six3/Six6 may act
via a different molecular network and require interacting
proteins different from those identified for sine oculis.
Experiments using yeast two hybrid screens have shown
that both Six3 and Six6 proteins interact with transcrip-
tional repressors of the groucho family [36–38]. More
recently, it has been reported that Six3 directly interacts
with the DNA replication inhibitor, geminin, acting
antagonistically with geminin to regulate the balance
between proliferation and differentiation in early vertebrate
eye development [39]. In the same study, Six6 was also
shown to interact with geminin [39], suggesting that both
Six3 and Six6 might function through similar mechanisms.
In addition, Six6 has been shown to bind to the Kip1
promoter in retinal cells in association with several
corepressors including Dach2 [40]. In the Six6 mutant
mouse, there is upregulation of Kip1 mRNA and protein in
the retinal progenitor cells and premature cell cycle exit
implicating a direct role of Six6 in regulating the cell cycle
[40].
Consistent with its role in cell proliferation, overexpres-
sion of Six3 in medaka, zebrafish, Xenopus, and mouse
results in the expansion of the rostral forebrain, formation of
ectopic optic vesicles in the neural tube region that gives
rise to the midbrain and hindbrain, and expansion of the
normal optic vesicles while inactivation of Six3 results in
the loss of forebrain structures including the retina
[34,35,41–43]. Overexpression of Six6 in Xenopus has
been shown to induce ectopic retinal tissue and expand the
optic vesicle and the rostral neuroepithelium [43,44]. These
studies suggest that the functional roles of Six3 and Six6 are
distinct from each other and implicate Six3 as having a
broader role in head development (including the forebrain
Table 1
Amplicon design parameters used in qRT-PCR
Gene Primer Sequence
SIX3 Forward primer CCTCCCACTTCTTGTTGCCA
Reverse primer CGCTACTCGCCAGAAGTATG
Probe CTTCGCCGATTCTCACCACC
SIX6 Forward primer CGGCCACTTCAGCCATCT
Reverse primer CCTGGATGGGCAACTCAGA
Probe CATCACGTCCAGCGACAGCG
GAPDH Forward primer GAAGGTGAAGGTCGGAGTC
Reverse primer CAGAGTTAAAAGCAGCCCTG
Probe TTTGGTCGTATTGGGCGCCT
and eye formation) while Six6 appears to have a more
specific role in eye development.
Human SIX3 and SIX6 have been mapped to chromo-
some 2p16-p21 and 14q22.3-q23, respectively [30,45].
Consistent with its role in head development, mutations in
SIX3 have been shown to cause holoprosencephaly type II
[46,47]. SIX6 has been associated with anophthalmia and
pituitary anomalies in one study [30], whereas in others
there is no etiological link with developmental eye
anomalies of the microphthalmia, anophthalmia, and colo-
boma spectrum [48,49]. Expression studies in medaka,
zebrafish, Xenopus, chick, and mouse have shown that early
expression of Six3 is localized in the anterior neuroectoderm
and is later maintained in the forebrain, eye anlage, lens
placode, olfactory primordium, and the hypothalamus
[17,50–53]. In contrast, the expression of Six6 begins later
than Six3 in a more restricted pattern and is detected in the
ventral diencephalon, pituitary gland, optic stalk, and neural
retina [23,26–28,51,54].
Human SIX3 mRNA has been localized in the fetal eye
from 5 weeks of gestation by RT-PCR in the ganglion cell
layer and in the inner nuclear layer of the 22 week fetal eye
as well as the adult human retina [45]. RT-PCR analyses of
human SIX6 expression have shown that it is expressed
between 12 and 14 weeks of gestation, and in situ
hybridization localized the RNA to the ganglion cell layer
and the inner nuclear layer of the 19 week fetal eye [30]. To
date, there are no reports of SIX6 expression in the adult
human eye. Moreover, despite their well-studied roles in
early eye development, there is no information regarding the
relative levels and distribution of SIX3 and SIX6 expression
in the adult human eye or other nonocular tissues. Here we
report, for the first time, real-time quantitative RT-PCR
analyses of SIX3 and SIX6 expression in the adult human
eye, and compare the ocular expression of these genes with
that in 72 different nonocular human body tissues. We have
also detected a novel correlation between the spatial
distribution and the quantitative expression of SIX3 and
IDH1, the gene encoding the NADP+-dependent enzyme
isocitrate dehydrogenase in both the eye and the rest of the
body and a further correlation between SIX3 and cadherin
18, type 2 (CDH14).
Amplicon size (bp) Annealing temperature
66 60-C
G
GC
67 60-C
AGTG
AAC 71 60-CGT
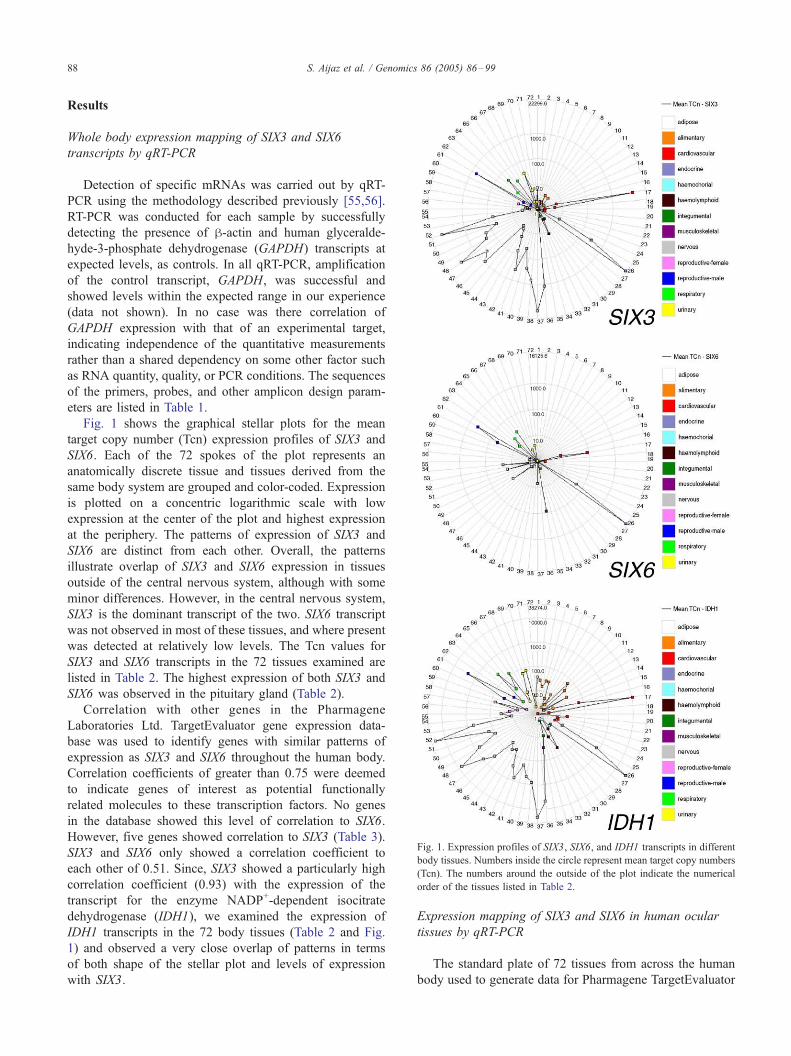
Fig. 1. Expression profiles of SIX3, SIX6, and IDH1 transcripts in different
body tissues. Numbers inside the circle represent mean target copy numbers
(Tcn). The numbers around the outside of the plot indicate the numerical
order of the tissues listed in Table 2.
S. Aijaz et al. / Genomics 86 (2005) 86–9988
Results
Whole body expression mapping of SIX3 and SIX6
transcripts by qRT-PCR
Detection of specific mRNAs was carried out by qRT-
PCR using the methodology described previously [55,56].
RT-PCR was conducted for each sample by successfully
detecting the presence of h-actin and human glyceralde-
hyde-3-phosphate dehydrogenase (GAPDH) transcripts at
expected levels, as controls. In all qRT-PCR, amplification
of the control transcript, GAPDH, was successful and
showed levels within the expected range in our experience
(data not shown). In no case was there correlation of
GAPDH expression with that of an experimental target,
indicating independence of the quantitative measurements
rather than a shared dependency on some other factor such
as RNA quantity, quality, or PCR conditions. The sequences
of the primers, probes, and other amplicon design param-
eters are listed in Table 1.
