Embed Size (px)
Citation preview

Exam tonight! Here! 6:30 pm-8:30 pm
Hints:
1. 15 points The Double Helix2. 18 points DNA structure (this requires intuitive thinking
and introduces you to the technique of nearest-neighbor analysis)
3. 17 points DNA repair4. 20 points life cycle strategies and RNA synthesis5. 15 points catabolite repression6. 15 points attenuation


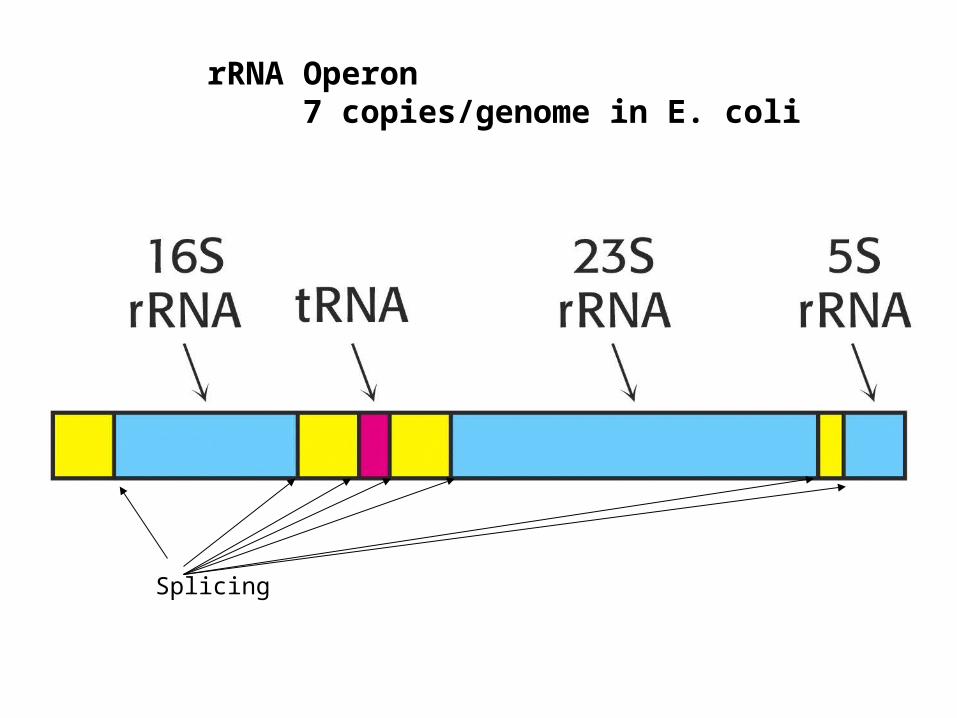
Splicing
rRNA Operon7 copies/genome in E. coli

Figure 32-8 Base sequence of yeast
tRNAAla drawn in the cloverleaf form.
Figure 32-9 Cloverleaf secondary structure of tRNA.

Figure 32-10 A selection of the modified nucleosides that occur in tRNAs together with their
standard abbreviations.
Pag
e 12
94

Figure 32-10 (continued) A selection of the modified nucleosides that occur in tRNAs together
with their standard abbreviations.
Pag
e 12
94

Nucleotidyl transferase(template independent)

Figure 32-11b Structure of yeast tRNAPhe. (b) The X-ray structure drawn to show how its base paired stems are arranged form the L-shaped molecule.
Pag
e 12
95

Figure 32-12 Tertiary base pairing interactions in yeast tRNAPhe.
Pag
e 12
96

Pag
e 12
99
Figure 32-16Experimentally observed
identity elements of RNAs.


An Aminoacyl–tRNA.P
age
1297

Figure 32-17aX-Ray structure of E. coli GlnRS · tRNAGln · ATP. (a) tRNA and ATP wireframe; tRNA sugar–phosphates green, bases magenta, ATP red.
Pag
e 13
00

PD molecule of the month_the_month

Different Approaches to the Same ProblemIn this picture, five complexes of an aminoacyl-tRNA synthetase with tRNA are shown, aligned so that the tRNA molecules are in the same orientation. Notice that the enzymes approach the tRNA from different angles. The isoleucine, valine and glutamine enzymes cradle the tRNA, gripping the anticodon loop (at the bottom in each tRNA), and placing the amino-acid acceptor end of the tRNA in the active site (at the top right in each tRNA). These all share a similar protein framework, known as "Type I," approaching the tRNA similarly and adding the amino acid to the last 2' hydroxyl group in the tRNA.
The phenlyalanine and threonine enzymes are part of a second class of enzymes, known as "Type II." They approach the tRNA from the other side, and add the amino acid to the 3’ hydroxyl on the last tRNA base.
Type I enzymes contain a Rossmann fold in the catalytic domain(alternating αβ structures)

Rossmann fold from a decarboxylase
What’s bound to this enzyme????? Nicotinaminde mononucleotide

BIOINFORMATICS DISCOVERY NOTE Vol. 26 no. 6 2010: 709–714, 2010Six Rossmannoid folds, including the Class I aminoacyl-tRNAsynthetases, share a partial core with the anti-codon-bindingdomain of a Class II aminoacyl-tRNA synthetaseStephen Cammer1, and Charles W. Carter Jr2∗

ATP binding site of Met tRNA synthetase
ε-amino group of his 21 is close to the phosphates of ATP.
Nucleic Acids Res. 1995 December 11; 23(23): 4793–4798. Transition state stabilization by the 'high' motif of class I aminoacyl-tRNA synthetases: the case of Escherichia coli methionyl-tRNA synthetase.E Schmitt, M Panvert, S Blanquet, and Y Mechulam

Figure 32-19a Comparison of the modes by which GlnRS and AspRS bind their cognate
tRNAs.
Pag
e 13
02
(b) AspRS, a Class II synthetase.(a) GlnRS, a Class I synthetase.

