Embed Size (px)
Citation preview

1
Evolutionfromadherenttosuspension–systems1
biologyofHEK293celllinedevelopment2
3MagdalenaMalm1*,RasoolSaghaleyni2*,MagnusLundqvist1,MarcoGiudici1,Ve-4ronique Chotteau1, Raymond Field3, Paul Varley3, Diane Hatton3, Luigi Grassi3,5ThomasSvensson1,4,MathiasUhlen1,5,6,JensNielsen1,5andJohanRockberg167*Jointfirstauthorscontributingequally891KTH-SchoolofEngineeringSciencesinChemistry,Biotechnology,andHealth,10Dept. of Protein Science, Royal Institute of Technology, SE-106 91 Stockholm,11Sweden122DepartmentofBiologyandBiologicalEngineering,ChalmersUniversityof13Technology,SE-41296Gothenburg,Sweden143 Biopharmaceutical Development, BioPharmaceuticals R&D, AstraZeneca,Mil-15steinBuilding,GrantaPark,CambridgeCB216GHUK164NBIS-BioinformaticsSystemsBiologySupport,ChalmersUniversityofTech-17nology,SE-41296Gothenburg,Sweden185NovoNordiskFoundationCenterforBiosustainability,TechnicalUniversityof19Denmark,2800KongensLyngby,Denmark206KTH - Science for Life Laboratory, Royal Institute of Technology, SE-171 21,21Stockholm,Sweden222324Correspondingauthors:25JensNielsen,Department of Biology and Biological Engineering, Chalmers Universi-26ty of Technology, 41296 Gothenburg, Sweden 27E-mail:[email protected],KTH-RoyalInstituteofTechnology,SchoolofEngineeringSci-30encesinChemistry,BiotechnologyandHealth,S-10691Stockholm,Sweden31E-mail:[email protected]:+468790998834Fax:+4685537848135
36
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

2
Abstract 1
The need for new safe and efficacious therapies has led to an increased focus on biologics 2produced in mammalian cells. The human cell line HEK293 has bio-synthetic potential 3for human-like production and is today used for manufacturing of several therapeutic pro-4teins and viral vectors. Despite this increasing popularity there is still limited knowledge 5of the detailed genetic and composition of derivatives of this strain. Here we present a 6genomic, transcriptomic and metabolic gene analysis of six of the most widely used 7HEK293 cell lines. Changes in gene copy and expression between industrial progeny cell 8lines and the original HEK293 were associated with cellular component organization, cell 9motility and cell adhesion. Changes in gene expression between adherent and suspension 10derivatives highlighted switching in cholesterol biosynthesis and expression of five key 11genes (RARG, ID1, ZIC1, LOX and DHRS3), a pattern validated in 63 human adherent 12or suspension cell lines of other origin. 1314
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

3
Introduction1Theproductionofproteintherapeuticsisafast-growingfieldasitallowsforthe2generation of sophisticatedmolecules with high specificity and activity in hu-3mans (Leader et al., 2008)(Bandaranayake and Almo, 2014)(Alex Philippidis,42017;Walsh, 2014)(AlexPhilippidis, 2017;Walsh, 2014). Theneed for proper5protein folding and glycosylation of therapeutic proteins has promoted their6productioninmammaliancells,especiallyChinesehamsterovary(CHO),butalso7humancellssuchasHEK293(Zhuetal.,2012)(Dumontetal.,2016)(Priolaetal.,82016; Sanchez-Garcia et al., 2016;Walsh, 2014). Even thoughCHO is themost9commonmammaliancell factoryused forproductionofadvancedrecombinant10proteins,thereisanincreasingdemandforimprovedandmoreefficientbiopro-11duction platforms. With an increasing number of difficult-to-express proteins12includingnext-generationbiologics, suchasbispecificantibodiesandantibody-13drug conjugates, entering clinical development, alternative or engineered ex-14pressionhostsarebeingexplored.Forinstance,extensiveomicsprofilingofthe15CHO cell has been carried out during recent years (Wlaschin et al.,162005)(Hammondetal.,2011)(Xuetal.,2011)(Brinkrolf etal.,2013)(Birzeleet17al.,2010)(Beckeretal.,2011)(Sellicketal.,2011)(Dietmairetal.,2012),which18haspavedthewayforcelllineengineeringeffortsaimingtoimprovebioproduc-19tionefficiencyandproductquality(Xiaoetal.,2014)(Leeetal.,2015)(Kildegaard20etal.,2013).Moreover,humanproductioncelllines,suchasHEK293,canserve21asconvenientexpressionhostsforproteinswithspecificrequirementforhuman22post-translational modifications (Dumont et al., 2016)(Lalonde and Durocher,232017).2425Thehumancell lineHEK293is themostcommonlyutilizedhumancell line for26expressionof recombinantproteins for various research applications. This cell27line originated from the kidney of an aborted human female embryo andwas28originallyimmortalizedin1973bytheintegrationofa4kbpadenoviral5(Ad5)29genome fragment including the genes encoding the E1A and E1B proteins, at30chromosome19(Grahametal.,1977)(Louisetal.,1997).TheexpressionofE1A31andE1BenablecontinuousculturingofHEK293cellsbyinhibitingapoptosisand32interferingwith transcription and cell cycle control pathways (Berk, 2005). In33addition,sinceE1AandE1Bareessentialhelperfactorsforadenoassociatedvi-34rus(AAV)production,thecontinuousexpressionofthesegenesmakesHEK29335cells attractive production hosts for recombinant AAV particles (Clément and36Grieger,2016)..ThegenomesandtranscriptomicprofilesofsixdifferentHEK29337cell lineswererecentlymapped,confirmingpreviousobservationsofapseudo-38triploidgenomewiththeadenoviralDNAinsertedonchromosome19(Y.C.Lin39etal.,2014)(Bylundetal.,2004)(Louisetal.,1997).Furthermore,itwasshown40thatstandardlaboratoryculturingofthecelllinesinmostcasesdidnotchange41the overall genomic composition of the cells although the organization of the42HEK293 genome is continuously evolving through the events of chromosomal43
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

4
translocationsandcopynumberalterations(Y.C.Linetal.,2014).However,the1chromosome number of HEK293 cells derived from different commercial2sources has been reported to show a high degree of variation, suggesting that3long-term cultivation and subcloning of cells result in karyotypic drift (A.A.4Stepanenko andDmitrenko, 2015). Such abnormalities and genomic instability5is,however,characteristicforimmortalizedcellsandhavealsobeenreportedfor6CHOcells(A.A.StepanenkoandDmitrenko,2015)(Väremoetal.,2014)(Vcelaret7al.,2018)(Wurm,2013).89SeveralHEK293celllineshavebeenestablishedfromtheparentalHEK293line-10age. Strategies to improve recombinantprotein expression include the genera-11tionoftransformedHEK293cellsconstitutivelyexpressingthetemperaturesen-12sitive allele of the large T antigen of Simian virus 40 (293T), (DuBridge et al.,131987)or theEpstein-Barr virusnuclear antigenEBNA1 (293E) (Murphyet al.,141992)(Swirskietal.,1992).Celllinesexpressingtheseviralelements,enableepi-15somalreplicationofplasmidscontainingtheSV40originofreplication(293T)or16EBV oriP (293E). One 293E cell line is host for the production of dulaglutide17(TRULICITY®)(Lalonde and Durocher, 2017). In addition, several HEK293 cell18lineshavebeenadaptedtosuspensiongrowthinserum-freemediumathighcell19density (Graham,1987)(Garnieret al., 1994)(Côtéet al., 1998), enabling large-20scale cultivation and bioproduction in bioreactors. Two industrially relevant21suspensioncelllinesare293-Fand293-H(Gibco,ThermoFisherScientific).The22293-Fclonewas initially isolatedby limitingdilutionofHEK293whereaclone23withfastgrowthandhightransfectivitywasobtained,whichwaslaterrecloned24andsubjected to suspensiongrowthadaptation in serum-freemedium(Figure251A).Interestingly,the293-Hcelllineisasuspensiongrowthadaptedcloneorig-26inally derived fromamore adherentHEK293 cell clone. The final 293-H clone27has high adherence during plaque assays, fast growth, high transfectivity and28productivity(Figure1A).The293-Fcelllineistodayusedfortheproductionof29the regulatory approved rhFVIII (NUWIQ®), whereas 293-H cells are used for30production of rFVIIIFc (ELOCTATE®) and rFIXFc (ALPROLIX®)(Dumont et al.,312016)(LalondeandDurocher,2017).Furthermore, theFreestyle293-Fcell line32has been generated by adaptation of 293-F cells to Freestyle medium (Gibco,33Thermo Fisher Scientific). Even though each of the above-mentioned cell lines34are all derived from the same originalHEK293 strain, significant genomic and35transcriptomic changes between parental and progenitor cell lines can be ex-36pectedduetothegenomicinstabilityofHEK293asdiscussedabove.3738DespiteextensiveusageofCHOandHEKinbothsuspensionandadherentmode39and several empirical protocols for adaptation in either direction, molecular40knowledgeof thekeygenes involved inthetransitionbetweenthetwogrowth41statesislimited.Whileadherentcellshavetraditionallybeenwidelyusedforthe42productionof viruses, e.g.AAVand lenti virus for clinical research, suspension43
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

5
cellsaretheplatformofchoiceforthebioproductionoftherapeuticproteins.The1main advantages of suspension cell bioproduction schemes are growth at very2high cell densities in serum-free medium at scale, enabling higher volumetric3yields.Easyadaptationbetweencellulargrowthinadherenttosuspensionmode4hastraditionallybeenakeyfeaturewhenselectingstrainsforbioprocessappli-5cations, including usage of CHO and HEK293. Whereas certain experimental6steps aremore efficient in adherentmode, e.g. chemical transfection and viral7infection,theabilitytoincreasethevolumetriccelldensitybygrowthinsuspen-8sionwithoutcellclumpformationthatresultsinoxygenlimitationsisakeystep9fromamanufacturingperspective.1011Here,wepresentagenomicandtranscriptomicanalysisoftheHEK293parental12celllinealongwithfivewidelyusedHEK293derivatives(Figure1A).Anoverall13analysisofthedifferencesingenomiclandscapeandtranscriptomicprofileswas14performedinordertoprovidenovelmolecularinsightsintothedifferencesbe-15tweencelllinesthathaveoccurredduringtheprocessofclonalisolationandex-16pansion.Furthermore,wefocusontranscriptomicdifferencesbetweenadherent17and suspension HEK293 cells and the impact of the differentially expressed18genesonmetabolicpathwaysandthephenotypeof thecells fromabioprocess19perspective.2021
Results2223Overallgenomicand transcriptomicprofilesemphasizeclonaldivergence24between all theprogeny cell lines compared to theparentalHEK293 cell25line26Inthisstudy,six industriallyrelevantHEK293cell lines(Figure1A)weresub-27jected toomicsprofiling.Thissetof cell lines includes theparentalHEK293as28wellasfiveadditionalcelllinesthathaveallbeenclonallyderivedfromparental29HEK293cells.Thecelllinescanbedividedintoeitheradherent(HEK293,293E30and293T)orsuspension(293-H,293-FandFreestyle293-F)cells.Thegenomes31andthetranscriptomesofthesesixcelllinesweresequencedusingIlluminaHiS-32eq.Table S1 provides full results of transcript levels (TPM) for all cell lines.33Comparisons of the genomes and transcription profiles between the cell lines34showoverallsimilarresults(Figure1Band1C).Hierarchicalclusteringdivided35the progeny cell lines into two different taxonomic groups, of either adherent36(293T, 293E) or suspension cell lines (293-H, 293-F and Freestyle 293-F), di-37verged from the parental HEK293. Interestingly, the original HEK293 cell line38wasthemostdistantfromallothercelllines.Asexpected,thetwo293-Flineages39(293-FandFreestyle293-F)showedverysimilarprofiles.Thesamepatternof40geneexpressionclusteringwasvisualizedbyprincipalcomponentanalysis(Fig-41ure 1D), where the suspension cell-lines grouped together in the plot, with a42
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