Fig. 1 shows the graphical stellar plots for the mean
target copy number (Tcn) expression profiles of SIX3 and
SIX6. Each of the 72 spokes of the plot represents an
anatomically discrete tissue and tissues derived from the
same body system are grouped and color-coded. Expression
is plotted on a concentric logarithmic scale with low
expression at the center of the plot and highest expression
at the periphery. The patterns of expression of SIX3 and
SIX6 are distinct from each other. Overall, the patterns
illustrate overlap of SIX3 and SIX6 expression in tissues
outside of the central nervous system, although with some
minor differences. However, in the central nervous system,
SIX3 is the dominant transcript of the two. SIX6 transcript
was not observed in most of these tissues, and where present
was detected at relatively low levels. The Tcn values for
SIX3 and SIX6 transcripts in the 72 tissues examined are
listed in Table 2. The highest expression of both SIX3 and
SIX6 was observed in the pituitary gland (Table 2).
Correlation with other genes in the Pharmagene
Laboratories Ltd. TargetEvaluator gene expression data-
base was used to identify genes with similar patterns of
expression as SIX3 and SIX6 throughout the human body.
Correlation coefficients of greater than 0.75 were deemed
to indicate genes of interest as potential functionally
related molecules to these transcription factors. No genes
in the database showed this level of correlation to SIX6.
However, five genes showed correlation to SIX3 (Table 3).
SIX3 and SIX6 only showed a correlation coefficient to
each other of 0.51. Since, SIX3 showed a particularly high
correlation coefficient (0.93) with the expression of the
transcript for the enzyme NADP+-dependent isocitrate
dehydrogenase (IDH1), we examined the expression of
IDH1 transcripts in the 72 body tissues (Table 2 and Fig.
1) and observed a very close overlap of patterns in terms
of both shape of the stellar plot and levels of expression
with SIX3.
Expression mapping of SIX3 and SIX6 in human ocular
tissues by qRT-PCR
The standard plate of 72 tissues from across the human
body used to generate data for Pharmagene TargetEvaluator
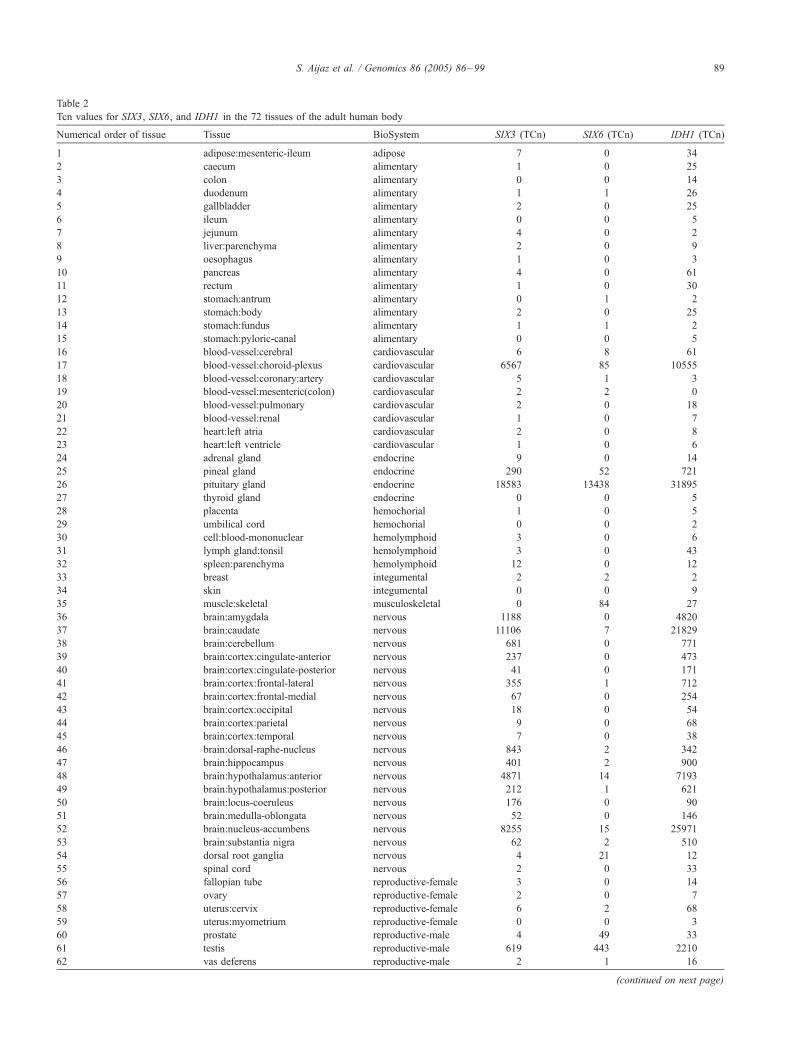
Table 2
Tcn values for SIX3, SIX6, and IDH1 in the 72 tissues of the adult human body
Numerical order of tissue Tissue BioSystem SIX3 (TCn) SIX6 (TCn) IDH1 (TCn)
1 adipose:mesenteric-ileum adipose 7 0 34
2 caecum alimentary 1 0 25
3 colon alimentary 0 0 14
4 duodenum alimentary 1 1 26
5 gallbladder alimentary 2 0 25
6 ileum alimentary 0 0 5
7 jejunum alimentary 4 0 2
8 liver:parenchyma alimentary 2 0 9
9 oesophagus alimentary 1 0 3
10 pancreas alimentary 4 0 61
11 rectum alimentary 1 0 30
12 stomach:antrum alimentary 0 1 2
13 stomach:body alimentary 2 0 25
14 stomach:fundus alimentary 1 1 2
15 stomach:pyloric-canal alimentary 0 0 5
16 blood-vessel:cerebral cardiovascular 6 8 61
17 blood-vessel:choroid-plexus cardiovascular 6567 85 10555
18 blood-vessel:coronary:artery cardiovascular 5 1 3
19 blood-vessel:mesenteric(colon) cardiovascular 2 2 0
20 blood-vessel:pulmonary cardiovascular 2 0 18
21 blood-vessel:renal cardiovascular 1 0 7
22 heart:left atria cardiovascular 2 0 8
23 heart:left ventricle cardiovascular 1 0 6
24 adrenal gland endocrine 9 0 14
25 pineal gland endocrine 290 52 721
26 pituitary gland endocrine 18583 13438 31895
27 thyroid gland endocrine 0 0 5
28 placenta hemochorial 1 0 5
29 umbilical cord hemochorial 0 0 2
30 cell:blood-mononuclear hemolymphoid 3 0 6
31 lymph gland:tonsil hemolymphoid 3 0 43
32 spleen:parenchyma hemolymphoid 12 0 12
33 breast integumental 2 2 2
34 skin integumental 0 0 9
35 muscle:skeletal musculoskeletal 0 84 27
36 brain:amygdala nervous 1188 0 4820
37 brain:caudate nervous 11106 7 21829
38 brain:cerebellum nervous 681 0 771
39 brain:cortex:cingulate-anterior nervous 237 0 473
40 brain:cortex:cingulate-posterior nervous 41 0 171
41 brain:cortex:frontal-lateral nervous 355 1 712
42 brain:cortex:frontal-medial nervous 67 0 254
43 brain:cortex:occipital nervous 18 0 54
44 brain:cortex:parietal nervous 9 0 68
45 brain:cortex:temporal nervous 7 0 38
46 brain:dorsal-raphe-nucleus nervous 843 2 342
47 brain:hippocampus nervous 401 2 900
48 brain:hypothalamus:anterior nervous 4871 14 7193
49 brain:hypothalamus:posterior nervous 212 1 621
50 brain:locus-coeruleus nervous 176 0 90
51 brain:medulla-oblongata nervous 52 0 146
52 brain:nucleus-accumbens nervous 8255 15 25971
53 brain:substantia nigra nervous 62 2 510
54 dorsal root ganglia nervous 4 21 12
55 spinal cord nervous 2 0 33
56 fallopian tube reproductive-female 3 0 14
57 ovary reproductive-female 2 0 7
58 uterus:cervix reproductive-female 6 2 68
59 uterus:myometrium reproductive-female 0 0 3
60 prostate reproductive-male 4 49 33
61 testis reproductive-male 619 443 2210
62 vas deferens reproductive-male 2 1 16
(continued on next page)
S. Aijaz et al. / Genomics 86 (2005) 86–99 89
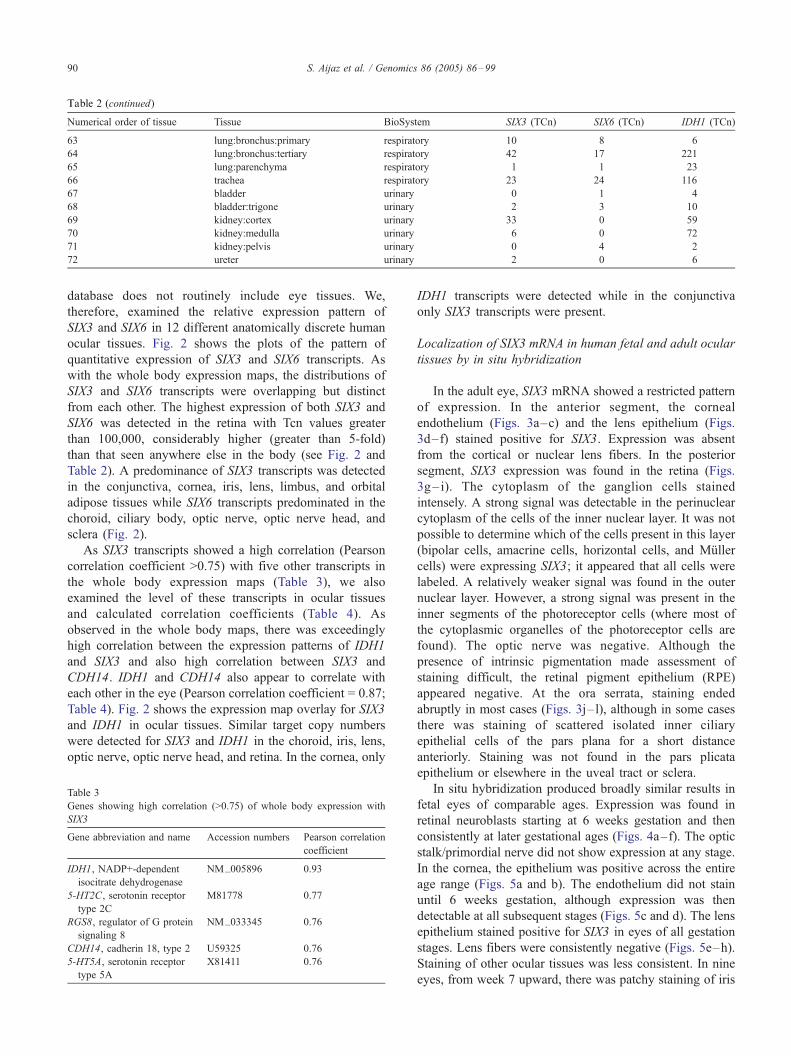
Numerical order of tissue Tissue BioSystem SIX3 (TCn) SIX6 (TCn) IDH1 (TCn)
63 lung:bronchus:primary respiratory 10 8 6