Figure 32-20 Comparison of the stereochemistries of
aminoacylation by Class I and Class II aaRSs.
Pag
e 13
03

Figure 32-22a Superposition of tRNAIle in the putative aminoacylation and editing
modes of IleRS · tRNAIle.
Pag
e 13
04

Figure 32-22b Cartoon comparison of the putative aminoacylation and editing modes of
IleRS · tRNAIle.
Pag
e 13
04
http://pubs.acs.org/doi/full/10.1021/ja9095208

Figure 32-23 Schematic diagram of the aminoacylation and editing mechanisms of
Class I and Class II aaRSs emphasizing the “mirror symmetry” of their overall
mechanisms.
Pag
e 13
04

Figure 32-24 The Glu-AdT–mediated synthesis of Gln– tRNAGln from Glu–tRNAGln.
Pag
e 13
06

Table 32-2 The “Standard” Genetic Codea.
Pag
e 12
90

Table 32-1 Amino Acid Incorporation Stimulated by a Random Copolymer of U and G in Mole Ratio 0.76:0.24.
Synthesized by polynucleotide phosphorylase (template independent)
Pag
e 12
89

mRNA/tRNA binding is antiparallel
• 5’
5’ 3’
mRNA
tRNA
CCA-amino acid (3’end)
5’end
Codon: 1 2 3
Anticodon 3 2 1
Anticodon 3’---UUG----5’ Codon 5’----CCU----3’
Wobble base

Table 32-5 Allowed Wobble Pairing Combinations in the Third Codon–Anticodon Position.
Pag
e 13
08

Figure 32-25a Wobble pairing. (a) U · G and I · A wobble pairs. Both have been observed in X-ray
structures.
Pag
e 13
07

Wobble

FIG. 1. Mechanisms of aminoacyl-tRNA formation. Both pathways, direct acylation and tRNA-dependent amino acid modification, are depicted for glutaminyl-tRNA formation. For example, E. coli uses glutaminyl-tRNA synthetase while B. subtilis employs Glu-tRNAGln amidotransferase for this purpose. http://mmbr.asm.org/cgi/content/full/64/1/202Microbiology and Molecular Biology Reviews, March 2000, p. 202-236, Vol. 64, No. 1Aminoacyl-tRNA Synthetases, the Genetic Code, and the Evolutionary Process . Carl R. Woese,1 Gary J. Olsen,1 Michael Ibba,2 and Dieter Söll3,

Species ARSs Function References
Escherichia coli TRS Translational control Romby et al., 1996; Brunel et al., 1993
E. coli AlaRS Transcription control Putney et al., 1981
Neurospora crassa Mitochondrial YRS Splicing Akins et al., 1987; Cherniack et al., 1990
Saccharomyces cerevisiae Mitochondrial LRS Splicing Herbert et al., 1988; Labouesse, 1990
Homo sapiens Cytoplasmic YRS Angiogenic cytokine Wakasugi et al., 2002a; Wakasugi et al., 2002b
H. sapiens Cytoplasmic WRS Angiostatic cytokine Otani et al., 2002; Wakasugi et al., 2002b
H. sapiens Cytoplasmic MRS Transcription of rRNA Ko et al., 2000
H. sapiens Cytoplasmic QRS Anti-apoptosis Ko et al., 2001a
Table 1. Non-canonical activities of ARSs

Activation


Pag
e 13
11
Figure 32-28 Two-dimensional gel electrophoretogram of E. coli small ribosomal subunit proteins.

Table 32-7Components of E. coli Ribosomes.
Pag
e 13
10

Figure 32-76 Reactions
involved in the attachment of ubiquitin to a
protein.

Figure 32-27a Secondary structures of the E. coli ribosomal RNAs. (a) 16S RNA.
Pag
e 13
11

