Embed Size (px)
Citation preview

BIO
PHYS
ICS
AN
DCO
MPU
TATI
ON
AL
BIO
LOG
Y
Experimental evaluation of the generalized vibrationaltheory of G protein-coupled receptor activationRoss D. Hoehna, David E. Nicholsb,1, John D. McCorvyb, Hartmut Nevenc, and Sabre Kaisa,d,1
aDepartment of Chemistry, Purdue University, West Lafayette, IN 47907; bDepartment of Pharmacology, University of North Carolina, Chapel Hill, NC 27514;cGoogle Los Angeles, Venice, CA 90291; and dSanta Fe Institute, Santa Fe, NM 87501
Edited by Leslie B. Vosshall, The Rockefeller University, New York, NY, and approved April 14, 2017 (received for review November 6, 2016)
Recently, an alternative theory concerning the method by whicholfactory proteins are activated has garnered attention. This the-ory proposes that the activation of olfactory G protein-coupledreceptors occurs by an inelastic electron tunneling mechanismthat is mediated through the presence of an agonist with anappropriate vibrational state to accept the inelastic portion ofthe tunneling electron’s energy. In a recent series of papers, somesuggestive theoretical evidence has been offered that this the-ory may be applied to nonolfactory G protein-coupled recep-tors (GPCRs), including those associated with the central ner-vous system (CNS). [Chee HK, June OS (2013) Genomics Inform11(4):282–288; Chee HK, et al. (2015) FEBS Lett 589(4):548–552;Oh SJ (2012) Genomics Inform 10(2):128–132]. Herein, we test theviability of this idea, both by receptor affinity and receptor acti-vation measured by calcium flux. This test was performed usinga pair of well-characterized agonists for members of the 5-HT2class of serotonin receptors, 2,5-dimethoxy-4-iodoamphetamine(DOI) and N,N-dimethyllysergamide (DAM-57), and their respec-tive deuterated isotopologues. No evidence was found that selec-tive deuteration affected either the binding affinity or the acti-vation by the selected ligands for the examined members of the5-HT2 receptor class.
protein | mechanism of action | pharmacology | quantum biology |electron tunneling
O lfaction—and other chemo-sensitive sensory processes—isan important information-gathering technique for organ-
isms of many clades and kingdoms. Specifically, human olfactionis known to occur by activation of olfactory receptors (ORs)—a subclass of G protein-coupled receptors (GPCRs)—locatedwithin the nasal epithelium and mediating responses within theolfactory bulb, where it is encoded and conveys information tothe amygdala, the orbitofrontal cortex, and the hippocampus(1, 2). The discovery and the cloning of ORs led to the joint2004 Nobel Prize in Physiology and Medicine to Richard Axeland Linda Buck. (3) GPCRs are 7-helical transmembrane pro-teins, facilitating communication from extracellular ligand sig-nals to the cellular interior through activation of (interior) Gproteins, while maintaining the integrity of the membrane (4).GPCRs are activated by an appropriate agonist moving into theprotein’s orthosteric binding site, resulting in a conformationalchange within the helical bundle. This structural change leads toaltered conformations of the intracellular loops that couple toappropriate signaling molecules within the cell, e.g., G proteins.A recent series of papers has experimentally determined acti-vated/inactivated states through isotopic-tagged receptors withNMR spectroscopy (5–7). An additional work used moleculardynamics to provide structural insights into how the agonist mayassist the interchange between conformations through severalproposed peptide sidechain pathways by examining the struc-tures of the activated and inactivated µ-opioid receptor (8).Additionally, photon-induced conformational changes in light-sensitive proteins have also been observed (9, 10).
Recently, an iteration of the vibrational theory of olfaction(VTO)—suggested and advocated by Luca Turin (11, 12)—
has arisen and has gained both supporters (13–16) and detrac-tors (17, 18); the novelty of this incarnation of the VTO isascribable to its nonthermal- and nonphoton-based mechanism.Turin’s theory is a contemporary reincarnation of the more clas-sical theory proposed by Dyson (19), Wright (20), and Wrightand Serenius (21), where the activation of the olfactory recep-tor is performed—or sensitive to—the molecular vibrations ofthe olfactant. Dyson suggested that the molecular vibrationsof the agonist were exactly responsible for the activation ofthe protein. These vibrations were entirely thermally excited,as no mechanism for photoexcitation is available within thebody. The modern incarnation of this theory suggests that theOR behaves as an electron-tunneling (ET) junction. Hypothet-ically, this electron transfer instigates the conformation changeof the olfactory GPCR, leading to the intracellular signal cas-cade. This theory both is contentious and lacks direct at-receptorevidence. Several of the proposed tests for this theory, includ-ing odor mixing, have been addressed to disprove this theorythrough controlled sensory tests (13, 15). Additionally, moremodern examinations of this theory with both humans andinsect subjects have provided mixed and controversial results.(14, 16–18, 22–24).
There have been several tests of the previous iterations of theVTO, including odor blending (25, 26) and isotope exchange (20,27–30); each of these methods—odor blending (11, 18) and iso-tope exchange (11, 31)—has been used to evaluate the modernVTO. Initial justifications of the VTO correlated characteristicsof the vibrational spectra with olfactory perception (12), contin-ued within ORs (32), and expanded into non-OR GPCRs (33–36). Isotope exchange—which provides the ability to alter thevibrations of a molecule while maintaining its chemistry—maymanifest itself as an alteration of either the intensity or the qual-ity of the scent. Isotope exchange tests of the VTO focused on1H→2H exchange, although 13C exchange has been suggested
Significance
Herein, we test the present iteration of the vibrational theoryof protein activation by comparing predictions obtained fromTurin’s vibrational theory for the activation of olfactory recep-tors measuring affinity and activation at a nonolfactory recep-tor family of G protein-coupled receptors. This was done at theCNS serotonin receptor family h5-HT2 and with both the 2,5-dimethoxy-4-iodoamphetamine and N,N-dimethyllysergamideagonists. Invalidation was performed through a comparativeanalysis of agonist behavior between isotopologues.