64 lung:bronchus:tertiary respiratory 42 17 221
65 lung:parenchyma respiratory 1 1 23
66 trachea respiratory 23 24 116
67 bladder urinary 0 1 4
68 bladder:trigone urinary 2 3 10
69 kidney:cortex urinary 33 0 59
70 kidney:medulla urinary 6 0 72
71 kidney:pelvis urinary 0 4 2
72 ureter urinary 2 0 6
Table 2 (continued)
S. Aijaz et al. / Genomics 86 (2005) 86–9990
database does not routinely include eye tissues. We,
therefore, examined the relative expression pattern of
SIX3 and SIX6 in 12 different anatomically discrete human
ocular tissues. Fig. 2 shows the plots of the pattern of
quantitative expression of SIX3 and SIX6 transcripts. As
with the whole body expression maps, the distributions of
SIX3 and SIX6 transcripts were overlapping but distinct
from each other. The highest expression of both SIX3 and
SIX6 was detected in the retina with Tcn values greater
than 100,000, considerably higher (greater than 5-fold)
than that seen anywhere else in the body (see Fig. 2 and
Table 2). A predominance of SIX3 transcripts was detected
in the conjunctiva, cornea, iris, lens, limbus, and orbital
adipose tissues while SIX6 transcripts predominated in the
choroid, ciliary body, optic nerve, optic nerve head, and
sclera (Fig. 2).
As SIX3 transcripts showed a high correlation (Pearson
correlation coefficient >0.75) with five other transcripts in
the whole body expression maps (Table 3), we also
examined the level of these transcripts in ocular tissues
and calculated correlation coefficients (Table 4). As
observed in the whole body maps, there was exceedingly
high correlation between the expression patterns of IDH1
and SIX3 and also high correlation between SIX3 and
CDH14. IDH1 and CDH14 also appear to correlate with
each other in the eye (Pearson correlation coefficient = 0.87;
Table 4). Fig. 2 shows the expression map overlay for SIX3
and IDH1 in ocular tissues. Similar target copy numbers
were detected for SIX3 and IDH1 in the choroid, iris, lens,
optic nerve, optic nerve head, and retina. In the cornea, only
Table 3
Genes showing high correlation (>0.75) of whole body expression with
SIX3
Gene abbreviation and name Accession numbers Pearson correlation
coefficient
IDH1, NADP+-dependent
isocitrate dehydrogenase
NM_005896 0.93
5-HT2C, serotonin receptor
type 2C
M81778 0.77
RGS8, regulator of G protein
signaling 8
NM_033345 0.76
CDH14, cadherin 18, type 2 U59325 0.76
5-HT5A, serotonin receptor
type 5A
X81411 0.76
IDH1 transcripts were detected while in the conjunctiva
only SIX3 transcripts were present.
Localization of SIX3 mRNA in human fetal and adult ocular
tissues by in situ hybridization
In the adult eye, SIX3 mRNA showed a restricted pattern
of expression. In the anterior segment, the corneal
endothelium (Figs. 3a–c) and the lens epithelium (Figs.
3d–f) stained positive for SIX3. Expression was absent
from the cortical or nuclear lens fibers. In the posterior
segment, SIX3 expression was found in the retina (Figs.
3g– i). The cytoplasm of the ganglion cells stained
intensely. A strong signal was detectable in the perinuclear
cytoplasm of the cells of the inner nuclear layer. It was not
possible to determine which of the cells present in this layer
(bipolar cells, amacrine cells, horizontal cells, and Muller
cells) were expressing SIX3; it appeared that all cells were
labeled. A relatively weaker signal was found in the outer
nuclear layer. However, a strong signal was present in the
inner segments of the photoreceptor cells (where most of
the cytoplasmic organelles of the photoreceptor cells are
found). The optic nerve was negative. Although the
presence of intrinsic pigmentation made assessment of
staining difficult, the retinal pigment epithelium (RPE)
appeared negative. At the ora serrata, staining ended
abruptly in most cases (Figs. 3j–l), although in some cases
there was staining of scattered isolated inner ciliary
epithelial cells of the pars plana for a short distance
anteriorly. Staining was not found in the pars plicata
epithelium or elsewhere in the uveal tract or sclera.
In situ hybridization produced broadly similar results in
fetal eyes of comparable ages. Expression was found in
retinal neuroblasts starting at 6 weeks gestation and then
consistently at later gestational ages (Figs. 4a–f). The optic
stalk/primordial nerve did not show expression at any stage.
In the cornea, the epithelium was positive across the entire
age range (Figs. 5a and b). The endothelium did not stain
until 6 weeks gestation, although expression was then
detectable at all subsequent stages (Figs. 5c and d). The lens
epithelium stained positive for SIX3 in eyes of all gestation
stages. Lens fibers were consistently negative (Figs. 5e–h).
Staining of other ocular tissues was less consistent. In nine
eyes, from week 7 upward, there was patchy staining of iris
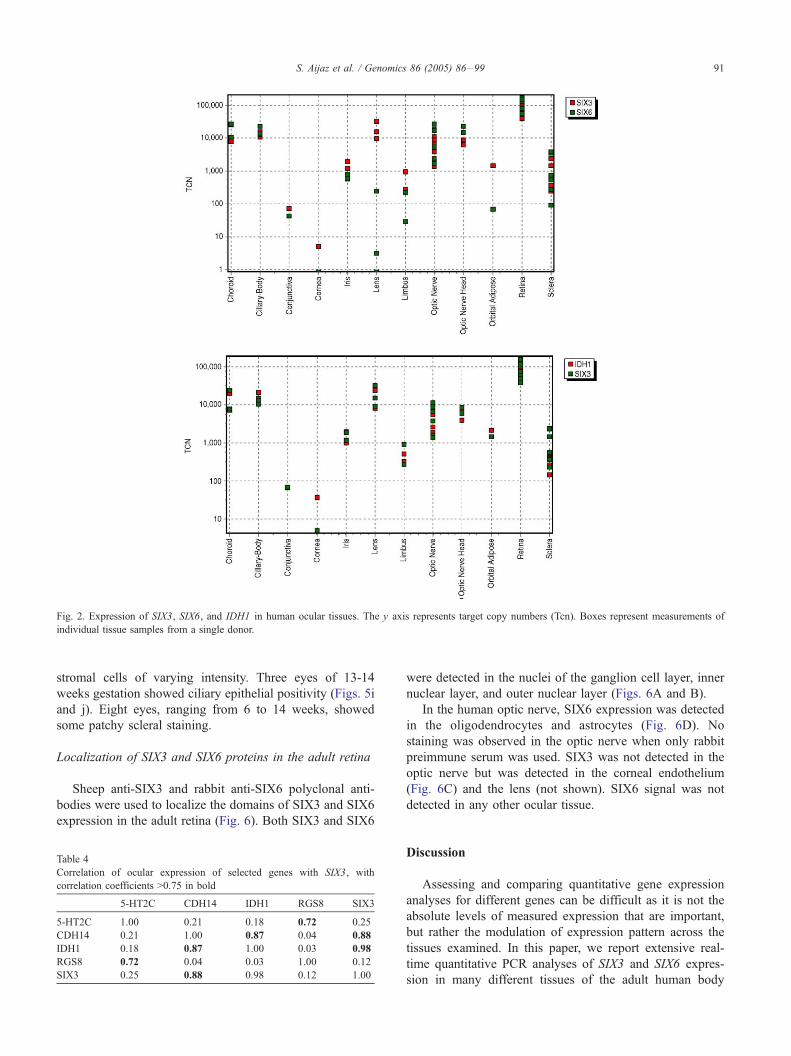
Fig. 2. Expression of SIX3, SIX6, and IDH1 in human ocular tissues. The y axis represents target copy numbers (Tcn). Boxes represent measurements of
individual tissue samples from a single donor.