Figure 32-30 Assembly map of the E. coli small subunit.
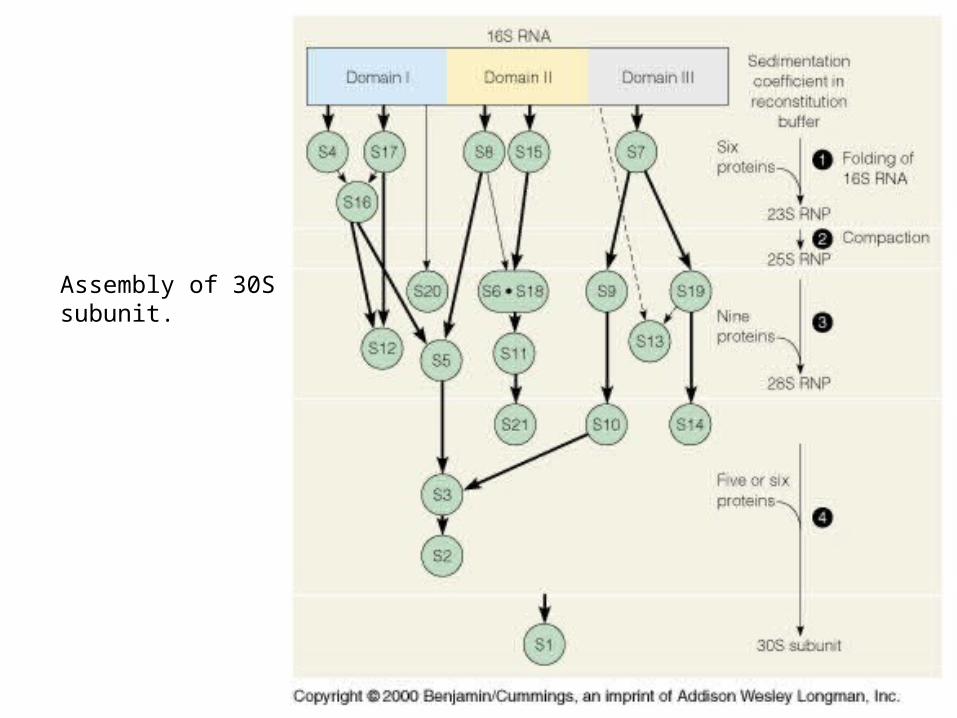
Assembly of 30Ssubunit.


Model from 0.9 Å x-ray data.

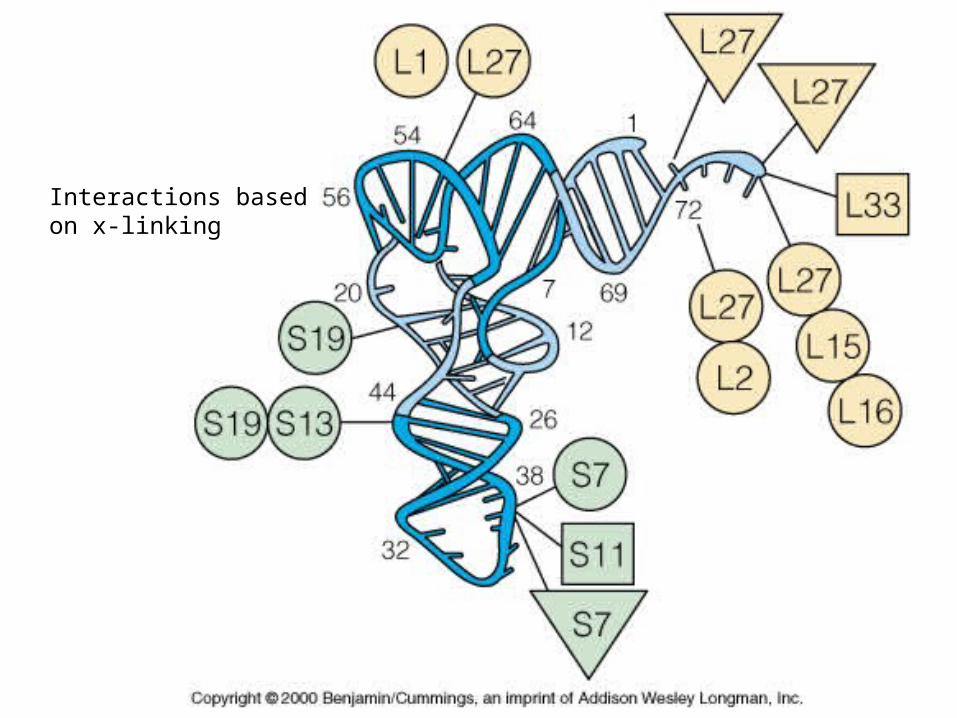
Interactions basedon x-linking
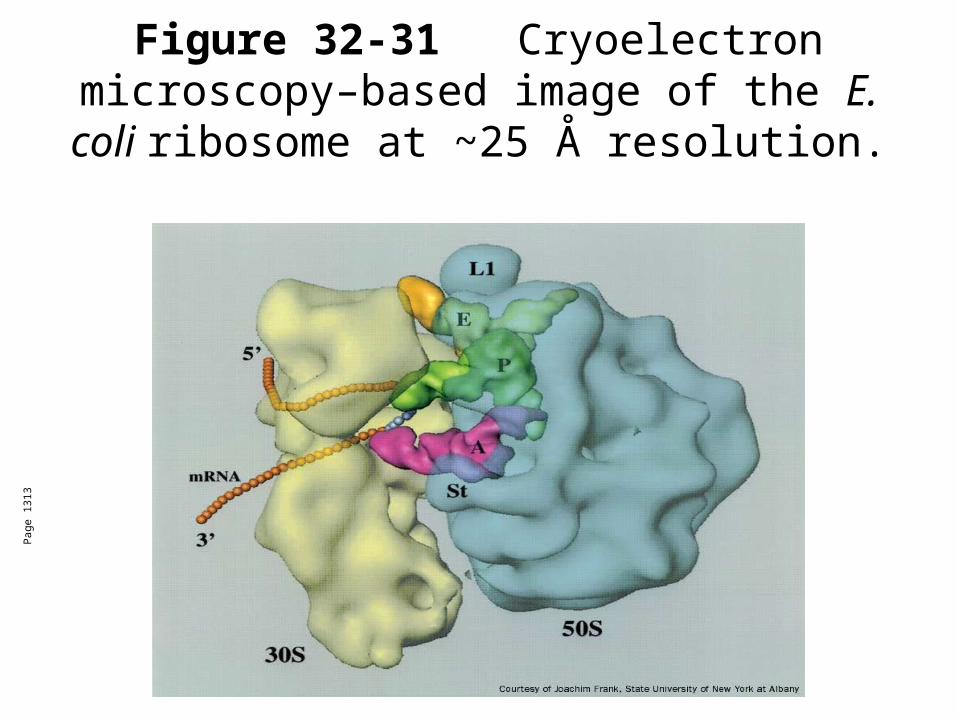
Figure 32-31 Cryoelectron microscopy–based image of the E. coli ribosome at ~25
Å resolution.
Pag
e 13
13

