Embed Size (px)
Citation preview

1
EVOLUTION OF THE GENETIC CONTROL OF LEAF DEVELOPMENT WITH AN EMPHASIS ON CARNIVOROUS PITCHER PLANTS
By
NICHOLAS WILLIAM MILES
A DISSERTATION PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY
UNIVERSITY OF FLORIDA
2013

2
© 2013 Nicholas William Miles

3
To those who believed in me, especially my Mom and Dad, Heather-Rose, and Susan Spaulding

4
ACKNOWLEDGMENTS
I would like to thank my advisors, Pam and Doug Soltis, for their tireless work
and support. This dissertation is testament to the saint-like patience they have had with
me over the years, and especially this year. I would also like to thank my committee
members, Drs. David Oppenheimer and Mark Settles, for the integral part they have
played in the forming of my ideas. Both of them and their labs have shared resources
that I cannot thank them for enough. Drs. Tsukaya and Yamaguchi, who showed
genuine rectitude as hosts to me in a foreign country and enabled me to collect data
that I had strived to do for 6 years, I am forever indebted to. The members of the Soltis
Lab, who I now view in likeness of family members, I will remember for the rest of my
life with great sentiment.

5
TABLE OF CONTENTS page
ACKNOWLEDGMENTS .................................................................................................. 4
LIST OF TABLES ............................................................................................................ 7
LIST OF FIGURES .......................................................................................................... 8
ABSTRACT ................................................................................................................... 11
CHAPTER
1 INTRODUCTION .................................................................................................... 12
Angiosperm Leaves as a System for the Study of Evolutionary Development (Evo-Devo) .......................................................................................................... 12
Nucleotide Sequencing Technology Leafs Out ....................................................... 15 Evo-Devo Studies of Carnivorous Pitcher Plants .................................................... 16
2 NEO- AND SUB-FUNCTIONALIZATION OF THE PLANT BODY PATTERNING GENE CLASS III HD-ZIP CORRELATES WITH LAND PLANT EVOLUTION ........ 19
Introductory Remarks.............................................................................................. 19
Methods .................................................................................................................. 24 Sequence Retrieval .......................................................................................... 24
Alignment ......................................................................................................... 25 Phylogenetic Analyses ..................................................................................... 25
Results .................................................................................................................... 26
BLAST results .................................................................................................. 26 Phylogenetic analyses ...................................................................................... 26
Discussion .............................................................................................................. 29
3 GENOMIC COEVOLUTION OF HD-ZIP III RELATED PATHWAY GENES ........... 46
Introductory Remarks.............................................................................................. 46
Methods .................................................................................................................. 51 Sequence retrieval ........................................................................................... 51 Alignment ......................................................................................................... 53
Phylogenetic analyses ...................................................................................... 53
Arabidopsis microarray expression .................................................................. 53 Results .................................................................................................................... 54
Paralog numbers .............................................................................................. 54 Pathway proportionality .................................................................................... 54 Lineage evolution ............................................................................................. 55
Discussion .............................................................................................................. 56

6
4 LEAF EVO-DEVO OF THE OF CARNIVOROUS PITCHER PLANT, CEPHALOTUS FOLLICULARIS (CEPHALOTACEAE) .......................................... 77
Introductory Remarks.............................................................................................. 77
Methods .................................................................................................................. 80 RNA extraction and gene amplification ............................................................ 80 Sequence alignment and phylogenetic analysis ............................................... 81 In situ hybridization ........................................................................................... 81
Probe construction ..................................................................................... 81
Tissue fixation ............................................................................................ 82 Hybridization .............................................................................................. 82
Results .................................................................................................................... 83 Discussion .............................................................................................................. 83
5 CONCLUSION ........................................................................................................ 96
APPENDIX
A FURTHER CARNIVOROUS PLANT EXPERIMENTS ............................................ 99
AS2 ......................................................................................................................... 99
VIGS Development ............................................................................................... 100
LIST OF REFERENCES ............................................................................................. 107
BIOGRAPHICAL SKETCH .......................................................................................... 116

7
LIST OF TABLES Table page 2-1 GenBank accession numbers of sequences from previous studies of Floyd
et al. (2006) and Prigge and Clark (2006) .......................................................... 33
2-2 Sampled Gymnosperm Taxa from 1KP Data.................................................... 34
2-3 Sampled Monilophyte taxa from 1KP data ....................................................... 35
3-1 Number of paralogs of the 4 gene families in the 6 sampled angiosperm genomes. ............................................................................................................ 60
3-2 Proportions of the number of paralogs for each of the 4 gene families in the genomes of the 6 sampled angiosperm genomes. ............................................. 61
3- 3 Places in the genomes of the sampled angiosperm genomes that contain the tandem precursor miR166 sequences. ......................................................... 61
4-1 Nucleotide sequence of primers used to amplify HistoneH4 and YABBY genes in Cephalotus follicularis. ......................................................................... 86

8
LIST OF FIGURES Figure page 1-1 Genetic model controlling leaf development and polarity in angiosperms.. ........ 18
2- 1 Evolution of HD-ZIP III and ZPR in land plants.. ................................................. 36
2-2 Protein domain structure of HD-ZIP III and ZPR................................................. 38
2-3 Phylogenetic tree of the topology found to have the highest likelihood by RAxML for sampled vascular plant HD-ZIP III sequences. ................................. 39
2-4 Subtree of HD-ZIP III vascular plant phylogeny containing lycophtye and PaleoHDZ3 clades. ............................................................................................. 40
2-5 Subtree HD-ZIP III vascular plant phylogeny containing gymnosperm ZPR clade. .................................................................................................................. 42
2-6 Subtree HD-ZIP III vascular plant phylogeny containing monilophyte RBVC8 as well as gymnosperm and angiosperm C8 clades. .......................................... 43
2-7 Gymnosperm RBVC8 as well as gymnosperm and angiosperm RBV subtree of of HD-ZIP III vascular plant phylogeny. .......................................................... 44
2-8 Alignment of gymnosperm ZPR sequences and the Homeodomain-Leucine-Zipper Region of HD-ZIP III. ............................................................................... 45
3-1 Phylogeny of major Angiosperm lineages and eudicot orders with the placement of sampled taxa. ................................................................................ 62
3-2 Phylogeny and ancestral genome evolution of sampled angiosperms and the number of copies of each of the sampled genes in their genome....................... 63
3-3 Bar graphs comparing the number of copies of HD-ZIP III with the number of copies of the genes that regulate it in the genomes of the six sampled species. .............................................................................................................. 64
3-4 Gene trees and species trees of the four leaf polarity genes in the six angiosperm genomes. ........................................................................................ 66
3-5 Phylogram of sampled HD-ZIP III genes. ........................................................... 67
3-6 Unrooted phylogeny of sampled GARP and KANADI genes from green plants and a protozoan. ...................................................................................... 68
3-7 Unrooted phylogram of sampled KANADI genes. ............................................... 69
3-8 Alignment of miR166 from the genomes of Amborella, Aquilegia, Vitis, and Arabidopsis. ........................................................................................................ 70

9
3-9 Syntenic regions of the Amborella and Aquilegia genomes that contain a discovered miR166 copy. ................................................................................... 71
3-10 Unrooted phylogram of sampled ZPR genes. ..................................................... 72
3-11 Expression from 454 read counts from Ancestral Angiosperm project (http://ancangio.uga.edu/) of Amborella HD-ZIP III paralogs. ............................. 73
3-12 Expression from microarray intensities of Arabidopsis HD-ZIP III paralogs. ....... 74
3-13 Expression from microarray intensities of Arabidopsis ZPR paralogs. ............... 75
3-14 Expression from microarray intensities of Arabidopsis KANADI paralogs. ......... 76
4-1 Front view of Cephalotus follicularis pitcher and basal rosette of flat leaves. ..... 88
4- 2 Photographs of Cephalotus pitcher, flat leaves, intermediate leaf, inflorescence, and flowers. ................................................................................. 89
4-3 Longitudinal section drawing of Cephalotus mature pitcher and shoot tip. ......... 90
4-4 Alignment of Cephalotus and Arabidopsis HistoneH4 amino acid sequences. ... 91
4-5 Phylogram of angiosperm and isolated carnivorous pitcher plant YABBY paralogs. ............................................................................................................. 92
4-6 In-situ hybridization of HistoneH4 in developing Cephalotus pitcher. ................. 93
4-7 In-situ hybrizations of CephFIL in serial longitudinal sections of a developing Cephalotus pitcher. ............................................................................................. 94
4-8 Stipules in the seedling of Geissois pruinosa (Cunoniaceae). ............................ 95
4-9 Expression of TAS3 in the vasculature, adaxial domain of developing leaves, and stipules of Arabidopsis thaliana. .................................................................. 95
A-1 Phylogeny of Arabidopsis AS2/LOB paralogs and AS2/LOB BLAST results form 1KP (www.onekp.com) transcripts for Nepenthes alata (abbreviated Nep), Cunonia capensis (Cun), and Cephalotus follicularis (Cep). ................... 101
A-2 Longitudinal section of in situ hybridization of Nepenthes alata AS2. ............... 102
A-3 Longitudinal sections of fluorescent In situ hybridization of Nephenthes alata AS2. .................................................................................................................. 103
A-4 Longitudinal sections of fluorescent in situ hybridization of Cephalotus AS2. .. 104
A-5 Longitudinal sections of fluorescent in situ hybridization of Cephalotus AS2. .. 105

10
A-6 Electrophoresis gel visualization of TRV1 and TRV2 PCR’s of Agrobacterium infected plants. ................................................................................................. 106

11
Abstract of Dissertation Presented to the Graduate School of the University of Florida in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy
EVOLUTION OF THE GENETIC CONTROL OF LEAF DEVELOPMENT WITH AN
EMPHASIS ON CARNIVOROUS PITCHER PLANTS By
Nicholas William Miles
August 2013
Chair: Pamela S. Soltis Cochair: Douglas E. Soltis Major: Botany
In this dissertation I explore, with various methods, the evolution of the genes
that control the development of leaves. First, the evolutionary history of the adaxial
identity gene Class III Homeodomain-Leucine Zipper (HD-ZIP III) and the microProtein
LITTLE ZIPPER (ZPR) is reconstructed, and ZPR is shown to be derived from HD-ZIP
III through a duplication event prior to the origin of extant seed plants. Second, a
genome-scale sampling of diverse angiosperms for four genes whose interactions
control leaf polarity is conducted. This approach allows predictions of
subfunctionalization of paralogs and tests a hypothesis of stoichiometry of genes in the
genome. The carnivorous plant, Utricularia gibba, is also sampled and shows extreme
amounts of duplications and losses of gene lineages. Third, the expression of an
abaxial identity gene, YABBY, is characterized in the carnivorous pitcher plant
Cephalotus follicularis and informs on the evolution of the leaf into a pitcher structure.

12
CHAPTER 1 INTRODUCTION
Angiosperm Leaves as a System for the Study of Evolutionary Development (Evo-Devo)
“Alles ist Blatt.”
Johann Wolfgang von Goethe (1790)
As one of the greatest thinkers of the 18th Century, Goethe had an undeniably
large effect on the field of evolutionary botany, and his famous quotation, translated as
“All is leaf”, highlights the special place that the leaf has in the evolution and
development of plants. Many of the adaptations in angiosperms that are thought to be
responsible for them being the most specious group of plants living, evolved as
developmental changes to leaves. These include the evolutionary transitions to most of
the reproductive organs, i.e., sepals, petals, stamens, carpels, the ovule, and most fruits
(reviewed in Mathews and Kramer, 2012).
Leaves also have taken on a multitude of forms in land plants, as they have
adapted to the biotic and abiotic interactions in the environment. As one of the central
questions of biology is how variation arises, leaves are especially suited to answer this
question because of the vast amounts of variation between clades, within clades, and
even within an individual.
Some of the most amazing forms that never cease to produce wonder in humans
are the extreme shapes adapted for interactions with animals. For instance, some
plants modify their leaves into pouches that serve as nests for ants, as seen in the
bullate leaves of Dischidia (Apocynaceae) or the modified stipules of some Acacia
species (Fabaceae). In Acacia, the stipules of some species have been transformed
into sharp spines to protect the plant from herbivores. Modified leaves in the form of

13
spines are also highly adapted in the succulent, arid-adapted Cactaceae and
Euphorbiaceae. The cup-shaped leaves of Ficus benghalinensis var. krishnae seem to
have the unique function of producing an animal interaction that has been adapted as a
religious symbol in the Hare Krishna religion (Juniper et al., 1989). And finally, in
carnivorous plants, all of the trapping mechanisms involve morphological and structural
changes to leaves (Juniper et al., 1989).
All of the above-mentioned instances of extreme structural and functional leaf
changes were studied in great detail by plant morphologists, like Goethe, and these
aberrations have served as interesting challenges to the models of plant morphology.
Today, the field of Evolutionary-Development, or “Evo-Devo”, typically applies a model
of development that involves the interaction of genes to developmental aberrations to
investigate how changes in those genes account for the changes in development and
evolutionary transitions in morphology. In the case of bizarre leaf shapes, many of the
changes involve a change from bifacial to unifacial polarity. The field of developmental
genetics has produced a model of bifacial leaf development (Figure 1-1), and applying
that model to the origin and development of various leaf forms was the main aim of this
dissertation work.
The model of leaf polarity consists of genes that have mutually exclusive adaxial
and abaxial expression domains. The initial cue for the orientation of these domains
comes from a signal in the meristem so that the adaxial domain is positioned next to the
meristem and the abaxial away. The juxtaposition of the expression domains in the leaf
primordia is essential for defining a plane a growth to create a flat leaf from the initially
radial primordia. The model is also predicated on the idea that the expression domains

14
of the genes in a bifacial leaf define the anatomical differentiation as well as the
morphological differentiation. When a gene of the model is mutated the disruption of the
repulsing behavior of the network usually leads to the domination of one domain over
the other and the subsequent leaf is radial in morphology with a uniform distribution of
the cell types that are characteristic of the dominate domain. Therefore, because
carnivorous pitcher plants have unifacial leaves without a flat growth plane, it is
expected that the leaf polarity model is acting differently during their development.
Plant leaves also show morphological and anatomical polarity in the proximal-
distal axis of the leaf. Most leaves have proceeding from the proximal to distal end
base, petiole, and lamina zones. The distance from the shoot apical meristem defines
this axis of polarity. One of the molecular signals for positioning the proximal-distal
zones is the class I knotted1-like homeobox (KNOX) gene (Freeling, 1992). Gain of
function mutations in KNOX cause changes to the proximal-distal placement of leaf
characteristics, like the leaf ligule of the maize (Zea mays) (Freeling, 1992). The gene
BLADE ON PETIOLE (BOP) is also known to control the proximal-distal axil, specifically
the boundary between the lamina and petiole (Ha et al., 2007). There is evidence that
the proximal-distal and adaxial-abaxial genes interact so that the two axes are
interlinked genetically (Ha et al., 2007). Carnivorous pitcher plants also show distinct
developmental changes in the proximal-distal axes. Nepenthaceae is interpreted
morphologically to have a very large base region that is expanded for light capture, a
twinning tendril-like petiole, and a pitcher lamina (Franck, 1976). The lamina region of
Sarraceniaceae can be very elongated to make a tall pitcher (Franck, 1976).

15
Nucleotide Sequencing Technology Leafs Out
Recent advances in nucleotide sequencing technology has allowed an
unprecedented amount of sequence data to be collected from diverse plant groups, with
most of the data being freely available to the public. Understanding the homology and
sequence evolution of the genes involved in the model of bifacial leaf development is a
fundamental first step in the study of leaf Evo-Devo and is the motivation for Chapters
two and three.
Chapter two describes the evolutionary history of two plant organ polarity genes,
Class III Homeodomain Leucine-Zipper (HD-ZIP III) and LITTLE ZIPPER (ZPR), in land
plants. These two genes have an interesting feedback regulatory interaction, in that,
HD-ZIP III is a transcription factor that enhances the transcription of down-stream
adaxial development genes, but also enhances ZPR, a microprotein whose protein
binds to HD-ZIP III proteins and inactivates their DNA bind ability (Wenkel et al., 2007).
My study provides new insights into the evolution of these genes in gymnosperm and
monilophytes, where the number of copies and history of the two genes is not well
understood because of the lack of fully sequenced genomes from taxa in those groups.
The presence or absence of ZPR outside of the Angiosperms was not known prior to
this study. By fully sampling from all major groups of land plants with data from the One
Thousand Plant Transcriptomes Project (1KP; www.onekp.com), this study was able to
better understand the make up of the main clades of these genes and the relations
among them. Perhaps, most importantly, this study was able to provide evidence that
ZPR evolved from HD-ZIP III just prior to the divergence of gymnosperms and
angiosperms.

16
Chapter three describes the evolution of HD-ZIP III and three of its regulatory
genes in the sequenced genomes of six angiosperm species, including the newly
available draft of the first carnivorous plant genome. My description of three regulatory
genes of HD-ZIP III includes a post-transcriptional microRNA, a post-translational micro-
Protein, and a repulsing transcription factor. With this sampling of a network of
interacting genes, I set out to find out if the network is behaving stoichiometrically by
quantifying the number of coding copies in the genomes and their proportions to the
gene they regulate, HD-ZIP III. My findings show that, indeed, they do have similar
proportionalities in all six sampled genomes, even in the fractionalized genome of the
carnivorous plant. My findings also show that the process of fractionalization in the
carnivorous plant genome has removed major lineages of the genes, which might be
related for the major loss of roots in the plant. The possibility of the lost lineages being
specialized for root development is further corroborated by the quantified expression of
those lineages in Arabidopsis and Amborella.
Evo-Devo Studies of Carnivorous Pitcher Plants
Chapter four describes at the expression of a paralog of the abaxial leaf identity
gene, YABBY, in the carnivorous pitcher plant Cephalotus follicularis. This investigation
relies on laborious molecular techniques for assessing the exact areas of expression in
the leaf. However, this data allows for the comparisons with the expression domain in
bifacial leaves and other non-carnivorous unifacial leaves. The development of
carnivorous pitcher plants has been most recently understood as being homologous
with the development of the unifacial peltate leaves (Franck, 1976) and expression of
YABBY has been previously described in a peltate leaf (Gleissberg, 2005). Therefore, I
test the hypothesis that the carnivorous pitcher plant would have a similar expression

17
domain of the homologous YABBY paralog as the peltate leaf. The findings from this
chapter allow for the tying together of morphological studies two centuries old with
genetic models of the past two decades.
Appendix A of this dissertation is a description of other laboratory experiments
done with carnivorous plants to try to better understand how the leaf polarity genes are
working in the three instances of convergent carnivorous pitcher plant evolution. Some
of these experiments include describing the expression domain of the adaxial leaf
identity gene, ASSYMENTRIC LEAF2 (AS2) in two of the pitcher plant families,
Cephalotaceae and Nepenthaceae. Other experiments tried to develop a protocol for
knocking down the expression of target genes in carnivorous plants. Knocking down the
expression of leaf polarity genes was the ultimate goal because it would have provided
better evidence for the role of leaf polarity genes in pitcher plants.
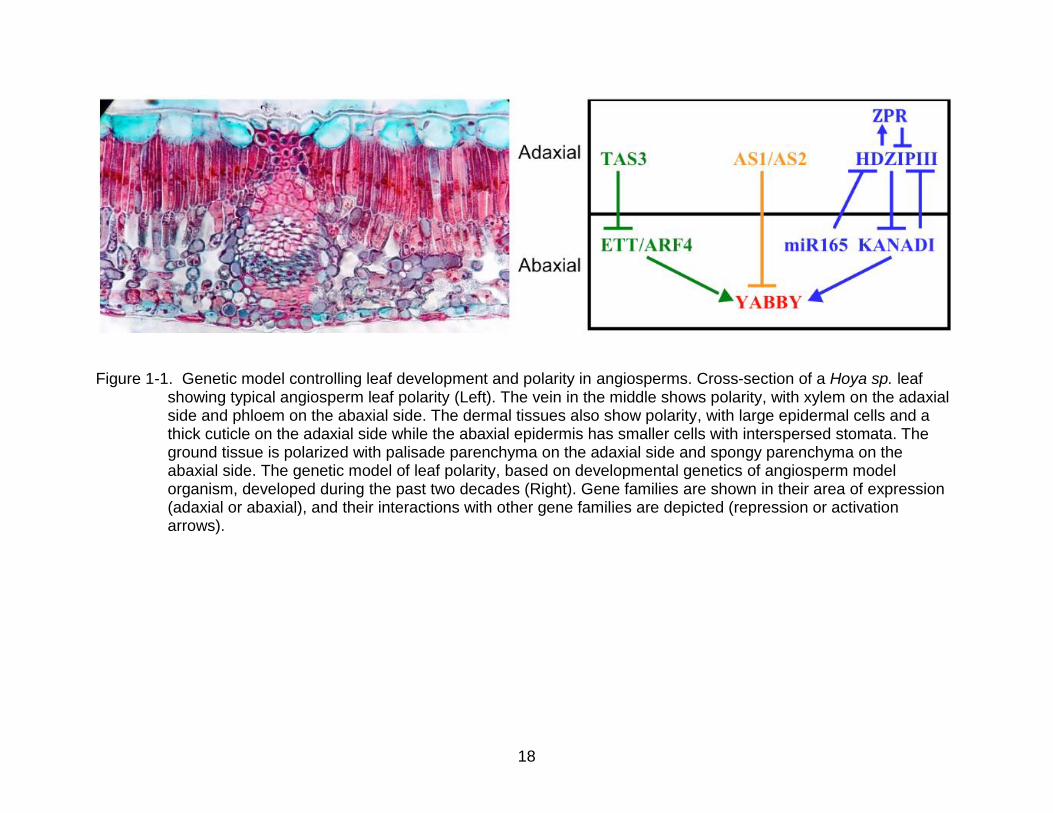
18
Figure 1-1. Genetic model controlling leaf development and polarity in angiosperms. Cross-section of a Hoya sp. leaf showing typical angiosperm leaf polarity (Left). The vein in the middle shows polarity, with xylem on the adaxial side and phloem on the abaxial side. The dermal tissues also show polarity, with large epidermal cells and a thick cuticle on the adaxial side while the abaxial epidermis has smaller cells with interspersed stomata. The ground tissue is polarized with palisade parenchyma on the adaxial side and spongy parenchyma on the abaxial side. The genetic model of leaf polarity, based on developmental genetics of angiosperm model organism, developed during the past two decades (Right). Gene families are shown in their area of expression (adaxial or abaxial), and their interactions with other gene families are depicted (repression or activation arrows).