6
verycloseclusteringof293-FandFreestyle293-Fcells.Ontheotherhand,the1adherentcell-linesHEK293EandHEK293Tshowedlargervariationsingeneex-2pressionpatternsbetweencelllines.TheparentalcelllineHEK293showedano-3table difference in transcriptome profile compared to all the other cell lines4along the firstprincipalcomponent (PC1).Theseresults indicateagenomicdi-5vergence of the clonal lineages compared to the parentalHEK293 and suggest6thepresenceofsimilartranscriptomictraitsbetweenHEK293progenycelllines7individuallyselectedforduringtheisolationofeachclone.Hierarchialclustering8ofthecelllinesbasedonSNPsgaveasimilartrendwithhighsimilaritybetween9293-FandFreestyle293-Fcelllines,whereastheparentalHEK293celllinehad10lowersimilaritiestotheprogenythecelllines(FigureS1A).However,adifferent11patternofoverallclusteringwasobserved,withtheoriginalHEK293and293E12celllinesseparatedfromtherestonaseparatebranchandthelowestsimilarity13scoresobservedforthe293Ecelllinecomparedtotherest.1415TheHEK293cell linewasoriginally immortalizedbytherandomintegrationof16viralgenomicDNAofadenovirus5(Grahametal.,1977),whichincludestheE1A17andE1Bgenes.Inthisstudy,overallhighmRNAlevelsofE1AandE1Bwereob-18served in allHEK293 cell lines (Figure 1E),with significantly (p<0.05) higher19expressionlevelsofE1AinHEK293,293Eand293-Hcelllinesovertheothercell20lines, whereas 293F had a significantly (p<0.05) higher E1B expression com-21pared to 293H and Freestyle 293-F (Figure S1B). No other significant differ-22enceswereobserved forE1B.Furthermore, the293Tand293Ecell lineswere23originally generated by the transfection and stable integration of plasmids ex-24pressing the SV40 Large T antigen and EBNA-1, respectively. As expected, the25geneexpressionofLargeTandEBNA-1wasdetectedin293Tand293E,respec-26tively(Figure1E).Interestingly,expressionoftheLargeTantigenwasalsoob-27servedin293E,whichisnotreportedbythesupplier(ATCC).Thepresenceofa28truncatedversionofLargeTinthe293Egenomewasconfirmedbydenovoas-29sembly of all reads notmapping to the human reference genome (Figure S1).30Tracingtheoriginofthe293Ecellline(Swirskietal.,1992),theLargeTexpres-31sion of 293Emay be derived from the pRSVneo plasmid thatwas used to co-32transfectHEK293cellsalongwiththepCMV-EBNAplasmidforthegenerationof33the stable EBNA-1 expressing clone (293c18) byG418 selection. The pRSVneo34plasmid contains a truncated version of the Large T gene (according to the35AddGenevectorDatabase),whichalignsperfectlywiththetruncatedLargeTse-36quencefoundinthe293Egenome(FigureS1C).3738CopynumbervariationbetweentheparentalHEK293cell lineand itsde-39rivativesconfirmedthegenomicinstabilityoftheHEK293strainandiden-40tified loci with conserved copy number gain/loss between progeny cell41lines42
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

7
InordertoevaluatethegenomicvariationbetweenHEK293anditsderivatives1further, overall genomic copy number variation of all progeny cell lines com-2paredtotheparentalHEK293wasperformed.Acomparisonofgainedandlost3regionsonallchromosomesbetweenallcelllinescanbefoundinFigure2and4TableS2.Interestingly,aconservedpatternofcopynumbergainorlossoflarge5regionshasoccurredonseveralchromosomesofallHEK293progenycellscom-6paredtoHEK293,whereasotherchangesaremorelocalorcelllinespecific.For7instance,onchromosome13,aregionof>15Mb(Figure2)hasbeenamplified8significantly in all cell lines compared to the parental HEK293 strain. All ele-9mentswithcopynumbergainof>1log2fold-changecommontoallprogenitor10cellsarelocatedinthisregion(TableS2).Amongstthese,fouroutofsevenpro-11tein-codinggenes (BORA,MZT1,PIBF1andKLHL1)belong to the cytoskeleton12geneset(GO:0005856).Onchromosome18,thereisaconservedpatternofcopy13numberlossofmostofthechromosomesequenceforallprogenycelllinescom-14paredtotheparentalHEK293strain,withtheexceptionofahighdegreeofcopy15number gainof a region close to the centromere for all cell lines except293E.16Withintheregionofconservedgain,severalgenesencodingcelladhesionmole-17culeswithinthedesmocollin(DSC)anddesmoglein(DSG)subfamilies,belonging18to the cell-cell adhesion gene set (GO: 0098609), are located.When analyzing19morelocalcopynumbervariationsbetweenprogenycell linesandtheparental20strain,someinteresting lossorgainof fullorpartialelementscomparedtothe21parentalHEK293was identified. For instance, copynumber losswasobserved22for the fumarate hydratase (FH) gene, which has previously been reported to23havelostseveralgenecopiesinHEK293andhencebeenhypothesizedtoplaya24roleinthephenotypictransformationofHEK293(Y.-C.Linetal.,2014).Interest-25ingly, the fumarate hydratase gene along with the neighboring kynurenine 3-26monooxygenase(KMO)gene,hadalog2-foldcopyratioof<-1in293T,293-Fand27Freestyle293-FcelllinescomparedtotheparentalHEK293(Figure2andFig-28ureS2A),suggestingthatthesecellshavehalfthenumberofcopiescomparedto29theparentalcellline.Moreover,the293Tand293-Hcelllineshaveagainofthe30genomiclocisurroundingtheFHgene,whilemaintainingthecopynumberofthe31FHgenecomparedtoHEK293. Interestingly, theresultingFHexpression levels32ofthecelllinesonlypartlycorrelatedwiththegenecopynumberchanges(Fig-33ure 2 and Figure S2A). Even though the gene copy number of the parental34HEK293strainisthesameasfor293Tand293-Hlineages,theFHmRNAlevels35of HEK293was as low as the expression levels of the lineages with only half36numberofFHgenecopies.Moreover,theexpressionlevelsofKMOwascompa-37rably lowinallcell linesbutdidnotcorrelatewithgenecopynumber.Besides38thechangesingenecopynumberoftheFHloci,alociaroundthetransducin-like39enhancerprotein4(TLE4)gene,encodingatranscriptionalco-repressorofWnt40signalingpathwaymembers,andthenon-codingRNALINC01507wasfoundto41have a log2-fold copy number gain of >1.5 in all progeny cell lines except for42293E(Figure2).ThisgainintheTLE4lociwasaccordinglyreflectedinthetran-43
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

8
scriptionlevelofthegenewithahigherlevelofexpressionin293T,293-H,293-F1andFreestyle293-Fcomparedto293EandHEK293(FigureS2A).Inaddition,a2majorlossofcopynumberoftheADAM3Apseudogenewasobservedforallcell3lines accept 293-Hwith amaintained lowor no expressionof thepsuedogene4observedinthecelllines(Figure2andFigureS2AB).56Due to theobservedpatternof commongenomicchanges toprogenycell lines7compared to theparentalHEK293,anevaluationof commonSNPsamongstall8progeny cell lines but not HEK293was performed. GO enrichment analysis of9commongeneswithhighormoderate impactSNPsdifferent inallprogenycell10linescomparedtotheoriginalHEK293(TableS3),showedsignificant(adjusted11p-value <0.05) enrichment of homophilic cell adhesion via plasma membrane12adhesion molecules (GO:0007156; adjusted p-value 0.025; fold enrichment1310.26; data not shown) and cell-cell adhesion via plasma-membrane adhesion14molecules(GO:0098742;adjustedp-value0.032;foldenrichment7.53;datanot15shown).AllgeneswithmoderateorhighimpactSNPsinprogenycelllinescom-16pared to HEK293 found amongst both these GO-terms were protocadherins17(PCDH12, PCDHB10, PCDHB13, PCDHB15, PCDHB16 PCDHGA2, PCDHGA3 and18PCDHGB2).Inaddition,theTeneurin-2gene(TENM2)(withinGO:0098742)also19hadan altered SNPallele in all progeny cell lines compared toHEK293.These20SNPs all result inmissensemutations with unknown biological impact on the21geneproducts.However, theenrichmentof commonSNPswithin thisgroupof22genes in all HEK293 progeny cell linesmay suggest an impact on the protein23functionanda selectiveadvantageof suchphenotypicchangesduringcontinu-24ouscelllinecultivation.2526Differential expression between all progeny cell lines and the parental27HEK293revealedconsensuschangesingeneexpressionofcellularcompo-28nent organization and integral components of the plasma membrane29amongstprogenycelllines30Based on the overall genomic and transcriptomic profiles of the different31HEK293celllines,theparentalHEK293strainstoodoutasdifferentcomparedto32allothercelllines.Inordertoevaluatecommonchangesbetweenallprogenycell33linesandtheparentalHEK293,differentialexpressionanalysiswasperformed.34Resultsshowedasignificantconsensusofdown-regulationofgenesinvolvedin35extracellularmatrixorganization,locomotionandcelladhesioninprogenycells36comparedtotheparentalHEK293strain(Figure3A).Moreover,aminoacidme-37tabolismandmetabolicprocessof smallmoleculeswere foundup-regulated in38allprogenycell-lines.Alongwithchangesinextracellularmatrixgenes,thereis39alsoaconsensusamongstprogenycelllinescomparedtoHEK293ofdifferential40expressionofgenes involved inother typesofcellularcomponentorganization41suchascellmorphogenesis, cytoskeletonorganization,membraneorganization42and cell junction organization. A comparison between gene expression fold43
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

9
changesandcopynumbervariationof thedifferentiallyexpressedgenes(log2-1fold change >+/- 1) for each progeny cell line compared toHEK293 showed a2trend of enrichment of gained gene copies amongst genes with up-regulated3mRNAlevels(FigureS2B).However,therewasnotacleartrendoflossingene4copynumberamongsttranscriptionallydown-regulatedgenesforallcelllines.56For further evaluation of the transcriptomic similarities and changes between7HEK293celllines,pairwisedifferentialexpressioncomparisonsbetweenallcell8lineswereperformed.Asexpected,theparentalcelllinehadthehighestnumber9ofdifferentiallyexpressedgeneswhencompared toallother cell lines (Figure103B andTable S4). In addition,when looking at differentially expressed genes11uniquetocertaincomparisons,thelargestgroupofgeneswerefoundcommonto12all pairwise comparisons between HEK293 and each of the progeny cell lines13(greenbarinFigure3B),againemphasizingarelativelyhighdegreeofcommon14transcriptomic changes amongst progeny cell lines differentiated from the pa-15rentalHEK293.Astheprogenycelllineshadanenrichmentofdifferentiallyex-16pressed genes associated with cellular component organization compared to17HEK293, we sought to evaluate to what cellular compartments the 329 genes18commontoallpairwisecomparisonsbetweenHEK293andprogenycelllineslo-19calize. In line with the overall differential expression evaluation (Figure 3A),20whichemphasizedchangesinforinstancecelladhesionandextracellularmatrix21organization,therewasasignificant(padj<0.05)enrichmentofgenesrelatingto22the integral compartment of plasma membrane (GO:0005887) amongst the23commondifferentiallyexpressed(DE)genesuniquetothecomparisonsbetween24HEK293andallprogenycelllines(Figure3C).2526Differential expression between suspension and adherent HEK293 cell27linesidentifiedkeychangesrelatedtocholesterolmetabolism28Thegrowthmorphologyofbioproductioncelllinesisofgreatimportanceforcul-29turemaintenanceandefficiencyofindustrialbioprocessing.Inordertolookinto30geneexpressionvariations correlatingwith adherent and suspensionHEK29331cell lines, differential expression analysis between adherent and suspension32HEK293progenycelllineswasperformed.Asresultsfromtheoverallcompari-33sonoftranscriptomicprofilesoftheHEK293celllinesshowedthattheparental34HEK293 cell line is highly differentiated from all of the progeny cell lines and35moreover,thattheFreestyle293-Fcelllineisverysimilartothe293-Fcellline,36HEK293andFreestyle293-Fwereexcludedfromthisanalysis,soasnottoskew37thedata.Enrichmentanalysisofthedifferentiallyexpressedgenesbetweenad-38herent (293T and293E) and suspension (293-H and293-F) progeny cell lines39showedsignificantexpressiondifferencesofsimilargenesetsasinthecompari-40son betweenprogeny cell lines and the parentalHEK293 (Figure 3A and 4A,41Table S5).For instance, thesuspensionprogenycell lineshadasignificantup-42regulation of gene sets involved in cellular compartment organization such as43
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