Author contributions: R.D.H., D.E.N., H.N., and S.K. designed research; R.D.H., D.E.N.,and J.D.M. performed research; R.D.H., D.E.N., J.D.M., H.N., and S.K. analyzed data; andR.D.H., D.E.N., and S.K. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1To whom correspondence may be addressed. Email: [email protected] or [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1618422114/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1618422114 PNAS | May 30, 2017 | vol. 114 | no. 22 | 5595–5600

N
NH
H
CH3
H
O
N
aCH3bCH3
I
OCH3
OCH3
CH3
H17H16
H18NH2
A B
Fig. 1. (A) Structure of the (R)-DOI molecule. In blue text, the atomicindexes for some specific sidechain hydrogen atoms initially consideredfor deuteration are shown. Additional sites considered for deuteration arethe hydrogens of the two methoxy groups. (B) Structure of the DAM-57 molecule. In blue text, each methyl amide carbon where deuterationexchange was undertaken is denoted as either carbon “a” or “b.”
(37). Unfortunately, measuring either quality or intensitychanges by sensory studies has presented itself as inconclusiveand difficult to quantify.
Recently, several recent studies—both in vitro and in vivo—have been conducted at a more physiological level. Block et al.(17) studied the activation of several ORs (importantly,OR5AN1 and MOR244-3) by both muscone and cyclopen-tadecanone; neither receptor displayed a significant differen-tial response to isotopologues of the musk odorants, suggest-ing failure of the VTO at mammalian ORs. However, criticismof this work has arisen on grounds of lacking the in situ envi-ronment (odorant binding proteins, cofactors, etc.) and possiblybeing too specific with respect to the repertoire of examined ORs(23, 24, 38, 39). Supporting previous behavioral studies (14, 16),Drimyl et al. (23) and Paoli et al. (24) studied direct electro-physical responses induced by odorant detection at the antennaeand glomeral lobe of several Drosophila species and Apis mel-lifera, respectively. These findings show unique spatial–temporalresponses with respect to families of isotopologues. Two thingsshould be further noted with respect to these in situ insect stud-ies: (i) Despite attempts at compensation, these results may notbe entirely independent of perireceptor effects (including trans-port, enzymes, and extracellular vestibule). (ii) Insect ORs areionotropic receptors (IRs) and not GPCRs; each class of recep-tors (GPCRs vs. IRs) shows specific evolutionary benefits (broadresponsiveness vs. speed of detection and processing) (40, 41).
Herein, we exploit the fact that ORs are a subclass of thebroader GPCR family with highly conserved sequences andstructural motifs. The nonexceptionalism of ORs within thebroader class of GPCRs was previously discussed within the con-text of the VTO (18) and is highlighted by Barwich (42), whoasserts ORs are a model system for the GPCRs within neuro-biology. Furthermore, ORs maintain up to 40% genetic similar-ity with rhodopsin (43). Additionally, ORs appear within areasof mammalian corpus that have no olfactory capacity (44–55).We therefore hypothesize that due to functional and morpholog-ical similarities, if ORs are activated through an ET mechanism,other GPCRs share the same fundamental mechanism. Examina-tion of another (better characterized) GPCR subclass may pro-vide insight into the possibility of the proposed ET mechanism.The serotonin 5-HT2 receptor class, notably a primary targetfor hallucinogenic compounds, was selected as the main test forthe contemporary VTO. (R)-2,5-dimethoxy-4-iodoamphetamine(R-DOI) is a well-characterized agonist for the serotonin 5-HT2A
receptor used as a standard ligand for studying the pharmacol-ogy of 5-HT2A receptors and is widely used as a radioligandto measure expression and affinity of ligands at 5-HT2A recep-tors, particularly in brain tissues. (R)-DOI is reported to havepsychedelic effects somewhat similar to those of lysergic aciddiethylamide (LSD) (56). A quintessential hallucinogenic ligandfor the 5-HT2A receptor is LSD; a highly related (non-Schedule1) molecule is generated by substituting the diethyl amide with adimethyl amide, creating N,N-dimethyllysergamide (DAM-57).DAM-57 is an assumed serotonergic psychedelic with limitedhallucinogenic capacity and very mild autonomic stimulation inhumans at dosages of ∼100µg (57).
The organization of this paper is as follows: Theoretical Pre-dictions gives a brief explanation of the theoretical predictionsfor (R)-DOI at members of the 5-HT2 serotonin receptor classaccording to Turin’s theory, while making direct reference toa previous work concerning similar predictions for DAM-57;Experimental Procedures briefly introduces methods of experi-mental analysis for determining affinity; Results and Discussionprovides a discussion of the experimental results and a compar-ison with theoretical predictions; and finally, in Conclusion weprovide concluding remarks.
Theoretical PredictionsTurin—within the contemporary VTO—hypothesized that theactive site of the GPCR [specifically an OR, although later worksconsidered generalizing this hypothesis (18, 36, 58)] acts as anET junction (11). According to the theory, an electron emergesfrom a donation site—likely a metal atom acting as a cofac-tor (11, 31), redox chemistry (59), or peptide sidechain (11, 60)capable of oxidation—and traverses the active site to an accep-tor site, which is likely a specific motif or residue sidechain. Asthe electron traverses the active site, it may undertake severalpaths: (i) elastic tunneling, where no energy is lost or gained bythe electron; (ii) inelastic tunneling (IET), where the electronmay donate or accept a quantum of energy during transfer; and(iii) subsequently higher ordered inelastic processes (61–63). Thehypothesized presence of a possible metal cofactor site—actingto assist either in binding or in a later activation step—at ORs,GPCRs, and non-GPCR chemokine receptors is supported byaltered behavioral response (64–66), physiological response (66–68), theory (69, 70), and in vitro observations (70–79).
Fig. 2. Plots of the tunneling probability as a function of energy, P(E), forvarious deuterated analogues of the DOI molecule. (Top Left) In orange,2H16 deuterated DOI; (Top Right) in green 2H18 deuterated DOI; (BottomLeft) in magenta, 2H16 and 2H17 deuterated DOI; and (Bottom Right) in cyan,2H16, 2H17, and 2H18 deuterated DOI. In all plots, blue is the all-protium DOItunneling spectrum.