19
CHAPTER 2 NEO- AND SUB-FUNCTIONALIZATION OF THE PLANT BODY PATTERNING GENE
CLASS III HD-ZIP CORRELATES WITH LAND PLANT EVOLUTION
Introductory Remarks
Class III Homeodomain-Leucine Zipper (HD-ZIP III) is a plant transcription factor
that plays an important role in patterning much of the plant body, including embryos,
vascular cambium, lateral organ polarity, and apical meristem maintenance. These
parts of the plant body have undergone significant adaptations during the evolution of
land plants (Embryophyta), producing characteristics of ecological and physiological
importance such as plant height, wood, seeds, and leaves. These traits also play
integral roles as the targets of agricultural crop improvement. Therefore, understanding
the evolution and regulation of HD-ZIP III is closely connected to both fundamental
knowledge about plant development and evolution, as well as the use of seeds, grains,
wood products, and lateral organs such as fruits and leaves by humans.
Specific actions of HD-ZIP III in plant organs include a role in patterning of early
development from zygote to young embryo. Normally developing embryos show a
polarized expression pattern of HD-ZIP III, while mutants of the gene fail to develop
important early body plan characteristics properly (Bowman and Floyd, 2008). In a
certain Arabidopsis thaliana HD-ZIP III mutant, the first embryonic seed leaves,
cotyledons, fail to develop (Prigge et al., 2005). This is of economic interest as
cotyledons are important products from many agricultural plants, such as seeds of the
legume family, Fabaceae (e.g., soybean and peanut). In peanut, the HD-ZIP III
regulator, miR166, is normally expressed in the shoot apical meristem, root, and leaves,
but lowly expressed in the cotyledons (Chi et al., 2011), providing evidence that HD-ZIP

20
III may have higher expression in the large peanut cotyledon and may contributes to its
size.
Another region of the embryo in which HD-ZIP III is expressed, in addition to the
adaxial side of the cotyledons, is the area that develops into the shoot apical meristem
(SAM) (Prigge et al., 2005). The SAM is a region of undifferentiated stem cells that give
rise to all the aboveground organs of the plant, including the stem and lateral organs.
The SAM shows increasing size and organization from the earliest land plants to
angiosperms (Floyd and Bowman, 2007). HD-ZIP III gene family members control SAM
patterning in the angiosperm model Arabidopsis thaliana, with mutations generating
plants with reduced or completely absent SAMs (Byrne, 2006).
The vascular cambium is the area of undifferentiated cells that undergo
secondary growth to produce phloem and xylem, the latter maturing into wood. True
secondary growth is only found in members of Embryophyta. HD-ZIP III has been
shown to be involved in vascular cambium growth in the rosid Arabidopsis thaliana
(Zhong and Ye, 2007), the rosid timber tree Populus trichocarpa (Robischon et al.,
2011; Zhu et al., 2013), the asterid Zinnia elegans (Ohashi-Ito and Fukuda, 2003), and
the conifer Picea glauca (Côte et al., 2010). Differential development of xylem over
phloem by the vascular cambium, and therefore overproduction of wood, has been
shown to be promoted by an HD-ZIP III paralog in Populus trichocarpa and is
seasonally regulated (Ko et al., 2006).
HD-ZIP III genes also play an important part in controlling the shape and
patterning of leaves and other lateral organs by patterning the adaxial side of those
developing organs. The horizontal growth of the leaf lamina is attributed to the

21
juxtaposition of the adaxial and abaxial expression domains in developing leaf
primordia. Triple mutants of the HD-ZIP III paralogs phabulosa, phavulota, and revoluta
generate Arabidopsis plants with leaf laminar growth abnormalities. Strong mutant
plants produce symmetrical needle-like leaves, and weak mutants have a rounded
lamina described as a trumpet or bell (McConnell et al., 2001).
The important role of HD-ZIP III in leaf development has made it a popular choice
for studies on the evolution of this highly adaptive organ of vascular plants. The main
photosynthetic, light-capturing, and respiratory organs, leaves are also commonly co-
opted for novel ecological and physiological functions. In vascular plants, leaves are
thought to have evolved multiple times (Figure 2-1). Lycophytes first evolved
rudimentary microphylls more than 400 million years ago (Kenrick and Crane, 1997).
Later, larger megaphylls evolved independently multiple times in the monilophytes and
once in seed plants. These lateral organs have also been modified into reproductive
structures, including floral organs (sepals, petals, stamens, and carpels) in angiosperms
(Mathews and Kramer, 2012). The question of homology and developmental
mechanisms in the variety of lateral organs in land plants has been debated for some
time (Kenrick and Crane, 1997; Kaplan, 2001). Molecular tools and data can be used
for interpreting the homology and evolution of lateral organs, facets of these organs that
have been hard to interpret based on the morphology of fossil and extant species
(Boyce and Knoll, 2002).
To understand the evolution of plant form, various studies have been conducted
on the evolution of molecular developmental mechanisms, with the evolution of leaf
developmental mechanisms widely investigated (reviewed in Yamaguchi et al., 2012).

22
The genome sequence of the lycophyte Selaginella moellendorffii and developmental
genetic data for the angiosperm model Arabidopsis thaliana have supplied the most
information about the genetic differences between the disparate types of leaves. In
general, a genome-wide comparison has revealed a general trend of increasing number
of gene families, with 1350 gene families being gained and 442 gene families lost during
the evolution between a hypothesized basal vascular plant gene set to the hypothesized
basal angiosperm gene set (Banks et al., 2011). Focusing just on the gene families
known to be involved in lateral organ initiation and patterning, Floyd and Bowman
(2007) found that only one of the 20 gene families they investigated was missing from
the Selaginella genome, but there was a general increase in the number of gene family
members between Selaginella and angiosperms. The patterning controlled by the
interplay of two of those gene families, KNOX and ASYMMETRIC LEAVES1/ROUGH
SHEATH2/PHANTASTICA (ARP), is similar between Selaginella and Arabidopsis,
indicating a possible independent recruitment of the same pathway for similar purposes
(Harrison et al., 2005). HD-ZIP III has also undergone independent diversification and
subfunctionalization of paralogs to pattern microphylls in Selaginella and megaphylls in
seed plants (Floyd et al., 2006). The micro-RNA-mediated post-transcriptional
regulation of HD-ZIP III was also found to be a mechanism of patterning that is shared
across all green plants (Floyd and Bowman, 2004).
HD-ZIP III proteins act as transcription factors whose patterns of expression
dictate the localized expression of downstream genes that promote the specific cell
growth shape and differentiation of the tissue type. The protein regulates downstream
genes either as a homo- or heterodimer (Sessa et al., 1993). There are four known

23
domains in HD-ZIP III proteins (Figure 2-2). Near the N terminus is the homeodomain, a
helix-loop-helix motif that binds to the DNA strand of the gene it is targeting for
regulation. Directly next to the homeodomain is the leucine-zipper domain where the
two HD-ZIP III proteins bind together to make a dimer (Staudt and Wenkel, 2011). The
START (STeroidogenic Acute Regulatory protein–related lipid Transfer) domain is a
putative lipid-binding site in the middle of the protein (Mukherjee et al., 2009). Next to
that is the HD-SAD domain, which is known to be associated with the START domain of
HD-ZIP III and HD-ZIP IV, but its activity is unknown (Mukherjee and Burglin, 2006). At
the C terminus is the MEKHLA domain, which is a PAS-like domain that regulates the
activity of the protein-protein dimer. It is the target of an unknown cellular signal that
binds to the domain and affects conformation of the HD-ZIP III peptide. This domain’s
evolution is curious as it is only found in cyanobacteria and chloroplast genomes and is
hypothesized to have fused to an HD-ZIP IV copy to produce the HD-ZIP III gene at
some time after the divergence of red algae from green plants but before the origin of
land plants (Mukherjee and Burglin, 2006; Magnani and Barton; 2011). Floyd et al.
(2006), in their sampling of charophycean algae, found one copy of HD-ZIP III in Chara
carollina (Charales) but no copies in Coleochaete scutata (Coleochaetales). Lack of
expression of the gene in the abaxial side is achieved through the post-transcriptional
degradation initiated by miR165/166. Mutational studies in Arabidopsis have shown that
mutations in the micro-RNA binding site cause a gain of function so that HD-ZIP III is
also expressed in the abaxial side and the subsequent organs are adaxialized.
Mutations in other areas of HD-ZIP III cause a loss of function, and the organ is
abaxialized (McConnell et al., 2001).

24
LITTLE ZIPPER (ZPR) is a small “microProtein” (Staudt and Wenkel, 2011), with
a conserved leucine-zipper region whose expression is activated by HD-ZIP III (Wenkel
et al, 2007). Once translated, the Leucine-Zipper region of the ZPR protein binds to the
Leucine-Zipper region of an HD-ZIP III protein, forming a non-functional heterodimer.
This negative feedback pathway has been found in Arabidopsis, and the paralogs have
been found in the genomes of rice (Oryza sativa) and corn (Zea mays), but little is
known about its evolution or presence in other plants (Staudt and Wenkel, 2011). The
evolutionary history of HD-ZIP III has been well studied in land plants (Prigge and Clark,
2006; Floyd et al., 2006), in the genomes of rice (Jain et al., 2008) and poplar (Populus
trichocarpa) (Hu et al., 2012), and in the context of the whole super-class of
Homeodomain genes in plants (Mukherjee et al., 2009). However, most studies of HD-
ZIP III have sparsely sampled the monilophyte clade and gymnosperms. With the recent
sequencing of a large sampling of green plant transcriptomes by the 1KP project (One
Thousand Plant Transcriptomes; www.onekp.com), we are now able to understand
better the evolution of HD-ZIP III and its regulator, ZPR. Here we investigate the
evolutionary history of HD-ZIP III and ZPR with extensive sampling across land plants,
and test the hypothesis that ZPR evolved from an HD-ZIP III paralog by truncation of
the gene down to the lone Leucine-Zipper region.
Methods
Sequence Retrieval
Nucleotide sequences from previous studies of HD-ZIP III (Prigge and Clark,
2006; Floyd et al., 2006) were downloaded from GenBank
(http://www.ncbi.nlm.nih.gov/genbank), and ZPR sequences were downloaded from The
Arabidopsis Information Resource (www.arabidopsis.org; Lamesch et al., 2011) (Table

25
2-1). Those nucleotide sequences were translated to amino acid sequences and used
as a query in tBLASTn searches of the Illumina-generated transcriptome sequence data
produced by the 1KP (One Thousand Plant Transcriptomes; www.onekp.com) project
for the taxa listed (Table 2-2; Table 2-3). TBLASTn searches were also performed on
the EST assemblies of Zamia vazquezii from the Ancestral Angiosperm Genome
Project (www.ancangio.uga.edu) and the assembled genome of Amborella trichopoda
(www.amborella.org).
Alignment
Previously published sequences and large 1KP BLAST hits with characteristic
HD-ZIP III motifs were translated and included in the final alignment using MAFFT
(Katoh et al., 2002) in Geneious version 5.6.5 (Biomatters, www.geneious.com). The
alignments were subsequently refined manually. The leucine-zipper regions of the ZPR
genes were aligned with leucine-zipper regions of the HD-ZIP III genes.
Phylogenetic Analyses
The amino acid alignment was phylogenetically analyzed using RAxML (RAxML-
VI-HPC) (Stamatakis, 2006) with the GTR + I + Gamma model of amino acid sequence
evolution. The topology with the highest maximum likelihood score was generated, and
support for that topology was assessed by 1000 bootstrap replicate searches using the
thorough bootstrap algorithm.
Amino acid sequences were used instead of nucleotide to account for the long
evolutionary distance of the sampled taxa. Amino acid data can be favored because the
20 characters of the amino acid data can code for more evolutionary information per site
than the 4-letter code of nucleotide data and amino acid data have less compositional
bias than DNA (Brocchieri, 2001). Past studies of HD-ZIP III evolution compared the

26
phylogenetic results of the same data represented in amino acid characters and
nucleotide characters and found few discrepancies (Prigge and Clark, 2006).
Results
BLAST results
Searches of the HD-ZIP III genes found putative copies in the transcriptomes of
the sampled lycophytes, monilophytes, and gymnosperms. The Amborella genome was
also queried with HD-ZIP III using tBLASTn, and three areas of the genome showed
characteristic HD-ZIP III motifs and intron-exon structure. TBLASTn searches with ZPR
found two copies in the genome of Amborella and generally one putative copy in each
of the gymnosperm taxa. However, no suitable hits were found in the transcriptomes of
the monilophytes, and the closest hits were of HD-ZIP III monilophyte transcripts. The
reverse searches with gymnosperm ZPR genes as a query of GenBank found only
angiosperm HD-ZIP III and ZPR genes. The leucine-zipper regions of the HD-ZIP III and
ZPR genes showed distinct motifs differing from the leucine-zipper regions of other HD-
ZIP III genes (Figure 2-4).
Phylogenetic analyses
Phylogenetic analyses of the combined data of Floyd et al. (2006) and Prigge
and Clark (2006) revealed that there is another lineage of HD-ZIP III in plants that was
not evident in either of the individual data sets. This new lineage is present in
monilophytes and gymnosperms but does not appear to exist in angiosperms (Figure 2-
1, Figure 2-3). This previously undetected lineage is sister to another exclusively
monilophyte + gymnosperm lineage termed “PaleoHDZ3” by Prigge and Clark (2006).
This lineage appears to have arisen from a duplication that occurred prior to the origin
of monilophytes but after the divergence of lycophytes from the remaining

27
tracheophytes (and thus is present in extant gymnosperms). This means that two
separate duplications of this gene occurred prior to the divergence of monilophytes and
seed plants. No HD-ZIP III paralogs that belonged to the “PaleoHDZ3” lineages were
found in angiosperms, making it appear that both copies were lost after the divergence
of angiosperms from extant gymnosperms but prior to the diversification of extant
angiosperms.
Each of the three HD-ZIP III sequences found in the genome of Amborella
appears in the phylogeny as sister to one of the three main lineages of HD-ZIP III found
in other angiosperms, including a “C8”, “REV”, and “PHX” copy (following the
nomenclature of Prigge and Clark (2006)). This means that of the five copies present in
the gymnosperms, the ancestor of angiosperms retained only two copies and that a
duplication of one of those two copies prior to the origin of the angiosperms produced
the “REV” and “PHX” lineages. The fact that no PaleoHDZ3 sequences could be found
in the genome of Amborella allows for a better understanding of the timing of the loss of
PaleoHDZ3 lineages in angiosperms. This loss seems to have taken place between 150
and 300 million years ago, oddly enough, when a major paleopolyploid event is believed
to have taken place before angiosperm diversification (Jiao et al., 2011). However, our
phylogenetic results show that a minimum of three duplication events happened prior to
the diversification of the seed plants, lending support to the presence of the whole-
genome duplication event before the diversification of the clade.
The inclusion of 110 gymnosperm HD-ZIP III sequences, 48 monilophyte HD-ZIP
III sequences, and 27 gymnosperm ZPR sequences further clarified and supported the
presence of an unknown monilophyte + seed plant lineage (Figure 2- 3). With this larger

28
sampling it is clear that the clade denoted as PaleoHDZ3 by Prigge and Clark (2006)
actuality represents two separate gene lineages in vascular plants, denoted here as
PaleoHDZ3a and PaleoHDZ3b. The monophyly of both the PaleoHDZIP3a and
PaleoHDZIP3b clades was supported by 82% and 92% bootstrap support, respectively.
On a finer scale, a paleo-duplication event in the PaleoHDZ3b lineage is evident in
leptosporangiate ferns. The Angiopteris evecta PaleoHDZ3b sequence is sister to the
two lineages of leptosporangiate PaleoHDZIP3b sequences with 94% bootstrap
support. This indicates the duplication happened after the divergence of
leptosporangiate and eusporangiate ferns and is consistent with a hypothesized whole-
genome duplication event prior to the diversification of leptosporangiate ferns (Barker,
2013).
The backbone of the “RBVC8” seed plant clade was poorly resolved (Figure 2-3)
with the Gymno-RBCVC8 clade sister to the Gymno-RBV clade but with only 21%
bootstrap support. The relationship between Gymno-RBVC8, Gymno-C8, and Gymno-
RBV has been a difficult relationship to resolve with high support in our study, as well as
in previous studies (Prigge and Clark, 2006; Floyd et al., 2006), possibly because of the
number of and time since the duplication events. We have chosen to depict the
relationship in Figure 2-1 with Gymno-RBVC8 sister to the RBV + C8 clades because of
the low support for our inferred relationship and the precedence of the two previous
studies (Prigge and Clark, 2006; Floyd et al., 2006). However, both topologies, with
Gymno-C8 sister to RBVC8 and RBV, or with Gymno-RBVC8 sister to C8 and RBV,
require an equally parsimonious number of duplication and loss events to explain the
presence in angiosperms.

29
The ZPR sequences of gymnosperms and angiosperms were consistently
grouped together with >98% bootstrap support when included in the phylogenetic
analyses with the HD-ZIP III sequences, indicating that gymnosperm and angiosperm
ZPR arose as the result of a single evolutionary event. When the ZPR sequences were
aligned with the HD-ZIP III sequences, the ZPR sequences grouped consistently with
the paleoHDZ3 lineages. The gymnosperm sequences were easier to align to the HD-
ZIP III sequences presumably because the amino acid residues around the leucine-
zipper were more conserved than the angiosperm ZPR sequences. When only
gymnosperm ZPR genes were included in the ZPR-HD-ZIP III alignment, the ZPR
sequences grouped with the paleoHDZ3b sequences with 84% bootstrap support and
grouped with the gymno-paleoHDZ3b sequences with 70% bootstrap support. This is
evidence that ZPR evolved from the duplication of a gymno-paleoHDZIP3b sequence
and was subsequently passed on to angiosperms.
Discussion
In previous studies (Floyd et al., 2006; Prigge and Clark, 2006), the number of
HD-ZIP III lineages in some angiosperms, lycophytes, and mosses was known with a
high degree of certainty because of the genome sequences available for species from
each group. However, no genomes or genomic resources were available for any
monilophytes or gymnosperms. The utility of recent sequences obtained from the
transcriptome sequencing project, 1KP , was well illustrated here. Two main factors of
the project contributed to the successful use of the sequences they generated. First, the
1KP sampling was of considerable breadth, including many species from each of the
main lineages of gymnosperms and monilophytes. Second, for this study, the partial,
random sequencing of the transcriptome employed by 1KP, rather than generating

30
traditional Expressed Sequence Tags (ESTs), allowed for better sampling of the highly
conserved regions of the HD-ZIP III genes. ESTs, in contrast, capture mostly 3’-UTR
regions of the gene that are rapidly evolving and difficult to align in large taxonomic
samplings. The 1KP transcriptome data were therefore more useful for large-scale
phylogenetic studies, like this one, and made it easier to find homologs with tBLASTn
searches queried with homologs from other lineages.
The 1KP transcriptome data detected more paralogs than did the two previous
studies based on Sanger sequencing (Floyd et al., 2006; Prigge and Clark, 2006). Our
findings also differ from the EST sequence study of Côte et al. (2010), whose Neighbor-
Joining analyses differ in the description of the relationship of the “PaleoHDZIP3”
lineage to the other seed plant lineages. This discrepancy may be due to the greater
susceptibility of Neighbor-Joining methods than maximum likelihood to long-branch
attraction (Felsenstein, 1978), which could be expected in data sets that have limited
taxonomic sampling (Graybeal, 1998; Soltis and Soltis, 2004).
The truncation of an HD-ZIP III paralog to the ZPR gene is reminiscent of the
other lateral organ development gene family, AUXIN RESPONSE FACTOR (ARF),
which also functions as a dimer and has evolved a diversity of isoforms mainly by
truncation and alternative splicing (Finet et al., 2013). In addition, a similar gene
truncation event happened in the Homeodomain gene KNOX, where the truncation also
involved the loss of the homeodomain region but a conservation of the dimer-binding
domain as a way to regulate the activity of the KNOX genes (Magnani and Hake, 2008).
This study helps explain not only when ZPR evolved, but also the circumstances
that may have necessitated its evolution. As a regulator of HD-ZIP III, ZPR origin may

31
be related to the fact that there was an unprecedentedly large number of functional
copies in the genome when ZPR originated. Given that HD-ZIP III evolved somewhere
in multicellular green algae, additional copies of HD-ZIP III have accumulated in the
genomes of mosses, and still more in lycophytes, and monilophytes. These groups
evolved more specialized organs, and patterning of those organs became more
complex as they adapted to conditions on land; concomitantly, the number of copies of
HD-ZIP III increased with an increase in the complexity of function. For instance, there
is no known copy of HD-ZIP III in Coleochaetales, and there is only one copy of HD-ZIP
III in Chara coralline (Charales), which represents one of the first lineages to evolve
apical growth (Floyd et al., 2006). In mosses there may be multiple copies but only one
that was shared between the most recent common ancestor of mosses and vascular
plants, and there is only one copy reported in hornworts (Floyd et al., 2006). Lycophytes
have two copies of HD-ZIP III, and these are subfunctionalized into two distinct
expression domains in response to two major adaptations, vasculature and microphylls
(Floyd et al., 2006). In monilophytes, there are three distinct lineages of HD-ZIP III and
this increase in the number of paralogs looks to have happened at the time as the
evolution of megaphylls in the group. In gymnosperms, there is a large jump to six HD-
ZIP III lineages, and happens around the same time as the evolution of secondary
growth. The origin of the micro-protein ZPR in gymnosperms may have been
advantageous for controlling the more complicated subfunctionalized actions of HD-ZIP
III. In angiosperms, an evolutionary trend is observed with duplications of the micro-
protein and a decrease in the number of copies of the transcription factor, with three
HD-ZIP III lineages and two ZPR lineages in the early angiosperm Amborella

32
trichopoda. In Arabidopsis thaliana, the ratio is similar, with five HD-ZIP III lineages and
four ZPR lineages.
This study is also germane to discussions on the homology of lateral organs. The
data suggest that the microphylls of lycophytes and the megaphylls found in
euphyllophytes are evolutionarily very different (Kenrick and Crane, 1997; Kaplan,
2001). There appears to be only one copy of HD-ZIP III that is found in the most recent
common ancestor of lycophytes and euphyllophytes. The gene lineage in each of these
two clades of vascular plants has independently diversified and subfunctionalized and
interacted with other genes to produce a leaf organ. With regard to the homology of
megaphylls of monilophytes and seed plants, the data do not disagree with the
possibility that these two groups co-inherited their HD-ZIP III leaf developmental
mechanism. There is no major loss of HD-ZIP III copies prior to the origin of extant
gymnosperms or in the various monilophyte clades, making it highly likely that the most
recent common ancestor of euphyllophytes had a leaf organ that was inherited by both
lineages. However, the fact that seed plants evolved the ZPR gene and monilophytes
did not suggests that the lateral organ developmental tool kit in seed plants is more
specialized and that ZPR represents a mechanism of control that monilophytes did not
evolve.