10
cellmorphogenesis, cell junctionorganization,cellmembraneorganizationand1cytoskeletonorganization.Interestingly,thereisnosignificantchangeintheex-2pression of the extracellularmatrix organization gene set between suspension3andadherentHEK293progenycelllines.Perhapsasexpected,therewassignifi-4cant differential expression observed for the cell adhesion, cell differentiation,5cell morphogenesis and cell motility gene sets. Interestingly, all of the above-6mentioned gene categories, including cell adhesion, were up-regulated in sus-7pensionHEK293cellsascomparedtoadherent.Whenlookingatthemostsignif-8icantlydifferentiallyexpressedgenes(adjustedp-value<0.01)amongstthecell9adhesiongeneset,manygenesofthecadherinsuperfamilyofcelladhesionmol-10eculeswere foundup-regulated in suspension cell lines compared to adherent11HEK293progenycells(TableS6).1213In order to evaluatewhatmetabolic impact the differentially expressed genes14mayhaveoncells insuspensioncompared to theadherentstate,agenerichu-15manmetabolicmodel,HMR2(Mardinogluetal.,2014),wasused togeneratea16setofmetabolicgenesandtheirassignedpathwaystofindmetabolicpathways17withalteredexpressionbetweenadherentandsuspensionHEK293progenycell18lines.AsshowninFigure4B,pathwaysrelatedtoaromaticaminoacidsandoxi-19dative phosphorylation were significantly down-regulated in suspension cells20comparedtoadherentandhadamongstthehighestnumberofdifferentiallyex-21pressedgenes.Inaddition,pathwaysrelatedtoretinol,linoleateandnucleotide22metabolismwerealso significantlydown-regulated in suspension cell-lines.On23the other hand, biosynthesis andmetabolism of cholesterol were found to be24most significantly up-regulated amongst metabolic pathways in suspension25compared to adherent cells. In addition, pathways related to proteinmodifica-26tionandfattyacidmetabolism(omega-3/6fattyacidmetabolism,fattyacidde-27saturation, fatty acid biosynthesis and fatty acid elongation) were also up-28regulated in suspension compared to adherent HEK293 progeny cell lines. All29resultsfromthemetabolicgenesetanalysisareprovidedinTableS7.3031Focusingon thepairwisecomparisonsbetweenHEK293cell lines,38differen-32tially expressed genes were identified common to all adherent to suspension33pairwisecomparisons(redcolumninFigure3B,Figure5A,TableS4).Looking34intopotentialunderlyinggenomicalterationsof thesegenes,nohigh impactor35moderatesinglenucleotidepolymorphism(SNP)wasdetectedinanyofcelllines36that could explain the differential expression (data not shown). Three of the37genes (ARRDC3, HMGCS1 and PCYOX1L) had the same directional gene copy38numbervariation (gainor loss) compared togeneexpression fold-changes (up39ordown)ofallsuspensioncomparedtoadherentcells,whichmayatleastpartly40explainthedifferentialexpressionofthesegenesbetweenthegroups.Geneen-41richmentanalysisofthissetof38differentiallyexpressedgenesbetweenadher-42entandsuspensioncells,performedusingEnrichr(Kuleshovetal.,2016)(Chen43
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

11
etal.,2013),predictedthecholesterolbiosyntheticprocesspathwayasthecellu-1larpathwaymostaffectedbythisexpressionvariation(Figure5B).Thisresult2furtheremphasizesthedifferentialexpressionbetweenadherentandsuspension3cellsofgenesinvolvedinthecholesterolpathway,mentionedabove.Amongthe438 common differentially expressed genes MSMO1, IDI1, NPC1L1, INSIG1 and5HMGCS1 are directly related to cholesterol biosynthesis (GO:0006695 and/or6GO:0008203,Figure5B).Eachofthesegeneshadatleastatwo-foldincreasein7expressioninthesuspensioncellscomparedwiththeadherentcell-lines.Based8onthesefindings,wesoughttopredicttheeffectofthedifferentiallyexpressed9genes between adherent and suspension HEK293 cell lines on the cholesterol10biosynthesis of the cells using Ingenuity pathway analysis (IPA). Although the11MSMO1,IDI1andHMGCS1geneswereallup-regulatedin293-Fand293-Hcom-12pared toHEK293, the down-regulation of the lathosterol oxidase gene (SC5D),13whichgeneproductisindownstreamstepsofthepathway,resultedinapredict-14ed reduction in cholesterol production in 293-F and 293-H cells compared to15HEK293 (Figure S3). Comparisons between suspension cell lines (293-F and16293-H)andadherentprogeny cell lines (293Eand293T)didnot result in any17predictedchangesincholesterolbiosynthesis(datanotshown).1819Asfouroutofthe38differentiallyexpressedgenes(LOX,SMAD7,ID1&TXNIP)20have previously been shown to have a role in the epithelial to mesenchymal21transition(EMT)pathway(Zhaoetal.,2015),wesoughttoevaluatetheroleof22thispathwayinthetransitionfromadherenttosuspensioncellgrowthofthese23HEK293cell lines.ThenormalizedexpressionofasetofEMTmarkersshowed24thattheparentalHEK293actuallyhadthehighestlevelofexpressionofvarious25mesenchymalmarkers (N-cadherin,vimentinand fibronectin)ofall thesixcell26lines(FigureS4A).Moreover,whenpredictingEMTpathwayoutcomesforsus-27pension cells (293F and293H) compared to theparentalHEK293 strainusing28Ingenuitypathwayanalysis(IPA),theresultspredictedreducedEMTinthesus-29pensionprogenycellscomparedtoHEK293(FigureS4B).However,suspension30cellswerepredictedtohaveincreaseddisruptionofadherencejunctions,which31isconsistentwiththesuspensioncellphenotype.Takentogetherthecomparison32between adherent and suspensionHEK293 progeny cell lines suggest that the33transitionbetweenadherent to suspensioncell growth isnotequivalent to the34epithelialtomesenchymaltransitioneventhoughseveralEMT-associatedgenes35maybekeytothedifferencebetweencelllines.Insteadkeychangeswerefound36associatedwithcholesterolbiosynthesisandfattyacidmetabolism.3738Extended validation of the 38 consistently differentially expressed genes39betweenadherentandsuspensionHEK293cell linesinasetof63human40cell lines identified fivekey genespotentially associatedwithdifferences41betweenadherentandsuspensionphenotypes42
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

12
For identificationofkeygenes involved in the transition fromadherent tosus-1pensionmorphology,expressiondatafromanadditionalsetof63differenthu-2mancelllinesdepositedintheHumanProteinAtlasdatabase(Uhlenetal.,2017)3wereanalyzed.Principalcomponentanalysisofthesecelllinesresultedinclus-4teringof suspension cell lines in adistinct group separated fromadherent cell5lines (Figure 6A).However, sincemostof thesuspensioncell linesareof lym-6phoidormyeloidoriginthisclusteringmaybearesultofasimilaroriginofsus-7pensioncelllines.Transcriptiondataofthe38previouslyidentifieddifferentially8expressedgenesfrom47adherentand16suspensioncelllines(TableS8)was9comparedbetweenthetwogroupsusingaMann-WhitneyU-test.Withinthisset,10nine genes (LOX, ID1, ADAMTS1, ZIC1, KCNMA1, DHRS3, RARG, COL4A6 and11ARRDC4)hadsignificantdifferentexpressionlevelsbetweenadherentandsus-12pensioncelllineswithp-values<0.01(Figure6Band6C).Fourofthesegenes13(ADAMTS1,KCNMA1,COL4A6andARRDC4)hadtheoppositedirectionalchange14in the extendeddata set compared to thedifferential expressionbetweenonly15HEK293 strains. Based on these findings, the remaining five genes (LOX, ID1,16ZIC1,DHRS3andRARG),whichshowedaconsistentdown-regulationinsuspen-17sioncelllinescomparedtoadherentcells,mayplayimportantrolesinthemor-18phologicaldifferencesbetweentheadherentandsuspensioncelllines.1920Methods2122CellcultivationforDNAandRNApreparation23Theadherentcell linesHEK293(ATCC-CRL-1573),HEK293T(ATCC-CRL-3216)24and 293E (ATCC-CRL-10852) were obtained from ATCC and propagated in25DMEM(D6429)supplementedwith10%FBSat37°Cinahumidifiedincubator26with 5% CO2 in air. Suspension cell lines 293-F, 293-H and FreestyleTM 293-F27(Gibco)wereobtainedfromThermoFisherScientificandcultivatedin293SFM28IImedium(Gibco)supplementedwithGlutamaxatafinalconcentrationof4mM29(Gibco). Suspension cells were cultivated in 125-ml Erlenmeyer shake flasks30(Corning)at37°Cand120rpminahumidifiedincubatorwith8%CO2inair.All31cellswerepropagatedfromfrozenstocksfornolongerthan20passages.3233RNAandDNApreparationandsequencing34Adherentcellsweredetachedbytrypsinizationandbothadherentandsuspen-35sion cellswereharvestedby centrifugation.GenomicDNAwasextractedusing36theBloodandCellCultureDNAMiniKit(Qiagen)accordingtothemanufactur-37er’s guidelines and concentrationswere determinedby using aNanoDropND-381000 spectrophotometer (Thermo Scientific). Genome sequencing was per-39formedattheNationalGenomicsInfrastructure(Scilifelab,Solna,Sweden)using40theIlluminaHiSeqXplatform.ForRNAextraction,cellsgrowninlogphasefrom41three biological replicates were collected (derived from successive propaga-42tions).CellpelletswereresuspendedinRNAlaterStabilizationSolution(Invitro-43
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