5596 | www.pnas.org/cgi/doi/10.1073/pnas.1618422114 Hoehn et al.
BIO
PHYS
ICS
AN
DCO
MPU
TATI
ON
AL
BIO
LOG
Y
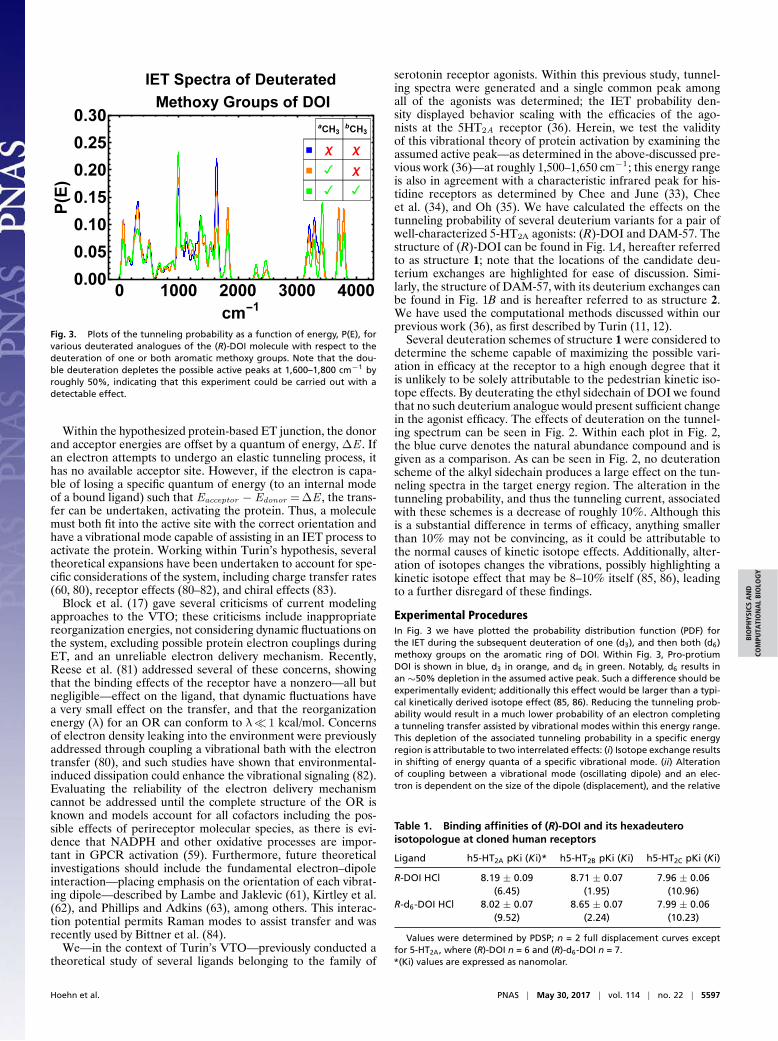
Fig. 3. Plots of the tunneling probability as a function of energy, P(E), forvarious deuterated analogues of the (R)-DOI molecule with respect to thedeuteration of one or both aromatic methoxy groups. Note that the dou-ble deuteration depletes the possible active peaks at 1,600–1,800 cm−1 byroughly 50%, indicating that this experiment could be carried out with adetectable effect.
Within the hypothesized protein-based ET junction, the donorand acceptor energies are offset by a quantum of energy, ∆E . Ifan electron attempts to undergo an elastic tunneling process, ithas no available acceptor site. However, if the electron is capa-ble of losing a specific quantum of energy (to an internal modeof a bound ligand) such that Eacceptor − Edonor = ∆E , the trans-fer can be undertaken, activating the protein. Thus, a moleculemust both fit into the active site with the correct orientation andhave a vibrational mode capable of assisting in an IET process toactivate the protein. Working within Turin’s hypothesis, severaltheoretical expansions have been undertaken to account for spe-cific considerations of the system, including charge transfer rates(60, 80), receptor effects (80–82), and chiral effects (83).
Block et al. (17) gave several criticisms of current modelingapproaches to the VTO; these criticisms include inappropriatereorganization energies, not considering dynamic fluctuations onthe system, excluding possible protein electron couplings duringET, and an unreliable electron delivery mechanism. Recently,Reese et al. (81) addressed several of these concerns, showingthat the binding effects of the receptor have a nonzero—all butnegligible—effect on the ligand, that dynamic fluctuations havea very small effect on the transfer, and that the reorganizationenergy (λ) for an OR can conform to λ� 1 kcal/mol. Concernsof electron density leaking into the environment were previouslyaddressed through coupling a vibrational bath with the electrontransfer (80), and such studies have shown that environmental-induced dissipation could enhance the vibrational signaling (82).Evaluating the reliability of the electron delivery mechanismcannot be addressed until the complete structure of the OR isknown and models account for all cofactors including the pos-sible effects of perireceptor molecular species, as there is evi-dence that NADPH and other oxidative processes are impor-tant in GPCR activation (59). Furthermore, future theoreticalinvestigations should include the fundamental electron–dipoleinteraction—placing emphasis on the orientation of each vibrat-ing dipole—described by Lambe and Jaklevic (61), Kirtley et al.(62), and Phillips and Adkins (63), among others. This interac-tion potential permits Raman modes to assist transfer and wasrecently used by Bittner et al. (84).