33
Table 2-1. GenBank accession numbers of sequences from previous studies of Floyd et al. (2006) and Prigge and Clark (2006)
Species Sequence Name
GenBank Accession Number
Species Sequence Name
Sequence Name
Selaginella kraussiana SkHDZ31 DQ657196
Cycas revoluta CreHDZ32 DQ657218
Selaginella kraussiana SkHDZ32 DQ657197
Ginkgo biloba GbHDZ31 DQ657215
Selaginella moellendorffii SmHDZ31 DQ657198
Ginkgo biloba GbHDZ32 DQ657216
Selaginella moellendorffii SmHDZ32 DQ657199
Ginkgo biloba GbHDZ33 DQ657217
Selaginella moellendorffii SmHDZ32 DQ657197
Taxus globosa TgC3HDZ1 DQ385530
Ceratopteris richardii CrC3HDZ1 DQ385524
Taxus globosa TgC3HDZ2 DQ385531
Ceratopteris richardii CrC3HDZ1 DQ385524
Pinus taeda PtaHDZ31 DQ657210
Marsilea minuta MmHDZ31 DQ657207
Pinus taeda PtaHDZ32 DQ657211
Marsilea minuta MmHDZ32 DQ657209
Pinus taeda PtaHDZ33 DQ657212
Psilotum nudum PnC3HDZ1 DQ385521
Pinus taeda PtaHDZ34 DQ657213.
Psilotum nudum PnC3HDZ2 DQ385522
Pinus taeda PtaHDZ35 DQ657214
Psilotum nudum PnC3HDZ3 DQ385523
Pseudotsuga menziesii PmC3HDZ1 DQ385528
Ceratopteris richardii CriHDZ32 DQ657206
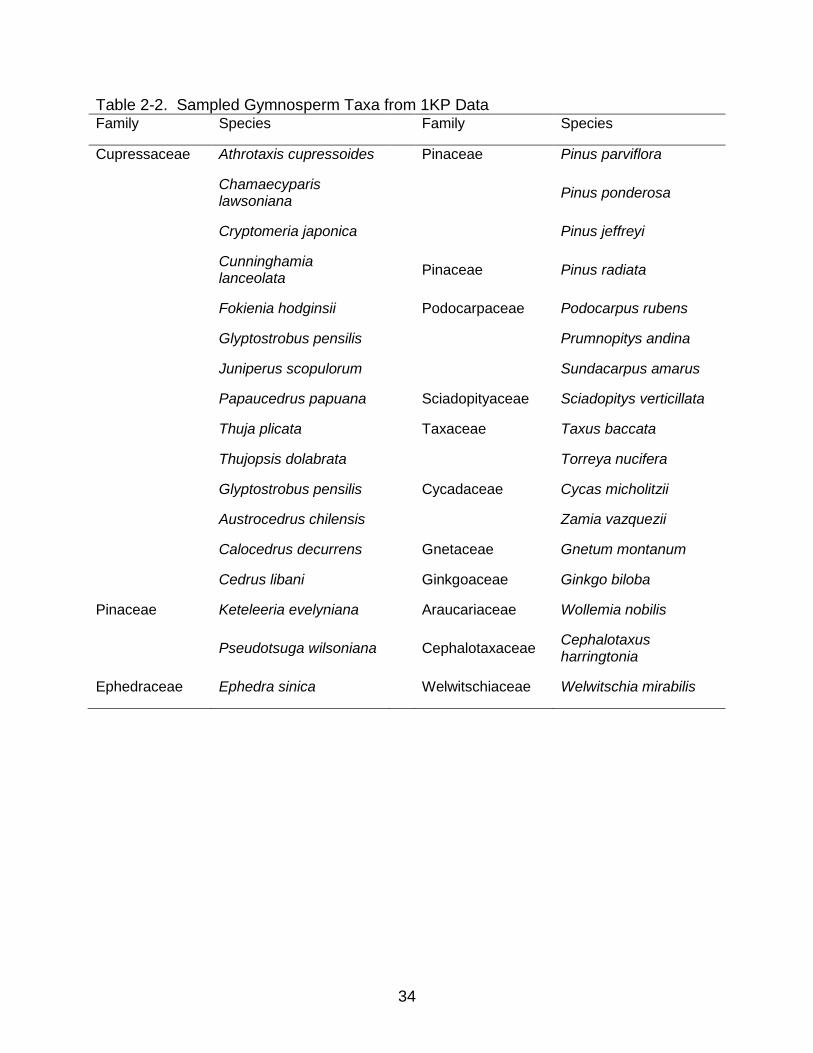
34
Table 2-2. Sampled Gymnosperm Taxa from 1KP Data Family Species Family Species
Cupressaceae Athrotaxis cupressoides Pinaceae Pinus parviflora
Chamaecyparis lawsoniana
Pinus ponderosa
Cryptomeria japonica Pinus jeffreyi
Cunninghamia lanceolata
Pinaceae Pinus radiata
Fokienia hodginsii Podocarpaceae Podocarpus rubens
Glyptostrobus pensilis Prumnopitys andina
Juniperus scopulorum Sundacarpus amarus
Papaucedrus papuana Sciadopityaceae Sciadopitys verticillata
Thuja plicata Taxaceae Taxus baccata
Thujopsis dolabrata Torreya nucifera
Glyptostrobus pensilis Cycadaceae Cycas micholitzii
Austrocedrus chilensis Zamia vazquezii
Calocedrus decurrens Gnetaceae Gnetum montanum
Cedrus libani Ginkgoaceae Ginkgo biloba
Pinaceae Keteleeria evelyniana Araucariaceae Wollemia nobilis
Pseudotsuga wilsoniana
Cephalotaxaceae Cephalotaxus harringtonia
Ephedraceae Ephedra sinica Welwitschiaceae Welwitschia mirabilis

35
Table 2-3. Sampled Monilophyte taxa from 1KP data
Family Species
Aspleniaceae Asplenium nidus
Asplenium platyneuron
Equisetaceae Equisetum diffusum
Marattiaceae Angiopteris evecta
Ophioglossaceae Ophioglossum vulgatum
Polypodiaceae Polypodium amorphum
Polypodium hesperium
Pteridaceae Adiantum aleuticum
Cheilanthes arizonica
Myriopteris eatonii
Woodsiaceae Cystopteris fragilis
Cystopteris protrusa
Gymnocarpium dryopteris
Woodsia scopulina

36
Figure 2- 1. Evolution of HD-ZIP III and ZPR in land plants. Black outline phylogeny represents a chronogram of land plant evolution (species tree) based on inferred dates of Jiao et al. (2011). Colored internal lines represent HD-ZIP III lineages (gene tree). Black arrows mark major land plant morphological adaptations. Green arrows mark currently accepted evolution of microphyll and megaphyll leaves. Starbursts represent gene duplication events.

37
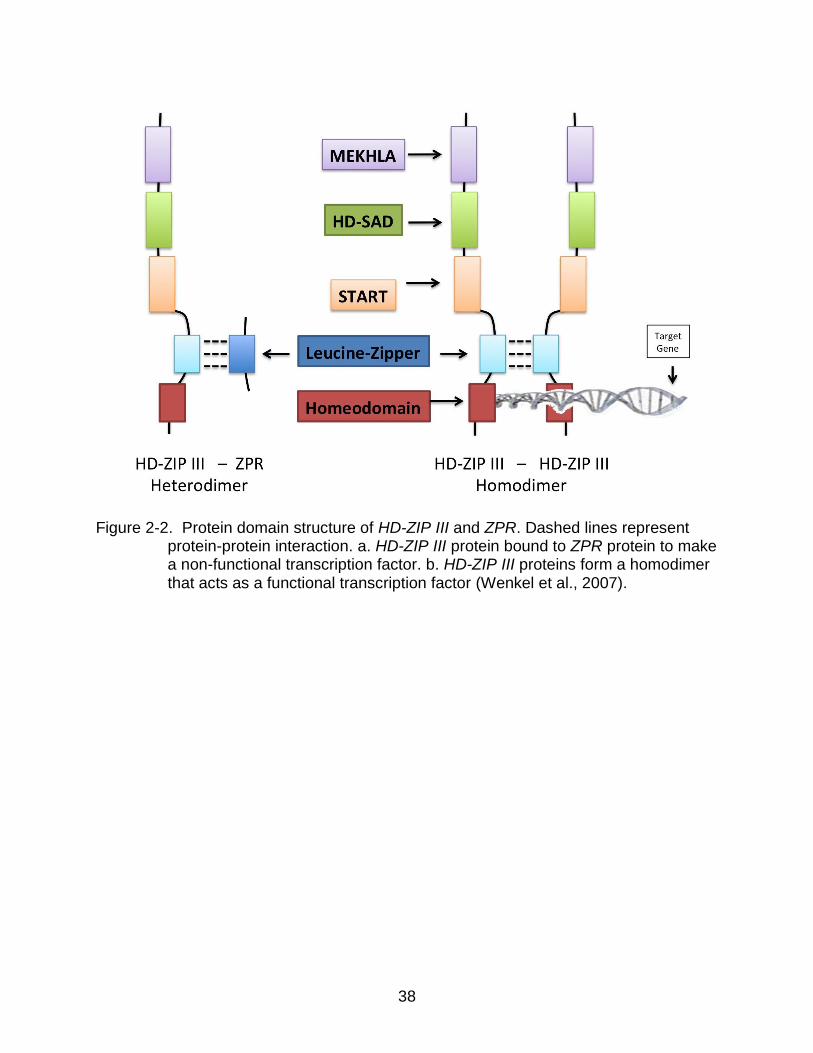
38
Figure 2-2. Protein domain structure of HD-ZIP III and ZPR. Dashed lines represent
protein-protein interaction. a. HD-ZIP III protein bound to ZPR protein to make a non-functional transcription factor. b. HD-ZIP III proteins form a homodimer that acts as a functional transcription factor (Wenkel et al., 2007).

39
Figure 2-3. Phylogenetic tree of the topology found to have the highest likelihood by
RAxML for sampled vascular plant HD-ZIP III sequences. Numbers at nodes represent the percentage of 1000 bootstrap analyses that support with this topology. The presence of two clades of paleoHDZ3 was not previously known. This phylogeny also represents the first placement of ZPR phylogenetically with HD-ZIP III.

40
Figure 2-4. Subtree of HD-ZIP III vascular plant phylogeny containing lycophtye and PaleoHDZ3 clades. Tips are labeled with their species name, 4-letter 1KP species identifier, and the scaffold number of the sequence. Tips that represent the sequences form Floyd et al. (2006) and Prigge and Clark (2006) are labeled with the sequence names from those publications.

41

42
Figure 2-5. Subtree HD-ZIP III vascular plant phylogeny containing gymnosperm ZPR clade. Tips are labeled with their species name, 4-letter 1KP species identifier, and the scaffold number of the sequence.
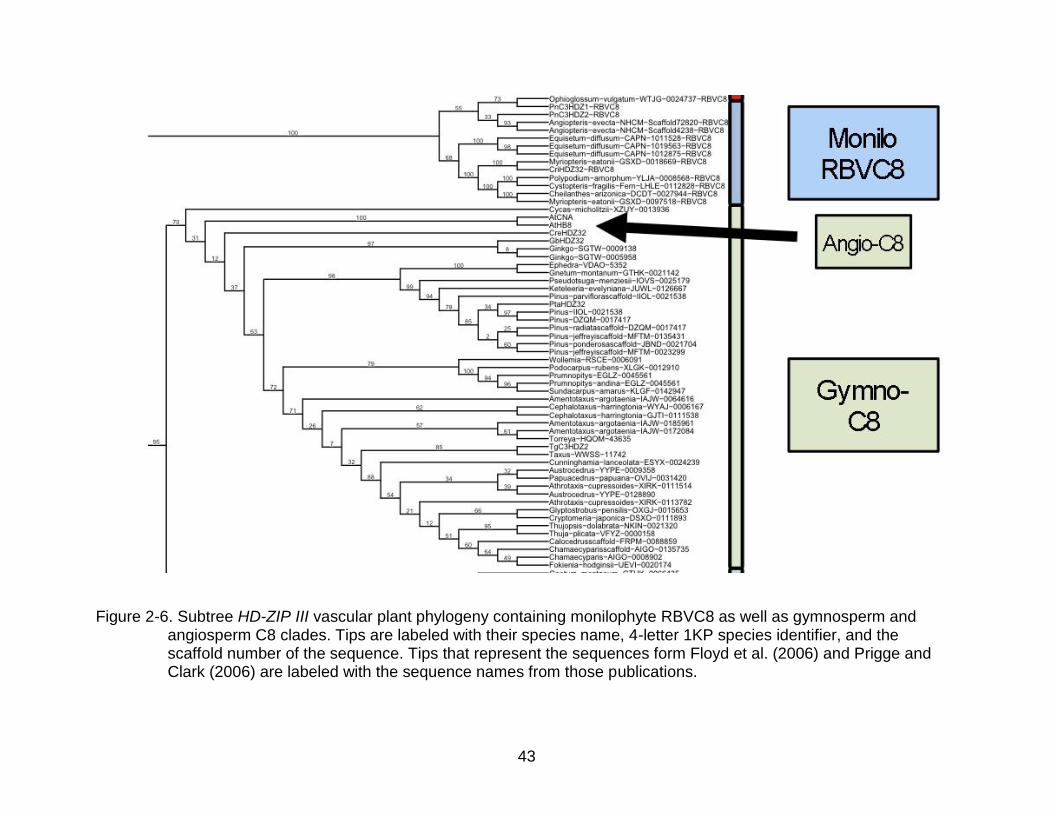
43
Figure 2-6. Subtree HD-ZIP III vascular plant phylogeny containing monilophyte RBVC8 as well as gymnosperm and angiosperm C8 clades. Tips are labeled with their species name, 4-letter 1KP species identifier, and the scaffold number of the sequence. Tips that represent the sequences form Floyd et al. (2006) and Prigge and Clark (2006) are labeled with the sequence names from those publications.
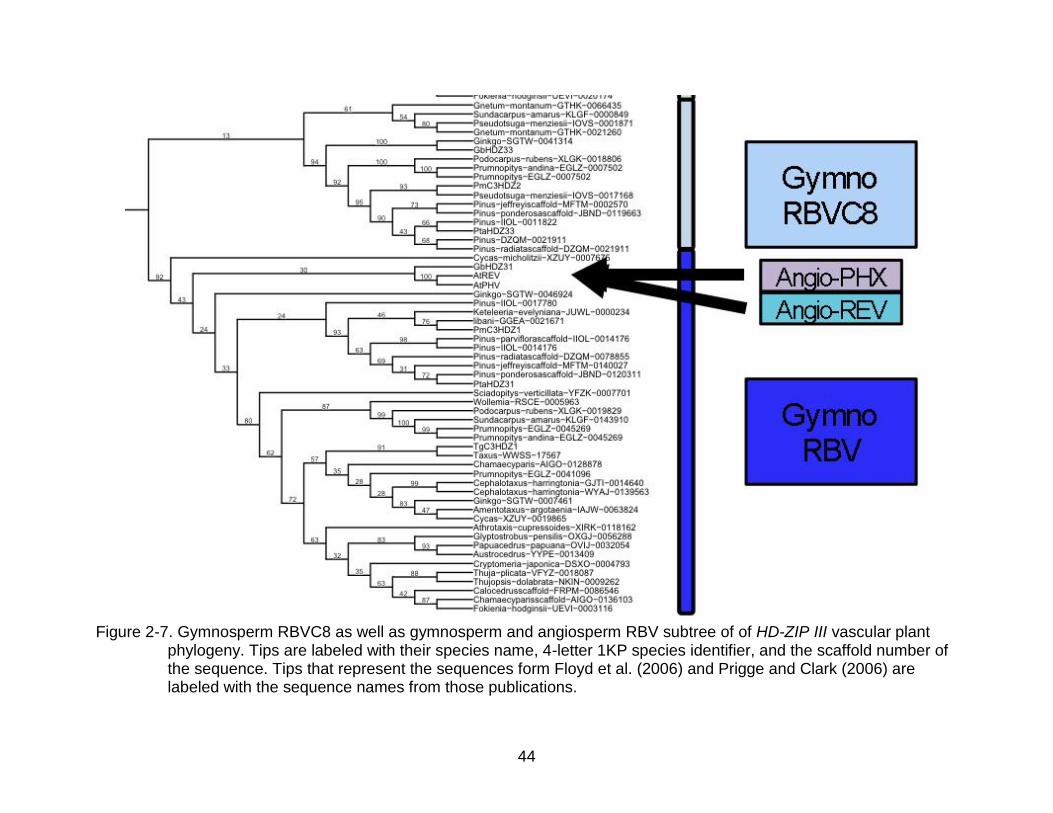
44
Figure 2-7. Gymnosperm RBVC8 as well as gymnosperm and angiosperm RBV subtree of of HD-ZIP III vascular plant
phylogeny. Tips are labeled with their species name, 4-letter 1KP species identifier, and the scaffold number of the sequence. Tips that represent the sequences form Floyd et al. (2006) and Prigge and Clark (2006) are labeled with the sequence names from those publications.

45
Figure 2-8. Alignment of gymnosperm ZPR sequences and the Homeodomain-Leucine-Zipper Region of HD-ZIP III. Yellow dots mark Leucine sites in HD-ZIP III that have become Isoleucine sites in ZPR. Black dots mark homologous leucine sights in HD-ZIP III and ZPR Leucine-Zipper domains. Red bar marks a highly conserved motif between ZPR and HD-ZIP III.

46
CHAPTER 3 GENOMIC COEVOLUTION OF HD-ZIP III RELATED PATHWAY GENES
Introductory Remarks
Polyploidy or whole-genome duplication (WGD) is a common phenomenon in
plants and is concurrent with the major diversifications in angiosperms (e.g., Soltis et
al., 2009; Van de Peer, 2011; Jiao et al., 2011). Polyploidy is known to be a source of
genetic innovations with the main mode of the evolution of genetic complexity in
angiosperms coming from gene and whole-genome duplications (Otto & Whitton, 2000;
Mable, 2003; Gregory & Mable, 2005; Freeling and Thomas, 2006; Soltis et al., 2010;
Buggs et al., 2011). The diversification of transcription-associated proteins, in particular,
has been implicated in the increase in complexity of land plants (Richardt et al., 2007)
and more recently the evolution of such structures as the seed and flower following
ancient polyploidy in the early evolution of seed plants and angiosperms (Zahn et al.,
2005; Jiao et al. 2011).
Many functionally characterized developmental genes in angiosperms have been
highly influenced by gene duplications stemming from ancient polyploidy events. These
genes and their duplication events have been implicated in helping to generate
morphological innovations in angiosperms such as flowers and fruits (Irish and Litt,
2005; Zahn et al., 2005). The process of subfunctionalization or neofunctionalization of
developmental gene duplicates has been shown or hypothesized to spur many of these
morphological novelties. A duplication of the leaf shape gene Asymmetric
Leaves1/Rough Sheath 2/Phantastica (ARP) gene has been hypothesized to be
involved in the morphologically diverse leaf shapes in the stone plants, Ruschioideae
(Iliing et al., 2009). A gene duplication event and subsequent subfunctionalization of the

47
leaf development gene PHANTASTICA have been implicated in the evolution of the
compound leaf in Lotus japonicus (Luo et al., 2005). Duplications in the organ polarity-
controlling gene family, YABBY, have also been involved in patterning angiosperm-
specific organs (Bartholmes et al., 2011). By sampling the genome of Utricularia, Ibarra-
Laclette et al. (2012) found an abundance of duplications in TCP (TEOSINTE
BRANCHED1/CYCLOIDEA/PCF) genes and hypothesize that they may be responsible
for the large amount of branching diversity seen in the genus.
Even though individual gene family duplications and polyploidy have been
implicated in a broad range of evolutionary development studies of land plants, few
studies have investigated at the effects of duplication events on pathways or sets of
interacting genes. Many questions still remain about what happens to interacting genes
after polyploidy. For instance, do other genes in a pathway become sub- or
neofunctionalized when a gene acquires a new developmental task? If a gene takes on
a new function leading to a new phenotype, do the other interacting genes
subfunctionalize to interact with that gene to produce that new phenotype? Also, are
interacting genes maintained in stoichiometric relationships? These same questions are
just as poorly understood when considering the opposite of gene duplication, gene loss.
Considering how many polyploidization events have occurred in angiosperm history
(Soltis et al., 2009; Jiao et al., 2012), but the relatively stable number of genes in the
angiosperm genome (Table 3-1) (Galbraith et al., 2011; Soltis and Soltis, 2013), gene
loss is a prevalent process, but the rules governing loss are poorly understood (Soltis et
al., 2010).