13
gen)accordingtothemanufacturer’srecommendationsuntilRNAextraction.To-1talRNAwasextractedfromthreereplicatesofeachcelllineusingQiagen’sRNe-2asy plusMini Kit according to themanufacturer’s instructions. Concentrations3weredeterminedwithaNanoDropND-1000spectrophotometerandRNAquali-4tywasassessedona2100Bioanalyzer(AgilentTechnologies)usingRNA60005Nanochips(AgilentTechnologies).AllsampleshadanRNAintegritynumberof6atleast9.9.RNAsequencingwasperformedatGATC(Konstanz,Germany)using7theInviewTranscriptomeAdvanceserviceandanIlluminaHiSeqinstrument.89DNA-sequencinganalysis10Genomesequencingreadswerealignedtothereference(human_g1k_v37.fasta)11usingbwa (0.7.12) (Li andDurbin, 2010).The rawalignmentshave thenbeen12deduplicated, recalibrated and cleaned using GATK (version 3.3-0-geee94ec,13gatk-bundle/2.8) (McKennaetal.,2010).Qualitycontrol informationwasgath-14eredusingQualimap (v2.2) (Okonechnikov et al., 2016). SNVs and indels have15beencalledusingtheHaplotypeCaller.Thesevariantswerethenfunctionallyan-16notatedusingsnpEff(4.1).ThePiperpipelinefromtheNationalGenomicsInfra-17structure was used. The correlation between BAM files was assessed using18multibamsummaryanditsplotCorrelationfunctionfromdeepTools2(Ramírezet19al.,2016).Spearmanwasusedtocalculatecorrelationcoefficientsbetweensam-20ples,andtheclustersarejoinedwiththeNearestPointAlgorithm.AnSNPcorre-21lationheatmapwas generatedusing seqCAT (Fasterius andAl-Khalili Szigyarto,222018).Theheatmapwasbasedonthesimilarityscores(Yuetal.,2015)betweenthe23cell lines andeuclideandistances.To compare theLargeTantigen sequencesof24293T and 293E, unmapped reads were extracted to new bam files using25SAMtools(Lietal.,2009),convertedto fastqwithBEDTools(QuinlanandHall,262010),anddenovoassembledwithMEGAHIT(Lietal.,2016).NCBIBLASTwas27used to identify theLargeTantigen in theassembled contigs.Toevaluateand28visualizecopynumbervariations,CNVkit(Talevichetal.,2016)wasusedwithits29whole-genome sequencing method, cbs segmentation (Olshen et al.,302011)(VenkatramanandOlshen,2007)andtheHEK293alignmentasreference.31GOenrichment analysis of geneswithhighormoderate impact SNPswasper-32formedusingPANTHERclassificationsystem(Mietal.,2019).3334RNA-sequencingdata35Kallisto(Brayetal.,2016)wasusedtoquantifytranscripts,andthetranscripts36weremappedto thehumanGrCh37genome.Log transformednormalizeddata37byDESeq2wasusedforcelllineclusteringandcalculationofEuclideansample38distances.SignificanttestingofdifferentialmRNAexpressionofE1A/Belements39wasdonebyWelsh twosample t-test.Fordifferentialexpressionanalysis, raw40countdata fromKallistowas importedusing the tximportpackage(Sonesonet41al., 2015) and analyzedwithDESeq2 (Love et al., 2014). In thedifferential ex-42pression analysis, all suspension cell-lineswere compared to all adherent cell-43
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

14
lines, and additionally, all pairwise combinations between suspension and ad-1herent cell-lines were evaluated. The expression comparison of the viral ele-2mentswasbasedonnormalizedcountsfromDESeq2.Forevaluatingdifferential3expressionof38commondifferentiallyexpressedgenesbetweenadherentand4suspensionHEK293celllinesinasetof63humancelllines,RNA-seqdatafrom5each cell line deposited in the HPA database was used. Based on the growth6characteristics, cellsweredivided into twogroupsof adherent and suspension7cells.AMann-WhitneyU-testwasusedtocomparenormalizedcountsbasedon8library sizebetweenthe twogroups foreachof the38differentiallyexpressed9genes.1011GeneSetanalysis12Todiscoversignificantalterationsofgenesetsandmetabolicpathwaysbetween13HEK293cell lines, thePianopackage inRwasused (Väremoetal., 2013).The14adjustedp-valuesandfoldchangesfromthedifferentialexpressionwasusedin15combinationwithagenesetcollectionbasedon“goslim_genericBiologicalPro-16cess”.Theheatmapfortheprogenycellslinesvs.HEK293wasbasedonthecon-17sensus score from all pairwise gene set statistics calculations with Wilcoxon18rank-sum test. The heatmap for suspension cells (293-H, 293-F) vs. adherent19cells(293E,293T)wasbasedontheconsensusscorefromgenesetstatisticscal-20culationswithmean,median,sum,stoufferandtailStrengthtests.Toproducethe21networkplot,genesetswereexportedfromHMR2(Mardinogluetal.,2014).For22finding differentially expressed pathways of genes between adherent and sus-23pensioncelllines,weusedWilcoxonasstatisticaltestandfilteredgenesetswith24adjustedp-value lower than0.05assignificantlychanged. Inaddition, forgene25setanalysisof38commonDEgenesbetweenadherentandsuspensioncelllines26weusedEnrichRandGObiologicalprocessasgenesetcollection(Kuleshovetal.,272016).2829Ingenuitypathwayanalysis30Inordertopredictthepathwaychangesbetweencelllinesbasedondifferential-31lyexpressedgenesfrompairwisecomparisons,ingenuitypathwayanalysis(IPA,32QIAGENInc.,)wasperformed.Toconsiderageneasdifferentiallyexpressedwe33usedlog2foldchange>1or<1andadjustedp-value<0.05.Forfilteringresultsof34genesetanalysisbyIPAweusedBenjamini-Hochbergmultipletestingcorrected35p-valueslowerthan0.05tofindgenesetswithadifferentexpressionpattern.36
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

15
Discussion1Due to the extensive usage of HEK293 cells as a bioproduction platform for2pharmaceutical proteins and AAV vectors, characterization of the HEK293 ge-3nome and transcriptome is relevant for bioprocess development. A deeper4knowledge of the HEK293 genomic and transcriptomic traits can for instance5pave theway formorerational cell lineengineeringapproaches,aiming to im-6prove bioproduction efficiency and quality of protein products. As different7HEK293lineagesarepropagatedunderdifferentconditionsandtheobservation8thatimmortalizedcontinuouslyculturedcelllines,suchasHEK293,haveahigh9degreeofgenomic instabilitywith frequentchromosomerearrangements(Y.C.10Linetal.,2014)(A.A.StepanenkoandDmitrenko,2015)itcanbeexpectedthat11differentHEK293lineagesaredifferentiatedatthegenomicandtranscriptomic12levelcomparedtotheparentalcellline.Here,thegenomesandtranscriptomesof13theoriginalHEK293alongwithfiveprogenycelllineageswereanalyzed(Figure141A).Theoverall comparisonof genomicand transcriptomicprofiles confirmed15the picture of clonally diverged progeny cells as compared to the parental16HEK293.Acommonhierarchicalclusteringbetweenthecelllineswasobserved17forbothgenomicandtranscriptomiccomparisonswiththeparentalHEK293cell18line,theprogenysuspensioncelllinesandtheprogenyadherentcellsclustering19onthreeseparatebranches(Figure1Band 1C).Asexpected, therewasahigh20degree of genomic and transcriptomic similarities of the Freestyle 293-F and21293-Fcelllines(Figure1B,1Cand1D).Itisunclearwhetherthesetwocelllines22are actually the same clone or if the Freestyle 293-F cell lineagewas isolated23throughsub-cloningof293-F.Theresultspresentedhereindeedshowthatthey24arehighlysimilarbothonagenomicandtranscriptomiclevelandconfirmsthe25previously reported findings that standard propagation of HEK293 cell lines26doesnotalter thegenomicprofile toa largeextent (Y.C.Linet al., 2014)Fur-27thermore, basedon thehierarchical clustering, the adherent progeny cell lines28showedahigherdegreeofdivergencefromeachothercomparedtosuspension29cells.Thismaybearesultoftheindependenttransformationandisolationofthe30293Tand293Elineagesbythestableintegrationofdifferentviralgenesindif-31ferentlabs.WhereasallthesixHEK293celllinesconstitutivelyexpresstheE1A32andE1Bgenes(Figure1E),which isbeneficial fortheproductionof lenti-and33AAV viruses for gene therapy, the 293T and 293E cell lines also constitutively34expresstheLargeTantigenandEBNA-1,respectively(Figure1E).Interestingly,35a truncated version of the Large T antigenwas also found to be expressed in36293E cells,which has to our knowledge not been reported previously (Figure37S1).TheLargeTantigensequencewaslikelyderivedfromthepRSVneoplasmid38thatwasco-transfectedwithpCMV-EBNA1duringtheisolationofthe293Ec1839clone(Swirskietal.,1992).40
Progeny HEK293 cell lines carry common genomic and transcriptomic41traitsindependentlyevolvedduringcelllinedevelopment42
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

16
Notably, theoverall comparisonof thegenomicand transcriptomicsprofilesof1HEK293cell linessuggests that theparentalHEK293strainhas thehighestdi-2vergence amongst the cell lines and suggests common changes independently3evolvedinallprogenycelllines.Inlinewiththesefindings,commonchangesin4genecopynumbergainorloss(Figure2)andconsensusdifferentialexpression5alterations(Figure3)wereobservedamongstallprogenycelllineswhencom-6paredtoHEK293.Especially,acommonpatternofcopynumbergainandlossfor7progenycelllineswasobservedonchromosome13and18(Figure2).Suchpat-8terns, foundacrossallorseveral lineagesofHEK293isolatedindependentlyby9differentmethods, suggests a selective advantage for altered copy numbers of10theselociinregardtothephenotypesofthecelllines.Onchromosome13,sev-11eral genes associated with the cytoskeleton (BORA, MZT1, PIBF1 and KLHL1)12had a copynumber gain in all progeny cell lines (Table S2). TheDACH1gene13wasalsofoundamongstthesegenes,whichhasnotablybeenshowntohavean14effect on cytoskeleton organization and to induce amore epithelial phenotype15upon overexpression through the upregulation of E-cadherin(F. Zhao et al.,162015).Inaddition,BORA,MZT1andDACH1werefoundamongstthe329genes17commonly differentially expressed between all adherent and suspension cell18lines(TableS2).Indeed,consensusgeneexpressionchangesassociatedwithcy-19toskeleton organization was observed between all progeny HEK293 cell lines20andHEK293intheoverallanalysisoftranscriptionprofiles(Figure3A).Within21the gained region of chromosome 18 common to all progeny cell lines except22293E, thereare severaldesmocollin anddesmogleingenes (DSC1,DSC2,DSC3,23DSG1, DSG2, DSG3 and DSG4) belonging to the cell-cell adhesion gene set24(GO:0098609),whichmayrenderthisregionpronetogenecopynumbervaria-25tion.Noneof these geneswere foundamongst the commonlydifferentially ex-26pressed genes between all progeny cell lines andHEK293 asmaybe expected27since293Ehadalossofcopynumberinthisregion(TableS4).However,several28of these genes (DSC2, DSC3 and DSG2) where found up-regulated in HEK29329suspension cells compared to adherent cells (Table S5), suggesting that they30play a role in the differences between adherent and suspension cell lines. The31observedenrichmentofcelladhesionGO-terms(GO:0007156andGO:0098742)32amongstgeneswithcommonhigh/moderateimpactSNPsuniquetoprogenycell33linescomparedtoHEK293cellsalsosupportedcommongenomicalterationsin34progenycelllinesinvolvedincelladhesion.Combinedwiththeobserveddown-35regulation of the expression of cell adhesion genes in progeny cell lines com-36paredtoHEK293(Figure3A),resultshighlightchangesincelladhesionduring37continuouscultivationandcelllinedevelopmentofHEK293cells.3839Moreover, specific genomic regionsofmore local gainor lossof specific genes40wereobserved,includingalossoffumaratehydratase(FH)genecopies.Theloss41ofFHcopieswaspreviouslyobservedforHEK293byLinandcoworkersandwas42suggestedtoplayanimportantpartinthetransformedphenotypeofthecellline43
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