We—in the context of Turin’s VTO—previously conducted atheoretical study of several ligands belonging to the family of
serotonin receptor agonists. Within this previous study, tunnel-ing spectra were generated and a single common peak amongall of the agonists was determined; the IET probability den-sity displayed behavior scaling with the efficacies of the ago-nists at the 5HT2A receptor (36). Herein, we test the validityof this vibrational theory of protein activation by examining theassumed active peak—as determined in the above-discussed pre-vious work (36)—at roughly 1,500–1,650 cm−1; this energy rangeis also in agreement with a characteristic infrared peak for his-tidine receptors as determined by Chee and June (33), Cheeet al. (34), and Oh (35). We have calculated the effects on thetunneling probability of several deuterium variants for a pair ofwell-characterized 5-HT2A agonists: (R)-DOI and DAM-57. Thestructure of (R)-DOI can be found in Fig. 1A, hereafter referredto as structure 1; note that the locations of the candidate deu-terium exchanges are highlighted for ease of discussion. Simi-larly, the structure of DAM-57, with its deuterium exchanges canbe found in Fig. 1B and is hereafter referred to as structure 2.We have used the computational methods discussed within ourprevious work (36), as first described by Turin (11, 12).
Several deuteration schemes of structure 1 were considered todetermine the scheme capable of maximizing the possible vari-ation in efficacy at the receptor to a high enough degree that itis unlikely to be solely attributable to the pedestrian kinetic iso-tope effects. By deuterating the ethyl sidechain of DOI we foundthat no such deuterium analogue would present sufficient changein the agonist efficacy. The effects of deuteration on the tunnel-ing spectrum can be seen in Fig. 2. Within each plot in Fig. 2,the blue curve denotes the natural abundance compound and isgiven as a comparison. As can be seen in Fig. 2, no deuterationscheme of the alkyl sidechain produces a large effect on the tun-neling spectra in the target energy region. The alteration in thetunneling probability, and thus the tunneling current, associatedwith these schemes is a decrease of roughly 10%. Although thisis a substantial difference in terms of efficacy, anything smallerthan 10% may not be convincing, as it could be attributable tothe normal causes of kinetic isotope effects. Additionally, alter-ation of isotopes changes the vibrations, possibly highlighting akinetic isotope effect that may be 8–10% itself (85, 86), leadingto a further disregard of these findings.
Experimental ProceduresIn Fig. 3 we have plotted the probability distribution function (PDF) forthe IET during the subsequent deuteration of one (d3), and then both (d6)methoxy groups on the aromatic ring of DOI. Within Fig. 3, Pro-protiumDOI is shown in blue, d3 in orange, and d6 in green. Notably, d6 results inan∼50% depletion in the assumed active peak. Such a difference should beexperimentally evident; additionally this effect would be larger than a typi-cal kinetically derived isotope effect (85, 86). Reducing the tunneling prob-ability would result in a much lower probability of an electron completinga tunneling transfer assisted by vibrational modes within this energy range.This depletion of the associated tunneling probability in a specific energyregion is attributable to two interrelated effects: (i) Isotope exchange resultsin shifting of energy quanta of a specific vibrational mode. (ii) Alterationof coupling between a vibrational mode (oscillating dipole) and an elec-tron is dependent on the size of the dipole (displacement), and the relative
Table 1. Binding affinities of (R)-DOI and its hexadeuteroisotopologue at cloned human receptors
Ligand h5-HT2A pKi (Ki)* h5-HT2B pKi (Ki) h5-HT2C pKi (Ki)
R-DOI HCl 8.19 ± 0.09 8.71 ± 0.07 7.96 ± 0.06(6.45) (1.95) (10.96)
R-d6-DOI HCl 8.02 ± 0.07 8.65 ± 0.07 7.99 ± 0.06(9.52) (2.24) (10.23)
Values were determined by PDSP; n = 2 full displacement curves exceptfor 5-HT2A, where (R)-DOI n = 6 and (R)-d6-DOI n = 7.*(Ki) values are expressed as nanomolar.
Hoehn et al. PNAS | May 30, 2017 | vol. 114 | no. 22 | 5597
Table 2. Comparison of (R)-DOI and DAM-57 and their hexadeutero isotopologues foractivation of 5-HT2A/2B/2C measuring Gq-mediated calcium flux, as illustrated in Fig. 4
Gq calcium flux
5-HT2A 5-HT2B 5-HT2C
EC50, nM Emax EC50, nM Emax EC50, nM Emax
Ligands (pEC50± SEM) % 5-HT (pEC50± SEM) % 5-HT (pEC50± SEM) % 5-HT
5-HT 0.83 100 1.29 100 0.25 100(9.08 ± 0.02) (9.01 ± 0.12) (9.62 ± 0.10)
(R)-DOI 0.58 95 ± 4 4.80 94 ± 1 2.19 101 ± 2(9.25 ± 0.08) (8.43 ± 0.12) (8.70 ± 0.14)
(R)-d6-DOI 0.63 96 ± 4 4.40 91 ± 1 2.23 100 ± 3(9.22 ± 0.10) (8.45 ± 0.10) (8.70 ± 0.15)
DAM-57 1.54 98 ± 1 13.6 73 ± 2 57.5 81 ± 6(8.82 ± 0.06) (7.87 ± 0.04) (7.24 ± 0.02)
d6-DAM-57 1.51 98 ± 1 12.6 74 ± 1 49.2 83 ± 6(8.84 ± 0.07) (7.91 ± 0.05) (7.31 ± 0.02)
Calcium flux data were acquired with human 5-HT2A, 5-HT2B, and 5-HT2C INI-expressing tetracycline-inducible HEK cells. Estimates of EC50 and Emax represent the average and SE of the mean (SEM) from threeindependent experiments performed in triplicate. Emax is defined as percentage of 5-HT maximum response.
magnitude of an IET PDF peak associated with a mode is dependent on themass of an atom. This depletion of electron transfer should result in fewersuccessful activations of the protein than the natural abundance compound.It is for these reasons that the deuteration schemes involving exchange ofall six of the methoxy hydrogens of DOI and the methyl chains on the amideof DAM-57 were selected as candidates.
d6-(R)-DOI was prepared by an asymmetric synthesis, as shown in Fig. S1.NMR, mass spectral, and melting-point data were consistent with a pre-viously published synthesis of the protio compound (87). d6-DAM-57 wasprepared by a standard route (Fig. S2), using either dimethylamine or d6-dimethylamine. First, DOI and d6-DOI were submitted to the National Insti-tute of Mental Health (NIMH)-sponsored Psychoactive Drug Screening Pro-gram (PDSP) (88) to determine their binding affinities at the human 5-HT2A
receptor and then all compounds were tested at 5-HT2A, 5-HT2B, and 5-HT2C
receptors, measuring Gq-mediated calcium flux. A complete discussion ofthe synthetic routes for all molecules, as well as a discussion of the biologi-cal assays, is given in Supporting Information.