S. Aijaz et al. / Genomics 86 (2005) 86–99 91
stromal cells of varying intensity. Three eyes of 13-14
weeks gestation showed ciliary epithelial positivity (Figs. 5i
and j). Eight eyes, ranging from 6 to 14 weeks, showed
some patchy scleral staining.
Localization of SIX3 and SIX6 proteins in the adult retina
Sheep anti-SIX3 and rabbit anti-SIX6 polyclonal anti-
bodies were used to localize the domains of SIX3 and SIX6
expression in the adult retina (Fig. 6). Both SIX3 and SIX6
Table 4
Correlation of ocular expression of selected genes with SIX3, with
correlation coefficients >0.75 in bold
5-HT2C CDH14 IDH1 RGS8 SIX3
5-HT2C 1.00 0.21 0.18 0.72 0.25
CDH14 0.21 1.00 0.87 0.04 0.88
IDH1 0.18 0.87 1.00 0.03 0.98
RGS8 0.72 0.04 0.03 1.00 0.12
SIX3 0.25 0.88 0.98 0.12 1.00
were detected in the nuclei of the ganglion cell layer, inner
nuclear layer, and outer nuclear layer (Figs. 6A and B).
In the human optic nerve, SIX6 expression was detected
in the oligodendrocytes and astrocytes (Fig. 6D). No
staining was observed in the optic nerve when only rabbit
preimmune serum was used. SIX3 was not detected in the
optic nerve but was detected in the corneal endothelium
(Fig. 6C) and the lens (not shown). SIX6 signal was not
detected in any other ocular tissue.
Discussion
Assessing and comparing quantitative gene expression
analyses for different genes can be difficult as it is not the
absolute levels of measured expression that are important,
but rather the modulation of expression pattern across the
tissues examined. In this paper, we report extensive real-
time quantitative PCR analyses of SIX3 and SIX6 expres-
sion in many different tissues of the adult human body

Fig. 3. Expression of SIX3 in adult human eye tissues demonstrated by RNA in situ hybridization. (a, d, g, j) Hematoxylin and eosin-stained sections for
orientation. (b, e, h, k) In situ hybridization using the antisense SIX3 riboprobe, with a positive signal indicated by blue/black staining. (c, f, i, l) Negative
controls by in situ hybridization using the sense SIX3 riboprobe. Expression is seen in the corneal endothelium (a–c, arrow in a), the lens epithelium (d– f,
arrow in d), and throughout the cellular layers of the retina (g– i, GC, ganglion cells; INL, inner nuclear layer; ONL, outer nuclear layer; PR, photoreceptors).
Retinal expression ends at the ora serrata (j – l, arrow in j indicates site of ora serrata). All scale bars = 50 Am.
Fig. 4. Expression of SIX3 in fetal human retinal tissues demonstrated by RNA in situ hybridization. (a, d) Hematoxylin and eosin-stained sections for
orientation. (b, e) In situ hybridization using the antisense SIX3 riboprobe, with a positive signal indicated by blue/black staining. (c, f) Negative controls by in
situ hybridization using the sense SIX3 riboprobe. a–c show expression in the retina at 9.5 weeks and d–f show retinal expression at 13 weeks (NBs,
neuroblasts; INBL, inner neuroblastic layer; ONBL, outer neuroblastic layer). All scale bars = 50 Am.
S. Aijaz et al. / Genomics 86 (2005) 86–9992
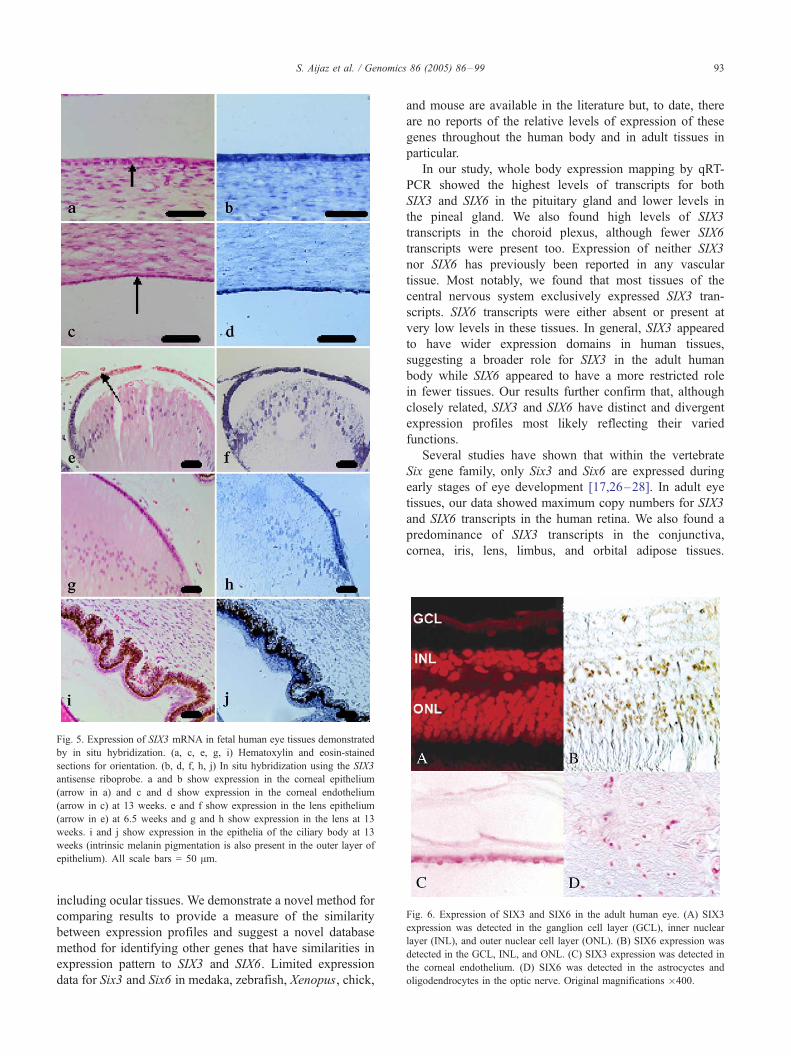
Fig. 5. Expression of SIX3 mRNA in fetal human eye tissues demonstrated
by in situ hybridization. (a, c, e, g, i) Hematoxylin and eosin-stained
sections for orientation. (b, d, f, h, j) In situ hybridization using the SIX3
antisense riboprobe. a and b show expression in the corneal epithelium
(arrow in a) and c and d show expression in the corneal endothelium
(arrow in c) at 13 weeks. e and f show expression in the lens epithelium
(arrow in e) at 6.5 weeks and g and h show expression in the lens at 13
weeks. i and j show expression in the epithelia of the ciliary body at 13
weeks (intrinsic melanin pigmentation is also present in the outer layer of
epithelium). All scale bars = 50 Am.
Fig. 6. Expression of SIX3 and SIX6 in the adult human eye. (A) SIX3
expression was detected in the ganglion cell layer (GCL), inner nuclear
layer (INL), and outer nuclear cell layer (ONL). (B) SIX6 expression was
detected in the GCL, INL, and ONL. (C) SIX3 expression was detected in
the corneal endothelium. (D) SIX6 was detected in the astrocyctes and
oligodendrocytes in the optic nerve. Original magnifications �400.
S. Aijaz et al. / Genomics 86 (2005) 86–99 93
including ocular tissues. We demonstrate a novel method for
comparing results to provide a measure of the similarity
between expression profiles and suggest a novel database
method for identifying other genes that have similarities in
expression pattern to SIX3 and SIX6. Limited expression
data for Six3 and Six6 in medaka, zebrafish, Xenopus, chick,
and mouse are available in the literature but, to date, there
are no reports of the relative levels of expression of these
genes throughout the human body and in adult tissues in
particular.