Table 32-10 Some Ribosomal Inhibitors.
Pag
e 13
39

Figure 32-65Selection of
antibiotics that act as translational
inhibitors.
Pag
e 13
40

Table 32-9 The Soluble
Protein Factors of E. coli Protein Synthesis.

Figure 32-45 Translational
initiation pathway in E.
coli.

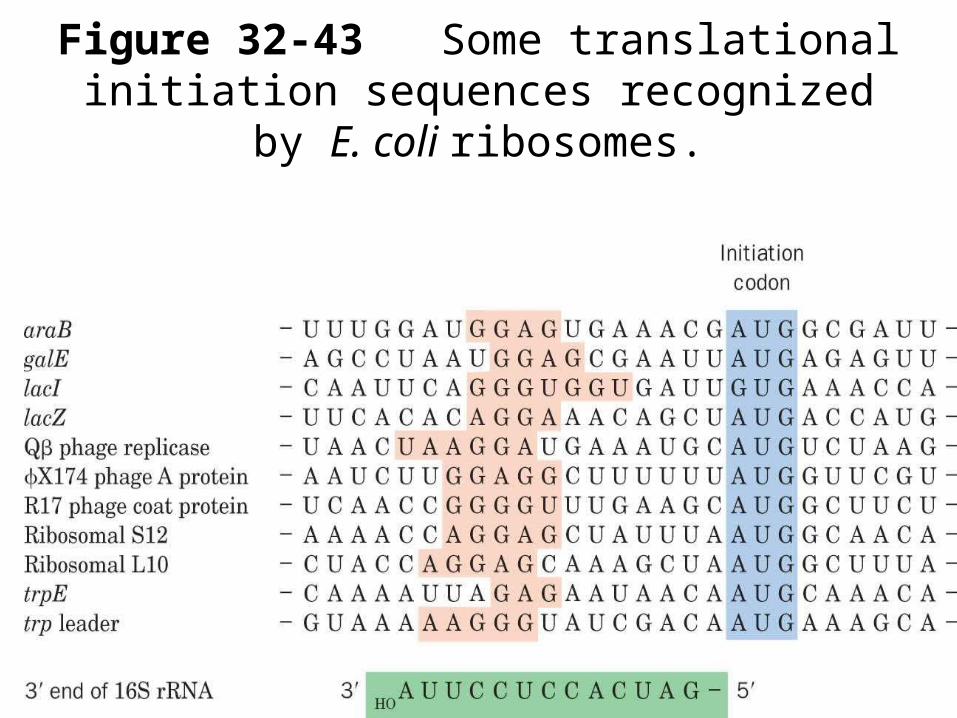
Figure 32-43 Some translational initiation sequences recognized by E. coli ribosomes.

SD=Shine-Dalgarno sequence complimentaryto rRNA L10

Figure 32-39 Demonstration that polypeptide synthesis proceeds from the N-
terminus to the C-terminus.
Pag
e 13
19

Figure 32-41 Ribosomal peptidyl transferase reaction forming a peptide bond.
Pag
e 13
20

Figure 32-52 Puromycin.P
age
1329

Figure 32-53 Ribosomal tetrahedral intermediate and its analog. (a) Tetrahedral intermediate (red C). (b) CCdA-p-Puro. Tetrahedral phosphoryl group (red P).