17
(Y.C. Lin et al., 2014). In linewith this, our results showed that several of the1HEK293progenycelllines(293E,293-FandFreestyle293-F)wasfoundtoonly2maintainhalf thenumberof FHgene copies compared to theoriginalHEK2933(FigureS2),supportinganadvantageouslossoftheFHgeneintheHEK293cell4lineages. Furthermore, a conserved pattern of substantial gain (>1.5 log2-fold5change) of theTLE4 gene and surrounding loci, including the non-codingRNA6LINC01507,was observed for all progeny cells except 293E (Figure S2). This7geneencodesthetransducin-likeenhancerprotein4,whichisatranscriptional8co-repressorofmembersoftheWntsignalingpathway.Down-regulationofthis9gene has been associatedwith the epithelial-to-mesenchymal transition (EMT)10phenotype (Wuetal.,2016), consistentwith theobservation in this study that11theoriginalHEK293strainseemstohaveamoremesenchymalexpressionpro-12filethentheothercelllines(FigureS4).Interestingly,TLE4haspreviouslybeen13reported to have both a tumor suppressor function and to be associatedwith14promoting tumor growth in different studies of different cancers (Shin et al.,152016)(Wanget al., 2016).Moreover, a significant loss of theADAM3Apseudo-16gene(<-1log2-foldchange),whichhaspreviouslybeenassociatedwithdifferent17cancers(Barrowetal.,2011)(Liuetal.,2012),wasobservedinallprogenycell18linesaccept293-HcomparedtoHEK293(FigureS2).ThespecificgainofTLE419and/orlossofADAM3Alociandtheirassociationwithtumordevelopment,sug-20gestimportantfunctionsofthesegenesintheevolutionofHEK293celllines.2122Taken together, the genomic and transcriptomic profiles of HEK293 and its23progeny cell lines suggest commongenomic and gene expression changes that24haveindependentlyevolvedinalltheprogenycelllineagescomparedtotheorig-25inalHEK293,likelyreflectedintraitsprovidingadvantagesduringsinglecelliso-26lationprocedures.Suchtraitscouldbeassociatedwithforinstancefastprolifera-27tionandmaintainingevasionofnormalcellsenescence.Moreover,genomicand28transcriptomictraitsassociatedwithforinstancecelladhesion,cellmotilityand29extracellularmatrix organizationmay verywell be under a selective pressure30during single cell cloning, potentially through altered expression of cell mem-31braneproteins. Indeed, theparentalHEK293cell line isdividing slower and is32more difficult to detach by trypsinization than the other adherent cell lines. It33should however be noted that the assumption that there are independently34evolved common changes inprogeny cell lines compared toHEK293, relies on35thattheHEK293celllineusedasreferenceinthisstudy(ATCC-CRL1573)isac-36tuallytheverysamecelllinethathasbeenusedforthegenerationofeachprog-37enycellline.IfanolderornewerversionofHEK293wasusedforsubcloningof38eitherprogenycellline,thenthedifferencesweseemaybeduetochangesthat39occurredinthisHEK293strainonlyorintheprocessofadaptation.Duetolim-40itedliteratureavailableonprogenycelllinedevelopment,wecannotcompletely41excludethispossibility.However,wehypothesizethatthiscelllineis,ifnotthe42
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

18
same,atleasthighlysimilartotheHEK293strainusedforprogenycelllinede-1velopment.23Key differences between adherent and suspension progeny HEK293 cell4linesincludechangesincelladhesiongeneexpression5In bioproduction processes for pharmaceutical proteins, suspension cell lines6enablelarge-scalecultivationinbioreactors,whichisrequiredinordertomeet7thedemandsformarketeddrugs.However,theadaptationofcellsfromadherent8to suspension growth and the differential cultivation procedures between ad-9herentandsuspensioncellsinducesphenotypicchangestothecelllines.Inorder10todevelopadeeperunderstandingofsuchchanges,weevaluateddifferencesin11geneexpressionlevelsbetweenadherentandsuspensionprogenyHEK293cells.12Consensusdifferentialexpressionresultswerefoundrelatedtoup-regulationof13genesassociatedwithcellcomponentorganizationsuchasmembrane,cytoskel-14eton and cell junction in suspension compared to adherent cells (Figure 5A).15Moreover, significant changeswere associatedwith cell adhesion, cellmotility,16cellmorphogenesisanddifferentiation.Noteworthy,celladhesionwasfoundup-17regulatedinsuspensionprogenycelllinesandwhenlookingintotheexpression18of themost significant differentially expressed genes (adjusted p-value <0.01)19amongst the cell adhesiongene set (Table S6), thereare several cell adhesion20molecules(CAMs)foundbothup-anddown-regulatedinsuspensioncellscom-21pared to adherent. Remarkably, severalmembers of the cadherin superfamily,22including many different protocadherins (PCDH), desmoglein 2 (DSG2) and23desmocollin2(DSC2)were foundsignificantlyup-regulated insuspensioncells24comparedtoadherentcells.Thisfamilyofgenesareinvolvedintheformationof25adherence junctions between cells (Bruner and Derksen, 2018). Interestingly,26amongstthemostsignificantdifferentiallyexpressedgeneswithinthecelladhe-27sion gene set, four protocadherinmembers showed the highest fold-change of28up-regulated genes in suspension cells compared to adherent (Table S6).29Amongst otherCAMs like integrins and Ig-superfamily cell adhesionmolecules30(IgSFCAMs)bothsignificantlyup-anddown-regulatedgeneswereobservedbe-31tweenadherentandsuspensioncelllines.Thehigherexpressionofsuchcellad-32hesion molecules in suspension cell lines compared to adherent progeny33HEK293cellsmaybeexplainedbythelossofculturedishsupporttogrowonin34case of suspension cells. Upon disruption of adhesion interactions with other35cellsandextracellularmatrix,anaturalcellularresponsemaybetoincreaseor36maintain the expression of adhesion molecules in an attempt to restore such37connections.Theabilityofthesuspensioncelllinestoformcellaggregatesdur-38ingsuspensioncultivationandtheeaseofthecellstoattachtoculturedishsur-39facesuponcultivationwithoutshaking,canbespeculatedtosupportthesefind-40ings. Such cell adhesionmolecules found up-regulated in suspension cell lines41may thus be appropriate cell line engineering targets for improvedbioprocess42performanceofsuspensioncelllines.43
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

19
1Metabolicchangesofthecholesterolbiosynthesismayprovidesupportfor2suspensiongrowthinserumfreemedium3Further evaluationof thedifferentially expressedgenesbetweenadherent and4suspension HEK293 progeny cell lines, based on metabolic gene set analysis,5highlightedmetabolicdifferencesbetweenthecelllines.Forinstance,processes6related to thebiosynthesisofaromaticaminoacids (tryptophan,phenylalanine7andtyrosine)weredown-regulated(Figure4B)insuspensioncellscomparedto8adherentcells.Furthermore,othermetabolicpathwayswithdifferencesbetween9suspensionandadherentcelllinesinvolvedcholesterolbiosynthesisandmetab-10olism,omega-3andomega-6fattyacidbiosynthesis, fattyacidbiosynthesis,de-11saturationandelongation,retinolmetabolismandlinoleatemetabolism(Figure124B).Allofthesepathwaysarerelatedtolipidsand/orcholesterolmetabolicpro-13cesses.Thesemetabolicchangesmaybearesultofdifferentgrowthmediacom-14positionsusedforthecultivationofeitheradherentorsuspensioncellsthatmay15imply different concentrations of for instance amino acids, glucose or serum.16Whenreducing thenumberofdifferentiallyexpressedgenes to those thatcon-17sistentlyshoweddifferentialexpressionbetweenadherentandsuspensioncells18inpairwisecomparisonsofallcell lines, thecholesterolandsterolbiosynthesis19andmetabolismpathwaywerefoundtobemostsignificantlydifferentbetween20thecell types(Figure 5B).Assuspensioncell linesarecultivatedunderserum21free conditions, the increased expressionof genes associatedwith for instance22cholesterolinsuspensioncelllinesmaybearesultofalowercholesterolcontent23in the medium. However, since cholesterol is a major component of the cell24membraneandhasan important functionformembranestructureandcellsig-25naling(MaxfieldandTabas,2005)thedifferentialexpressionofgenesassociated26withcholesterolsynthesisandmetabolismmayalsobeofimportanceforthedif-27ferent morphologies between adherent and suspension HEK293 cells. Indeed,28previous studieshave shown that cholesterol plays a critical role in regulating29theformationofcell-to-cellinteractionsinendothelialcells(Corveraetal.,2000)30andthatdepletionofcholesterolreducescelladhesionandincreasesendothelial31cell stiffness (Byfield et al., 2004; Norman et al., 2010). Increased cell surface32stiffnesshasbeenreportedforHEK293cells insuspensioncomparedtoadher-33entstateasaresultofup-regulationandre-organizationoftheactincytoskele-34ton(Haghparastetal.,2015).Thismaypartlybearesultofalteredcholesterol35levelsinthecellmembranesincecholesterolisaregulatoroftheactincytoskele-36tonandcholesteroldepletionhasbeenshowntoinduceactinpolymerization(Qi37etal.,2009).Threeoftheconsistentlyup-regulatedgenes(MSMO1,HMGCS1and38IDI1) in suspensionHEK293 compared to adherent encode enzymes that have39direct roles in the cholesterol biosynthesis pathway (Mazein et al., 2013).Me-40thylsterolmonooxygenase 1 (MSMO1), an endoplasmatic reticulum transmem-41brane protein, and hydroxymethylglutaryl-CoA synthase (HMGCS1), located in42thecytoplasm,arecatalyzingstepsinthecholesterolsynthesispathway,where-43
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

20
asisopentenyl-diphosphatedelta-isomerase(IDI1)catalyzetheisomerizationof1onethecholesterolbuildingblocks,isopentenyldiphosphate.Moreover,twoad-2ditional genes (NPC1L1 and INSIG1) found up-regulated in suspension cells3comparedtoadherent,arealsoassociatedwithcholesterolmetabolismbyvari-4ousprocesses.TheNPC1L1geneencodesatransporterprotein(NPC1-likeintra-5cellularcholesteroltransporter1)requiredforcholesterolabsorptioninthein-6testineandisalsosuggestedtoplayanimportantroleinintracellularcholesterol7traffickingbetweenvesicles(HowlesandHui,2012).Insulin-inducedgene1pro-8tein(INSIG1) isanegativeregulatorofcholesterolsynthesisand important for9cholesterol homeostasis (Dong et al., 2012) andknockout of INSIG1hasprevi-10ouslybeenshowntoresultincholesterolaccumulation(Engelkingetal.,2005).11Asthisgenewasfoundtobeupregulatedinsuspensioncellscomparedtoadher-12entHEK293,thiswouldhavethereverseimpactonthecholesterolbiosynthesis13pathwayinsuspensioncellscomparedtotheabove-mentionedMSMO1,HMGCS114andIDI1genes,leadingtoareductionofthecholesterolcontentofthecell.Nota-15bly, a lower cholesterol biosynthesis in suspension cell lines compared to the16originalHEK293strainwasindeedpredictedusingIPA(SupplementaryFigure177).Itshouldhoweverbenotedthatthispredictiondoesnottakeintoconsidera-18tion theeffectof INSIG1, instead thepredictedreduction incholesterolbiosyn-19thesisinsuspensioncellscomparedtotheHEK293celllineisaresultofdown-20regulationofSC5D(lathosteroloxidase).2122From the transcriptomic data of these different HEK293 cell lines, differences23betweenadherentandsuspensioncellswerefoundtoberelatedtodifferencesin24themetabolismoflipidsingeneralandcholesterolspecifically.Thisemphasizes25apotentiallyimportantroleofcholesterolandlipidbiosynthesisinthetransition26fromadherent to serum free suspension growth ofHEK293 cell types. Froma27bioprocessperspective,differencesinintracellularcholesterolsynthesisandme-28tabolismareofinterestwithregardstothesecretorycapacityofacelllinesince29cholesterol content of a cell can impact its productivity. Previous findings has30shownthatcholesterolisessentialforERtoGolgitransportwithinthesecretory31pathway (Ridsdale et al., 2006) and that secretedproductivity of CHO cells in-32creases upon elevated intracellular cholesterol levels, through silencing of33INSIG1,possiblyduetoincreasingthevolumeoftheGolgicompartment(Lohet34al.,2017).Itwouldthereforebeofinteresttogainfurtherknowledgeaboutthe35cholesterol content and distribution within HEK293 cell lines and potentially36evaluateifthisthispathwaycanbeatargetforenhancedbioproductivitywith-37outhavingadeleteriousimpactonsuspensiongrowthorcellmorphology.3839Transcriptomicandgenomicresults indicatedthatsuspensionadaptation40ofHEK293doesnotfollowtheepithelialtomesenchymaltransition41Moreover,4ofthe38genes(ID1,SMAD7,TXNIPandLOX)thatwereconsistently42differentiallyexpressedbetweenadherentandsuspensionHEK293haveprevi-43
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