In our study, whole body expression mapping by qRT-
PCR showed the highest levels of transcripts for both
SIX3 and SIX6 in the pituitary gland and lower levels in
the pineal gland. We also found high levels of SIX3
transcripts in the choroid plexus, although fewer SIX6
transcripts were present too. Expression of neither SIX3
nor SIX6 has previously been reported in any vascular
tissue. Most notably, we found that most tissues of the
central nervous system exclusively expressed SIX3 tran-
scripts. SIX6 transcripts were either absent or present at
very low levels in these tissues. In general, SIX3 appeared
to have wider expression domains in human tissues,
suggesting a broader role for SIX3 in the adult human
body while SIX6 appeared to have a more restricted role
in fewer tissues. Our results further confirm that, although
closely related, SIX3 and SIX6 have distinct and divergent
expression profiles most likely reflecting their varied
functions.
Several studies have shown that within the vertebrate
Six gene family, only Six3 and Six6 are expressed during
early stages of eye development [17,26–28]. In adult eye
tissues, our data showed maximum copy numbers for SIX3
and SIX6 transcripts in the human retina. We also found a
predominance of SIX3 transcripts in the conjunctiva,
cornea, iris, lens, limbus, and orbital adipose tissues.

S. Aijaz et al. / Genomics 86 (2005) 86–9994
SIX6 expression predominated in the choroid, ciliary body,
optic nerve, optic nerve head, and sclera. Expression of
both Six3 and Six6 in the retina is extensively documented,
as is the exclusive expression of Six3 in the lens and Six6
in the optic stalk [17,23,26–28,50–54]. Expression in
other ocular tissues has not been previously reported. The
expression profile we observed for SIX3 in qRT-PCR
experiments was consistent with the localization of SIX3
mRNA in in situ hybridization experiments. As seen in
qRT-PCR maps, SIX3 mRNA signal was strongly detected
in the cornea, lens, and retina. In the fetal eyes, SIX3
mRNA was first detected in the retinal neuroblasts from 6
weeks of gestation, which is consistent with previous
reports [45].
To further characterize the expression domains of SIX3
and SIX6 at the protein level, we generated polyclonal
antibodies against specific C-terminal regions of human
SIX3 and SIX6 and stained sections of the adult human
retina with these antibodies. We observed similar expression
profiles for SIX3 and SIX6. Both proteins were localized in
the ganglion cell layer (GCL), inner nuclear layer (INL), and
the outer nuclear layer (ONL). For SIX3, this was in
contrast to the mRNA localization, since we did not observe
a very strong signal in the ONL (compared to the GC and
the INL) by in situ hybridization. This may reflect differ-
ences in the expression domains of SIX3 mRNA and
protein. Previous studies have also shown both human
SIX3 and SIX6 mRNA to be localized in the GCL and INL
but not the ONL [30,45]. Moreover, the continued
expression of SIX6 in the ocular tissues of the adult human
body, observed by us, is also in contrast to most previous
studies that have reported that Six6 is expressed early in eye
development and is then down-regulated [23,26,27,40,51].
These studies had suggested that Six6 is only required in
early eye development and are supported by a more recent
report that showed a proliferation defect in the retina of Six6
mutant mice which was only detectable in early stages of
retinal development [40]. Since Six6 mutant mice display
retinal hypoplasia concomitant with reduced BrdU labeling
and premature cell cycle exit, a role in cell proliferation has
been suggested for Six6 [40]. However, our data clearly
show the presence of very high copy numbers of SIX6
transcripts in the adult human retina, suggesting a later role
for SIX6 in the adult retina. A dual role has been suggested
for Six3, whereby it is required in the induction of the optic
vesicle and forebrain formation in early development and in
proximo-distal patterning of the retina during late develop-
ment [34,41,42]. A role in patterning has not been
demonstrated for Six6 so far but its presence in the adult
human retina, observed by us, suggests a similar role. Since
Six6 mutant mice exhibit reduced or absent optic nerves
[40], we also examined the expression of SIX6 protein in
the human optic nerve using anti-SIX6 antibodies. We
observed SIX6 expression only in the oligodendrocytes and
astrocytes of the human optic nerve, the significance of
which is unclear.
The whole body and ocular expression profile mapping
of SIX3 showed a high and unexpected correlation between
SIX3 transcripts and those of IDH1, the gene encoding the
NADP+-dependent isocitrate dehydrogenase, a key enzyme
in the Krebs cycle and we suggest that this gene be
investigated further for a functional link to SIX3. Interstitial
deletions of the long arm of chromosome 2 (where IDH1 is
localized) have been described which result in micro-
phthalmia, corneal anomalies ,and cleft palate [57]. More-
over, IDH1 is highly expressed in the bovine cornea and
was identified as a corneal epithelial crystallin involved in
maintaining corneal transparency [58]. This is in contrast to
our data where we observe low expression of IDH1
transcripts in the human cornea, suggesting that this protein
is unlikely to play this role in human cornea. Expression of
IDH1 isoforms has also been reported in cultured rat
neurons, astrocytes, oligodendrocytes, and microglial cells
[59] as well as the heart and the kidney [60]. These
observations are significant because we have detected high
levels of SIX3 transcripts in many nervous and renal tissues
of the human body, suggesting a link between the two.
Interestingly, in the rat kidney and heart, a small proportion
of IDH1 is reported to be expressed in the nuclei where it is
associated with heterochromatin [60], although it is unclear
as to how it is transported to the nucleus. The function of
IDH1 in these tissues is not well understood. It may be that
IDH1 transcription is modulated by SIX3 through a
mechanism similar to that reported in Drosophila imaginal
discs where homeotic genes were shown to regulate the
expression pattern of IDH1 especially in the eye where it
correlated with the differentiation of the ommatidia [61]. A
previous study has suggested that the cofactors that interact
with Six3 during late development (where it is involved in
patterning of the retina) may be different from those that
interact with it in early brain/eye formation, since the basic
helix loop helix proteins NeuroD, Ath5, Ath3, and Ash1
were shown to be coexpressed with Six3 only during late
retinal differentiation [62]. Therefore, one hypothesis
emerging from our observations could be that any SIX3/
IDH1 association only occurs in the adult tissue and this
needs to be examined in future studies.
Another novel correlation we have detected in our study
is between SIX3 and CDH14. The functional link between
SIX3 and CDH14 is not clear. CDH14 belongs to the type II
subclass of cadherins and its expression is restricted to the
central nervous system [63]. Since we have also observed
SIX3 expression in the CNS it is possible that there may be
an interaction between the two. Notably, high levels of
CDH14 expression have been reported in small-cell lung
carcinoma cell lines, which have neuroectodermal cell
phenotypes [64] and it was suggested that CDH14 has a
role in tumor suppression and/or regulation of proliferation.
Since SIX3 also plays a role in cell proliferation, it would be
interesting to speculate that the proliferative phenotype
observed upon overexpression of SIX3 might be mediated
through CDH14.

S. Aijaz et al. / Genomics 86 (2005) 86–99 95
In conclusion, while more work is needed to address the
postulated functional relevance of the coexpression of SIX3
with IDH1, CDH14, or the other genes showing highly
correlated expression profiles in adult human ocular and/or
other body tissues, correlation of gene expression studies
such as those described here are a powerful tool that will be
useful in identifying possible interactions between genes or
gene products.
Materials and methods
Human tissue acquisition and processing
All experiments involving the use of human tissue
conformed to the guidelines set forth in the Declaration of
Helsinki and we had prior approval from several appropriate
local ethics review boards for the collection and the use of
human tissues in this study. Briefly, adult human tissues
were collected from a variety of sources that included
hospitals and research tissue banks. Tissues were collected
from surgical specimens, postmortem examinations, or from
tissues deemed unsuitable for transplantation. In all cases,
informed consent was obtained from the donor or their next
of kin (in the case of deceased donors) for use of tissues in
biomedical research. Tissues were specifically dissected to
provide discrete anatomical structures and these were then
either snap-frozen in liquid nitrogen or fixed for processing
and embedding in paraffin to produce histological sections
by standard methods. The fetuses used in the study were
from normal pregnancies and had no detectable structural
abnormalities or other reasonable clinical indicators of
abnormality. They were collected for use in research, with
ethics committee approval, after medical terminations of
pregnancy and obtained by us from the hospital pathology
laboratory as surplus to diagnostic requirements.
For RT-PCR analysis, frozen tissue was used to extract
total RNA using TriZol reagent (Life Technologies, USA)
by methods adapted from the manufacturer’s protocol and
after a variety of methods of homogenization according to
tissue type. RNA was subjected to standard quality control
analyses prior to being deemed suitable for quantitative
transcript analysis. This quality control step involved the
assessment of RNA quantity, purity, and integrity using
spectrophotometry, gel electrophoresis, and assessment on a
Bioanalyzer (Agilent Technologies, USA). An additional
test of suitability for RT-PCR was conducted for each
sample by successfully detecting the presence of h-actinand human glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) transcripts at expected levels. This standard
approach is employed to produce consistency in all gene
expression mapping during the production of a database of
human gene expression maps by Pharmagene Laboratories
Ltd. Further details of these can be obtained from the
company website (http://www.pharmagene.com). All RNA
samples were diluted to a known standard working
concentration in nuclease-free water with the addition of
RNase inhibitor (Applied Biosystems, USA).