Peptidyl transferase activity not assoicated with proteins!!!
The ribosome is a ribozyme!!!!!
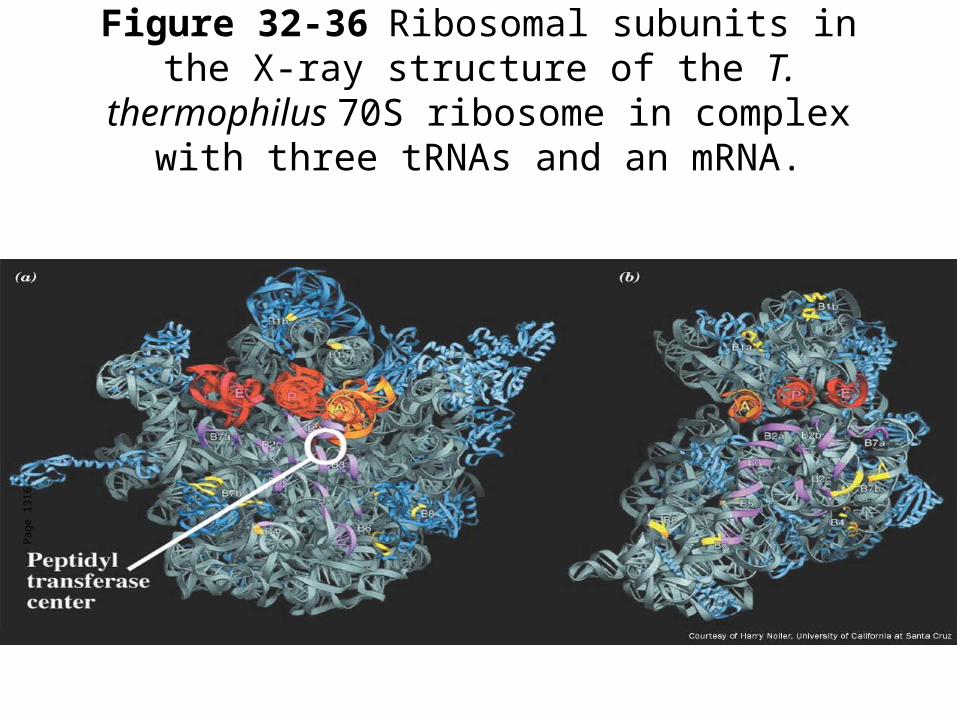
Figure 32-36 Ribosomal subunits in the X-ray structure of the T. thermophilus 70S ribosome in
complex with three tRNAs and an mRNA.
Pag
e 13
16

Figure 32-54 Proposed
mechanism of ribosomal peptide
synthesis.
Pag
e 13
31

Figure 32-48Elongation cycle in E. coli ribosomes. The E site, to which discharged tRNAs are transferred before being released into solution, is not shown.
Pag
e 13
27

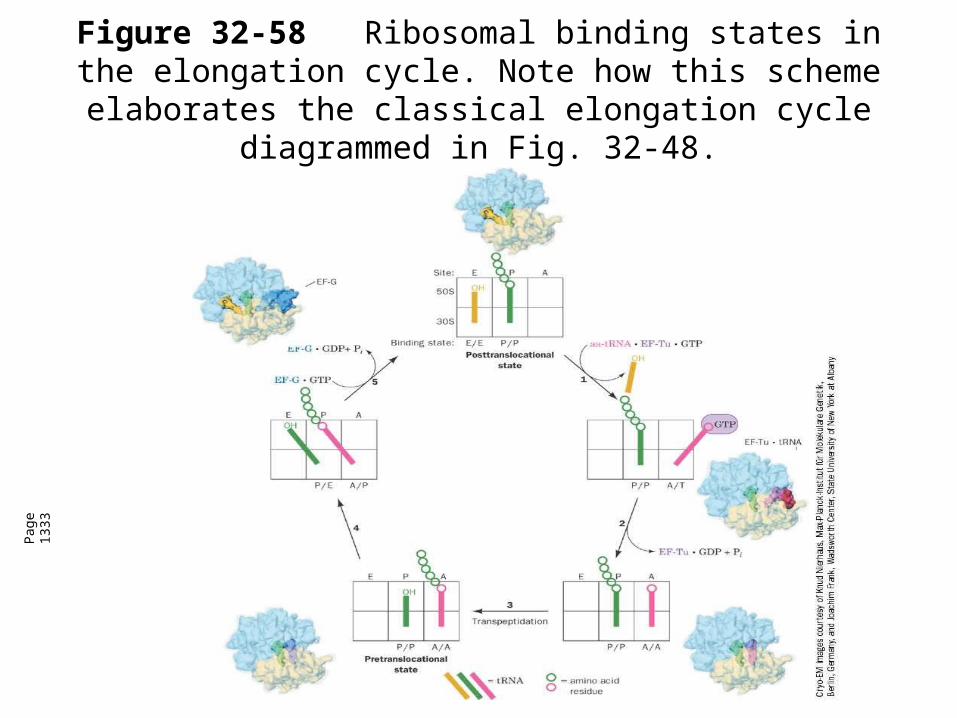
Figure 32-58Ribosomal binding states in the elongation cycle. Note how this scheme elaborates the classical
elongation cycle diagrammed in Fig. 32-48.
Pag
e 13
33

Figure 32-60Termination pathway in E. coli ribosomes. RF-1 recognizes the
Stop codons UAA and UAG, whereas
RF-2 (not shown) recognizes UAA and
UGA.
Pag
e 13
35

Figure 32-61 Ribosome-catalyzed hydrolysis of peptidyl–tRNA to form a polypeptide and free
tRNA.