Results and DiscussionHerein, we operate under the following null hypothesis: Avibration-sensitive mechanism is shared between all membersof the GPCR class of proteins, possibly due to conserved topo-graphical structures even without conservation of sequence iden-tities, implying that general topographical structures of theGPCRs are essential in performing their biological task (89). Fol-lowing this logic, it seems reasonable that the family of GPCRslikely shares a common fundamental activation mechanism. Fur-thermore, forms of both the lock-and-key and the glove-and-hand models exist in olfactory research, admitting acceptanceof common aspects. Due to the likelihood of a common fun-damental activation mechanism, findings at CNS GPCRs willhave implications at mammalian ORs. We have used a seriesof well-characterized GPCRs (explicitly h5-HT2A, h5-HT2B, andh5-HT2C) with two established ligands: (R)-DOI and DAM-57.
IET spectra were generated for both molecules, using meth-ods prescribed by Turin in previous works (11, 12). Within aprevious work (36), we examined the IET spectrum of severalagonists and determined a consistently shared peak between theagonist’s spectra, the area of this shared spectral aspect roughlyscaled with the agonist’s efficacy. This peak was consistent withthe works of Chee and June (33), Chee et al. (34), and Oh(35), who attempted to generate a novel pharmacophore toolbased on shared vibrational peaks. From the candidate deuter-ation schemes discussed above, we sought the greatest possiblealteration in efficacy at the previously suggested activation peakto focus our experimental effort (36).
Both (R)-DOI and (R)-d6-DOI were synthesized and wereassayed for binding affinity (Table 1). All four compounds wereassayed for functional activity, using calcium flux assays (Fig.4 and Table 2) at the 5-HT2A, 5-HT2B, and 5-HT2C receptors.Neither (R)-d6-DOI nor d6-DAM-57 presented significant alter-ations in either binding or potency and efficacy (as noted by EC50
and Emax , respectively) compared with the protium versions. Theendogenous agonist 5-HT was included in the calcium flux assaysfor comparison.
The process of protein agonism/antagonism involves, at min-imum, two steps: binding of the ligand to the active site of theprotein and the activation of the protein. It should be noted thatthese actions may happen in concert as proposed in the hand-in-glove/multiconformation (5–8) models or as two individual steps(11, 60, 67, 80, 90). In Table 1, we present the results of bind-ing displacement assays comparing the relative binding affinityof both (R)-DOI and (R)-d6-DOI at several serotonin GPCRs.Alteration in the binding kinetics, as defined by pKi, shows no sig-nificant difference between the protonated and deuterated vari-ants; explicitly, any difference found in the power-scaled equilib-rium constant exists within the standard errors of the number, andtherefore no claim of a difference can be made with confidence.The kinetic isotope effect may also affect the binding kinetics ofthe G protein and thus could appear independently of ligand-binding effects but would be apparent in activation studies.
Fig. 4. Calcium flux responses at human 5-HT2A, 5-HT2B, and 5-HT2C
INI receptors for (R)-DOI (red), (R)-d6-DOI (orange), DAM-57 (blue), andd6-DAM-57 (purple). Data are representative at each receptor type per-formed in triplicate and in parallel.
5598 | www.pnas.org/cgi/doi/10.1073/pnas.1618422114 Hoehn et al.

BIO
PHYS
ICS
AN
DCO
MPU
TATI
ON
AL
BIO
LOG
Y
Activation analysis was conducted through a series of exper-iments at human tetracycline-inducible HEK cells expressingall members of the h5-HT2 receptor subclass. Receptor acti-vation was determined by calcium flux assay, dependent onGαq dissociation. The experiment was performed in triplicate(n = 3) to achieve relevant statistics. Results of the Gαq fluxcan be found in both Fig. 4 and Table 2 for five species. Itis clear from Fig. 4 that DOI is a near full agonist at allthree 5-HT2 receptors examined in this assay, whereas DAM-57shows partial agonist activity at 5-HT2B and 5-HT2C receptors.Additionally—and more prescient—there is no significant differ-ence in calcium flux between either pair of parent–isotopologuecompounds at any of the 5-HT2 receptors. This conclusion isillustrated in Table 2, which gives both the EC50 and the per-centage of Emax . The power-scaled pEC50, which estimates dif-ferences in potency between each pair of parent–isotopologuecompounds, is within the SE and likewise for the percentage ofmaximum response (where 100% is taken to be the response tothe endogenous 5-HT ligand). Therefore, no significant ligand-binding or activation (due to either ligand–receptor or ligand–receptor-G protein interactions) effects were found.
Herein, we found no exceptional dependency on isotopicexchange for the activity of either (R)-DOI or DAM-57. Thesefindings do not argue for the irrelevancy of isotopic exchange,but do relegate the majority of this concern to binding. Isotopeeffects on the binding of a ligand with a protein have been exam-ined extensively (85, 86) and were the subject of a recent paperon the H2 histamine receptor (58). Whereas Krzan et al. (58)detected small alterations in the binding of ligand to receptor dueto isotope exchange and conclude with possibly relevant com-ments concerning the VTO, we considered both the affinity andthe functional activity. By considering both aspects, we are ableto draw conclusions regarding the activation of the protein—addressing the VTO directly—while retaining knowledge of pos-sible binding effects. As this work was to evaluate the depen-dency of activation on vibrational modes of ligands in GPCRswithin the CNS through an IET-esque mechanism, we state that
we found no evidence suggestive of the plausibility of said theory.We believe that the findings within this paper argue very stronglyagainst the VTO. Furthermore, this places our work in agree-ment with the transition state theories (recent works: refs. 5–8).Physiochemical properties of the ligand—other than vibrationalquanta—are likely involved in the activation of both ORs—assuggested for Drosophila receptors by Saberi and Seyed-allaei(91)—and GPCRs in general.