48
Here I investigate these questions by characterizing the duplications and losses
in perhaps the best functionally characterized family of transcription factors involved in
plant body plan, Class III Homeodomain-Leucine Zipper (HD-ZIP III) (Prigge et al.,
2005), and three of its closely interacting genes, Little Zipper (ZPR), KANADI (KAN),
and microRNA 166/165 (miR166/165) in six fully sequenced angiosperm genomes.
The six angiosperm species compared here, Amborella trichopoda
(Amborellaceae), Aquilegia coerulea (Ranunculaceae), Vitis vinifera (Vitaceae),
Arabidopsis thaliana (Brassicaceae), Mimulus guttatus (Phyrmaceae), and Utricularia
gibba (Lentibulariaceae), span a large swath in the phylogenetic and morphological
diversity of angiosperms and have varying numbers of inferred paleopolyploidy events
in their history (Figure 3-1, Figure 3-2). Amborella trichopoda is restricted to the island
of New Caledonia (Thien et al., 2003) and represents a monotypic lineage that is sister
to all other extant angiosperms (Soltis et al., 1999). It has only the single polyploidy
event that is shared among all angiosperms, thought to have taken place prior to the
divergence of all extant angiosperms (Jiao et al., 2012) and displays many putatively
ancestral characteristics of angiosperms, including small, gradually differentiated floral
organs (Endress and Igersheim, 2000; Buzgo et al., 2004) and a lack of vessels
(Carlquist and Schneider, 2001). Aquilegia coerulea is small herb in the diverse
Ranunculales, a clade that occupies a phylogenetically important place as sister to the
rest of the eudicots (Soltis et al. 2011), having originated after the divergence of
magnoliids and monocots. Ranunculales are also important because this lineage
diverged from other eudicots prior to a paleopolyploid event that characterizes core
eudicots (Jiao et al., 2012). Vitis vinifera, grape vine, is a member of the Vitales, an-

49
early diverging rosid. Arabidopsis thaliana is an herbaceous annual in Rosidae that is a
well-developed genetic model plant with two inferred paleopolyploid events in the
history of its order, Brassicales (Simillion et al. 2002; Soltis et al., 2009). Mimulus
guttatus is a model plant for ecological diversification and an asterid with few
paleopolyploid events. Except for the paleohexaploid event (or two WGD events)
inferred to have happened early in eudicot evolution (Jiao et al., 2012), Mimulus may
have only one other recent paleopolyploid event inferred to share with the recently
sequenced genome of the carnivorous plant Utricularia (Ibarra-Laclette et al., 2013).
Utricularia gibba is an aquatic, and morphologically ambiguous, carnivorous bladderwort
(Rutishauser and Isler, 2001) that lacks true roots. It has one of the smallest nuclear
genomes of all sampled angiosperms, with an estimated size of only 88 megabase pairs
(Greilhuber et al., 2006), but has undergone two polyploidy events since its most recent
common ancestor with Mimulus. Based on genomic synteny, the contradictory number
of genome duplications and small genome size of Utricularia have been explained by
extreme fractionation of its genome (Ibarra-Laclette et al., 2013).
Perhaps one of the plant gene families whose paralogs are best functionally
characterized in Arabidopsis and other model plants is HD-ZIP III (Prigge et al., 2005). It
is also part of a pathway that is functionally and evolutionarily well described, with both
post-transcriptional and post-translational regulation understood (Kinder and
Timmermans, 2007; Bowman, 2004; Wenkel et al., 2007).
HD-ZIP III is a transcription factor that controls the polar patterning of many plant
developmental stages, including the embryo, meristem, lateral organs, and vascular
cambium (Prigge et al., 2009). It acts as a homodimer that binds to target genes and

50
has three protein domains that act as sites for upstream chemical and physical stimuli
(Magnani and Barton, 2011). In lateral organs and vascular cambium it is expressed on
the adaxial side (Ohashi-Ito and Fukuda, 2003).
ZPR is a microprotein that has a lone leucine-zipper domain. It is activated by
HD-ZIP III, and its mature proteins in turn bind to HD-ZIP III proteins, forming a
nonfunctional heterodimer, a process that seems to be an adaptation for finely
regulating the action of HD-ZIP III.
KANADI is a GARP-domain transcription factor that is abaxially expressed by a
mutually antagonistic interaction with HD-ZIP III, and it is this juxtaposition of expression
domains that is thought to define the plain for lamina outgrowth (Kinder and
Timmermans, 2011). Few studies have investigated the evolutionary history of the
KANADI gene, although it has been suggested that its evolution has played a
fundamental role in the evolution of the plant body plan (Yamaguchi et al., 2012).
MicroRNA166/165 is a regulator of HD-ZIP III that has a complementary 21-
nucleotide binding site that acts with ARGONAUTE1 and other microRNA machinery to
direct cleavage of target HD-ZIP III mRNAs (Kinder and Timmermans; 2011). It is
expressed abaxially, thereby post-transcriptionally excluding HD-ZIP III expression from
the abaxial domain (Bowman, 2004). MicroRNA165 differs from 166 by one nucleotide,
and separate functions of the two microRNAs are not highly evident (Maher et al., 2006)
By fully sampling the genomes of multiple species for genes that interact together
in a pathway, the following specific questions can be addressed:
1. Are duplications of the genes noted above correlated with known paleopolyploid events?
2. Do genes in a pathway show stoichiometry of paralogs in the genome?

51
3. Can co-subfunctionalization among different gene families be deduced from similarly timed duplications or losses?
Also, by sampling the genome of the carnivorous plant, Utricularia gibba, a plant with
highly aberrant leaf, embryo, and shoot development and a debated absence of roots,
these questions can be addressed:
4. Is a plant with an extremely different morphology and development constrained to the same gene stoichiometry as other, more morphologically ‘typical’ plants?
5. Can any gene losses be attributed to the loss of typical root development?
6. Do the sampled gene families inferred to have a role in either carnivory or the highly modified morphology of U. gibba show similar fractionation patterns as the whole genome?
Methods
Sequence Retrieval
Arabidopsis sequences were downloaded from The Arabidopsis Information
Resource (TAIR) based on the curated names of CORONA (CNA), HOMEOBOX8 (HB-
8), REVOLUTA (REV), PHABULOSA (PHB), PHAVULOTA (PHV). TBLASTn searches
of the other angiosperm genomes were performed with queries of conserved domains
of the three genes and the binding site of the microRNA. Amborella trichopoda and
Utricularia gibba searches and sequence downloading were performed at CoGe
genome sequence database (genomevolution.org). Vitis vinifera, Aquilegia caerulea,
and Mimulus guttatus searches and sequence downloading were performed at
Phytozome (phytozome.org).
HD-ZIP III tBLASTn searches were performed with the conserved homeodomain-
leucine zipper region of the Arabidopsis HD-ZIP III paralogs. Matching sequences were
further pared down by accessing the results by sets of genes with similar e-values
(around -15) and correct HD-ZIP III exon-intron structure.

52
MicroRNA 166/165 searches were performed with the ultra-conserved 19-
nucleotide sequence “TCGGACCAGGCTTCATTCC” at high e-value settings of -0.1
and -0.01 to account for the short sequence. Matching sequences were further pared
down by the criteria of not being in predicted coding regions, as microRNAs should not
code for proteins, and having a loosely conserved matching sequence within 200 bp
that would correspond to the complementary binding site in the preprocessed RNA
hairpin stem.
ZPR tBLASTn searches were performed with the conserved leucine zipper
domain, “IIRENEKLKKKALLLHQENKTL”.
KANADI tBLASTn searches were performed with the first half of the conserved
GARP domain, “APRMRWTTTLHAHFVHAVELLGGHE”, located on the N-terminus of
an exon-intron splice site in the middle of the GARP domain. E-value parameters were
set to E=0.001 in CoGe and -1 in Phytozome. Matching sequences were further filtered
down by accessing the results by sets of genes with similar e-values and correct
KANADI exon-intron structure.
Deeper relationships of KANADI and GARP-domain genes of other green plants
were investigated by searching for homologues in the genome of the moss,
Physcomitrella patens, on the Phytozome website, as well as the transcriptomes of the
green algae, Chara vulgaris and Coleochaete irregularis, from the One Thousand Plant
Transcriptome project (1KP; www.onekp.com) data set. Previously published GARP
sequences from the protozoan, Giardia lamblia (Sun et al., 2006), were downloaded
from GenBank.

53
Alignment
Translation of the nucleotide sequences of the three sets of genes, HD-ZIP III,
ZPR, and KANADI, to amino acid sequences was done Geneious version 5.6.5
(Biomatters, www.geneious.com) with the Geneious progressive pairwise alignment
algorithm and then manually aligned by highly favoring homology based on amino acid
polarity and conservatively using gaps based on the assumption that amino acids
evolve by transmutations of whole codons rather than insertions/deletion of individual
sites.
Phylogenetic Analyses
The amino acid alignments were phylogenetically analyzed using RAxML
(RAxML-VI-HPC) (Stamatakis, 2006) with the GTR + I + Gamma model of amino acid
sequence evolution to account for the evolutionary distance of the sampled taxa
(Brocchieri, 2001). The topology with the highest maximum likelihood score was
generated, and support for that topology was assessed by 1000 bootstrap replicates
using the thorough bootstrap algorithm.
Arabidopsis Microarray Expression
Expression patterns of 10 of the 13 paralogs from the three coding genes are
present on the Arabidopsis AtGenExpress Visualization Tool (Schmid et al., 2005).
Locus identifiers for HD-ZIP III (At5g60690, At4g32880, At1g30490, At1g52150,
At2g34710); ZPR (At2g45450, At3g52770); KANADI (At4g17695, At5g42630,
At5g16560) genes were entered into the website, and expression charts were
generated and printed from the website.

54
Results
Paralog Numbers
The number of paralogs of each gene increases with the known paleopolyploid
events in the species history. Amborella, the plant with the fewest paleopolyploid
events, has the smallest number of each of the genes, and Utricularia, the plant with the
most paleopolyploid events, has the largest number of genes, despite its small genome
size (Table 3- 2). The number of copies of HD-ZIP III and miR166 in the genome of the
six sampled species correlates well, with a correlation coefficient of 0.961 (p-value =
0.0021) (Figure 3-3A). The number of copies of HD-ZIP III and KANADI correlated
equally well, with a correlation coefficient of 0.961 (p-value = 0.0022) (Figure 3-3B).
However, the number of copies of HD-ZIP III and ZPR does not correlate, with a
correlation coefficient of 0.195 (p-value = 0.7106) (Figure 3-3C). Mimulus shows a
reduction in the number of ZPR and KANADI paralogs with one paralog of KANADI
showing signs of pseudogenesis. The number of miR166/miR165 precursor copies
found in the genomes did show a steady increase in genomes (Figure 3-3D).
Pathway Proportionality
The proportions of each of the genes, defined as the total number of copies of a
gene, divided by the total number of copies in the pathway, in the six sampled plant
genomes are very similar (Table 3-3, Figure 3-2). The microRNAs show the highest
proportion in each of the genomes, varying between 0.38 - 0.43, with an average of 0.4.
KANADI and HD-ZIP III have the exact same number of paralogs in four of the six
genomes (Amborella, Aquilegia, Vitis, and Utricularia) with proportions between 0.17
and 0.27, and an average of 0.22 and 0.2, respectively. ZPR had the smallest

55
proportion of the genes in all six genomes, varying between 0.14 - 0.24, with an
average of 0.17.
Lineage Evolution
HD-ZIP III was inferred to have undergone nine duplication events and three
losses in the sampled lineages, with four of the duplications and all three of the losses
happening in the Utricularia lineage (Figure 3-4) and one loss occurring in the CNA
lineage, leaving only one paralog in its genome from the C8 lineage (Figure 3-5).
ZPR was inferred to have undergone 10 duplication events and four losses; one
of the losses was inferred to be a loss of a major lineage at the base of the sampled
asterids, with only one loss in the Utricularia lineage (Figure 3-4).
The sampled KANADI sequences were inferred to be a separate lineage from
other GARP-domain genes based on phylogenetic distance from other Arabidopsis
GARP-domain genes and similar green algae and protozoan sequences (Figure 3-6).
KANADI was inferred to have experienced 10 duplication events and two losses, with
one duplication at the base of the rosids and a subsequent loss of one duplicate paralog
in the Arabidopsis lineage. Five of the duplications that were inferred to have occurred
in the Utricularia lineage were restricted to the Group II KANADI lineage. The one loss
in the Utricularia lineage left the genome without any paralogs of the Group II KANADI
lineage (Figure 3-7).
MiR166/165 sequences show high amounts of variation between the two binding
sites in the prepossessed hairpin structure, prohibiting an alignment with confident
homology assessments and subsequent phylogenetic analyses (Figure 3-8). It was
evident, however, that the miR165 sequence evolved recently in the Arabidopsis
lineage. Homology between the genomic regions containing the microRNAs was

56
possible by comparing the synteny of the surrounding genomic regions (Figure 3-9). In
every genome sampled, there was evident homology between an area of the genome
that contained a tandem duplication of two of the microRNA 166 precursors (Table 3-3).
In Arabidopsis this area was where miR166C & D are found on chromosome 5,
separated by 1.8 kb.
The ZPR phylogeny showed two main lineages in angiosperms with a loss of one
of those lineages in the sampled species from the asterids (Figure 3-10).
Discussion
This study has shown that polyploidy was indeed acting as a driver of genetic
innovations in this sampling of interacting genes, especially in the HD-ZIP III gene
family. Many of the duplicate paralogs that came about as a result of paleopolyploid
events have persisted in angiosperm genomes. The other gene families, ZPR and KAN,
show more duplications that do not correlate with paleopolyploid events, specifically the
duplications in the common ancestors of the Aquilegia, rosid, and Mimulus lineages.
However, the phylogenies that were used to infer the evolution of those genes were not
well supported due to the limited sizes of the conserved domain used in the analysis.
Other methods of assessing homology and evolutionary events in these genes, like
gene synteny and phylogenetic analysis of introns, may help better resolve the
relationships.
The finding of conserved ratios of the 4 gene families in the sampled angiosperm
genomes is surprising considering the genomic rearrangements, evolutionary distance,
and morphological differences of the sampled taxa. In Utricularia this is especially
unexpected because of the fractionation that has led to massive genomic shuffling. One
could have also expected that the asterid species, Mimulus and Utricularia, would have

57
been unconstrained in HD-ZIP III pathway stoichiometry because mutants in the model
asterid species Antirrhinum majus have been hypothesized to rely on an alternative
pathway for leaf polarity (Kinder and Timmermans, 2007).
Deducing subfunctionalizations of interacting genes and previously unknown
gene functions was possible in this data set with these methods. Perhaps the most
exciting and obvious of these deductions is the role of certain lineages in the
development of roots. Because Utricularia lacks true root development, we can
hypothesize that the loss of the CNA lineage of HD-ZIP III, Clade I of ZPR, and Group II
of KANADI is related to the loss of a root-specific developmental pathway. I further
corroborated this hypothesis by examining the expression levels of these genes in
unduplicated forms in Amborella and in the better sampled Arabidopsis microarray data.
The preduplicated homolog of CNA and HB8 is the CNAHB8 homolog in Amborella
(and Aquilegia). In the transcriptome data of Amborella, CNAHB8 has by far the highest
number of reads sampled from the root tissue (Figure 3-11). CNAHB8 is also highly
expressed in the stem of Amborella, suggesting that the preduplicated homolog is
patterning vascular development in both the stem and roots. In the Arabidopsis
expression data, CNA is the most highly expressed HD-ZIP III paralog in root tissues
(Figure 3-12). Clade I of the ZPR family was also lost in the genomes of both Mimulus
and Utricularia. Only two of the four ZPR paralogs were present in the Arabidopsis
microarray sampling, but those two paralogs were from Clades I and II of ZPR, with the
Clade I paralog more highly expressed in the root and stem tissues (Figure 3-13).
Group II of KAN was present in all sampled genomes except Utricularia. In the
Arabidopsis expression data, three of the four paralogs were present, and each of the

58
paralogs was from a separate group. The lost Utricularia paralog of KAN corresponds to
the most highly expressed root paralog in the Arabidopsis expression data. The
expression of these KAN paralogs shows a clear case of subfunctionalized expression
of this group in the roots of Arabidopsis (Figure 3-14); however, the mutant of this
paralog, aberrant testa shape, is well characterized as an ovule developmental mutant
(McAbee et al., 2006).
The genomic losses and Arabidopsis expression patterns of those paralogs
provide evidence of subfunctionalization of those lineages for root development. This is
a substantial finding, as root development has not been described for any of these gene
families, even the well-characterized HD-ZIP III (Prigge et al., 2005), and the description
of root development is commonly ignored in mutant screens. However, root
development is controlled by a meristem just as shoots are, and it would follow that
similar genes have been subfunctionalized to control development of this important land
plant organ. This finding also carries great weight on the age-old debate of
morphological homology in Lentibulariaceae. The morphological definitions of the
organs of Utricularia have been debated by every major plant morphologist from the
beginning of modern botany (see Rutishauser and Isler, 2001). A modern “fuzzy”
definition of its organs has been proposed, with a mix of leaf, stem, and root structural
definitions used to explain the organs in the genus. Based on this study, it seems that
the developmental genetics of roots has been lost and that the root-like stolons found in
the genus are not produced by the conserved angiosperm root-specific paralogs of the
studied developmental genes. The modern morphological definition of Utricularia does
seem to be correct in assuming that Utricularia has undergone an extensive

59
canalization and a subsequent decanalization (much like the “fractionation” of Ibarra-
Lacette et al. (2013)) of its genome that is reflected in a “decanalization” of its
morphology (Rutishauser and Isler, 2001).
This study has shown the utility of whole genome sequencing from
phylogenetically important taxa for understanding gene function and evolution,
particularly of genes controlling aberrant morphologies, such as those in carnivorous
plants. The many copies of genes in angiosperm genomes can make it difficult to find a
specific mutant that shows a phenotypic difference and therefore implicate it in the
development of a specific part of the plant because of functional overlap of gene
duplicates. This limitation makes the approach used in this study a valuable alternative
for identifying genes with specific phenotypic effects. This study has also shown that a
more holistic view of gene evolution, by including interacting genes and investigating
losses as well as duplications, can lead to a better understanding of plant evolutionary
development

60
Table 3-1. Number of paralogs of the 4 gene families in the 6 sampled angiosperm genomes. Total number of genes, calculated by adding the number of all 4 gene families for that plant, is reported in column 6. The number of paleopolyploid (PP) events known in each species’ history is given in column 7, and the total number of genes in each species’ genome is given in column 8.
HD-
ZIP III
ZPR KANADI miR166 TOTAL # of
PP
events
Total
# of
genes
Amborella 3 2 3 6 14 1 26.8k
Aquilegia 3 4 3 7 17 1 24.8k
Vitis 4 4 4 8 20 2 26.3k
Arabidopsis 5 4 4 9 22 4 27.4k
Mimulus 7 3 6 10 26 3 25.5k
Utricularia 7 4 7 11 29 5 28.5k

61
Table 3-2. Proportions of the number of paralogs for each of the 4 gene families in the genomes of the 6 sampled angiosperm genomes. Proportions were calculated by dividing the number of paralogs for each gene family by the total number of genes in the 4 gene families. The average proportions among the 6 genomes are reported in row 8.
HD-ZIP III ZPR KANADI miR166
Amborella 0.21 0.14 0.21 0.43
Aquilegia 0.17 0.24 0.17 0.41
Vitis 0.2 0.2 0.2 0.4
Arabidopsis 0.23 0.18 0.18 0.41
Mimulus 0.27 0.16 0.23 0.38
Utricularia 0.24 0.14 0.24 0.4
Average 0.22 0.17 0.2 0.4
Table 3- 3. Places in the genomes of the sampled angiosperm genomes that contain
the tandem precursor miR166 sequences.
Species Location in genome assembly
Amborella Scaffold 128
Aquilegia Scaffold 69
Vitis Chromosome 7
Arabidopsis Chromosome 5
Mimulus Scaffold 76
Utricularia Scaffold 127

62
Figure 3-1. Phylogeny of major Angiosperm lineages and eudicot orders with the placement of sampled taxa. Based on
Angiosperm Phylogeny Group III (APG III) (Bremmer et al., 2009)
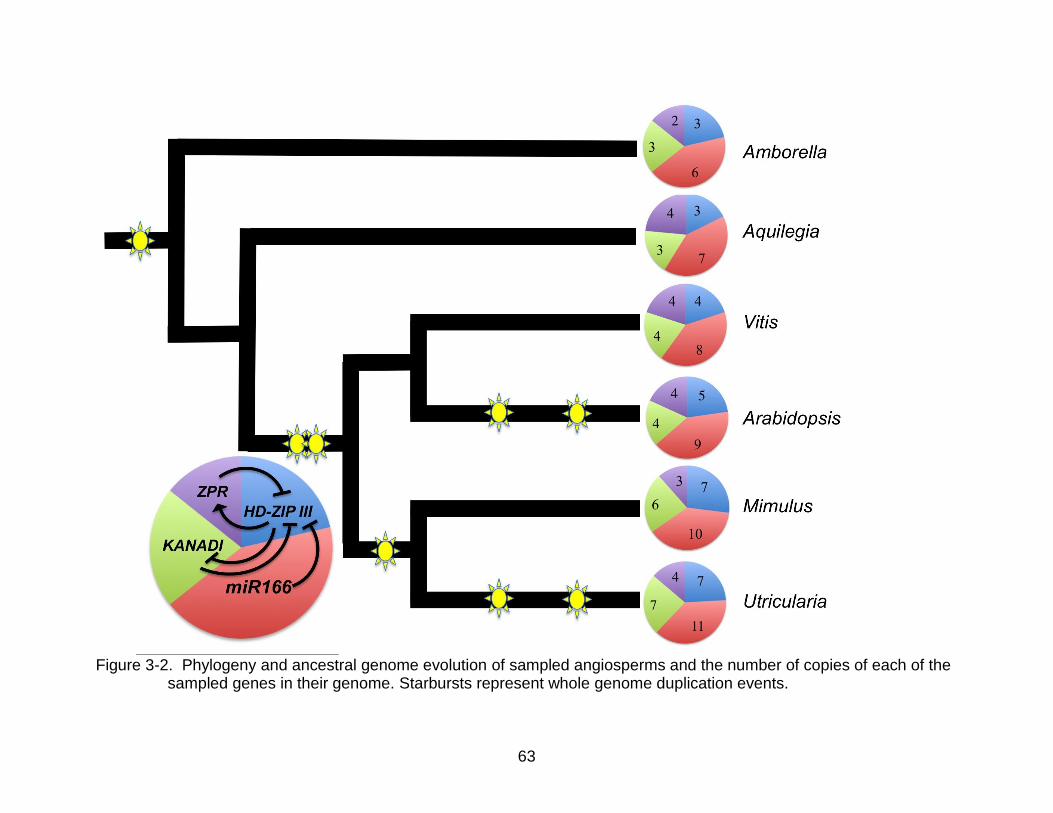
63
Figure 3-2. Phylogeny and ancestral genome evolution of sampled angiosperms and the number of copies of each of the
sampled genes in their genome. Starbursts represent whole genome duplication events.