MVA Fig. 27.29 Polyribosomes

Energetics: 4 high energy bonds/aa
YOU do the math!!!

MVA Fig. 27.30 Secretion

Figure 32-46Translational initiation pathway in eukaryotes.

MVA Fig.28.36ADP-ribosylated dipthamide derivative of his in eEF2

MVA Fig. 28.37

MVA Fig. 28.38
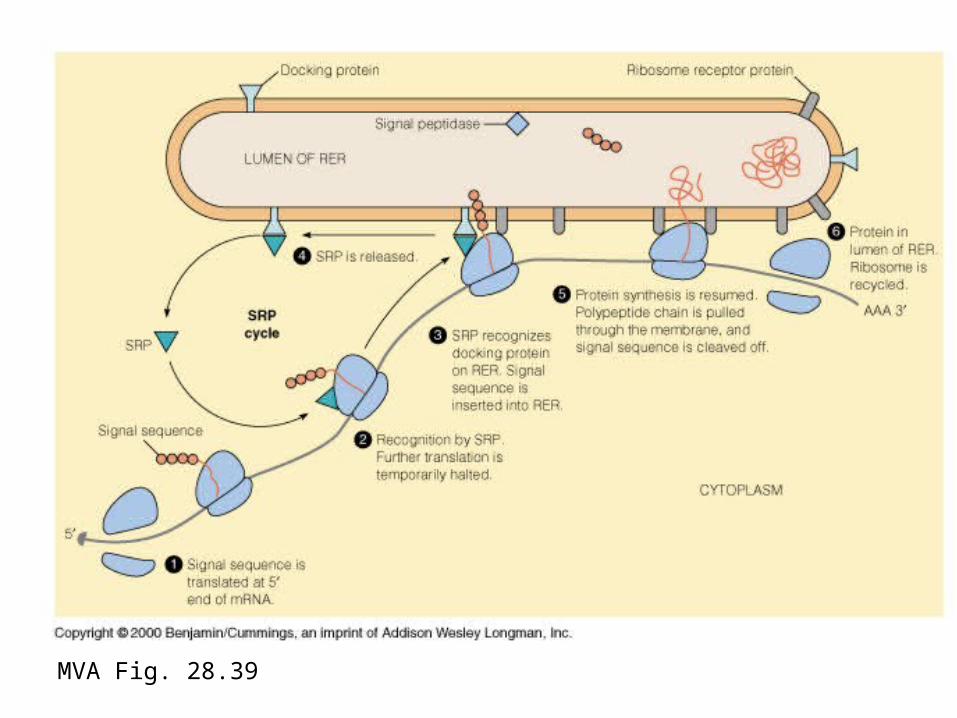
MVA Fig. 28.39

MVA Fig. 28.40

MVA Fig. 28.41
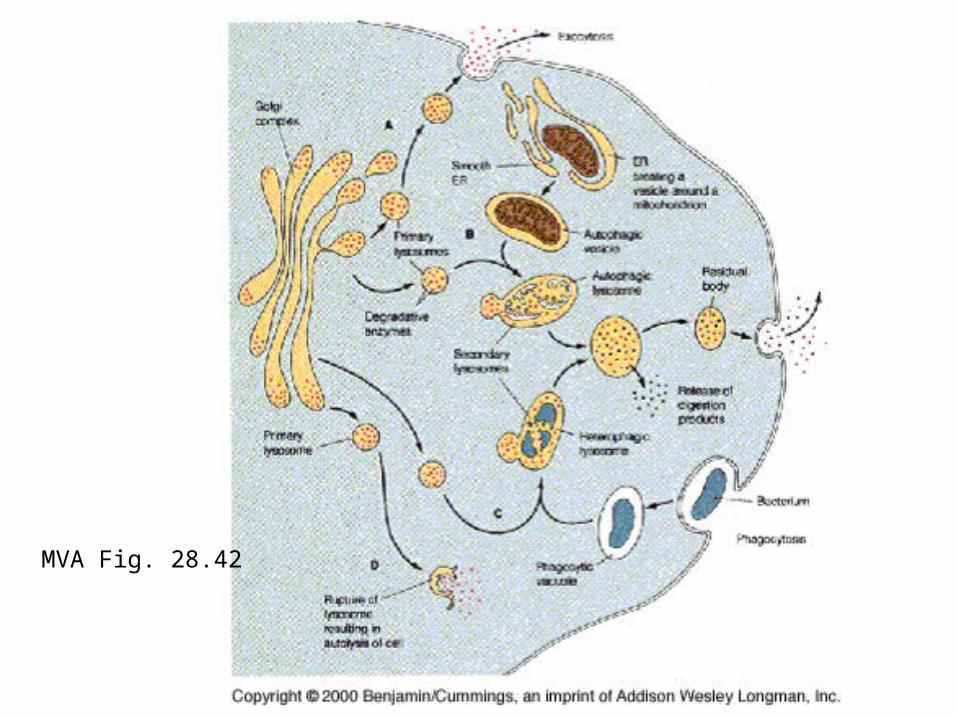
MVA Fig. 28.42
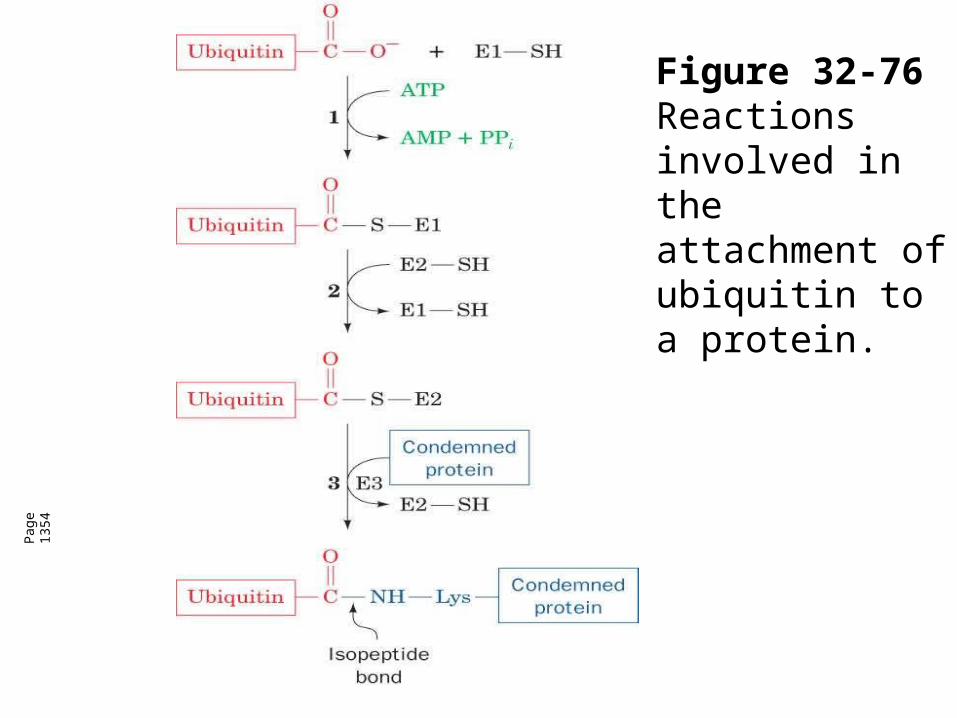
Figure 32-76 Reactions involved in the attachment of ubiquitin to a protein.
Pag
e 13
54