Quantitative RT-PCR analysis
Detection of specific mRNAs was carried out by qRT-
PCR using the ABI Prism 7700 Sequence Detection System
(Applied Biosystems, USA). Pairs of forward and reverse
primers and fluorogenic probes to target sequences (human
SIX3 sequence, Accession Number AF049339; human SIX6
sequence, Accession Number AF141651) were designed by
computer using Primer Express software (Applied Biosys-
tems) and are listed in Table 1. The primer/probe sets were
homology searched to ensure that they were specific using
an NCBI BLAST search (http://www.ncbi.nlm.nih.gov/).
Each primer/probe set was validated for size of amplicon
and for efficiency of amplification prior to being employed
in experimental RT-PCR analysis.
For each RT-PCR to detect a target sequence, the reaction
was duplexed with the detection of a 71-bp region of the
human GAPDH transcript that spans an exon/exon boun-
dary. This transcript is ubiquitously expressed and its
detection above a specific threshold level of 10,000 copies
was used as an internal positive control for each reaction
well within the assay. An initial enzyme activation step of
94-C for 12 min was followed by 40 cycles of 94-C for 15 s
(denaturing) and 60-C for 30 s (annealing and extending) of
PCR. SIX3 and SIX6 target probes were labeled with FAM
(6-carboxyfluorescein) (Applied Biosystems) while GAPDH
was labeled with VIC (Applied Biosystems) to allow for
target-specific fluorescent signal to be distinguished in the
duplexed reactions. Both fluors were quenched with
TAMRA (6-carboxytetramethylrhodamine). The transcript
quantification data were transferred to the Pharmagene data
visualization database, TargetEvaluator, where it was dis-
played as stellar and scatter plots. Further details of the RT-
PCR, transcript detection, and transcript quantification were
as previously described [56].
Expression measurements of SIX3 and SIX6 transcripts
in 72 anatomically discrete adult human tissues (compo-
site human whole body maps) were repeated in triplicate
for each gene. Different donors were used to provide
RNA for each tissue type for each measurement (i.e., 3
distinct composite human whole body maps were used for
the measurement of each gene transcript). Target copy
number was determined as the number of copies of each
gene per 100 ng of total RNA. This is derived from the
threshold fluorescence level by interpolation from the
global standard curve generated by Pharmagene Labora-
tories. Mean expression profiles were obtained by
calculating the mean target (gene) copy number from 3
measurements for each tissue and both the mean and the
individual measurements (not shown or reported in this
paper) for each tissue were plotted on stellar graphs for
ease of data visualization. Eye expression maps were
generated from data obtained from 12 anatomically

S. Aijaz et al. / Genomics 86 (2005) 86–9996
discrete eye tissues and plotted as individual tissue
observations.
Correlations were calculated for investigating the rela-
tionship between quantitative gene expression profiles. The
Pearson correlation coefficient is a statistical measure of the
similarity of two or more sets of numerical measurements to
each other. The closer the value of the coefficient to 1.0, the
greater the chance the two sets of measurements vary in
accordance with each other [65]. Pearson correlation
coefficients were calculated using the mean expression
levels of SIX3 and SIX6 genes at each anatomical site and
the corresponding data in the Pharmagene TargetEvaluator
database comprising whole body gene expression profiles of
1641 genes were searched for other genes showing high
correlation to either SIX3 or SIX6 where the correlation
coefficient was �0.75. A selection of specific genes thought
to correlate with either gene throughout the whole body was
then remapped in ocular tissues and correlation of expres-
sion within ocular tissues was assessed.
In situ hybridization of human eye tissues
Paraffin blocks of 10 adult human eyes, which had been
received routinely for pathological examination and consid-
ered histologically normal, were selected. These eyes had
been fixed in 3% glutaraldehyde in cacodylate buffer. Each
paraffin block contained a standard pupil-optic nerve (PO)
tissue block. An additional series of paraffin blocks of 28
normal human fetal eyes, ranging from 4.5 to 14 weeks
gestation, were also studied. The gestation and age of each
had been previously estimated by measurement of crown-
rump length (CRL) and foot length (FL). All paraffin
sections were cut at 5 Am and mounted on glass slides
coated with aminopropyltriethoxysilane (APES) (Sigma).
To generate the SIX3 probe, a fragment containing 280
bp of the 5UTR and the first 460 bp of the coding region of
the human SIX3 gene was subcloned into Bluescript II SK
vector (Stratagene). Sense and antisense SIX3 probes were
prepared for nonisotopic in situ hybridization by tran-
scription in the presence of digoxigenin-11-UTP (Roche
Biochemicals). Sections were dewaxed in xylene and
hydrated in phosphate-buffered saline/diethyl pyrocarbonate
(PBS/DEPC). A prehybridization treatment was carried out
with 0.2 N HCl, 0.3% Triton X (Sigma), and proteinase K
(Promega). The tissues were then further fixed in 2%
paraformaldehyde for 5 min. Prior to hybridization, the
labeled riboprobes were heated to 95-C and quenched on
ice. The slides were heated to 70-C and overnight hybrid-
ization was carried out at this temperature. After hybrid-
ization, the sections were washed and hybridization was
detected immunohistochemically using alkaline phospha-
tase-conjugated anti-digoxigenin antiserum and visualized
using the nitroblue tetrazolium/bromochloroindolyl phos-
phate (NBT/BCIP) substrate. Sections were not counter-
stained. They were examined with a BX50 light microscope
(Olympus) using bright-field and differential interference
contrast (Nomarski) optics and photographed with a DP10
digital camera (Olympus). Comparable attempts to localize
SIX6 transcripts on paraffin embedded sections by in situ
hybridization were unsuccessful despite numerous attempts
(perhaps due to the relative instability of the SIX6 mRNA).
The stability of the mRNA is better preserved in frozen
cryostat sections. However, due to limited availability of
high-quality human tissues suitable for the production of
well-orientated frozen sections of the retina (which the
banked frozen tissues used for qRT-PCR were not), we were
unable to gain access to cryosections.
Antibody generation
The SIX3 antibody was raised against recombinant
protein derived from the last 68 amino acids (aa 265-332)
of the human SIX3 sequence (Accession Number
AF049339). The DNA fragment of the SIX3 gene corre-
sponding to these amino acids was cloned with a GST-tag
using the Univector plasmid system [66]. The GST-tagged
SIX3 protein was purified using glutathione Sepharose
beads (Pharmacia). Sheep polyclonal antiserum (anti-SIX3)
was obtained by subcutaneous injections of purified
recombinant protein every 4 weeks (Diagnostics Scotland,
UK). A similar approach using recombinant protein derived
from the last 60 amino acids (aa 187-246) of the human
SIX6 sequence (Accession Number AF141651) did not
yield antibodies of high specificity. Therefore, an alternative
approach was taken to generate antibodies against SIX6.
The SIX6 antibody was raised against a C-terminal peptide
corresponding to amino acids 198-215 (SQGSGRAL-
RAEGDGTPEV) derived from the published human SIX6
sequence. This peptide was conjugated to keyhole limpet
hemocyanin (KLH) through an artificial cysteine placed at
the N-terminal (Genosys Biotechnologies, UK). Rabbit
polyclonal antiserum (anti-SIX6) was obtained by subcuta-
neous injection of the conjugated peptide into two rabbits
every 4 weeks (Genosys Biotechnologies). The specificity
of all antibodies was confirmed by Western blots using
recombinant SIX3 protein (for anti-SIX3) and either
recombinant SIX6 protein or the synthetic SIX6 peptide
(for anti-SIX6) as well as total protein extracts from the
human retina. As a negative control, GST alone was used in
the same blotting experiments.
Immunohistochemistry of human eye tissues
SIX3 staining
A piece of unaffected retina was excised from an eye
with malignant melanoma that was submitted for patho-
logical examination. The retina was fixed in 10% neutral-
buffered formalin and embedded in 5% agarose. The block
was mounted on a vibratome and 100-Am-thick sections
were cut. Free-floating sections were blocked with 5%
nonfat milk overnight at 4-C. The sections were then briefly
washed with PBS and incubated with the anti-SIX3

S. Aijaz et al. / Genomics 86 (2005) 86–99 97
antibody (1:500 dilution) at 4-C overnight. Subsequently,
the sections were washed with PBS and incubated with the
anti-sheep antibody conjugated to Cy3 (1:500; Jackson
ImmunoResearch Laboratories, USA) for 6 h at room
temperature (RT). After a brief rinse, the sections were
mounted on glass slides using fluorescent mounting medium
(Dako, UK). To verify the specificity of the immunostain-
ing, sections were also stained with sheep preimmune
serum. The mounted sections were visualized and photo-
graphed on a confocal microscope (Zeiss LSM 510). In
addition, pupil-optic nerve block sections of a 10% neutral-
buffered formalin-fixed, paraffin embedded, normal human
donor eye were cut at 6 Am and mounted on precoated
(APES) glass slides. Immunohistochemical staining using
anti-SIX3 antibody (1:1000) was conducted using the
methods described for SIX6 below.