64
Figure 3-3. Bar graphs comparing the number of copies of HD-ZIP III with the number of copies of the genes that regulate it in the genomes of the six sampled species. A) HD-ZIP III compared with miR166. The number of copies of the two genes correlates well, with a correlation coefficient of 0.961 (p-value = 0.0021). B) HD-ZIP III compared with KANADI. The number of copies of the two genes correlates well, with a correlation coefficient of 0.961 (p-value = 0.0022). C) HD-ZIP III compared with ZPR. The number of copies of the two genes does not correlate well, with a correlation coefficient of 0.195 (p-value = 0.7106). D) Stacked bar graph showing the sum of all four genes added together.
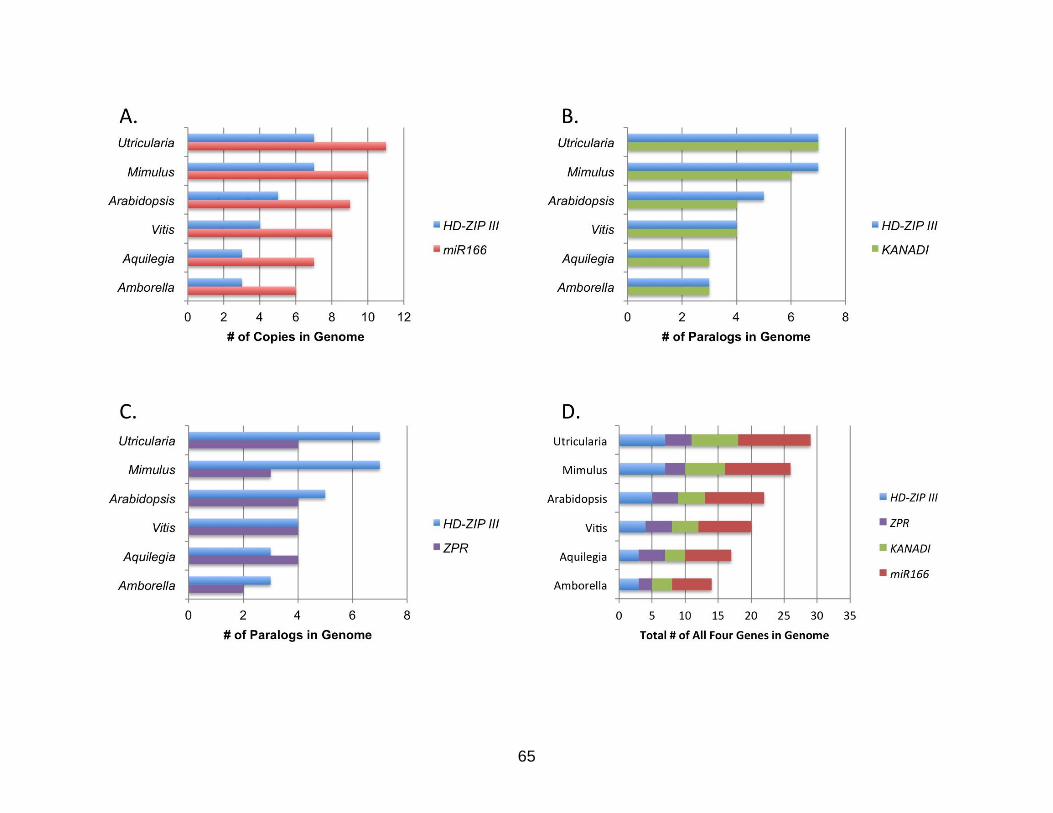
65
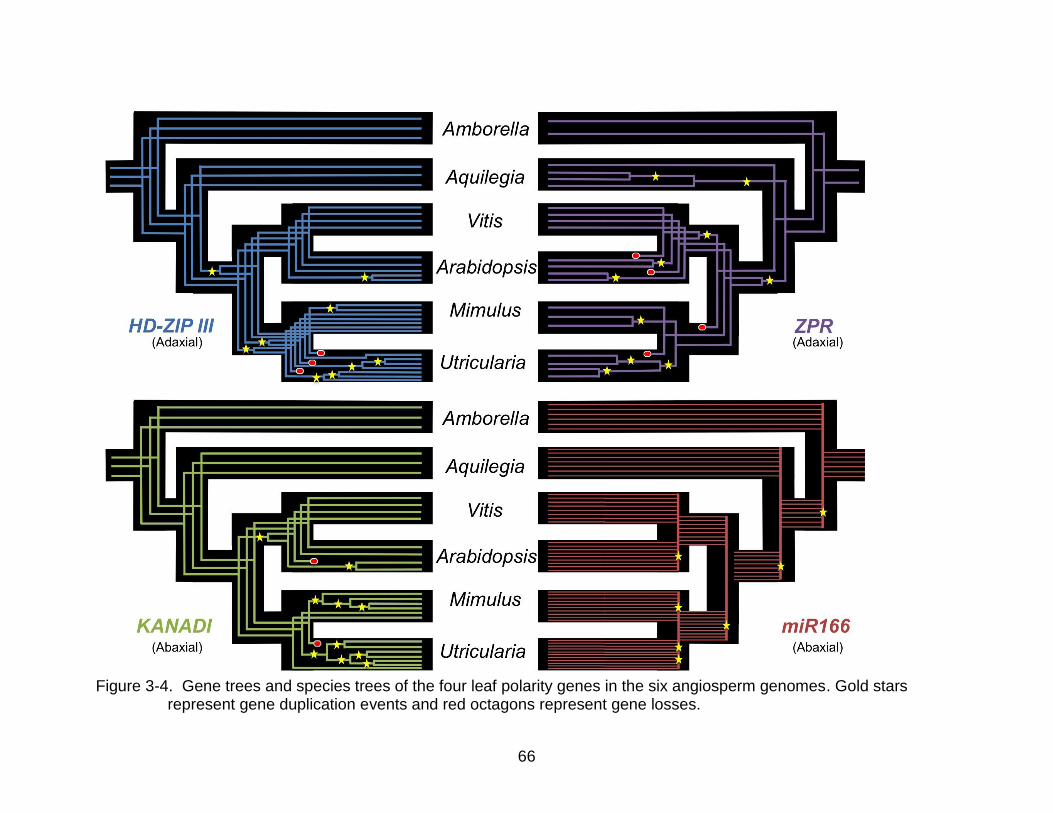
66
Figure 3-4. Gene trees and species trees of the four leaf polarity genes in the six angiosperm genomes. Gold stars
represent gene duplication events and red octagons represent gene losses.

67
Figure 3-5. Phylogram of sampled HD-ZIP III genes. Blue lines represent Amborella genes; red represents Aquilegia;
purple represents Vitis; green represents Arabidopsis; orange represents Mimulus, and yellow represents Utricularia. The labeling of the main clades adapted from Prigge and Clark (2006).

68
Figure 3-6. Unrooted phylogeny of sampled GARP and KANADI genes from green plants and a protozoan.

69
Figure 3-7. Unrooted phylogram of sampled KANADI genes. Blue dots represent Amborella genes; red represents
Aquilegia; purple represents Vitis; green represents Arabidopsis; orange represents Mimulus. Yellow represents Utricularia.

70
Figure 3-8. Alignment of miR166 from the genomes of Amborella, Aquilegia, Vitis, and Arabidopsis. Identity graph displays
the agreement between sequences. The mature microRNA binding site is in 100% agreement at sites 408 – 421 of this alignment. The complimentary binding site in the precursor hairpin structure is in high agreement at sites 245 – 276.
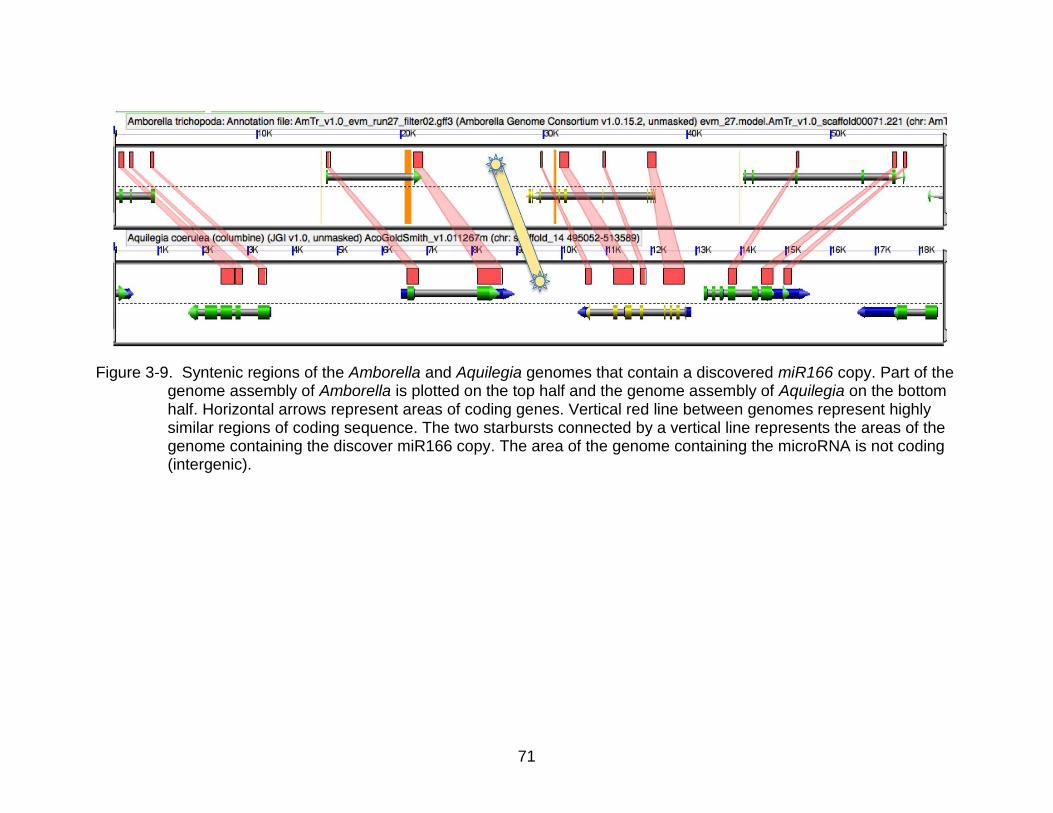
71
Figure 3-9. Syntenic regions of the Amborella and Aquilegia genomes that contain a discovered miR166 copy. Part of the
genome assembly of Amborella is plotted on the top half and the genome assembly of Aquilegia on the bottom half. Horizontal arrows represent areas of coding genes. Vertical red line between genomes represent highly similar regions of coding sequence. The two starbursts connected by a vertical line represents the areas of the genome containing the discover miR166 copy. The area of the genome containing the microRNA is not coding (intergenic).
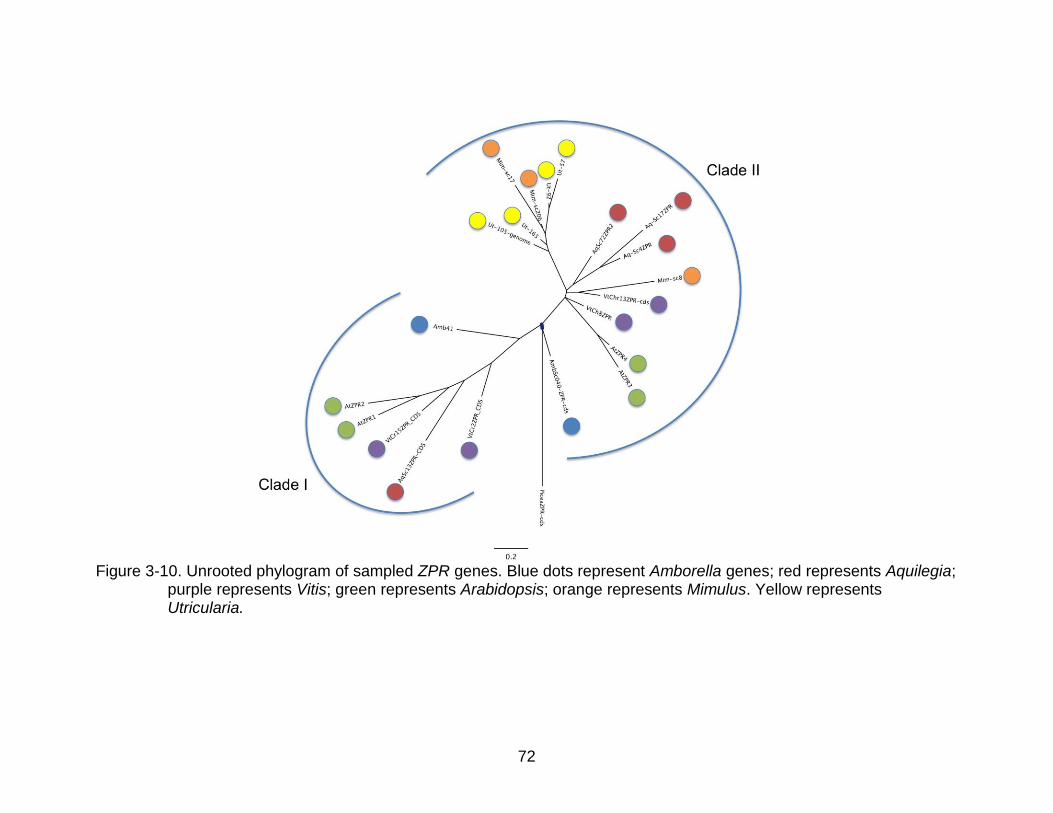
72
Figure 3-10. Unrooted phylogram of sampled ZPR genes. Blue dots represent Amborella genes; red represents Aquilegia;
purple represents Vitis; green represents Arabidopsis; orange represents Mimulus. Yellow represents Utricularia.

73
Figure 3-11. Expression from 454 read counts from Ancestral Angiosperm project (http://ancangio.uga.edu/) of Amborella
HD-ZIP III paralogs. The sampled tissues are grouped on the x-axis. The number of reads in the assembled HD-ZIP III paralogs found in Amborella is plotted on the y-axis. The number of reads is indicative of the expression level of that paralog in the tissue. The CNAHB8 paralog is the highest expressed paralog in the root, stem, and female flower tissue, however Utricularia has lost the CNA paralog.
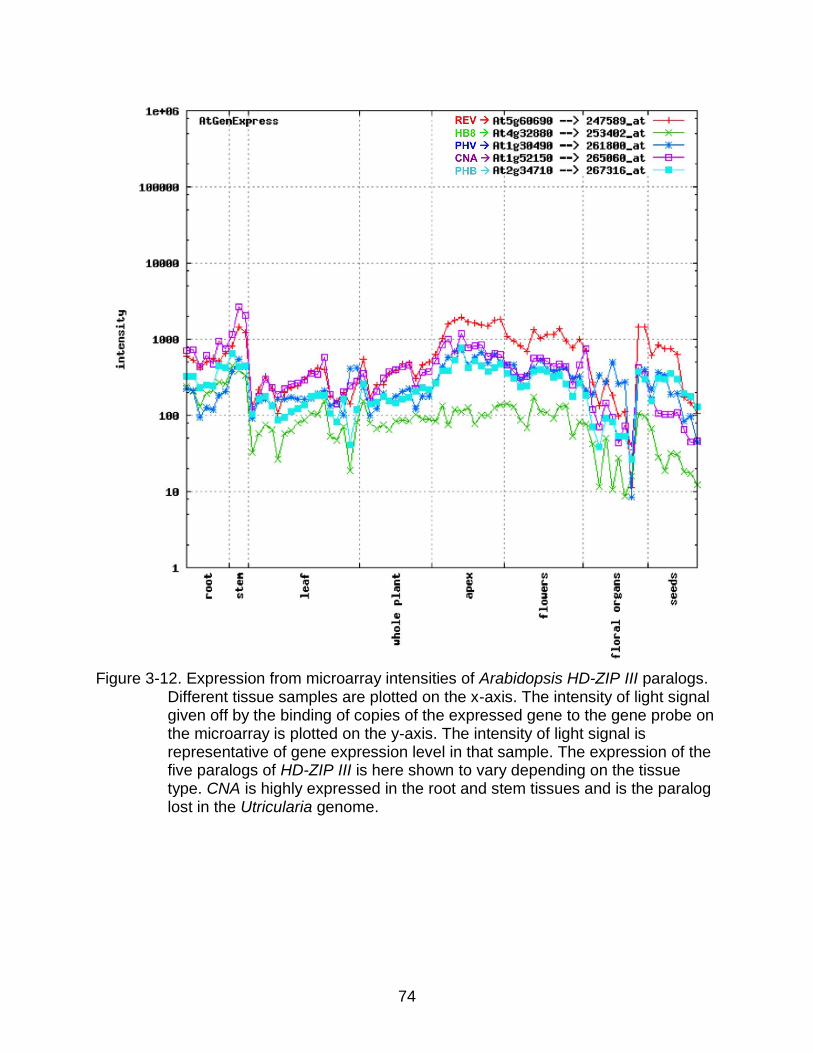
74
Figure 3-12. Expression from microarray intensities of Arabidopsis HD-ZIP III paralogs.
Different tissue samples are plotted on the x-axis. The intensity of light signal given off by the binding of copies of the expressed gene to the gene probe on the microarray is plotted on the y-axis. The intensity of light signal is representative of gene expression level in that sample. The expression of the five paralogs of HD-ZIP III is here shown to vary depending on the tissue type. CNA is highly expressed in the root and stem tissues and is the paralog lost in the Utricularia genome.

75
Figure 3-13. Expression from microarray intensities of Arabidopsis ZPR paralogs.
Different tissue samples are plotted on the x-axis. The intensity of light signal given off by the binding of copies of the expressed gene to the gene probe on the microarray is plotted on the y-axis. The intensity of light signal is representative of gene expression level in that sample. The expression of the two paralogs of ZPR is here shown to vary depending on the tissue type. The ZPR paralog from Clade I is highly expressed in the root and stem tissues, however there are no Mimulus or Utricularia Clade I paralogs in their genome.
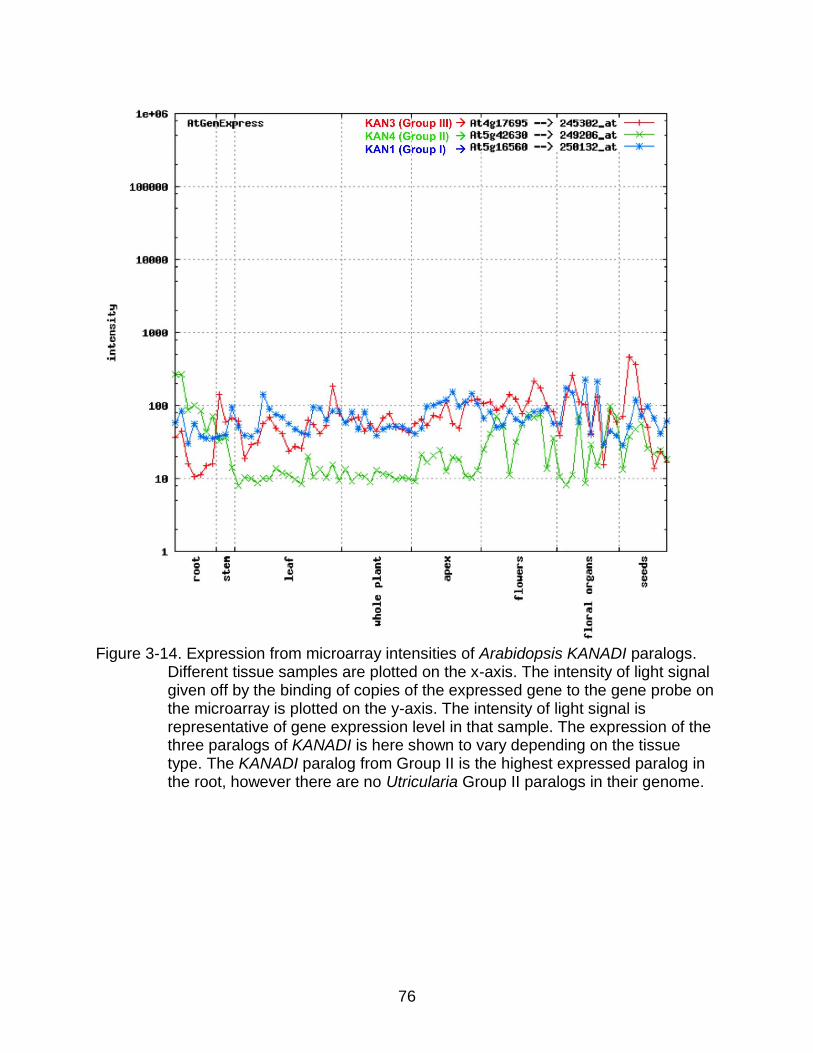
76
Figure 3-14. Expression from microarray intensities of Arabidopsis KANADI paralogs.
Different tissue samples are plotted on the x-axis. The intensity of light signal given off by the binding of copies of the expressed gene to the gene probe on the microarray is plotted on the y-axis. The intensity of light signal is representative of gene expression level in that sample. The expression of the three paralogs of KANADI is here shown to vary depending on the tissue type. The KANADI paralog from Group II is the highest expressed paralog in the root, however there are no Utricularia Group II paralogs in their genome.