21
ously been annotated to play a role in epithelial to mesenchymal transition1(EMT) (Zhaoetal.,2015), theeventwherestationaryepithelial cells lose their2cell-celladhesionandchangeintomotileandinvasivemesenchymalcells(Yang3and Weinberg, 2008). However, when evaluating the expression of common4markersformesenchymalandendothelialphenotypesaswellaspredictingthe5outcomeoftheEMTpathwayusingingenuitypathwayanalysis(IPA),theparen-6talHEK293strainshowedthemostmesenchymal-likephenotypewhereassus-7pension cell lineswerepredicted tohave reduced transition fromepithelial to8mesenchymal phenotype compared HEK293 (Figure S4). These results, along9withabove-mentioneddetailsregardingcopynumbervariationoftheTLE4and10DACH1genes,indicatethatthesuspensionadaptationofHEK293lineagesdoes11notfollowtheEMTpathway.However,suspensioncellswerepredictedtohavea12higher degree of disruption of adherence junctions compared to the parental13HEK293strain(datanotshown),whichmaybeimportantforthesuspensioncell14phenotype.1516Identificationoffivegeneswithpotentialkeyrolesindifferencesbetween17adherentandsuspensionhumancelllines18Altogethernineofthepreviouslyidentified38genes(LOX,ID1,ADAMTS1,ZIC1,19KCNMA1,DHRS3,RARG,COL4A6andARRDC4)wereshowntohavesignificantly20differentexpressionlevelsalsoinanextendedvalidationofthegenesinasetof2163humancelllinesfromtheHPAdatabase(Uhlenetal.,2017)(Figure6).How-22ever,inthecaseoffourofthegenes(ADAMTS1,KCNMA1,COL4A6andARRDC4)23thedirectionofexpressionbetweenadherentandsuspensioncellswasreversed24comparedtothepairwisecomparisonofHEK293celllinesonly(Figure5B).The25remainingfivegenes(LOX,ID1,ZIC1,DHRS3andRARG)showedaconsistentex-26pressionprofilebetweenadherentandsuspensioncellscomparedtotheresults27presentedinFigure5B,suggestingakeyroleofthesegenesinthemorphologies28ofadherentandsuspensioncells.ID1belongstotheTGF-betasignalingpathway29andup-regulationof thisgene,as found inadherentcellscompared tosuspen-30sioncelllines,hasbeenassociatedwiththemesenchymal-to-epithelialtransition31(ID1) (Stankic et al., 2013). Consistently, ID1 silencinghas alsobeen shown to32significantly reduce adhesion of neural stem cells (Tan et al., 2012) and con-33versely,increasedID1expressioninepithelialcellshasbeenrelatedtoincreased34adhesion(Qiuetal.,2011).Inaddition,lysyloxidase(LOX),anenzymeresponsi-35bleforthecovalentcross-linkingbetweenelastinandcollagenintheextracellu-36larmatrix,hasbeenshowntobe important forcell-matrixadhesion formation,37supportingtheadherentphenotypeofadherentcellsbutisalsoassociatedwith38cellinvasionandinductionofEMT(Payneetal.,2005)(Schietkeetal.,2010).Be-39sides the EMT-related genes, the additional three genes (RARG, DHRS3 and40ZIC1), consistently up-regulated in adherent cells compared to suspension cell41lines, have previously been associatedwith cell adhesion through the retinoic42acid signaling pathway. Indeed, retinol metabolism was found to be down-43
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

22
regulatedinsuspensioncellsalso inthemetabolicgenesetanalysis.TheRARG1gene,which isupregulated in adherent cells compared to suspension cells, en-2codestheretinoicacidreceptorgammathatcontrolstheexpressionofgenesin-3volvedincelladhesionandplaysanimportantroleinpromotingcelladhesionin4mouseembryonicfibroblasts(AlTanouryetal.,2013)(Kelleyetal.,2017).Fur-5thermore, the transcription factorZIC1hasbeenassociatedwith theupregula-6tionofgenesassociatedwithretinoicacidsignalingandadhesion(Cornishetal.,72009;Ganetal.,2011)whereasretinoicacid-inducibledehydrogenasereductase83(DHRS3)isanenzymeimportantforretinoicacidhomeostasisandactsanega-9tive regulatorof retinonicacid synthesis.Thisenzymehasaweakcatalytic ca-10pacitytoconvertretinaldehydetoretinol,whichresultsinreducedretinoicacid11levels, unless co-expressedwithRDH10,which induces the full capacity of the12DHRS3 enzyme, resulting in decreased retinoic acid synthesis (Adams et al.,132014).Furthermore,signalingofretinonicacidreceptorsonlyintheabsenceret-14inoicacidisassociatedwithincreasedadhesioninmouseembryonicfibroblasts15(AlTanouryetal.,2013).ThelevelofRDH10,alongwithDHRS3,wassignificant-16lylowerintheHEK293suspensioncellscomparedtoadherent(TableS5),sug-17gestingmoreefficientcatalyticactivityoftheDHRS3enzymeintheadherentcell18lines,whichmay lead to retinoic acid signaling in the absenceof retinoic acid,19resultinginamoreadherentphenotypeinthesecelllines.2021Conclusions22Ourstudyhasoutlinedthegenomic,transcriptomicandmetabolomicvariations23betweensixindustriallyrelevantHEK293celllines,inanattempttoimprovethe24understandingoftheirrespectivedifferencesinphenotype.Wereportaselective25pressuretodevelopcertainexpressionprofilesduringtheevolutionandcontin-26uouscultivation,evidencedbythenumerousgenesandpathwaysdetailedhere.27ThekeycommonchangesbetweenHEK293anditsprogenycelllinesinvolvein28particularcellmembraneproteinsandprocessesrelatedtocelladhesion,motili-29tyandtheorganizationofvariouscellularcomponentssuchasthecytoskeleton30and extracellular matrix. In addition, changes associated with differences be-31tween adherent and suspension cell growth in particularly involve changes in32cell adhesion protein expression, cholesterol metabolism and a set of six key33genes(RARG,ID1,ZIC1,LOXandDHRS3)withpotentiallykeyrolesinthediffer-34entiationbetween the two groups. These results couldbe of importancewhen35pursuing further cell line engineering or bioprocess optimization of these and36otherhumancelllines.37
Acknowledgements38
ThisworkwassupportedbytheKnutandAliceWallenbergFoundation,Astra-39Zeneca, SSF, Vinnova and the Novo Nordisk Foundation (grant no.40NNF10CC1016517).41
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

23
AuthorContributions1
Conceptualization, R.F., P.V., M.U., J.N., and J.R; Methodology, M.M., R.S. and J.R. 2Formal analysis, M.M., R.S., and M.L.; Investigation, M.M., R.S., and M.L., Writing 3– Original Draft, M.M, R.S., M.L., and J.R.; Writing – Review & Editing, M.M., 4M.L., D.H., J.N., and J.R.; Visualization, M.M., R.S., and M.L., Supervision, R.F., 5P.V., D.H., T.S., M.U., J.N., and J.R., Funding Acquisition, R.F., P.V., M.U. J.N, and 6J.R.7
Declarationofinterests8
The authors declare no competing interests910
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

24
Figures1
2Figure1. Comparisonsof genomicand transcriptomicprofilesofHEK2933cellsshowedtaxonomicdivergencebetweenparentalHEK293andprogeny4celllines.5(A)AschematicoverviewofthelineagerelationshipofthesixHEK293celllines6usedinthisstudy.Bluedotsrepresentadherentcellswhereasgreydotsrepre-7sent suspensioncell lines. (B)Genomic comparisonbetweenHEK293cell lines8basedonSpearmancorrelationcoefficientsofreadcounts.Darkerbluecolorin-9dicateshighercorrelation.(C)Sample-to-samplecomparisonbetweentranscrip-10tomesillustratedbyaheatmapandhierarchicalclusteringoftaxonomicaldiver-11gencebetweensamples.Darkerbluecolor indicatesshorterEuclideandistance12between samples andmore similarity. (D) PCAplot showing the separation in13
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

25
expression pattern between samples. (E) RNA expression levels of stably inte-1gratedviralgenes(EBNA-1,LargeT,E1AandE1B)invariousHEK293cellline-2agesdeterminedbyRNAsequencing.3
4
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

26
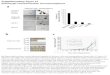
1Figure2.CopynumbervariationanalysisofHEK293progenycellscom-2paredtotheparentalHEK293revealedconservedpatternsofcopynumber3gainandloss.A)Copynumbergain(red)orloss(blue)overallchromosomesof4progenyHEK293celllines.Thecolorintensivenesscorrespondtolog2fold-5changecomparedtoHEK293cells.Theasterixsymbol(*)marksregionsof6commoncopynumbergain(>1log2foldchange)commonbetweenseveral7progenyHEK293celllinesonchromsome13and18.Identifiedgeneswithlocal8
1
293E
293T
293-H
293-F Free
style
293-F
2
3
4
5
293E
293T
293-H
293-F Free
style
293-F
7
8
9
10
11
6
12
14
15
13
293E
293T
293-H
293-F Free
style
293-F
16
17
18
19
20
21
22
X
Gain Loss
FH, KMOADAM3A
TLE4, LINC01507
*
*
A)
B)
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

27
copynumbergainorlossofthefullgenesequencecommonbetweenseveral1HEK293progenycelllinesaremarkedwiththegenename.B)Copynumbergain2orloss(log2foldchangecomparedtoHEK293foreachcelllineoftheFH,3KMO,TLE4andADAM3Agenes.4 5
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

28
1Figure3.Differentialexpressionanalysisemphasizedprocessesandgenes2with common changes in all progeny cell lines compared to the parental3HEK2934(A) Consensus heatmap of GO biological processeswith a different expression5patternbetweenprogenycell linescomparedtotheparentalHEK293(B)Com-6mondifferentiallyexpressed(DE)genesinpairwisecomparisonsofallHEK2937cells.Bluebars shownumberofDEgenes in eachpairwise comparison.Green8bar shows 329 common DE genes in pairwise comparisons of progeny with9HEK293parentalcells.Redbarshowscommon38DEgenesinthecomparisonof10suspensioncellsagainstadherentcells.(C)ToptensignificantGOcellularcom-11ponentsofthe329commonDEgenesinpairwisecomparisonsbetweenprogeny12cellsandHEK293.13 14
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

29
1Figure4.Differentialexpressionanalysisbetweensuspensionandadher-2ent HEK293 progeny cell lineages identified significant up-regulation of3gene sets associated with cell adhesion, motility, differentiation, cellular4compartmentorganizationandcholesterolmetabolisminsuspensioncells.5(A) Consensus heatmap of GO biological processeswith a different expression6pattern between suspension cells (293-H and 293-F) against adherent cells7(293Eand293T).(B)Metabolicgenessetanalysisforcomparingmetabolicdif-8ferencesbetweensuspensionandadherentHEK293progenycelllines.Thesize9of each node corresponds to the number of genes in each of these pathways,10thickness of connections between nodes corresponds to the number shared11genesbetweenpathwaysandthecolorsofthenodesshowsthep-valueforthe12givenmetabolicprocess.13 14
A
B
P-value up
P-value down
Max node size:256 genes
Min node size:6 genes
Max edge with:23 genes
Min edge width:1 gene
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