ConclusionHerein, we attempted to examine the viability of a contentioustheory of protein activation—originally proposed for ORs—witha series of serotonin receptors widely expressed in the CNS:h5-HT2A, h5-HT2B, and h5-HT2C. We have tested two well-characterized ligands—and specific isotopologues—at the 5-HTreceptor family: (R)-DOI and DAM-57. Our calculations ofthe tunneling probability—based on Turin’s theory—predictedas much as a 50% loss of potency for both (R)-d6-DOI andd6-DAM-57, compared with their protium counterparts. Theminor deviations in the binding between (R)-DOI and (R)-d6-DOI were within the SE and therefore cannot be assumed to beof any significance, as were the changes in receptor function mea-sured by calcium flux. Similarly, no deviation in the potency orefficacy was detected for d6-DAM-57. As all values were withinthe SE, we can only conclude that alteration of the hydrogen iso-topes at (R)-DOI and DAM-57 agonists has no significant effecton the activity at the series of 5-HT receptors studied herein.Clearly, our results are not consistent with predictions madeunder the VTO acting within nonolfactory GPCRs. This callsinto question the viability of the IET mechanism within non-ORGPCRs, while additionally making it more difficult to argue infavor of the VTO without invoking exceptionalism of ORs withinthe GPCR class of proteins.
ACKNOWLEDGMENTS. The authors gratefully acknowledge assistance withthe NMR and MS analyses by Jarod Waybright. The authors acknowledgesupport from the National Institute of Mental Health-sponsored Psychoac-tive Drug Screening Program.
1. Kesner RP, Hunsaker MR, Ziegler W (2011) The role of the dorsal and ventral hip-pocampus in olfactory working memory. Neurobiol Learn Mem 96:361–366.
2. Rolls ET (2000) The orbitofrontal cortex and reward. Cereb cortex 10:284–294.3. Buck L, Axel R (1991) A novel multigene family may encode odorant receptors: A
molecular basis for odor recognition. Cell 65:175–187.4. Jastrzebska B, Debinski A, Filipek S, Palczewski K (2011) Role of membrane integrity
on G protein-coupled receptors: Rhodopsin stability and function. Prog Lipid Res50:267–277.
5. Manglik A, et al. (2015) Structural insights into the dynamic process of β2-adrenergicreceptor signaling. Cell 161:1101–1111.
6. Sounier R, et al. (2015) Propagation of conformational changes during µ-opioidreceptor activation. Nature 524:375–378.
7. Nygaard R, et al. (2013) The dynamic process of β2-adrenergic receptor activation.Cell 152:532–542.
8. Huang W, et al. (2015) Structural insights into µ-opioid receptor activation. Nature524:315–321.
9. Pande K, et al. (2016) Femtosecond structural dynamics drives the trans/cis isomeriza-tion in photoactive yellow protein. Science 352:725–729.
10. Nango E, et al. (2016) A three-dimensional movie of structural changes in bacteri-orhodopsin. Science 354:1552–1557.
11. Turin L (1996) A spectroscopic mechanism for primary olfactory reception. ChemSenses 21:773–791.
12. Turin L (2002) A method for the calculation of odor character from molecular struc-ture. J Theor Biol 216:367–385.
13. Haffenden L, Yaylayan V, Fortin J (2001) Investigation of vibrational theory of olfac-tion with variously labelled benzaldehydes. Food Chem 73:67–72.
14. Franco MI, Turin L, Mershin A, Skoulakis EMC (2011) Molecular vibration-sensingcomponent in Drosophila melanogaster olfaction. Proc Natl Acad Sci USA 108:3797–3802.
15. Gane S, et al. (2013) Molecular vibration-sensing component in human olfaction. PLoSOne 8:1–7.
16. Gronenberg W, et al. (2014) Honeybees (Apis mellifera) learn to discriminate the smellof organic compounds from their respective deuterated isotopomers. Proc Biol Sci281:20133089.
17. Block E, et al. (2015) Implausibility of the vibrational theory of olfaction. Proc NatlAcad Sci USA 112:E2766–E2774.
18. Keller A, Vosshall LB (2004) A psychophysical test of the vibration theory of olfaction.Nat Neurosci 7:337–338.
19. Dyson GM (1928) Some aspects of the vibration theory of odor. Perfum Essent Oil Rec19:456–459.
20. Wright RH (1954) Odour and molecular vibration. I. Quantum and thermodynamicconsiderations. J Appl Chem 4:611–615.
21. Wright RH, Serenius RSE (1954) Odour and molecular vibration. II. Raman spectra ofsubstances with the nitrobenzene odour. J Appl Chem 4:615–621.
22. Kim J, Matsuyama S, Suzuki T (2004) Deuterated analogues of 4,8-dimethyldecanal,the aggregation pheromone of Tribolium castaneum: Synthesis and pheromonalactivity. J Labelled Compd Radiopharm 47:921–934.
23. Drimyli E, Gaitanidis A, Maniati K, Turin L, Skoulakis EMC (2016) Differential electro-physiological responses to odorant isotopologues in drosophilid antennae. eNeuro3:015215.
24. Paoli M, et al. (2016) Differential odour coding of isotopomers in the honeybee brain.Sci Rep 6:21893.
25. Wright R (1977) Odor and molecular vibration: Neural coding of olfactory informa-tion. J Theor Biol 64:473–502.
26. Wright RH (1983) Molecular vibration and odour blending. Chem Senses 8:103–106.27. Doolittle RE, Beroza M, Keiser I, Schneider EL (1968) Deuteration of the melon fly
attractant, cue-lure, and its effect on olfactory response and infra-red absorption. JInsect Physiol 14:1697–1712.
28. Blum MS, Doolittle RE, Beroza M (1971) Alarm pheromones: Utilization in evaluationof olfactory theories. J Insect Physiol 17:2351–2361.