SIX6 staining
A separate eye from a 68-year-old donor was obtained
from an Eye bank. This eye was fixed in 10% neutral-
buffered formalin and processed for embedding in paraffin
wax by standard methods. Sections were cut at 6 Am and
mounted on APES-coated glass slides. The sections were
dewaxed in xylene and rehydrated with 100, 90, and 70%
industrial methylated spirit. The sections were then pre-
treated with 0.1% trypsin in O.2 M Tris, pH 7.8, for 10 min.
After washing with PBS, the endogenous peroxidase
activity was blocked by incubating the sections with 0.5%
hydrogen peroxide in 50% methanol for 30 min at RT. The
sections were then blocked with 10% normal goat serum
(Dako) in PBS for 20 min at RT, followed by incubation
with anti-SIX6 antibody (1:1000 dilution) in PBS contain-
ing 10% normal goat serum overnight at 4-C. Subsequently,the sections were rinsed with PBS and incubated with goat
anti-rabbit antibody conjugated to horseradish peroxidase
(1:500; Jackson ImmunoResearch Laboratories) for 45 min
at RT. The immunoreaction was visualized by incubating
with either a red substrate (Vector Laboratories Inc., USA)
or a solution containing 3,3-diaminobenzidine (DAB)
(Sigma) and 0.03% hydrogen peroxide in PBS. Subse-
quently, the sections were dehydrated in an ascending series
of alcohols, equilibrated in xylene, and mounted in
dibutylthalate xylene (DPX) mounting medium (Merck,
UK). Some sections were also stained with rabbit preim-
mune serum to detect nonspecific staining. The stained
sections were visualized with a BX50 light microscope
(Olympus) using bright-field and differential interference
contrast (Nomarski) optics and photographed with a DP10
digital camera (Olympus).
Acknowledgments
We are grateful to Mike Cheetham for his help, support,
and encouragement. We are also grateful to Peter Munro for
help with confocal microscopy. We thank Dee Hornell,
Rosalind Hart, and Robert Alexander for assistance in
immunohistochemical techniques. This research was sup-
ported by a grant from the Birth Defects Foundation, United
Kingdom. I. H. holds a Career Development Award from the
Medical Research Council, United Kingdom.
References
[1] I.M. Hanson, Mammalian homologues of the Drosophila eye
specification genes, Semin. Cell Dev. Biol. 12 (2001) 475–484.
[2] J.P. Kumar, Signalling pathways in Drosophila and vertebrate retinal
development, Nature Rev. Genet. 2 (2001) 846–857.
[3] S. Wawersik, R.L. Maas, Vertebrate eye development as modeled in
Drosophila, Hum. Mol. Genet. 9 (2000) 917–925.
[4] G. Halder, P. Callaerts, W.J. Gehring, Induction of ectopic eyes by
targeted expression of the eyeless gene in Drosophila, Science 267
(1995) 1788–1792.
[5] G. Halder, P. Callaerts, S. Flister, U. Walldorf, U. Kloter, W.J.
Gehring, Eyeless initiates the expression of both sine oculis and eyes
absent during Drosophila compound eye development, Development
125 (1998) 2181–2191.
[6] N.M. Bonini, W.M. Leiserson, S. Benzer, The eyes absent gene:
genetic control of cell survival and differentiation in the developing
Drosophila eye, Cell 72 (1993) 379–395.
[7] B.N.R. Cheyette, P.J. Green, K. Martin, H. Garren, V. Hartenstein,
S.L. Zipurski, The Drosophila sine oculis locus encodes a homeo-
domain-containing protein required for the development of the entire
visual system, Neuron 12 (1994) 977–996.
[8] M.A. Serikaku, J.E. O’Tousa, Sine oculis is a homeobox gene required
for Drosophila visual system development, Genetics 138 (1994)
1137–1150.
[9] G. Mardon, N.M. Solomon, G.M. Rubin, Dachshund encodes a
nuclear protein required for normal eye and leg development in
Drosophila, Development 120 (1994) 3473–3486.
[10] R. Chen, M. Amoui, Z. Zhang, G. Mardon, Dachshund and eyes
absent proteins form a complex and function synergistically to
induce ectopic eye development in Drosophila, Cell 91 (1997)
893–903.
[11] W. Shen, G. Mardon, Ectopic eye development in Drosophila induced
by directed dachshund expression, Development 124 (1997) 45–52.
[12] R. Quiring, U. Walldorf, U. Kloter, W.J. Gehring, Homology of the
eyeless gene of Drosophila to the Small eye gene in mice and Aniridia
in humans, Science 265 (1994) 785–789.
[13] R.E. Hill, et al., Mouse small eye results from mutations in a paired-
like homeobox-containing gene, Nature 354 (1991) 522–525.
[14] K.L. Hammond, I.M. Hanson, A.G. Brown, L.A. Lettice, R.E. Hill,
Mammalian and Drosophila dachshund genes are related to the Ski
proto-oncogene and are expressed in eye and limb, Mech. Dev. 74
(1998) 121–131.
[15] P.-X. Xu, I. Woo, H. Her, D.R. Beier, R.L. Maas, Mouse Eya
homologues of the Drosophila eyes absent gene require Pax6 for
expression in lens and nasal placode, Development 124 (1997)
219–231.
[16] G. Borsani, et al., EYA4, a novel vertebrate gene related to Drosophila
eyes absent, Hum. Mol. Genet. 8 (1999) 11–23.
[17] G. Oliver, A. Mailhos, R.Wehr, N.G. Copeland, N.A. Jenkins, P. Gruss,
Six3, a murine homologue of the sine oculis gene, demarcates the most
anterior border of the developing neural plate and is expressed during
eye development, Development 121 (1995) 4045–4055.
[18] C.A. Boucher, et al., A novel homeodomain-encoding gene is
associated with a large CpG island interrupted by the myotonic
dystrophy unstable (CTG)n repeat, Hum. Mol. Genet. 4 (1995)
1919–1925.
[19] S.K. Heath, S. Carne, C. Hoyle, K.J. Johnson, D.J. Wells, Character-

S. Aijaz et al. / Genomics 86 (2005) 86–9998
ization of expression of mDMAHP, a homeodomain-encoding gene at
the murine DM locus, Hum. Mol. Genet. 6 (1997) 651–657.
[20] K. Kawakami, H. Ohto, K. Ikeda, R.G. Roeder, Structure, function
and expression of a murine homeobox protein AREC3, a homologue
of Drosophila sine oculis gene product, and implication in develop-
ment, Nucleic Acid Res. 24 (1996) 303–310.
[21] K. Kawakami, H. Ohto, T. Takizawa, T. Saito, Identification and
expression of Six family genes in mouse retina, FEBS Lett. 393 (1996)
259–263.
[22] G. Oliver, et al., Homeobox genes and connective tissue patterning,
Development 121 (1995) 693–705.
[23] J. Toy, J.-M. Yang, G.S. Leppert, O.H. Sundin, The Optx2 homeobox
gene is expressed in early precursors of the eye and activates retina-
specific genes, Proc. Natl. Acad. Sci. USA 95 (1998) 10643–10648.
[24] S. Rodriguez de Cordoba, M.E. Gallardo, J. Lopez-Rios, P. Bovolenta,
The human SIX family of homeobox genes, Curr. Genomics 2 (2001)
231–242.
[25] K. Kawakami, S. Sato, H. Ozaki, K. Ikeda, Six family genes-structure
and function as transcription factors and their roles in development,
Bioassays 22 (2000) 616–626.
[26] D. Jean, G. Bernier, P. Gruss, Six6 (Optx2) is a novel murine Six3-
related homeobox gene that demarcates the presumptive pituitary/
hypothalamic axis and the ventral optic stalk, Mech. Dev. 84 (1999)
31–40.
[27] J. Lopez-Rios, M.E. Gallardo, S. Rodriguez de Cordoba, P. Bovolenta,
Six9 (Optx2), a new member of the Six gene family of transcription
factors, is expressed at early stages of vertebrate ocular and pituitary
development, Mech. Dev. 83 (1999) 155–159.
[28] J. Toy, O.H. Sundin, Expression of the optx2 homeobox gene during
mouse development, Mech. Dev. 83 (1999) 183–186.