30
1Figure5.Pairwisecomparisonoftranscriptiondataidentifiedcholesterol2biosynthesisasthemainenrichedpathwaybetweenadherentandsuspen-3sionHEK293cells.4(A)HeatmapwithTPMvaluesforeachDEgenecommontoalladherenttosus-5pensioncomparisons.(B)ThetoptenmostenrichedbiologicalGOtermsofthe638commonDEgenesbetweenadherentandsuspensioncellsbasedongeneen-7richmentanalysis.Lengthandcolorofbarsbothshowsignificanceofadjustedp-8value for the hypergeometric test. Also, genes mentioned in each bar are the9genesthatbelongtoenrichedGOtermandpresentinthelistof38DEgenes.10 11
Suspension Cells
AdherentCells
A)
cholesterol biosynthetic process (GO:0006695) - IDI1;HMGCS1;NPC1L1;MSMO1;INSIG1
secondary alcohol biosynthetic process (GO:1902653) - IDI1;HMGCS1;NPC1L1;MSMO1;INSIG1
sterol biosynthetic process (GO:0016126) - IDI1;HMGCS1;NPC1L1;MSMO1; INSIG1
cholesterol metabolic process (GO:0008203) - IDI1;HMGCS1;NPC1L1;MSMO1;INSIG1
regulation of ubiquitin-protein transferase activity (GO:0051438) - ARRDC4;ARRDC3;SMAD7
cellular response to sterol depletion (GO:0071501) - NPC1L1;INSIG1
generation of neurons (GO:0048699) - TUBB2B;ID1;ID3;MAPT 0 2 4
-log10(Padj)
positive regulation of protein ubiquitination (GO:0031398) - ARRDC4;ARRDC3;SMAD7
central nervous system neuron development (GO:0021954) - NPY;MAPT
negative regulation of transferase activity (GO:0051348) - MAPT;SMAD7
B)
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

31
1
2Figure6. Gene expression validation of 38previously identifieddifferen-3tiallyexpressedgenes in63humancell lines, identifiedninesignificantly4differentiallyexpressedgenesbetweensuspensionandadherentcelllines.5(A)PCAtranscriptomicdataof63humancell-linesfromtheHumanProteinAt-6lasshowsaclearseparationofsuspensionandadherentcelllinesfromdifferent7tissues.(B)RangeofnormalizedcountsinHPAcelllinesforeachoftheprevious-8lyidentified38genes,differentiallyexpressedbetweenalladherentandsuspen-9sionHEK293cell lines.Theblack line ineachboxshowsmedianofnormalized10countsforthegene.(C)Genesthataredifferentiallyexpressedbetweenadherent11andsuspensioncellsusingaMann-WhitneyU-test,withp-values<0.01,arehigh-12lightedinpurple,wherelengthofbarsshowslogarithmicfoldchangeofmedian13between twogroups and the colorof barsdenotesdegreeof significanceofp-14value.Non-significantgeneshavegraybars.15 16
Nor
mal
ized
cou
nts
0"12
34"&
*+5%
-1)"
43$)
*'%-
Morphology
B
A
-Log10 (p value)
C
Log2
fold
cha
nge
of m
edia
n
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

32
TableS1.Transcriptomicdata(TPMvalues)ofallgenesinsixHEK293celllines12TableS2.Geneswithfullorpartialcopynumbergainorloss(>1/<-1log2fold-3change)inHEK293progenycelllinescomparedtotheparentalHEK293.Related4toFigure2.56TableS3.HighandmoderateimpactSNPscommonanduniquetoallprogenycell7linescomparedtotheparentalHEK293.89TableS4.Resultsofdifferential expressionanalysisbetweenprogeny cell lines10compared to HEK293 and intersects between results. Related to Figure 3 and11Figure5.1213TableS5.Resultsofdifferentialexpressionanalysisbetweensuspensionandad-14herentprogenyHEK293celllines.RelatedtoFigure4.1516TableS6.Listofdifferentiallyexpressedgenes(padj<0.01)insuspensioncom-17paredtoadherentHEK293cellline.RelatedtoFigure4.1819TableS7.Fullresultsofmetabolicgenesetanalysis.RelatedtoFigure4.2021TableS8.Normalized countsof38differentially expressedgenesbetween sus-22pensionandadherentHEK293celllinesinHPAcelllines.RelatedtoFigure623 24
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

33
References1Adams,M.K.,Belyaeva,O.V,Wu,L.,Kedishvili,N.Y.,2014.Theretinaldehyde2
reductaseactivityofdhrs3isreciprocallyactivatedbyretinol3dehydrogenase10tocontrolretinoidhomeostasis.J.Biol.Chem.289,414868–14880.doi:10.1074/jbc.M114.5522575
AlTanoury,Z.,Piskunov,A.,Andriamoratsiresy,D.,Gaouar,S.,Lutzing,R.,Ye,T.,6Jost,B.,Keime,C.,Rochette-Egly,C.,2013.Genesinvolvedincelladhesion7andsignaling:anewrepertoireofretinoicacidreceptortargetgenesin8mouseembryonicfibroblasts.J.CellSci.127,521–533.9doi:10.1242/jcs.13194610
AlexPhilippidis,2017.TheTop15Best-SellingDrugsof2016|TheLists|GEN11[WWWDocument].12
Bandaranayake,A.D.,Almo,S.C.,2014.Recentadvancesinmammalianprotein13production.FEBSLett.588,253–60.doi:10.1016/j.febslet.2013.11.03514
Barrow,J.,Adamowicz-Brice,M.,Cartmill,M.,MacArthur,D.,Lowe,J.,Robson,K.,15Brundler,M.A.,Walker,D.A.,Coyle,B.,Grundy,R.,2011.Homozygouslossof16ADAM3Arevealedbygenome-wideanalysisofpediatrichigh-gradeglioma17anddiffuseintrinsicpontinegliomas.Neuro.Oncol.13,212–222.18doi:10.1093/neuonc/noq15819
Becker,J.,Timmermann,C.,Jakobi,T.,Rupp,O.,Szczepanowski,R.,Hackl,M.,20Goesmann,A.,Tauch,A.,Borth,N.,Grillari,J.,Pühler,A.,Noll,T.,Brinkrolf,K.,212011.Next-generationsequencingoftheCHOcelltranscriptome.BMCProc.225Suppl8,P6.doi:10.1186/1753-6561-5-S8-P623
Berk,A.J.,2005.Recentlessonsingeneexpression,cellcyclecontrolandcell24biologyfromadenovirus.Oncogene24,7673–7685.25doi:10.1038/sj.onc.120904026
Birzele,F.,Schaub,J.,Rust,W.,Clemens,C.,Baum,P.,Kaufmann,H.,Weith,A.,27Schulz,T.W.,Hildebrandt,T.,2010.Intotheunknown:expressionprofiling28withoutgenomesequenceinformationinCHObynextgeneration29sequencing.NucleicAcidsRes.38,3999–4010.doi:10.1093/nar/gkq11630
Bray,N.L.,Pimentel,H.,Melsted,P.,Pachter,L.,2016.Near-optimalprobabilistic31RNA-seqquantification.Nat.Biotechnol.34,525–527.doi:10.1038/nbt.351932
Brinkrolf,K.,Rupp,O.,Laux,H.,Kollin,F.,Ernst,W.,Linke,B.,Kofler,R.,Romand,33S.,Hesse,F.,Budach,W.E.,Galosy,S.,Müller,D.,Noll,T.,Wienberg,J.,Jostock,34T.,Leonard,M.,Grillari,J.,Tauch,A.,Goesmann,A.,Helk,B.,Mott,J.E.,Pühler,35A.,Borth,N.,2013.Chinesehamstergenomesequencedfromsorted36chromosomes.Nat.Biotechnol.31,694–695.doi:10.1038/nbt.264537
Bruner,H.C.,Derksen,P.W.B.,2018.LossofE-cadherin-dependentcell–cell38adhesionandthedevelopmentandprogressionofcancer.ColdSpringHarb.39Perspect.Biol.doi:10.1101/cshperspect.a02933040
Byfield,F.J.,Aranda-Espinoza,H.,Romanenko,V.G.,Rothblat,G.H.,Levitan,I.,412004.Cholesteroldepletionincreasesmembranestiffnessofaortic42endothelialcells.Biophys.J.87,3336–3343.43doi:10.1529/biophysj.104.04063444
Bylund,L.,Kytölä,S.,Lui,W.-O.,Larsson,C.,Weber,G.,2004.Analysisofthe45cytogeneticstabilityofthehumanembryonalkidneycellline293by46cytogeneticandSTRprofilingapproaches.Cytogenet.GenomeRes.106,28–4732.doi:10.1159/00007855648
Chen,E.Y.,Tan,C.M.,Kou,Y.,Duan,Q.,Wang,Z.,Meirelles,G.,Clark,N.R.,Ma’ayan,49
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

34
A.,2013.Enrichr:interactiveandcollaborativeHTML5genelistenrichment1analysistool.BMCBioinformatics14,128.doi:10.1186/1471-2105-14-1282
Clément,N.,Grieger,J.C.,2016.Manufacturingofrecombinantadeno-associated3viralvectorsforclinicaltrials.Mol.Ther.MethodsClin.Dev.3,16002.4doi:10.1038/mtm.2016.25
Cornish,E.J.,Hassan,S.M.,Martin,J.D.,Li,S.,Merzdorf,C.S.,2009.Amicroarray6screenfordirecttargetsofZIC1identifiesanaquaporingene,aqp-3b,7expressedintheneuralfolds.Dev.Dyn.238,1179–1194.8doi:10.1002/dvdy.219539
Corvera,S.,DiBonaventura,C.,Shpetner,H.S.,2000.Cellconfluence-dependent10remodelingofendothelialmembranesmediatedbycholesterol.J.Biol.11Chem.275,31414–31421.doi:10.1074/jbc.M00170820012
Côté,J.,Garnier,A.,Massie,B.,Kamen,A.,1998.Serum-freeproductionof13recombinantproteinsandadenoviralvectorsby293SF-3F6cells.14Biotechnol.Bioeng.59,567–575.doi:10.1002/(SICI)1097-150290(19980905)59:5<567::AID-BIT6>3.0.CO;2-816
Dietmair,S.,Hodson,M.P.,Quek,L.E.,Timmins,N.E.,Chrysanthopoulos,P.,Jacob,17S.S.,Gray,P.,Nielsen,L.K.,2012.MetaboliteprofilingofCHOcellswith18differentgrowthcharacteristics.Biotechnol.Bioeng.109,1404–1414.19doi:10.1002/bit.2449620
Dong,X.Y.,Tang,S.Q.,Chen,J.D.,2012.DualfunctionsofInsigproteinsin21cholesterolhomeostasis.LipidsHealthDis.doi:10.1186/1476-511X-11-17322
DuBridge,R.B.,Tang,P.,Hsia,H.C.,Leong,P.M.,Miller,J.H.,Calos,M.P.,1987.23AnalysisofmutationinhumancellsbyusinganEpstein-Barrvirusshuttle24system.Mol.Cell.Biol.7,379–87.25
Dumont,J.,Euwart,D.,Mei,B.,Estes,S.,Kshirsagar,R.,2016.Humancelllinesfor26biopharmaceuticalmanufacturing:history,status,andfutureperspectives,27CriticalReviewsinBiotechnology.Taylor&Francis.28doi:10.3109/07388551.2015.108426629
Durocher,Y.,Perret,S.,Kamen,A.,2002.High-levelandhigh-throughput30recombinantproteinproductionbytransienttransfectionofsuspension-31growinghuman293-EBNA1cells.NucleicAcidsRes.30,E9.32
Engelking,L.J.,Liang,G.,Hammer,R.E.,Takaishi,K.,Kuriyama,H.,Evers,B.M.,Li,33W.P.,Horton,J.D.,Goldstein,J.L.,Brown,M.S.,2005.Schoenheimereffect34explained-Feedbackregulationofcholesterolsynthesisinmicemediated35byInsigproteins.J.Clin.Invest.115,2489–2498.doi:10.1172/JCI2561436
Foxman,B.,2011.OmicsAnalysesinMolecularEpidemiologicStudies,in:37MolecularToolsandInfectiousDiseaseEpidemiology.AcademicPress,pp.3899–116.doi:10.1016/b978-0-12-374133-2.00007-139
Gan,L.,Chen,S.,Zhong,J.,Wang,X.,Lam,E.K.Y.,2011.ZIC1IsDownregulated40throughPromoterHypermethylation,andFunctionsasaTumorSuppressor41GeneinColorectalCancer.PLoSOne6,16916.42doi:10.1371/journal.pone.001691643
Garnier,A.,Côté,J.,Nadeau,I.,Kamen,A.,Massie,B.,1994.Scale-upofthe44adenovirusexpressionsystemfortheproductionofrecombinantproteinin45human293Scells.Cytotechnology15,145–55.46
Graham,F.L.,1987.Growthof293CellsinSuspensionCulture.J.Gen.Virol.68,47937–940.doi:10.1099/0022-1317-68-3-93748
Graham,F.L.,Smileyt,J.,Russell,W.C.,Nairn,R.,1977.CharacteristicsofaHuman49
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