29. Barker RJ, Berdel RL, Waller GD (1973) The molecular basis for scent discrimina-tion: Response to nitrobenzene-d5 of honey bees (Apis mellifera L.) conditioned withnitrobenzene. Experientia 29:418–419.
30. Wright RH (1975) Odor and molecular vibration: Response to nitrobenzene-d5 ofhoney bees (Apis mellifera L.) conditioned with nitrobenzene. Experientia 31:530.
31. Turin L, Yoshii F (2003) Structure–odor relations: A modern perspective. Handbook ofOlfaction and Gustation, ed Doty RL (CRC, Oxford), 2nd Ed, pp 457–515.
32. Maia ER, Magalhaes DRB, Lerner DA, Berthomieu D, Bernassau JM (2014) Quantumcalculation for musk molecules infrared spectra towards the understanding of odor.Adv Chem 2014:1–13.
33. Chee HK, June OS (2013) Molecular vibration-activity relationship in the agonism ofadenosine receptors. Genomics Inform 11:282–288.
Hoehn et al. PNAS | May 30, 2017 | vol. 114 | no. 22 | 5599

34. Chee HK, Yang JS, Joung JG, Zhang BT, Oh SJ (2015) Characteristic molecular vibra-tions of adenosine receptor ligands. FEBS Lett 589:548–552.
35. Oh SJ (2012) Characteristics in molecular vibrational frequency patterns between ago-nists and antagonists of histamine receptors. Genomics Inform 10:128–132.
36. Hoehn RD, Nichols D, Hartmut N, Sabre K (2015) Neuroreceptor activation byvibration-assisted tunneling. Sci Rep 5:9990.
37. Klika KD (2013) The potential of 13C isotopomers as a test for the vibrational theoryof olfactory sense recognition. ISRN Org Chem 2013:515810.
38. Turin L, Gane S, Georganakis D, Maniati K, Skoulakis EMC (2015) Plausibility of thevibrational theory of olfaction. Proc Natl Acad Sci USA 112:E3154.
39. Block E, Jang S, Matsunami H, Batista VS, Zhuang H (2015) Reply to turin et al.: Vibra-tional theory of olfaction is implausible. Proc Natl Acad Sci USA 112:E3155.
40. Kaupp UB (2010) Olfactory signalling in vertebrates and insects: Differences and com-monalities. Nat Rev Neurosci 11:188–200.
41. Pellegrino M, Nakagawa T (2009) Smelling the difference: Controversial ideas in insectolfaction. J Exp Biol 212:1973–1979.
42. Barwich AS (2016) What is so special about smell? Olfaction as a model system inneurobiology. Postgrad Med J 92:27–33.
43. Crasto CJ (2009) Computational biology of olfactory receptors. Curr Bioinform 4:8–15.44. Abaffy T (2015) Human olfactory receptors expression and their role in non-olfactory
tissues a mini-review. J Pharmacogenomics Pharmacoproteomics 6:152.45. Kang N, Koo J (2012) Olfactory receptors in non-chemosensory tissues. BMB Rep
45:612–622.46. Feldmesser E, et al. (2006) Widespread ectopic expression of olfactory receptor genes.
BMC Genomics 7:121–121.47. Vanderhaeghen P, Schurmans S, Vassart G, Parmentier M (1993) Olfactory receptors
are displayed on dog mature sperm cells. J Cell Biol 123:1441–1452.48. Flegel C, et al. (2016) Characterization of the olfactory receptors expressed in human
spermatozoa. Front Mol Biosci 2:73.49. Malki A, et al. (2015) Class I odorant receptors, TAS1R and TAS2R taste receptors, are
markers for subpopulations of circulating leukocytes. J Leukoc Biol 97:533–545.50. Gaudin JC, Breuils L, Haertle T (2006) Mouse orthologs of human olfactory-like recep-
tors expressed in the tongue. Gene 381:42–48.51. Blache P, Gros L, Salazar G, Bataille D (1998) Cloning and tissue distribution of a new
rat olfactory receptor-like (OL2). Biochem Biophys Res Commun 242:669–672.52. Weber M, Pehl U, Breer H, Strotmann J (2002) Olfactory receptor expressed in ganglia
of the autonomic nervous system. J Neurosci Res 68:176–184.53. Pluznick JL, et al. (2009) Functional expression of the olfactory signaling system in the
kidney. Proc Natl Acad Sci USA 106:2059–2064.54. Goto T, Salpekar A, Monk M (2001) Expression of a testis-specific member of the olfac-
tory receptor gene family in human primordial germ cells. Mol Hum Reprod 7:553–558.
55. Drutel G, et al. (1995) Cloning of OL1, a putative olfactory receptor and its expressionin the developing rat heart. Recept Channels 3:33–40.
56. Shulgin A, Shulgin A (1991) PIHKAL: A Chemical Love Story (Transform Press, Berkeley,CA).
57. Shulgin A, Shulgin A (1997) TIHKAL: The Continuation (Transform Press, Berkeley, CA).58. Krzan M, et al. (2016) The quantum nature of drug-receptor interactions: Deuteration
changes binding affinities for histamine receptor ligands. PLoS One 11:1–16.59. Ushio-Fukai M (2009) Vascular signaling through G protein coupled receptors - new
concepts. Curr Opin Nephrol Hypertens 18:153–159.60. Brookes JC, Hartoutsiou F, Horsfield AP, Stoneham AM (2007) Could humans recognize
odor by phonon assisted tunneling? Phys Rev Lett 98:038101.61. Lambe J, Jaklevic RC (1968) Molecular vibration spectra by inelastic electron tunnel-
ing. Phys Rev 165:821–832.62. Kirtley J, Scalapino DJ, Hansma PK (1976) Theory of vibrational mode intensities in
inelastic electron tunneling spectroscopy. Phys Rev B 14:3177–3184.63. Phillips WA, Adkins CJ (1985) A theory for the intensities of inelastic electron-
tunnelling spectra. Philos Mag B 52:739–750.64. Frazier JL, Heitz JR (1975) Inhibition of olfaction in the moth Heliothis virescens by
the sulfhydryl reagent fluorescein mercuric acetate. Chem Senses 1:271–281.65. Jia H, et al. (2016) Enhancement of odor-induced activity in the canine brain by zinc
nanoparticles: A functional MRI study in fully unrestrained conscious dogs. ChemSenses 41:53–67.