[29] H.C. Seo, J. Curtiss, M. Mlodzik, A. Fjose, Six class homeobox genes
in Drosophila belong to three distinct families and are involved in
head development, Mech. Dev. 83 (1999) 127–139.
[30] M.E. Gallardo, et al., Genomic cloning and characterization of the
human homeobox gene SIX6 reveals a cluster of SIX genes in
chromosome 14 and associates SIX6 hemizygosity with bilateral
anophthalmia and pituitary anomalies, Genomics 61 (1999) 82–91.
[31] F. Pignoni, B. Hu, K.H. Zavitz, J. Xiao, P.A. Garrity, S.L. Zipursky,
The eye-specification proteins So and Eya form a complex and
regulate multiple steps in Drosophila eye development, Cell 91 (1997)
881–891.
[32] H. Ohto, et al., Cooperation of six and eya in activation of their target
genes through nuclear translocation of Eya, Mol. Cell. Biol. 19 (1999)
6815–6824.
[33] M. Seimiya, W.J. Gehring, The Drosophila homeobox gene optix is
capable of inducing ectopic eyes by an eyeless-independent mecha-
nism, Development 127 (2000) 1879–1886.
[34] F. Loosli, S. Winkler, J. Wittbrodt, Six3 overexpression initiates the
formation of ectopic retina, Genes Dev. 13 (1999) 649–654.
[35] O. Lagutin, C.C. Zhu, Y. Furuta, D.H. Rowitch, A.P. McMahon, G.
Oliver, Six3 promotes the formation of ectopic vesicle-like structures
in mouse embryos, Dev. Dyn. 221 (2001) 342–349.
[36] M. Kobayashi, K. Nishikawa, T. Suzuki, M. Yamamoto, The homeo-
box protein Six3 interacts with the Groucho corepressor and acts as a
transcriptional repressor in eye and forebrain formation, Dev. Biol.
232 (2001) 315–326.
[37] C.C. Zhu, M.A. Dyer, M. Uchikawa, H. Kondoh, O.V. Lagutin, G.
Oliver, Six3-mediated auto repression and eye development requires
its interaction with members of the Groucho-related family of co-
repressors, Development 129 (2002) 2835–2849.
[38] J. Lopez-Rıos, K. Tessmar, F. Loosli, J. Wittbrodt, P. Bovolenta, Six3
and Six6 activity is modulated by members of the groucho family,
Development 130 (2003) 185–195.
[39] F.D. Bene, K. Tessmar-Raible, J. Wittbrodt, Direct interaction
of geminin and Six3 in eye development, Nature 427 (2004)
745–749.
[40] X. Li, V. Perissi, F. Liu, D.W. Rose, M.G. Rosenfeld, Tissue-specific
regulation of retinal and pituitary precursor cell proliferation, Science
297 (2002) 1180–1183.
[41] M. Kobayashi, R. Toyama, H. Takeda, I.B. Dawid, K. Kawakami,
Overexpression of the forebrain-specific homeobox gene six3 induces
rostral forebrain enlargement in zebrafish, Development 125 (1998)
2973–2982.
[42] M. Carl, F. Loosli, J. Wittbrodt, Six3 inactivation reveals its essential
role for the formation and patterning of the vertebrate eye, Develop-
ment 129 (2002) 4057–4063.
[43] G. Bernier, F. Panitz, X. Zhou, T. Hollemann, P. Gruss, T. Pieler,
Expanded retina territory by midbrain transformation upon over-
expression of Six6 (Optx2) in Xenopus embryos, Mech. Dev. 93
(2000) 59–69.
[44] M.E. Zuber, M. Perron, A. Philpott, A. Bang, W.A. Harris, Giant eyes
in Xenopus laevis by overexpression of XOptx2, Cell 98 (1999)
341–352.
[45] B. Granadino, et al., Genomic cloning, structure, expression pattern,
and chromosomal location of the human SIX3 gene, Genomics 55
(1999) 100–105.
[46] L. Pasquier, et al., A new mutation in the six-domain of SIX3 gene
causes holoprosencephaly, Eur. J. Hum. Genet. 8 (2000) 797–800.
[47] D.E. Wallis, et al., Mutations in the homeodomain of the human SIX3
gene cause holoprosencephaly, Nature Genet. 22 (1999) 196–198.
[48] M.E. Gallardo, S. Rodriguez De Cordoba, A.S. Schneider, M.A.
Dwyer, C. Ayuso, P. Bovolenta, Analysis of the developmental SIX6
homeobox gene in patients with anophthalmia/microphthalmia, Am. J.
Med. Genet. 129A (1) (2004) 92–94.
[49] S. Aijaz, et al., Absence of SIX6 mutations in microphthalmia,
anophthalmia and coloboma, Invest. Ophthalmol. Vis. Sci. 45 (2004)
3871–3876.
[50] F. Loosli, R.W. Koster, M. Carl, A. Krone, J. Wittbrodt, Six3, a
medaka homologue of the Drosophila homeobox gene sine oculis is
expressed in the anterior embryonic shield and the developing eye,
Mech. Dev. 74 (1998) 159–164.
[51] H.C. Seo, O. Drivenes, S. Ellingsen, A. Fjose, Expression of two
zebrafish homologues of the murine Six3 gene demarcates the initial
eye primordia, Mech. Dev. 73 (1998) 45–57.
[52] X. Zhou, T. Hollemann, T. Pieler, P. Gruss, Cloning and expression of
xSix3, the Xenopus homologue of murine Six3, Mech. Dev. 91 (2000)
327–330.
[53] P. Bovolenta, A. Mallamaci, L. Puelles, E. Boncinelli, Expression
pattern of cSix3, a member of the six/sine oculis family of tran-
scription factors, Mech. Dev. 70 (1998) 201–203.
[54] H. Ghanbari, H.C. Seo, A. Fjose, A.W. Brandli, Molecular cloning and
embryonic expression of Xenopus six homeobox genes, Mech. Dev.
101 (2001) 271–277.
[55] W.P. Bowen, et al., Measurement of cytochrome P450 gene induction
in human hepatocytes using quantitative real-time reverse transcriptase-
polymerase chain reaction, Drug Metab. Dispos. 28 (2000) 781–787.
[56] D.J. Collison, R.A. Coleman, R.S. James, J. Carey, G. Duncan,
Characterization of muscarinic receptors in human lens cells by
pharmacologic and molecular techniques, Invest. Ophthalmol. Vis.
Sci. 41 (2000) 2633–2641.
[57] I.A. Glass, C.A. Swindlehurst, D.A. Aitken, W. McCrea, E. Boyd,
Interstitial deletion of the long arm of chromosome 2 with normal
levels of isocitrate dehydrogenase, J. Med. Genet. 26 (1989) 127–130.
[58] L. Sun, T.-T. Sun, R.M. Lavker, Identification of a cytosolic NADP+-
dependent isocitrate dehydrogenase that is preferentially expressed in
bovine corneal epithelium, J. Biol. Chem. 274 (1999) 17334–17341.
[59] T. Minich, S. Yokota, R. Dringen, Cytosolic and mitochondrial
isoforms of NADP+-dependent isocitrate dehydrogenases are
expressed in cultured rat neurons, astrocytes, oligodendrocytes, and
microglial cells, J. Neurochem. 86 (2003) 605–614.
[60] C.M. Haraguchi, T. Mabuchi, S. Yokota, Localization of a mitochon-
drial type of NADP-dependent isocitrate dehydrogenase in kidney
and heart of rat: an immunocytochemical and biochemical study,
J. Histochem. Cytochem. 51 (2003) 215–226.

S. Aijaz et al. / Genomics 86 (2005) 86–99 99
[61] D.T. Kuhn, G.N. Cinningham, Isocitrate dehydrogenase in D.
melanogaster imaginal discs: pattern development and alteration by
homeotic mutant genes, Dev. Genet. 7 (1986) 21–34.
[62] K. Tessmar, F. Loosli, J. Wittbrodt, A screen for co-factors of Six3,
Mech. Dev. 117 (2002) 103–113.
[63] T. Shibata, Y. Shimoyama, M. Gotoh, S. Hirohashi, Identification of
human cadherin-14, a novel neurally specific type II cadherin, by
protein interaction cloning, J. Biol. Chem. 272 (1997) 5236–5240.
[64] Y. Shimosato, et al., Biological, pathological and clinical features of
small cell lung cancer, Cancer Lett. 33 (1986) 241–258.
[65] D.G. Altman, Practical Statistics for Medical Research, Chapman
Hall/ CRC, London, 1990.
[66] Q. Liu, M.Z. Li, D. Leibham, D. Cortez, S.J. Elledge, The
univector plasmid-fusion system, a method for rapid construction
of recombinant DNAwithout restriction enzymes, Curr. Biol. 8 (1998)
1300–1309.





