Table 32-12 Half-lives of Cytoplasmic Enzymes as a Function of Their N-Terminal
Residues.
Pag
e 13
57

MVA Fig.28.36ADP-ribosylated dipthamide derivattive of his in eEF2
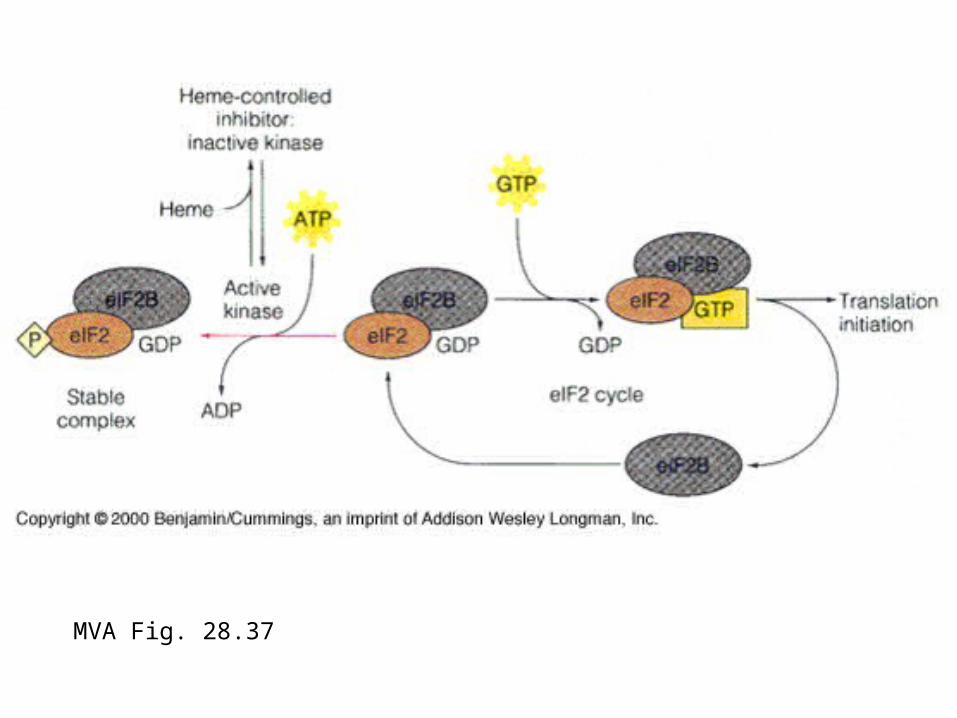
MVA Fig. 28.37

MVA Fig. 28.38

MVA Fig. 28.39

MVA Fig. 28.40

MVA Fig. 28.41

MVA Fig. 28.42

Figure 32-76 Reactions
involved in the attachment of ubiquitin to a
protein.