66. Moore CH, et al. (2012) Olfactory responses to explosives associated odorants areenhanced by zinc nanoparticles. Talanta 88:730–733.
67. Wang J, Luthey-Schulten ZA, Suslick KS (2003) Is the olfactory receptor a metallopro-tein? Proc Natl Acad Sci USA 100:3035–3039.
68. Viswaprakash N, et al. (2009) Enhancement of odorant-induced responses in olfactoryreceptor neurons by zinc nanoparticles. Chem Senses 34:547–557.
69. Vodyanoy V (2010) Zinc nanoparticles interact with olfactory receptor neurons.BioMetals 23:1097–1103.
70. Hu C, Chan SI, Sawyer EB, Yu Y, Wang J (2014) Metalloprotein design using geneticcode expansion. Chem Soc Rev 43:6498–6510.
71. Seebungkert B, Lynch JW (2001) A common inhibitory binding site for zinc and odor-ants at the voltage-gated K+ channel of rat olfactory receptor neurons. Eur J Neu-rosci 14:353–362.
72. Kinouchi K, Standifer KM, Pasternak GW (1990) Modulation of µ1, µ2, and δ opioidbinding by divalent cations. Biochem Pharmacol 40:382–384.
73. Standifer KM, Clark JA, Pasternak GW (1993) Modulation of µ1 opioid bindingby magnesium: Evidence for multiple receptor conformations. J Pharm Exp Ther266:106–113.
74. Holst B, Elling CE, Schwartz TW (2002) Metal ion-mediated agonism and agonistenhancement in melanocortin MC1 and MC4 receptors. J Biol Chem 277:47662–47670.
75. Li S, et al. (2016) Smelling sulfur: Copper and silver regulate the response of humanodorant receptor OR2T11 to low-molecular-weight thiols. J Am Chem Soc 138:13281–13288.
76. Block E (2017) Fifty years of smelling sulfur: From the chemistry of garlic to the molec-ular basis for olfaction. Phosphorus Sulfur Silicon Relat Elem 192:141–144.
77. Duan X, et al. (2012) Crucial role of copper in detection of metal-coordinating odor-ants. Proc Natl Acad Sci USA 109:3492–3497.
78. Gerlach LO, et al. (2003) Metal ion enhanced binding of AMD3100 to Asp262 in theCXCR4 receptor. Biochemistry 42:710–717.
79. Rodrıguez FI, et al. (1999) A copper cofactor for the ethylene receptor ETR1 fromArabidopsis. Science 283:996–998.
80. Solov’yov IA, Chang PY, Schulten K (2012) Vibrationally assisted electron transfermechanism of olfaction: Myth or reality? Phys Chem Chem Phys 14:13861–13871.
81. Reese A, List NH, Kongsted J, Solov’yov IA (2016) How far does a receptor influencevibrational properties of an odorant? PLoS One 11:1–21.
82. Checinska A, Pollock FA, Heaney L, Nazir A (2015) Dissipation enhanced vibrationalsensing in an olfactory molecular switch. J Chem Phys 142:025102.
83. Tirandaz A, Taher Ghahramani F, Shafiee A (2015) Dissipative vibrational model forchiral recognition in olfaction. Phys Rev E 92:032724.
84. Bittner ER, Madalan A, Czader A, Roman G (2012) Quantum origins of molecularrecognition and olfaction in Drosophila. J Chem Phys 137:22A551.
85. Rakowski K, Paneth P (1996) Isotope effects on binding. J Mol Struct 378:35–43.86. Swiderek K, Paneth P (2013) Binding isotope effects. Chem Rev 113:7851–7879.87. Mathis CA, Hoffman AJ, Nichols DE, Shulgin AT (1988) Synthesis of high specific activ-
ity 125I- and 123I-labelled enantiomers of 2,5-dimethoxy-4-iodophenylisopropylamine(DOI). J Labelled Comp Radiopharm 25:1255–1265.
88. Besnard J, et al. (2012) Automated design of ligands to polypharmacological profiles.Nature 492:215–220.
89. Zhang Z, Wu J, Yu J, Xiao J (2012) A brief review on the evolution of GPCR: Conserva-tion and diversification. Open J Genet 2:11–17.
90. Brookes JC, Horsfield AP, Stoneham AM (2012) The swipe card model of odorantrecognition. Sensors 12:15709–15749.
91. Saberi M, Seyed-allaei H (2016) Odorant receptors of Drosophila are sensitive to themolecular volume of odorants. Sci Rep 6:25103.
92. Curphey TJ (1979) Trifluoroacetylation of amino acids and peptides by ethyl trifluo-roacetate. J Org Chem 44:2805–2807.
93. Chambers JJ, Kurrasch-Orbaugh DM, Parker MA, Nichols DE (2001) Enantiospecificsynthesis and pharmacological evaluation of a series of super-potent, conformation-ally restricted 5-HT2A/2C receptor agonists. J Med Chem 44:1003–1010.
94. Stoll A, Hofmann A (1955) Amide der stereoisomeren lysergsauren und dihydro-lysergsauren. 38. Mitteilung uber mutterkornalkaloide [Amides of the stereoisomersof lysergic acid and dihydrolysergic acid. 38. Report about Ergot alkaloids]. Helv ChimActa 38:421–433. German.
5600 | www.pnas.org/cgi/doi/10.1073/pnas.1618422114 Hoehn et al.















![Experimental demonstration of Generalized Phase Contrast ......Generalized Phase Contrast technique (GPC) [14]. GPC which is a pure phase modulation technique can be considered as](https://img.pdfslide.us/doc/110x75/60e813995a6cca3dbd45a9d9/experimental-demonstration-of-generalized-phase-contrast-generalized-phase.jpg)