77
CHAPTER 4 LEAF EVO-DEVO OF THE OF CARNIVOROUS PITCHER PLANT, CEPHALOTUS
FOLLICULARIS (CEPHALOTACEAE)
Introductory Remarks
Charles Darwin saw the phenomena of carnivorous plants as an evolutionary
puzzle in which natural selection of structures in plants could explain their evolution. He
extensively studied the natural variation in glands of carnivorous plants and the
adaptations of their structure (Darwin, 1875). Today we have the opportunity to
understand the evolution of carnivorous plants from the point of view of the genetic
changes that produced their odd structures. With genetic research in model plants and
current genetic techniques, we can test hypotheses about what genetic regulatory
changes took place during the evolution of carnivorous plants and better understand the
homology of structures.
Because of the extensive morphological change that occurred during the
evolution of carnivorous plants, placing them in plant classification and knowing their
closest non-carnivorous relatives has been historically difficult. Prior to classifications
based on explicitly constructed phylogenetic hypotheses, deducing the morphological
homology of structural features of carnivorous plants to those of non-carnivorous plants
proved challenging (Franck, 1976). With molecular phylogenetics we now know that
carnivory evolved a minimum of four times in angiosperms (Albert et al., 1992; Soltis et
al., 2005). Despite their separate origins from distantly related angiosperms, these
instances of independent evolution reveal strong similarities in native habitats and
trapping mechanisms among carnivorous plants.
Although the different groups of carnivorous plants are found in almost all areas
of the globe, the type of habitats they live in is very similar. Carnivorous plants typically

78
grow in mesic soils that are low in nutrients, particularly nitrogen (D’Amato, 1998).
Carnivorous plants obtain nutrients from insects that they capture, while still carrying out
photosynthesis for energy (D’Amato, 1998). The multiple, independent origins of
carnivorous lineages can therefore be seen as an adaptation to similar environmental
pressures.
The change to carnivory typically has involved large structural changes to leaves,
resulting in flypaper traps, snap traps, lobster pot traps, and pitchers (Soltis et al.,
2005). A given trap type may be found in multiple distantly related clades, with pitcher
traps having originated in three different groups including Sarraceniaceae in the
Ericales, Nepenthaceae in the Caryophylalles, and Cephalotaceae in the Oxalidales.
Thus, the structural evolution of carnivorous plants is an interesting study system
because there is both convergence producing similar yet separate carnivorous groups
and divergence producing variation within the groups.
The modern interpretation of all carnivorous pitchers, including Cephalotus, is
that the unifacial epiascidiate tubular structure of the pitcher (Figure 4-1, Figure 4-2,
Figure 4-3) is achieved through a developmental sequence similar to that of peltate
leaves with outgrowths of the margins of the leaf growing towards the apex and uniting
to form a ring in cross section (Franck, 1976). Various interpretations of the homology of
the pitcher lid have been made, including as an extension of the collar (Arber, 1941), as
a congenitally fused pair of pinnae of a pinnately compound leaf (Troll, 1939; Froebe
and Baur, 1988), and as a ligule (Eichler, 1881).
Molecular genetics of the past two decades have provided insights into the
control of the development of leaf shapes (Tsukaya, 2006; Yamaguchi et al., 2012), with

79
the mechanism of the bifacial polarity of the leaf being closely connected to the lamina’s
morphological shape and marginal growth (Waites and Hudson, 1995; Kinder and
Timmermans, 2010). Gleissberg et al. (2005) examined this model of the genetic control
of leaf polarity in the peltate leaf of Tropaeolum majus (Tropaeolaceae) and found that
the marginal outgrowth that unites the leaf edges and forms the peltate structure shows
expression and shows expression of the abaxial identity gene, YABBY. YABBY is a
gene family with five well-characterized paralogs in Arabidopsis, including CRABS
CLAW (CRC), YABBY2 (YAB2), YABBY5 (YAB5), INNER NO OUTER (INO), and
FILAMENTOUS FLOWER (FIL), and all five have been shown to control the abaxial
identity of lateral organs in Arabidopsis (Siegfried et al., 1999).
Based on the leaf polarity model and evidence of how the model works in other
unifacial leaves, I hypothesize that the epiascidiate leaves of carnivorous pitchers are
also produced by marginal outgrowths associated with abaxial identity gene expression.
I have chosen to test this hypothesis by investigating the expression of a YABBY gene
homolog in Cephalotus follicularis. Cephalotus belongs to the monotypic family,
Cephalotaceae, which is a member of the Oxalidales, making it the most closely related
pitcher plant to Brassicales, the order to which both Arabidopsis thaliana (Brassicaceae)
and Tropaeolum majus (Tropaeolaceae) belong.
To test the hypothesis that the pitcher development in Cephalotus is associated
with a changed YABBY expression domain, I first isolated and described the expression
domain of the HistoneH4 gene as a common positive control for the in-situ protocol
(Gleissberg et al., 2005; Yamaguchi et al., 2012). As this is the first reported in situ
study for Cephalotus and carnivorous plants are known to be difficult material for

80
molecular work (Fleischmann and Heubl, 2009), this is an important first step for
collecting and interpreting in situ hybridization data. Next, YABBY genes were isolated
and their orthology deduced from the phylogenetic placement with previously identified
Arabidopsis and Oryza sativa orthologs (Yamaguchi et al., 2012). The expression of the
isolated Cephalotus FIL ortholog (CephFIL) was then used to identify the adaxial
genetic identity domain in the serial longitudinal section of the developing Cephalotus
pitcher, as was done in the peltate leaf of Tropaeolum (Gleissberg et al., 2005). If
CephFIL expression is seen in the outgrowths that fuse to form the lid and the adaxial
region of the petiole, then I can conclude that the pitcher is genetically similar to peltate
leaves. However, if no CephFIL expression is seen in the lid or adaxial petiole region,
then I would conclude that the lid is an outgrowth of the adaxial region and that it and
the petiole are not formed by the same genetic causes as peltate leaves.
Methods
RNA Extraction and Gene Amplification
Total RNA was isolated from shoot tips and young pitchers of Cephalotus plants
using Plant RNA Isolation Reagent (Invitrogen, Carlsbad, USA), with subsequent
treatment with DNase I (Invitrogen). Total RNA (1 μg) was used for first-strand cDNA
synthesis using the SuperScript III first-strand synthesis system (Invitrogen), and 0.5 μL
of this reaction was used as the template for PCR amplification. Target genes were
amplified using degenerate primers (Table 4-1). Amplified DNA fragments were gel-
extracted and cloned into pCRII-TOPO (Invitrogen), and at least 16 clones per fragment
were sequenced.

81
Sequence Alignment and Phylogenetic Analysis
HistoneH4 and YABBY clones were sequenced with forward and reverse plasmid
cloning site primers, M13F and M13R. The forward and reverse sequences of each
gene were then aligned together and checked for agreement. The HistoneH4
sequences were then translated and aligned with the translated sequence of
Arabidopsis thaliana HistoneH4 sequence downloaded from TAIR
(http://www.arabidopsis.org, AT2G28740.1). The YABBY sequences were translated
and aligned with sequences from Arabidopsis thaliana, Oryza sativa, and Amborella
trichopoda, representing the five major lineages of YABBY genes (Bartholmes et al.,
2012) using Geneious version 5.6.5 (Biomatters, www.geneious.com) with the
alignment algorithm developed by Biomatters and implemented in Geneious. The
YABBY amino acid alignment was phylogenetically analyzed using RAxML (RAxML-VI-
HPC) (Stamatakis, 2006) with the GTR + I + Gamma model of amino acid sequence
evolution. The topology with the highest maximum likelihood score was generated, and
support for that topology was assessed by 1000 bootstrap replicates using the thorough
bootstrap algorithm.
In situ Hybridization
Probe construction
Plasmids containing the sequenced HistoneH4 and the CephFIL paralogs were
amplified using the M13 priming sites in the plasmid and cleaned using a Wizard SV
PCR clean-up kit (Promega, Madison, USA). Appropriate sample dilutions were then
used for RNA probe synthesis using a DIG RNA labeling kit with a T3 and SP6 RNA
polymerase (Roche, Indianapolis, USA).

82
Tissue fixation
Shoot apices and young pitchers were fixed in 3% paraformaldehyde and 0.25%
glutaraldehyde in 0.1 M sodium phosphate buffer for 16 hours at 4°C. Tissue was then
dehydrated in a graded ethanol series, replaced with xylene, and embedded in
Paraplast Plus (Oxford Labware, Mansfield, Massachusetts, USA).
Hybridization
In situ hybridization was performed with a protocol adapted from Kramer (2005)
and Kouchi and Hata (1993). Microtome sections of paraplast-embedded material were
cut at 7 μm and mounted on Probe-On-Plus Slides (Fisher, Waltham, MA). Slides were
de-waxed and re-hydrated in increasing dilutions of ethanol, and proteins were digested
in proteinase K (1 μg/mL) at 37 °C for 40-60 minutes in 100 mM Tris-HCl (pH 7.5), 50
mM EDTA and then washed in distilled water for 5 minutes. After washing in 0.2%
phosphate-buffered glycine solution and phosphate-buffered saline (PBS: 7 mM
Na2HPO4, 3 mM NaH2PO4, 130 mM NaCl, pH 7.0), the tissue was re-fixed in 4%
paraformaldehyde. Positive charges were masked by treatment with 16 mM acetic
anhydrate in 0.1 M triethanolamine buffer. DIG-labeled probes were used for
hybridization at a concentration of 2.5 ng in hybridization buffer consisting of 50%
deionized formamide, 0.3 M NaC1, 10% dextran sulfate, 10 mM TRIS- C1,pH 7.4, 1mM
EDTA, 100mM DTT and 500~tg/ml Escherichia coli tRNA. Sections were incubated at
42° C for more than 16 h with coverslips in hybridization buffer (40-50 gl per slide)
containing the probes. The probe was detected by an alkaline-phosphatase-coupled
anti-digoxigenin-antibody and visualized by NBT/X phosphate blue color substrate.

83
Results
The isolated HistoneH4 transcript from Cephalotus that was used as the template
for the in situ hybridization probe showed 100% amino acid sequence identity with that
of the Arabidopsis thaliana amino acid sequence at 72 amino acid residues which
accounts for 68% percent of the 105 amino acid long protein (Figure 4-4).
Isolated YABBY transcripts from Cephalotus were phylogenetically placed in two
of the major lineages, FIL and CRC (Figure 4-5). All major lineages of YABBY genes
were recovered in the phylogeny, although not with high bootstrap support. In addition
to overall sequence-based orthology assessment, the amino acid sequences of the
Cephalotus transcripts show characteristic motifs of the lineages to which they belong.
The HistoneH4 transcripts were localized in areas of active cell division, as this
gene is expressed during the S phase of the cell cycle (Gaudin et al., 2000).
Specifically, transcripts were detected as bluish-purplish signal in all major areas of the
developing pitcher, including the inner (adaxial) region of pitcher, the tip of the pitcher,
the median cells of the lid, the tip of the lid, and the median cells of the keel (Figure 4-
6).
The CephFIL transcripts were localized in the outer (abaxial) region of the pitcher
but not in the abaxial keel. There was also no CephFIL expression detected in either the
inner (adaxial) region of the pitcher or anywhere in the developing lid (Figure 4-7).
Discussion
These results do not support the hypothesis that the lid and outgrowing margins
of the Cephalotus pitcher would show YABBY expression. This is surprising as the lid is
the only part of the pitcher that is formed by an outgrowth and congenital fusion of the
opposite leaf margins, like the growth that produces peltate leaves and the pitchers of

84
the other two pitcher plant lineages, Sarraceniaceae and Nepenthaceae (Franck, 1976).
However, these data do agree with previous interpretations of the developmental origins
of the pitcher of Cephalotus. The lid has been described by Lloyd (1942) as “an
outgrowth from the upper [adaxial] surface of the petiole below the pitcher proper while
the pitcher has been produced by a ventro-dorsal invagination of the upper, more distal
region.” Because of the CephFIL expression domain, I predict that the lid has genetic
adaxial identity given that the abaxial identity gene is not expressed in the lid and
abaxial-adaxial identity genes have mutually exclusive expression domains (reviewed in
Kinder and Timmermans, 2010). Eichler’s (1881) interpretation of the lid as a ligule also
fits my data, as ligules are adaxial outgrowths.
As reviewed in Bartholmes et al. (2011), the expression domain of FIL has been
documented in some of the model angiosperms and variation in the location of that
domain is present. In the monocots it is the most labial, most likely because of the
multiple paralogs that appear to have come from duplications of the gene prior to the
monocot diversification. In Oryza sativa, the three paralogs YAB3, YAB4, and YAB5
have no expression in the leaf, the abaxial side of the leaf, and throughout the whole
leaf, respectively. In the only isolated paralog of FIL from Triticum aestivum, expression
is throughout the leaf. And in the maize, two of the paralogs are expressed adaxially,
and the precise location of the third paralog is not known. In eudicots, however, FIL is
expressed in the abaxial side of all taxa sampled including Arabidopsis thaliana,
Anthirrhinum majus, Streptocarpus rexii, and Lotus japonicus. Therefore, because
Cephalotus is a member of the eudicots, I am interpreting the expression of CephFIL to
be in the abaxial part of the leaf.

85
No interpretations of the homology of the Cephalotus pitcher have been made in
the years following the studies that have used molecular phylogenetics to place
Cephalotus in the Oxalidales. The exact location of Cephalotus in the Oxalidales has
not been recovered with high support (Soltis et al., 2011) but it is related to
Cunoniaceae, Bruneliaceae, and Elaeocarpaceae. The fact that this pitcher plant is a
rosid and that the lid arises at the base of the leaf and has adaxial identity would allow
for the lid to be interpreted as a pair of fused stipules, as stipules are common in rosids.
Stipules in the Oxalidales are very diverse (Dickinson and Rutishauser, 1990), and
there are no other structures on Cephalotus that have been interpreted as stipules. The
recently described stipules at different growth stages of Geissois pruinosa
(Cunoniaceae) (Fogliania et al., 2009) show similarities (Figure 4-8) to the morphology
of the lid in intermediate pitcher leaves in Cephalotus (Figure 4-2 c-f).
Interpreting the lid as a pair of stipules would also require a change in the
proximal-distal axis since stipules are usually located at the base of the leaf. Mutants of
the KNOX gene in maize show the gene controls the proximal-distal placement of the
ligule with parts of the ligule moving farther towards the distal end of the leaf (Freeling,
1992). A similar change may be happening in Cephalotus to place the lid at the end of
the petiole, near the pitcher. The placement of the stipules in the Oxalidales is labial,
with stipules at the base of the petiole, as in Geissois pruinosa (Figure 4-8) or at the
distal end of the petiole, as in Pseudoweinmannia lachnocarpa (Cunoniaceae)
(Dickinson and Rutishauser, 1990).
The stipules of Arabidopsis have been shown to have genetic adaxial identity
with the trans-acting small interfering RNA Gene, TAS3, showing high expression

86
throughout the whole stipule (Garcia et al., 2006) (Figure 4-9). TAS3 is an indirect
suppressor of FIL, acting through the abaxially expressed AUXIN RESPONSE FACTOR
3 (ARF3/ETT) and ARF4 (Garcia et al., 2006). Therefore, based on the model of
adaxial-abaxial identity gene interactions and the lack of CephFIL expression in the lid
and the predicted adaxial identity of the lid, I am hypothesizing that the lid of Cephalotus
is genetically a pair of fused stipules. This interpretation would also mean that the
pitcher is not genetically homologous to a peltate leaf and that its unifacial qualities are
not produced by a cross-zone formation fusing the two abaxial margins together, as is
seen in the other two independent evolutions of pitchers in Sarraceniaceae and
Nepenthaceae (Franck, 1976). In those two families the lid is at the distal end of the
pitcher, indicating that they are most likely not of stipular origin, and thus marks a
distinctive difference in the evolutionary changes of the leaf that happened during the
convergent evolution of the pitcher in the three different angiosperm clades.
Table 4-1. Nucleotide sequence of primers used to amplify HistoneH4 and YABBY genes in Cephalotus follicularis.
Primer Name 5’ to 3’ sequence
dYABu2 GGGGAAGTGGGCCCARTTYTTNG dYABd1 TGTTCAAGACCGTGACCGTNMGNTGYGG dH4-d1 GGAAGGTGCTGCGGGAYAAYATHCA dH4-u2 CCGCTTCAGGGCGTACACNACRTCCAT
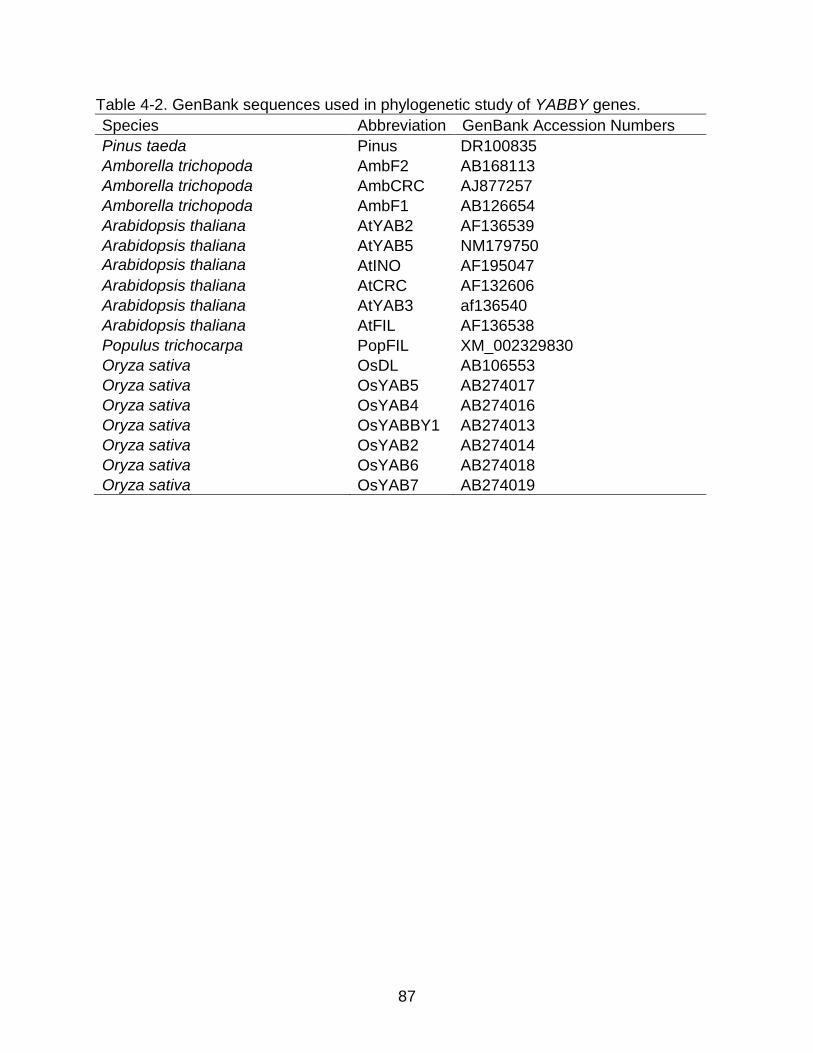
87
Table 4-2. GenBank sequences used in phylogenetic study of YABBY genes.
Species Abbreviation GenBank Accession Numbers
Pinus taeda Pinus DR100835
Amborella trichopoda AmbF2 AB168113
Amborella trichopoda AmbCRC AJ877257
Amborella trichopoda AmbF1 AB126654
Arabidopsis thaliana AtYAB2 AF136539
Arabidopsis thaliana AtYAB5 NM179750
Arabidopsis thaliana AtINO AF195047
Arabidopsis thaliana AtCRC AF132606
Arabidopsis thaliana AtYAB3 af136540
Arabidopsis thaliana AtFIL AF136538
Populus trichocarpa PopFIL XM_002329830
Oryza sativa OsDL AB106553
Oryza sativa OsYAB5 AB274017
Oryza sativa OsYAB4 AB274016
Oryza sativa OsYABBY1 AB274013
Oryza sativa OsYAB2 AB274014
Oryza sativa OsYAB6 AB274018
Oryza sativa OsYAB7 AB274019

88
Figure 4-1. Front view of Cephalotus follicularis pitcher and basal rosette of flat leaves.

89
Figure 4- 2. Photographs of Cephalotus pitcher, flat leaves, intermediate leaf, inflorescence, and flowers. The orientation
of the pitcher (A, B) is depicted in the position that it emerges as a leaf primordium with the keel at the distal end and the lid attached at the proximal base of the petiole.

90
Figure 4-3. Longitudinal section drawing of Cephalotus mature pitcher and shoot tip. Left-most pitcher represents orientation of pitcher at maturity and the structures on either side of the shoot apical meristem represent developing pitchers (not to scale). Lid (L); Ascidium (A); Pitcher (P); Keel (K); and Shoot Apical Meristem (SAM).