Figure 32-27a Secondary structures of the E. coli ribosomal RNAs. (a) 16S RNA.
Pag
e 13
11

Figure 32-30 Assembly map of the E. coli small subunit.

Assembly of 30Ssubunit.

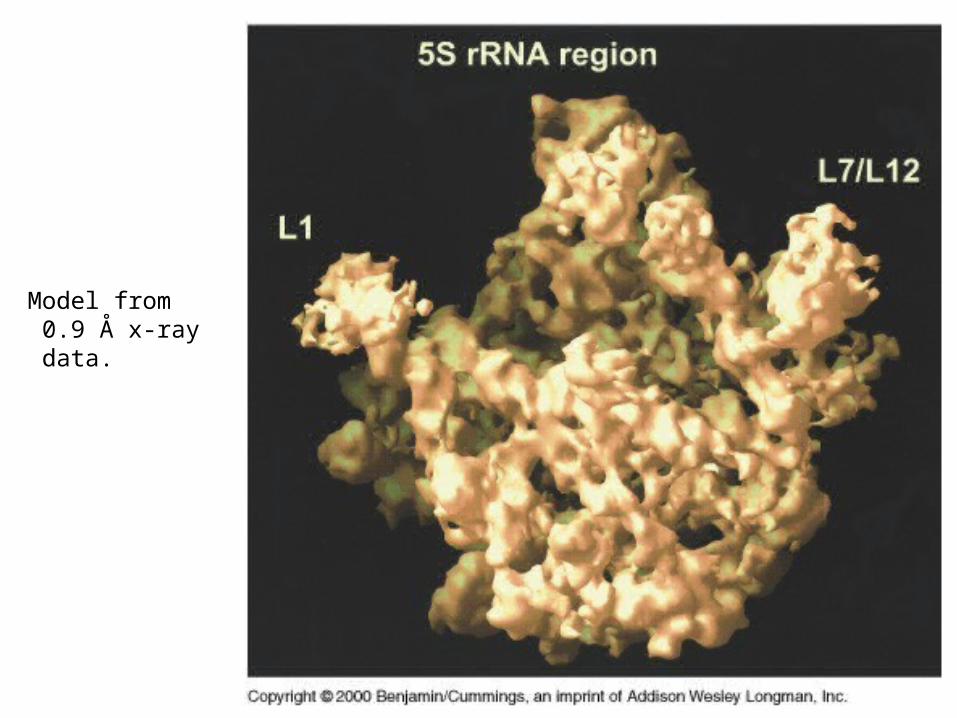
Model from 0.9 Å x-ray data.

Figure 32-36 Ribosomal subunits in the X-ray structure of the T. thermophilus 70S ribosome in
complex with three tRNAs and an mRNA.
Pag
e 13
16


Interactions basedon x-linking

Figure 32-31 Cryoelectron microscopy–based image of the E. coli ribosome at ~25
Å resolution.
Pag
e 13
13

Lee, S. W. et al. J Cell Sci 2004;117:3725-3734
Three-dimensional structure of the human multi-synthetase complex
Three-dimensional structure of the human multi-synthetase complex. (A) `Front' view.
(B) `Side' view created by –90° rotation about the vertical axis. (C) `Top' view created by –90° rotation about the horizontal axis. The
multi-synthetase complex was isolated from cultured human cells and prepared for
electron microscopy by negatively staining with NanoVan (Wolfe et al., 2003). The
volume was calculated from 8437 images, filtered to its resolution limit of 33 Å and
presented at a threshold corresponding to a particle mass of 1.2x106 Da.

Left and right panels of the figure represent sequence and structural comparison of bacterial type Plasmodium TyrRS (PF11_0181) with human mitochondrial TyrRS (2PID) and the cytosolic Plasmodium TyrRS (Mal8p1.125) with human cytosolic TyrRS (1N3L). a) A structure-based sequence alignment of the catalytic domain of Plasmodium TyrRSs with human TyrRSs. Insertions in Pf and human sequences are colored in light blue and orange respectively. Class I synthetase conserved motifs are colored red. Residues involved in tRNA recognition and catalysis are indicated in green (same residues in Pf and Hs) and violet & boxed (different in Pf and Hs). The secondary structural elements are shown above the sequence alignments. Conserved residues are indicated by asterisk below the sequence alignment

(b) Superposition of Pf-TyrRS and Hs-TyrRS depicting the structural differences. Pf-Tyr is colored grey and Hs-TyrRS is colored tan. Insertions in Pf-TyrRSs are highlighted in blue whereas Hs-TyrRS insertions are in orange. Motif 1 in Pf (PF11_0181 - HLGN and Mal8p1.125 - HIAQ) and Hs (2PID - HVGH and 1N3L - HVAY) TyrRSs has been encircled red whereas Motif 2 in Pf (PF11_0181 - KLGKS and Mal8p1.125 - KMSKS) and Hs (2PID - KYSKS and 1N3L - KMSSS) is encircled green.
(c) Snapshot of the active sites of Pf and Hs TyrRSs (superimposed) structures. Non-conserved active site residues colored violet are encircled.Bhatt et al. BMC Genomics 2009 10:644 doi:10.1186/1471-2164-10-644





