91
Figure 4-4. Alignment of Cephalotus and Arabidopsis HistoneH4 amino acid sequences.

92
Figure 4-5. Phylogram of angiosperm and isolated carnivorous pitcher plant YABBY paralogs. Ceph22YAB represents the
CephFIL sequence used as a probe for in situ hybridization. Sequence abbreviations explained in Table 4-2. Cephalotus clones represented by Ceph abbreviation. Sarracenia flava clones represented by Sar, and Nepenthes alata clones represented by Nep.

93
Figure 4-6. In-situ hybridization of HistoneH4 in developing Cephalotus pitcher. Purple staining indicates areas of
HistoneH4 expression. Keel (K); pitcher (P); ascidium (A); Lid (L).

94
Figure 4-7. In-situ hybrizations of CephFIL in serial longitudinal sections of a developing Cephalotus pitcher. The median
of the pitcher is shown first (A) and successive images represent serial sections proceeding to the margin of the pitcher (I). The early keel is located at the distal (top) of the pitcher with the ascidium in the center and the lid at the proximal end of the pitcher. Keel (K); lid (L); ventral side of pitcher (VP); dorsal side of pitcher (DP).

95
Figure 4-8. Stipules in the seedling of Geissois pruinosa (Cunoniaceae). Free stipules
(FrSt) and fused stipules (FuSt) are indicated with arrows. Used with permission from Fogliani et al. (2007).
Figure 4-9. Expression of TAS3 in the vasculature, adaxial domain of developing
leaves, and stipules of Arabidopsis thaliana. Used with permission from Garcia et al. (2006). Blue staining expression of inserted of GUS reporter-gene 37bp of TAS3 (AT3g17185).

96
CHAPTER 6 CONCLUSION
In conclusion, this dissertation has used the leaf polarity genetic model to explore
the evolution of leaves throughout land plant evolution (Chapter Two), genome
dynamics and the loss of roots in angiosperms (Chapter Three), and finally, the
evolution of a highly modified and enigmatic carnivorous pitcher plant (Chapter Four).
These chapters reflect the ever-changing methods of science as most of this data was
collected outside of the lab and required no funding. This research can be seen as just
the beginning of our understanding of the evolution of the genes controlling leaf
development and provides a base on which to explore these findings further.
The findings in Chapter Two are based on the newly available gymnosperm and
monilophyte transcriptomes from the 1KP project. These data fill a substantial hole in
the sequence data available across the spectrum of plants. However, even today
genome sequencing of the gymnosperms and monilophytes is underway, and my
findings will provide hypotheses to test in those genomes. Specifically, better
understanding of the duplication that gave rise to the ZPR gene from HD-ZIP III will be
possible with gymnosperm genome sequences. My findings predict that the ZPR gene
will show genomic synteny with the PaleoHDZa paralog if ZPR did indeed originate from
a duplication of PaleoHDZa. The exact function of the five different HD-ZIP III paralogs
in the development of gymnosperms would also be a key question to address in the
future. As only three of those paralogs were passed on to angiosperms, one wonders
what the other copies do and if their function is similar to or different from HD-ZIP III
functions in angiosperms. Answering functional questions, unlike questions of lineage
evolution, currently requires a large amount of time and funding, but may be of special

97
interest to the timber industry as these genes most likely play a role in wood
development in gymnosperms.
Chapter Three makes predictions about previously unreported functions of HD-
ZIP III in root development. Even though this dissertation set out to improve our
understanding of leaf development, it is a welcome finding that carnivorous plants,
especially Utricularia, might be natural models for root development as well. As a plant
that gets its water and nutrients foliarly, Utricularia is, in a way, a root development
knock-out. Testing the subfunctionalization of the sampled polarity genes by functionally
characterizing their role in the model plant Arabidopsis may be of relative ease given
that the mutants are easily and cheaply available from TAIR. Understanding the losses
of the supposed root development paralogs in Utricularia may also be a plausible
avenue of future research because there is a gradient of root loss in Lentibulariaceae
(the family to which Utricularia belongs). Furthermore, transcriptome or genome
sequencing may be financially feasible for this question and many others about genome
evolution in the family, as the genome size is very small.
Appendix A, if anything, is a testament to the difficulty that carnivorous plants
pose to the techniques of molecular biology, ironically as they also did for morphologists
in the past. Their nucleotide-binding secondary compounds, which are well documented
to impede even the simplest of molecular biology techniques, including nucleotide
extraction and amplification (Bekesiova et al., 1999; Fleischmann and Heubl, 2009),
may be exacerbated in Cephalotus follicularis, the only known carnivorous plant in the
Rosidae, a clade of ~70,000 species (Soltis et al., 2005) also known to be difficult
molecularly (Zamboni et al., 2008). However, understanding the expression domains of

98
leaf polarity genes in carnivorous plants is an important endeavor but one that may be
better approached using laser dissection RT-PCR (Nelson et al., 2006) as secondary
compounds can be removed after dissection during the RNA extraction (Johnson et al.,
2012). New in situ methods may also hold promise (Wang et al., 2013).

99
APPENDIX A FURTHER CARNIVOROUS PLANT EXPERIMENTS
AS2
ASYMMETRIC LEAVES2 (AS2) is a member of the AS2/LOB gene family and
functions in both meristem maintenance and adaxial leaf identity (Iwakawa et al., 2007).
Knowing the expression domain of AS2 in carnivorous pitcher plants would have helped
better understand genetically how the pitcher relates to the flat leaves. Phylogenetic
analyses with Arabidopsis thaliana sequences Nepenthes alata, Cephalotus follicularis,
and Cunonia capensis, a closely related non-carnivorous plant to Cephalotus, were
performed to identify homologous transcripts for in situ hybridization probes (Figure A-
1). The sequences placed phylogenetically close to the Arabidopsis AS2 paralog, were
used for designing PCR primers for isolating AS2 paralogs from Nepenthes and
Cephalotus, and synthesizing probes for in situ hybridization.
The Nepenthes AS2 in situ hybridizations that used Western Blue alkaline-
phospotase conjugation to detect the DIG-labeled probes, showed high background
noise and identical staining patterns to the negative control sense probes (Figure A-2).
The only published studies of gene expression in Nepenthaceae used a
fluorescent detection of DIG-labeled probes (Schulze et al., 1999; An et al., 2001; An et
al., 2002) presumably because the method produces less background noise. This
method was used to try to detect the Nepenthes and Cephalotus AS2 probes, however
no signal could be detected in Nepenthes meristem, young developing pitcher, or older
developing pitchers (Figure A-3). This detection method was also tried on Cephalotus
meristem and young leaf primordia (Figure A-4) but no signal was clearly seen. No
signal could be detected in the developing pitcher either (Figure A-5).

100
VIGS Development
The best way to know how leaf development pathways are functioning in
carnivorous pitcher plants is by knocking down the expression of the pathway genes to
see what affect it has on the pitcher development. On of the most common ways for do
this in non-model plants is by a method called Virus Induced Gene Silencing (VIGS)
(Gould and Kramer, 2007). This method has not been tried on carnivorous pitcher
before. One of the first step is developing this method is finding a strain of
Agrobacterium that can successfully insert a virus vector into the genome of the target
plant. I tested the ability of Agrobacterium tumefaciens AGL1 to insert the commonly
used VIGS virus vectors, TRV1 and TRV2, into the genome of Nepenthes alata,
Sarracenia flava, Cephalotus follicularis, as well as the close relative of Sarracenia,
Actinidia chinensis, and the close relative of Cephalotus, Oxalis corniculata. The
protocol described in Gould and Kramer (2007) was used, specifically the vacuum
infiltration method. To test if the vectors were successfully inserted into the genome, a
DNA extraction was performed 5-days after Agrobacterium infection and tested with
PCR for the presence of TVR1 and TRV2 vectors. The PCR results showed that TRV
vectors were best inserted into the genomes of Sarracenia Alta, Oxalis corniculata, and
Actinidia chinensis (Figure A-6). The vector was not well inserted into the genomes of
Cephalotus, or Nepenthes (Figure A-6).
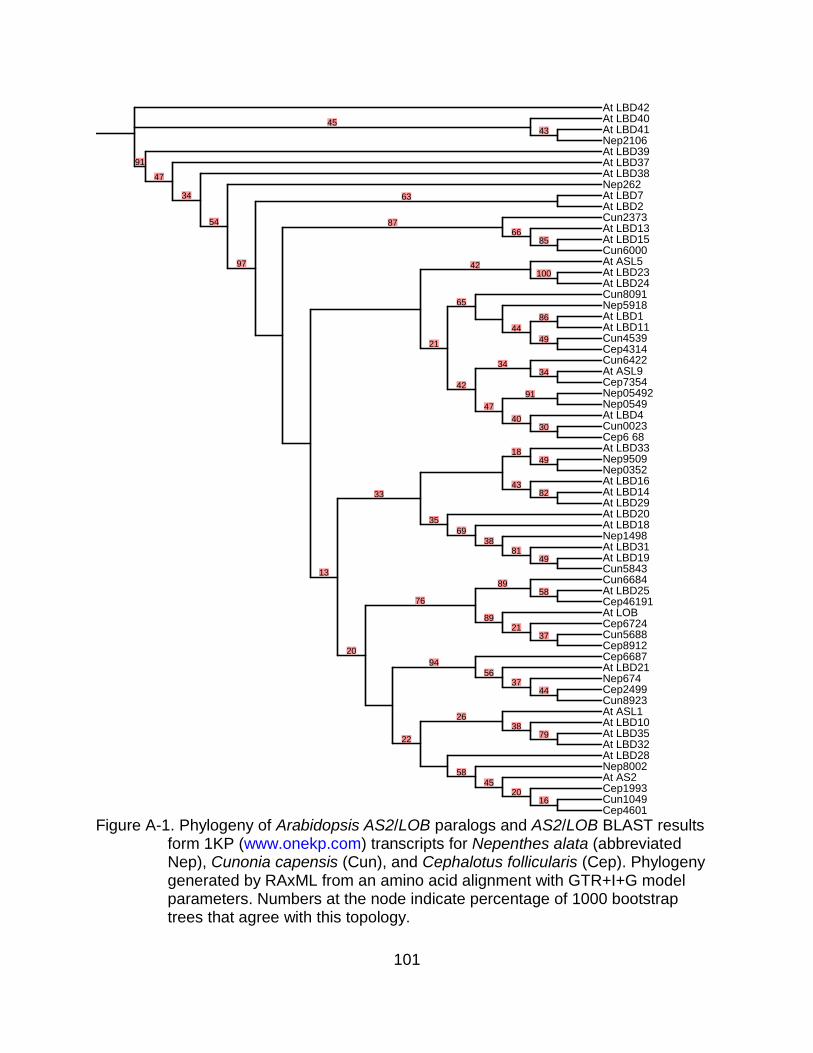
101
At LBD41 Nep2106
At LBD40
At LBD39 At LBD37 At LBD38 Nep262At LBD7 At LBD2
Cun6422At ASL9 Cep7354Nep05492Nep0549At LBD4 Cun0023Cep6 68
Nep5918At LBD1 At LBD11 Cun4539Cep4314
Cun8091
At ASL5 At LBD23 At LBD24
Cun5688Cep8912
Cep6724At LOB
Cun6684At LBD25 Cep46191
At LBD35 At LBD32
At LBD10 At ASL1
Nep8002
Cun1049Cep4601
Cep1993At AS2
At LBD28
Cep6687
Cep2499Cun8923
Nep674At LBD21
Nep9509Nep0352
At LBD33
At LBD16 At LBD14 At LBD29 At LBD20 At LBD18
At LBD31 At LBD19 Cun5843
Nep1498
At LBD15 Cun6000
At LBD13 Cun2373
At LBD42
4543
91
47
34
54
97
63
21
42
3434
47
91
4030
65
44
86
49
42100
13
20
76
8921
37
8958
22
2638
79
58
4520
16
94
5637
44
33
1849
4382
35
69
3881
49
8766
85
Figure A-1. Phylogeny of Arabidopsis AS2/LOB paralogs and AS2/LOB BLAST results
form 1KP (www.onekp.com) transcripts for Nepenthes alata (abbreviated Nep), Cunonia capensis (Cun), and Cephalotus follicularis (Cep). Phylogeny generated by RAxML from an amino acid alignment with GTR+I+G model parameters. Numbers at the node indicate percentage of 1000 bootstrap trees that agree with this topology.

102
Figure A-2. Longitudinal section of in situ hybridization of Nepenthes alata AS2. A)
section through the meristem (arrow head) and leaf primordium (arrow) with putative expression signal from anti-sense probe in the epidermal layers of the meristem and the adaxial side of the primodium. B) section through base of a pitcher sheathing around the meristem. C) section through the meristem and developing pitcher on the right side of the meristem. D) section a few section distal of the meristem with expression signal from the sense probe as a negative control
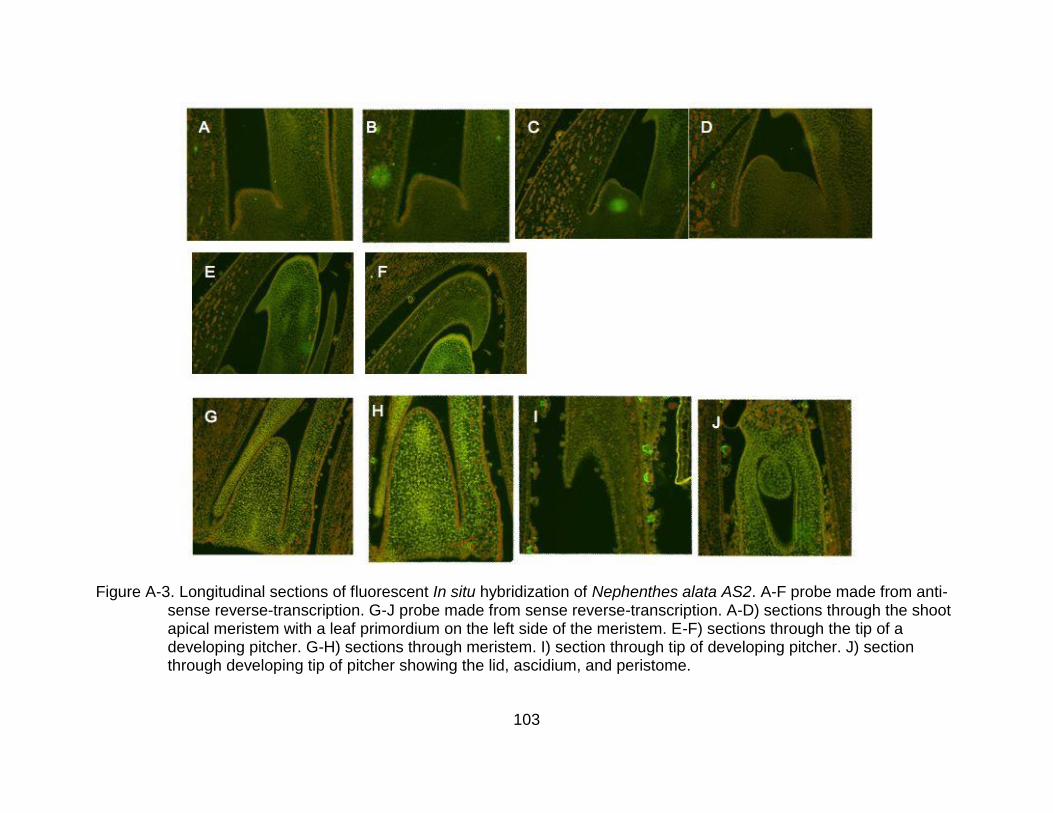
103
Figure A-3. Longitudinal sections of fluorescent In situ hybridization of Nephenthes alata AS2. A-F probe made from anti-
sense reverse-transcription. G-J probe made from sense reverse-transcription. A-D) sections through the shoot apical meristem with a leaf primordium on the left side of the meristem. E-F) sections through the tip of a developing pitcher. G-H) sections through meristem. I) section through tip of developing pitcher. J) section through developing tip of pitcher showing the lid, ascidium, and peristome.
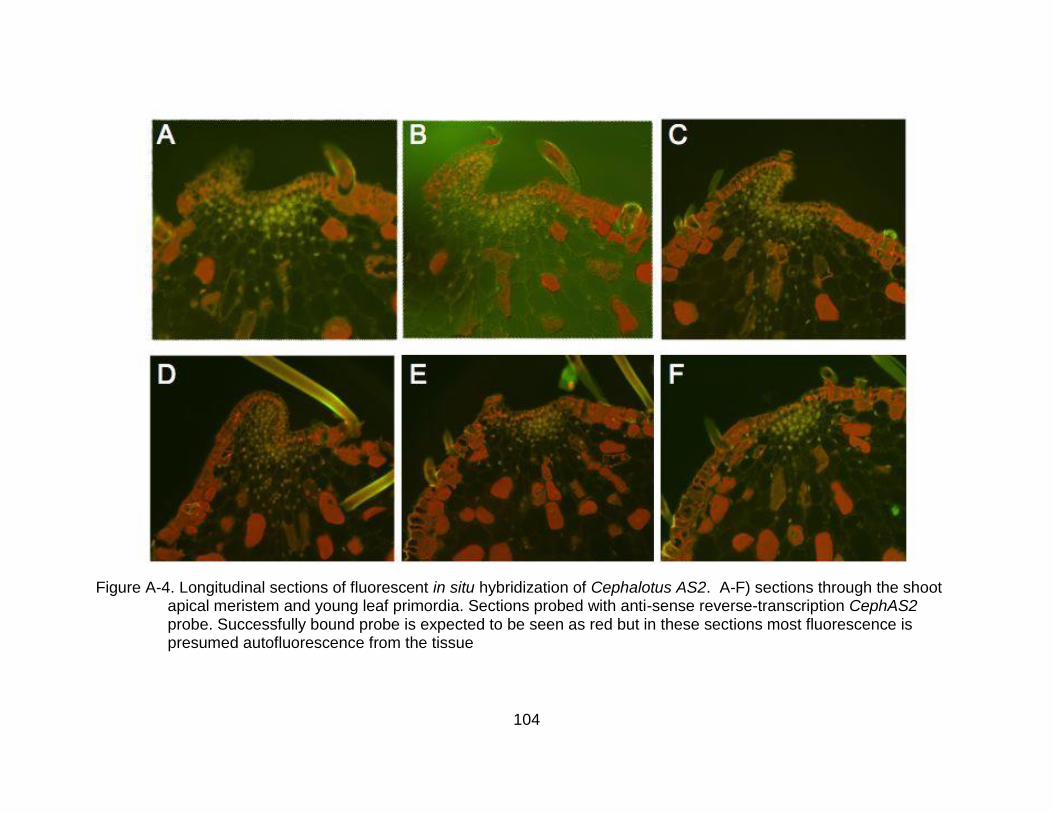
104
Figure A-4. Longitudinal sections of fluorescent in situ hybridization of Cephalotus AS2. A-F) sections through the shoot
apical meristem and young leaf primordia. Sections probed with anti-sense reverse-transcription CephAS2 probe. Successfully bound probe is expected to be seen as red but in these sections most fluorescence is presumed autofluorescence from the tissue

105
Figure A-5. Longitudinal sections of fluorescent in situ hybridization of Cephalotus AS2.
A-L) Developing Cephalotus pitcher in the same developmental stage and position as in Figure 4-7. Sections probed with anti-sense reverse-transcription CephAS2 probe. Successfully bound probe is expected to be seen as red but in these sections most fluorescence is presumed autofluorescence from the tissue

106
Figure A-6. Electrophoresis gel visualization of TRV1 and TRV2 PCR’s of Agrobacterium infected plants. Each well
represents a different plant sample from which the DNA was extracted and used as the template for the PCR. Presence of a band in the well indicates presence of the vector in the genome.

107
LIST OF REFERENCES
Albert, V. A., Williams, S. E., & Chase, M. W. (1992, September). Carnivorous plants: phylogeny and structural evolution. American Association for the Advancement of Science.
Alvarez-Ponce, D., & Fares, M. A. (2012). Evolutionary rate and duplicability in the Arabidopsis thaliana protein–protein interaction network. Genome biology and evolution, 4(12), 1263-1274.
An, C. I., Fukusaki, E. I., & Kobayashi, A. (2001). Plasma-membrane H+-ATPases are expressed in pitchers of the carnivorous plant Nepenthes alata Blanco. Planta, 212(4), 547-555.
An, C. I., Fukusaki, E. I., & Kobayashi, A. (2002). Aspartic proteinases are expressed in pitchers of the carnivorous plant Nepenthes alata Blanco. Planta, 214(5), 661-667.
Arber, A. (1941). On the morphology of the pitcher-leaves in Heliamphora, Sarracenia, Darlingtonia, Cephalotus, and Nepenthes. Annals of Botany, 5(4), 563-578.
Banks, J., Nishiyama, T., Hasebe, M., Bowman, J., Gribskov, M., Depamphilis, C., et al. (2011). The Selaginella Genome Identifies Genetic Changes Associated with the Evolution of Vascular Plants. Science , 332 (6032), 960-963.
Barker, M. S. (2013). Karyotype and genome evolution in pteridophytes. In Plant Genome Diversity Volume 2 (pp. 245-253). Springer Vienna.
Bartholmes, C., Hidalgo, O., & Gleissberg, S. (2012). Evolution of the YABBY gene family with emphasis on the basal eudicot Eschscholzia californica (Papaveraceae). Plant Biology, 14(1), 11-23.
Bekesiova, I., Nap, J. P., & Mlynarova, L. (1999). Isolation of high quality DNA and RNA from leaves of the carnivorous plant Drosera rotundifolia. Plant Molecular Biology Reporter, 17(3), 269-277.
Bowman, J. L. (2004). Class III HD‐ Zip gene regulation, the golden fleece of ARGONAUTE activity?. Bioessays, 26(9), 938-942.
Bowman, J. L., & Floyd, S. K. (2008). Patterning and polarity in seed plant shoots. Annu. Rev. Plant Biol., 59, 67-88.
Boyce, C. K., & Knoll, A. H. (2002). Evolution of developmental potential and the multiple independent origins of leaves in Paleozoic vascular plants. Paleobiology, 28(1), 70-100.

108
Bremer, B., Bremer, K., Chase, M., Fay, M., Reveal, J., Soltis, D., ... & Stevens, P. (2009). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Botanical Journal of the Linnean Society.
Brocchieri, L. (2001). Phylogenetic inferences from molecular sequences: review and critique. Theoretical population biology, 59(1), 27-40.
Byrne, M. E. (2006). Shoot meristem function and leaf polarity: the role of class III HD–ZIP genes. PLoS genetics, 2(6), e89.
Carlquist, S. J., & Schneider, E. L. (2001). Vegetative anatomy of the New Caledonian endemic Amborella trichopoda: relationships with the Illiciales and implications for vessel origin. Pacific Science, 55(3), 305-312.
Chi, X., Yang, Q., Chen, X., Wang, J., Pan, L., Chen, M., ... & Yu, S. (2011). Identification and characterization of microRNAs from peanut (Arachis hypogaea L.) by high-throughput sequencing. PLoS One, 6(11), e27530.
Côté, C., Boileau, F., Roy, V., Ouellet, M., Levasseur, C., Morency, M. J., ... & MacKay, J. (2010). Gene family structure, expression and functional analysis of HD-Zip III genes in angiosperm and gymnosperm forest trees. BMC Plant Biology, 10(1), 273.
Cuatrecasas, J. (1970). Brunelliaceae. Flora Neotropica, 2, 1-188.
D’Amato, P. (1998). The savage garden: cultivating carnivorous plants. Ten Speed Press.
Darwin, C. (1875). Insectivorous Plants. London: John Murray.
Dickson, A. (1883, January). On the morphology of the pitcher of Cephalotus follicularis. In Transactions of the Botanical Society of Edinburgh (Vol. 14, No. 1-4, pp. 172-181). Taylor & Francis Group.
Dickison, W. C., & Rutishauser, R. (1990). Developmental morphology of stipules and systematics of the Cunoniaceae and presumed allies. II, Taxa without interpetiolar stipules and conclusions. Botanica helvetica, 100(1), 75-95.
Eichler, A.W. (1881). Uber die Schlauchblatter von Cephalotus follicular (On the trapping leaves of C. follicular). Jahrbuch des Berliner Botanish Garten 1, 193-197.
Endress, P. K., & Igersheim, A. (2000). The reproductive structures of the basal angiosperm Amborella trichopoda (Amborellaceae). International Journal of Plant Sciences, 161(S6), S237-S248.

109
Felsenstein, J. (1978). Cases in which parsimony or compatibility methods will be positively misleading. Systematic Biology, 27(4), 401-410.
Finet, C., Berne-Dedieu, A., Scutt, C. P., & Marlétaz, F. (2013). Evolution of the ARF Gene Family in Land Plants: Old Domains, New Tricks. Molecular biology and evolution, 30(1), 45-56.
Fleischmann, A., & Heubl, G. (2009). Overcoming DNA extraction problems from carnivorous plants. In Anales del Jardín Botánico de Madrid (Vol. 66, No. 2, pp. 209-215). Real Jardín Botánico.
Floyd, S. K., & Bowman, J. L. (2004). Gene regulation: ancient microRNA target sequences in plants. Nature, 428(6982), 485-486.
Floyd, S. K., & Bowman, J. L. (2007). The ancestral developmental tool kit of land plants. International Journal of Plant Sciences, 168(1), 1-35.
Floyd, S. K., Zalewski, C. S., & Bowman, J. L. (2006). Evolution of class III homeodomain–leucine zipper genes in streptophytes. Genetics, 173(1), 373-388.
Fogliani, B., Hopkins, H. C., Bouraïma-Madjèbi, S., & Medevielle, V. (2009). Morphological development of Geissois pruinosa (Cunoniaceae) from seed to adult, and the expression of plesiomorphic characters in seedlings. Flora-Morphology, Distribution, Functional Ecology of Plants, 204(1), 7-16.
Franck, D. H. (1976). The morphological interpretation of epiascidiate leaves—An historical perspective—. The Botanical Review, 42(3), 345-388.
Freeling, M. (1992). A conceptual framework for maize leaf development. Developmental biology, 153(1), 44-58.
Freeling, M., and Thomas, B.C. (2006). Gene-balanced duplications, like tetraploidy, provide predictable drive to increase morphological complexity. Genome Res. 16, 805–814.
Froebe, H. A., & Baur, N. (1988). Die Morphogenese der Kannenblätter von Cephalotus follicularis Labill (No. 3). Akademie der Wissenschaften und der Literatur.
Galbraith, D. W., Bennetzen, J. L., Kellogg, E. A., Pires, J. C., & Soltis, P. S. (2011). The genomes of all angiosperms: a call for a coordinated global census. Journal of Botany, 2011.
Garcia, D., Collier, S. A., Byrne, M. E., & Martienssen, R. A. (2006). Specification of Leaf Polarity inArabidopsis via the trans-Acting siRNA Pathway. Current Biology, 16(9), 933-938.

110
Gaudin, V., Lunness, P. A., Fobert, P. R., Towers, M., Riou-Khamlichi, C., Murray, J. A., ... & Doonan, J. H. (2000). The expression of D-cyclin genes defines distinct developmental zones in snapdragon apical meristems and is locally regulated by the Cycloidea gene. Plant Physiology, 122(4), 1137-1148.
Gleissberg, S., Groot, E. P., Schmalz, M., Eichert, M., Kölsch, A., & Hutter, S. (2005). Developmental events leading to peltate leaf structure in Tropaeolum majus (Tropaeolaceae) are associated with expression domain changes of a YABBY gene. Development genes and evolution, 215(6), 313-319.
Gould, B., & Kramer, E. M. (2007). Virus-induced gene silencing as a tool for functional analyses in the emerging model plant Aquilegia (columbine, Ranunculaceae). Plant Methods, 3(1), 6.
Graybeal, A. (1998). Is it better to add taxa or characters to a difficult phylogenetic problem?. Systematic Biology, 47(1), 9-17.
Gregory, T. R., & Mable, B. K. (2005). Polyploidy in animals. The evolution of the genome, 171, 427-517.
Ha, C. M., Jun, J. H., Nam, H. G., & Fletcher, J. C. (2007). BLADE-ON-PETIOLE1 and 2 control Arabidopsis lateral organ fate through regulation of LOB domain and adaxial-abaxial polarity genes. The Plant Cell Online, 19(6), 1809-1825.
Harrison, C. J., Corley, S. B., Moylan, E. C., Alexander, D. L., Scotland, R. W., & Langdale, J. A. (2005). Independent recruitment of a conserved developmental mechanism during leaf evolution. Nature, 434(7032), 509-514.
Hu, R., Chi, X., Chai, G., Kong, Y., He, G., Wang, X., ... & Zhou, G. (2012). Genome-wide identification, evolutionary expansion, and expression profile of homeodomain-leucine zipper gene family in poplar (Populus trichocarpa). PloS one, 7(2), e31149.
Ibarra-Laclette, E., Lyons, E., Hernández-Guzmán, G., Pérez-Torres, C. A., Carretero-Paulet, L., Chang, T. H., ... & Herrera-Estrella, L. (2013). Architecture and evolution of a minute plant genome. Nature.
Iwakawa, H., Iwasaki, M., Kojima, S., Ueno, Y., Soma, T., Tanaka, H., ... & Machida, C. (2007). Expression of the ASYMMETRIC LEAVES2 gene in the adaxial domain of Arabidopsis leaves represses cell proliferation in this domain and is critical for the development of properly expanded leaves. The Plant Journal, 51(2), 173-184.
Jain, M., Tyagi, A. K., & Khurana, J. P. (2008). Genome‐ wide identification, classification, evolutionary expansion and expression analyses of homeobox genes in rice. FEBS journal, 275(11), 2845-2861.

111
Jiao, Y., Leebens-Mack, J., Ayyampalayam, S., Bowers, J. E., McKain, M. R., McNeal, J., ... & W dePamphilis, C. (2012). A genome triplication associated with early diversification of the core eudicots. Genome Biol, 13(1), R3.
Jiao, Y., Wickett, N. J., Ayyampalayam, S., Chanderbali, A. S., Landherr, L., Ralph, P. E., ... & Leebens-Mack, J. (2011). Ancestral polyploidy in seed plants and angiosperms. Nature, 473(7345), 97-100.
Johnson, M. T., Carpenter, E. J., Tian, Z., Bruskiewich, R., Burris, J. N., Carrigan, C. T., ... & Wong, G. K. S. (2012). Evaluating Methods for Isolating Total RNA and Predicting the Success of Sequencing Phylogenetically Diverse Plant Transcriptomes. PloS one, 7(11), e50226.
Juniper, B. E., Robins, R. J., & Joel, D. M. (1989). The carnivorous plants . London, etc.: Academic Press.
Kaplan, D. R. (2001). The science of plant morphology: definition, history, and role in modern biology. American Journal of Botany, 88(10), 1711-1741.
Katoh, K., Misawa, K., Kuma, K. I., & Miyata, T. (2002). MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic acids research, 30(14), 3059-3066.
Kenrick, P., & Crane, P. R. (1997). The origin and early diversification of land plants. A cladistic study (Vol. 560). Smithsonian Institute Press Washington DC.: A cladistic study. Smithsonian Institute Press.
Kidner, C. A., & Timmermans, M. C. (2007). Mixing and matching pathways in leaf polarity. Current opinion in plant biology, 10(1), 13-20.
Ko, J. H., Prassinos, C., & Han, K. H. (2006). Developmental and seasonal expression of PtaHB1, a Populus gene encoding a class III HD‐ Zip protein, is closely associated with secondary growth and inversely correlated with the level of microRNA (miR166). New Phytologist, 169(3), 469-478.
Kouchi, H., & Hata, S. (1993). Isolation and characterization of novel nodulin cDNAs representing genes expressed at early stages of soybean nodule development. Molecular and General Genetics MGG, 238(1-2), 106-119.
Kramer, E. M. (2005). Methods for studying the evolution of plant reproductive structures: comparative gene expression techniques. Methods in enzymology, 395, 617-636.
Lamesch, P., Berardini, T. Z., Li, D., Swarbreck, D., Wilks, C., Sasidharan, R., ... & Huala, E. (2012). The Arabidopsis Information Resource (TAIR): improved gene annotation and new tools. Nucleic acids research, 40(D1), D1202-D1210.

112
Liu, T., Reinhart, B. J., Magnani, E., Huang, T., Kerstetter, R., & Barton, M. K. (2013, May). Of Blades and Branches: Understanding and Expanding the Arabidopsis Ad/Abaxial Regulatory Network through Target Gene Identification. In Cold Spring Harbor symposia on quantitative biology. Cold Spring Harbor Laboratory Press.
Lloyd, F. E. (1942). The carnivorous plants. Waltham, Massachusetts: Chronica Botanica Company.
Luo, J.H., Yan, J., Weng, L., Yang, J., Zhao, Z., Chen, J.H. et al. (2005) Different expression patterns of duplicated PHANTASTICA-like genes in Lotus japonicus suggest their divergent functions during compound leaf development. Cell Res. 15: 665–677.
Mable, B. K. (2003). Breaking down taxonomic barriers in polyploidy research. Trends in plant Science, 8(12), 582-590.
Magnani, E., & Barton, M. K. (2011). A per-ARNT-sim-like sensor domain uniquely regulates the activity of the homeodomain leucine zipper transcription factor REVOLUTA in Arabidopsis. The Plant Cell Online, 23(2), 567-582.
Magnani, E., & Hake, S. (2008). KNOX lost the OX: the Arabidopsis KNATM gene defines a novel class of KNOX transcriptional regulators missing the homeodomain. The Plant Cell Online, 20(4), 875-887.
Maher, C., Stein, L., & Ware, D. (2006). Evolution of Arabidopsis microRNA families through duplication events. Genome research, 16(4), 510-519.
Mathews, S., & Kramer, E. M. (2012). The evolution of reproductive structures in seed plants: a re‐ examination based on insights from developmental genetics. New Phytologist, 194(4), 910-923.
Matthews, M. L., & Endress, P. K. (2002). Comparative floral structure and systematics in Oxalidales (Oxalidaceae, Connaraceae, Brunelliaceae, Cephalotaceae, Cunoniaceae, Elaeocarpaceae, Tremandraceae). Botanical Journal of the Linnean Society, 140(4), 321-381.
McConnell, J. R., Emery, J., Eshed, Y., Bao, N., Bowman, J., & Barton, M. K. (2001). Role of PHABULOSA and PHAVOLUTA in determining radial patterning in shoots. Nature, 411(6838), 709-713.
Moore, R. C., & Purugganan, M. D. (2005). The evolutionary dynamics of plant duplicate genes. Current opinion in plant biology, 8(2), 122-128.
Mukherjee, K., Brocchieri, L., & Bürglin, T. R. (2009). A comprehensive classification and evolutionary analysis of plant homeobox genes. Molecular biology and evolution, 26(12), 2775-2794.

113
Mukherjee, K., & Bürglin, T. R. (2006). MEKHLA, a novel domain with similarity to PAS domains, is fused to plant homeodomain-leucine zipper III proteins. Plant Physiology, 140(4), 1142-1150.
Nelson, T., Tausta, S. L., Gandotra, N., & Liu, T. (2006). Laser microdissection of plant tissue: what you see is what you get. Annu. Rev. Plant Biol., 57, 181-201.
Ohashi-Ito, K., & Fukuda, H. (2003). HD-zip III homeobox genes that include a novel member, ZeHB-13 (Zinnia)/ATHB-15 (Arabidopsis), are involved in procambium and xylem cell differentiation. Plant and Cell Physiology, 44(12), 1350-1358.
Otto, S. P., & Whitton, J. (2000). Polyploid incidence and evolution. Annual review of genetics, 34(1), 401-437.
Prigge, M. J., & Clark, S. E. (2006). Evolution of the class III HD‐ Zip gene family in land plants. Evolution & development, 8(4), 350-361.
Prigge, M. J., Otsuga, D., Alonso, J. M., Ecker, J. R., Drews, G. N., & Clark, S. E. (2005). Class III homeodomain-leucine zipper gene family members have overlapping, antagonistic, and distinct roles in Arabidopsis development. The Plant Cell Online, 17(1), 61-76.
Richardt, S., Lang, D., Reski, R., Frank, W., & Rensing, S. A. (2007). PlanTAPDB, a phylogeny-based resource of plant transcription-associated proteins. Plant Physiology, 143(4), 1452-1466.
Robischon, M., Du, J., Miura, E., & Groover, A. (2011). The Populus class III HD ZIP, popREVOLUTA, influences cambium initiation and patterning of woody stems. Plant physiology, 155(3), 1214-1225.
Schmid, M., Davison, T.S., Henz, S.R., Pape, U.J., Demar, M., Vingron, M., Schölkopf, B., Weigel, D. and Lohmann, J.U. (2005). A gene expression map of Arabidopsis thaliana development. Nat. Genet. 37, 501-6.
Schulze, W., Frommer, W. B., & Ward, J. M. (1999). Transporters for ammonium, amino acids and peptides are expressed in pitchers of the carnivorous plantNepenthes. The Plant Journal, 17(6), 637-646.
Sessa, G., Morelli, G., & Ruberti, I. (1993). The Athb-1 and-2 HD-Zip domains homodimerize forming complexes of different DNA binding specificities. The EMBO journal, 12(9), 3507.
Siegfried, K. R., Eshed, Y., Baum, S. F., Otsuga, D., Drews, G. N., & Bowman, J. L. (1999). Members of the YABBY gene family specify abaxial cell fate in Arabidopsis. Development, 126(18), 4117-4128.

114
Simillion, C., Vandepoele, K., Van Montagu, M. C., Zabeau, M., & Van de Peer, Y. (2002). The hidden duplication past of Arabidopsis thaliana. Proceedings of the National Academy of Sciences, 99(21), 13627-13632.
Soltis, D. E., Albert, V. A., Leebens-Mack, J., Bell, C. D., Paterson, A. H., Zheng, C., ... & Soltis, P. S. (2009). Polyploidy and angiosperm diversification. American Journal of Botany, 96(1), 336-348.
Soltis, P. S., Burleigh, J. G., Chanderbali, A. S., Yoo, M. J., & Soltis, D. E. (2010). Gene and genome duplications in plants. Evolution after Gene Duplication, 269-298.
Soltis, D. E., & Soltis, P. S. (2004). Amborella not a “basal angiosperm”? Not so fast. American Journal of Botany, 91(6), 997-1001.
Soltis, P. S., & Soltis, D. E. (2013). A conifer genome spruces up plant phylogenomics. Genome biology, 14(6), 1-5.
Soltis, P. S., Soltis, D. E., & Chase, M. W. (1999). Angiosperm phylogeny inferred from multiple genes as a tool for comparative biology. Nature, 402(6760), 402-404.
Soltis, D. E., Soltis, P. S., Endress, P. K., & Chase, M. W. (2005). Phylogeny and evolution of angiosperms. Sinauer Associates Incorporated.
Soltis, D. E., Smith, S. A., Cellinese, N., Wurdack, K. J., Tank, D. C., Brockington, S. F., ... & Soltis, P. S. (2011). Angiosperm phylogeny: 17 genes, 640 taxa. American Journal of Botany, 98(4), 704-730.
Stamatakis, A. (2006). RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics, 22(21), 2688-2690.
Staudt, A. C., & Wenkel, S. (2010). Regulation of protein function by ‘microProteins’. EMBO reports, 12(1), 35-42.
Sun, C. H., Su, L. H., & Gillin, F. D. (2006). Novel plant-GARP-like transcription factors in< i> Giardia lamblia</i>. Molecular and biochemical parasitology, 146(1), 45-57.
Thien, L. B., Sage, T. L., Jaffré, T., Bernhardt, P., Pontieri, V., Weston, P. H., ... & Dupre, J. L. (2003). The population structure and floral biology of Amborella trichopoda (Amborellaceae). Annals of the Missouri Botanical Garden, 466-490.
Toriba, T., Harada, K., Takamura, A., Nakamura, H., Ichikawa, H., Suzaki, T., & Hirano, H. Y. (2007). Molecular characterization the YABBY gene family in Oryza sativa and expression analysis of OsYABBY1. Molecular Genetics and Genomics, 277(5), 457-468.

115
Troll, W. (1939). Vergleichende Morphologie der höheren Pflanzen, vol. 1 Gebrüder Borntraeger.
Tsukaya, H. (2006). Mechanism of leaf-shape determination. Annu. Rev. Plant Biol., 57, 477-496.
Van de Peer, Y., Fawcett, J. A., Proost, S., Sterck, L., & Vandepoele, K. (2009). The flowering world: a tale of duplications. Trends in plant science, 14(12), 680-688.
Waites, R., & Hudson, A. (1995). phantastica: a gene required for dorsoventrality of leaves in Antirrhinum majus. Development, 121(7), 2143-2154.
Wang, J., Zhang, G., Li, Q., Jiang, H., Liu, C., Amatore, C., & Wang, X. (2013). In vivo self-bio-imaging of tumors through in situ biosynthesized fluorescent gold nanoclusters. Scientific reports, 3.
Wenkel, S., Emery, J., Hou, B. H., Evans, M. M., & Barton, M. K. (2007). A feedback regulatory module formed by LITTLE ZIPPER and HD-ZIPIII genes. The Plant Cell Online, 19(11), 3379-3390.
Yamaguchi, T., Nukazuka, A., & Tsukaya, H. (2012). Leaf adaxial–abaxial polarity specification and lamina outgrowth: evolution and development. Plant and Cell Physiology, 53(7), 1180-1194.
Zahn, L. M., Kong, H., Leebens-Mack, J. H., Kim, S., Soltis, P. S., Landherr, L. L., ... & Ma, H. (2005). The Evolution of the SEPALLATA Subfamily of MADS-Box Genes A Preangiosperm Origin With Multiple Duplications Throughout Angiosperm History. Genetics, 169(4), 2209-2223.
Zamboni, A., Pierantoni, L., & De Franceschi, P. (2008). Total RNA extraction from strawberry tree (Arbutus unedo) and several other woody-plants. iForest-Biogeosciences and Forestry, 1(1), 122-125.
Zhong, R., & Ye, Z. H. (2007). Regulation of HD-ZIP III genes by microRNA 165. Plant signaling & behavior, 2(5), 351-353.
Zhu, Y., Song, D., Sun, J., Wang, X., & Li, L. (2013). PtrHB7, a class III HD-Zip gene, plays a critical role in regulation of vascular cambium differentiation in Populus. Molecular plant.

116
BIOGRAPHICAL SKETCH
Nicholas William Miles was born in Indianapolis, Indiana on the day of the great
mid-west blizzard of 1985. His father was a custodian at the local hospital and his
mother a sales associate at an electronics store. As a child he was very active in sports
including baseball, soccer, and golf. His first interest in science was at 10 years of age
in the areas of astronomy and chemistry, with his favorite book at the time being A Brief
History of Time by Stephen Hawking. He also learned to play his first musical
instrument, the violin, at 10, taught to him by his mother. At 14 years of age he started
his first job as dishwasher. He was rejected entry to the University of Florida for
undergraduate studies and attended Florida State University beginning in 2003. In 2007
he graduated with Honors, Cum Laude, from FSU and started graduate school at the
University of Florida. During his time at the University of Florida he applied for 9 grants,
4 of which were successfully funded, 2 of those from The National Science Foundation.
He mentored 9 undergraduate and 4 high school students from various backgrounds
and majors. And he taught 14 semesters of classes, 7 of them upper level plant classes,
2 of them vertebrate centered classes, and one of them attended by a Heisman Trophy
Winner.






![Genetic Dissection of Leaf Development inBrassica rapa · Genetic Dissection of Leaf Development inBrassica rapa Using a Genetical Genomics Approach1[W] Dong Xiao, Huange Wang, Ram](https://img.pdfslide.us/doc/110x75/5f08df687e708231d4242257/genetic-dissection-of-leaf-development-inbrassica-genetic-dissection-of-leaf-development.jpg)






