35
CellLineTransformedbyDNAfromHumanAdenovirusType5.J.gen.Virol136,59–7.doi:10.1099/0022-1317-36-1-592
Haghparast,S.M.A.,Kihara,T.,Miyake,J.,2015.Distinctmechanicalbehaviorof3HEK293cellsinadherentandsuspendedstates.PeerJ3,e1131.4doi:10.7717/peerj.11315
Hammond,S.,Swanberg,J.C.,Kaplarevic,M.,Lee,K.H.,2011.Genomicsequencing6andanalysisofaChinesehamsterovarycelllineusingIlluminasequencing7technology.BMCGenomics12,67.doi:10.1186/1471-2164-12-678
Howles,P.N.,Hui,D.Y.,2012.PhysiologicalroleofhepaticNPC1L1inhuman9cholesterolandlipoproteinmetabolism:Newperspectivesandopen10questions.J.LipidRes.53,2253–2255.doi:10.1194/jlr.e03182311
Kelley,M.D.,Phomakay,R.,Lee,M.,Niedzwiedz,V.,Mayo,R.,2017.Retinoicacid12receptorgammaimpactscellularadhesion,Alpha5Beta1integrinexpression13andproliferationinK562cells.PLoSOne12,e0178116.14doi:10.1371/journal.pone.017811615
Kildegaard,H.F.,Baycin-Hizal,D.,Lewis,N.E.,Betenbaugh,M.J.,2013.The16emergingCHOsystemsbiologyera:harnessingthe‘omicsrevolutionfor17biotechnology.Curr.Opin.Biotechnol.24,1102–1107.18doi:10.1016/J.COPBIO.2013.02.00719
Kuleshov,M.V.,Jones,M.R.,Rouillard,A.D.,Fernandez,N.F.,Duan,Q.,Wang,Z.,20Koplev,S.,Jenkins,S.L.,Jagodnik,K.M.,Lachmann,A.,McDermott,M.G.,21Monteiro,C.D.,Gundersen,G.W.,Ma’ayan,A.,2016.Enrichr:a22comprehensivegenesetenrichmentanalysiswebserver2016update.23NucleicAcidsRes.44,W90–W97.doi:10.1093/nar/gkw37724
Lalonde,M.-E.E.,Durocher,Y.,2017.Therapeuticglycoproteinproductionin25mammaliancells.J.Biotechnol.251,128–140.26doi:10.1016/j.jbiotec.2017.04.02827
Leader,B.,Baca,Q.J.,Golan,D.E.,2008.Proteintherapeutics:asummaryand28pharmacologicalclassification.Nat.Rev.DrugDiscov.7,21–39.29doi:10.1038/nrd239930
Lee,J.S.,Kallehauge,T.B.,Pedersen,L.E.,Kildegaard,H.F.,Joung,J.K.,2015.Site-31specificintegrationinCHOcellsmediatedbyCRISPR/Cas9andhomology-32directedDNArepairpathway.Sci.Rep.5,8572.doi:10.1038/srep0857233
Lin,Y.-C.,Boone,M.,Meuris,L.,Lemmens,I.,Roy,N.Van,Soete,A.,2014.Genome34dynamicsofthehumanembryonickidney293lineageinresponsetocell35biologymanipulations.JokeReumers9101013101.36doi:10.1038/ncomms576737
Lin,Y.C.,Boone,M.,Meuris,L.,Lemmens,I.,VanRoy,N.,Soete,A.,Reumers,J.,38Moisse,M.,Plaisance,S.,Drmanac,R.,Chen,J.,Speleman,F.,Lambrechts,D.,39VanDePeer,Y.,Tavernier,J.,Callewaert,N.,2014.Genomedynamicsofthe40humanembryonickidney293lineageinresponsetocellbiology41manipulations.Nat.Commun.5,4767.doi:10.1038/ncomms576742
Liu,J.,Lee,W.,Jiang,Z.,Chen,Z.,Jhunjhunwala,S.,Haverty,P.M.,Gnad,F.,Guan,43Y.,Gilbert,H.N.,Stinson,J.,Klijn,C.,Guillory,J.,Bhatt,D.,Vartanian,S.,44Walter,K.,Chan,J.,Holcomb,T.,Dijkgraaf,P.,Johnson,S.,Koeman,J.,Minna,45J.D.,Gazdar,A.F.,Stern,H.M.,Hoeflich,K.P.,Wu,T.D.,Settleman,J.,De46Sauvage,F.J.,Gentleman,R.C.,Neve,R.M.,Stokoe,D.,Modrusan,Z.,Seshagiri,47S.,Shames,D.S.,Zhang,Z.,2012.Genomeandtranscriptomesequencingof48lungcancersrevealdiversemutationalandsplicingevents.GenomeRes.22,49
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

36
2315–2327.doi:10.1101/gr.140988.1121Loh,W.P.,Yang,Y.,Lam,K.P.,2017.miR-92aenhancesrecombinantprotein2
productivityinCHOcellsbyincreasingintracellularcholesterollevels.3Biotechnol.J.12,1600488.doi:10.1002/biot.2016004884
Louis,N.,Evelegh,C.,Graham,F.L.,1997.CloningandSequencingoftheCellular–5ViralJunctionsfromtheHumanAdenovirusType5Transformed293Cell6Line.Virology233,423–429.doi:10.1006/viro.1997.85977
Mardinoglu,A.,Agren,R.,Kampf,C.,Asplund,A.,Uhlen,M.,Nielsen,J.,2014.8Genome-scalemetabolicmodellingofhepatocytesrevealsserinedeficiency9inpatientswithnon-alcoholicfattyliverdisease.Nat.Commun.5,3083.10doi:10.1038/ncomms408311
Maxfield,F.R.,Tabas,I.,2005.Roleofcholesterolandlipidorganizationin12disease.Nature.doi:10.1038/nature0439913
Mazein,A.,Watterson,S.,Hsieh,W.Y.,Griffiths,W.J.,Ghazal,P.,2013.A14comprehensivemachine-readableviewofthemammaliancholesterol15biosynthesispathway.Biochem.Pharmacol.86,56–66.16doi:10.1016/j.bcp.2013.03.02117
Mootha,V.K.,Lindgren,C.M.,Eriksson,K.-F.,Subramanian,A.,Sihag,S.,Lehar,J.,18Puigserver,P.,Carlsson,E.,Ridderstråle,M.,Laurila,E.,Houstis,N.,Daly,M.J.,19Patterson,N.,Mesirov,J.P.,Golub,T.R.,Tamayo,P.,Spiegelman,B.,Lander,20E.S.,Hirschhorn,J.N.,Altshuler,D.,Groop,L.C.,2003.PGC-1α-responsive21genesinvolvedinoxidativephosphorylationarecoordinately22downregulatedinhumandiabetes.Nat.Genet.34,267–273.23doi:10.1038/ng118024
Murphy,A.J.M.,Kung,A.L.,Swirski,R.A.,Schimke,R.T.,1992.cDNAexpression25cloninginhumancellsusingthepλDR2episomalvectorsystem.Methods4,26111–131.doi:10.1016/1046-2023(92)90044-927
Norman,L.L.,Oetama,R.J.,Dembo,M.,Byfield,F.,Hammer,D.A.,Levitan,I.,28Aranda-Espinoza,H.,2010.Modificationofcellularcholesterolcontent29affectstractionforce,adhesionandcellspreading.Cell.Mol.Bioeng.3,151–30162.doi:10.1007/s12195-010-0119-x31
Payne,S.L.,Fogelgren,B.,Hess,A.R.,Seftor,E.A.,Wiley,E.L.,Fong,S.F.T.,Csiszar,32K.,Hendrix,M.J.C.,Kirschmann,D.A.,2005.Lysyloxidaseregulatesbreast33cancercellmigrationandadhesionthroughahydrogenperoxide-mediated34mechanism.CancerRes.65,11429–11436.doi:10.1158/0008-5472.CAN-3505-127436
Priola,J.J.,Calzadilla,N.,Baumann,M.,Borth,N.,Tate,C.G.,Betenbaugh,M.J.,372016.High-throughputscreeningandselectionofmammaliancellsfor38enhancedproteinproduction,BiotechnologyJournal.WILEY‐VCHVerlag.39doi:10.1002/biot.20150057940
Qi,M.,Liu,Y.,Freeman,M.R.,Solomon,K.R.,2009.Cholesterol-regulatedstress41fiberformation.J.Cell.Biochem.106,1031–1040.doi:10.1002/jcb.2208142
Qiu,J.,Wang,G.,Peng,Q.,Hu,J.,Luo,X.,Zheng,Y.,Teng,Y.,Tang,C.,2011.Id143inducestubulogenesisbyregulatingendothelialcelladhesionand44cytoskeletalorganizationthroughβ1-integrinandRho-kinasesignalling.Int.45J.Mol.Med.28,543–548.doi:10.3892/ijmm.2011.74146
Ridsdale,A.,Denis,M.,Gougeon,P.-Y.,Ngsee,J.K.,Presley,J.F.,Zha,X.,2006.47Cholesterolisrequiredforefficientendoplasmicreticulum-to-Golgi48transportofsecretorymembraneproteins.Mol.Biol.Cell17,1593–605.49
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

37
doi:10.1091/mbc.e05-02-01001Sanchez-Garcia,L.,Martín,L.,Mangues,R.,Ferrer-Miralles,N.,Vázquez,E.,2
Villaverde,A.,2016.Recombinantpharmaceuticalsfrommicrobialcells:a32015update.Microb.CellFact.15,33.doi:10.1186/s12934-016-0437-34
Schietke,R.,Warnecke,C.,Wacker,I.,Schödel,J.,Mole,D.R.,Campean,V.,Amann,5K.,Goppelt-Struebe,M.,Behrens,J.,Eckardt,K.U.,Wiesener,M.S.,2010.The6lysyloxidasesLOXandLOXL2arenecessaryandsufficienttorepressE-7cadherininHypoxia:Insightsintocellulartransformationprocesses8mediatedbyHIF-1.J.Biol.Chem.285,6658–6669.9doi:10.1074/jbc.M109.04242410
Sellick,C.A.,Croxford,A.S.,Maqsood,A.R.,Stephens,G.,Westerhoff,H.V.,11Goodacre,R.,Dickson,A.J.,2011.MetaboliteprofilingofrecombinantCHO12cells:Designingtailoredfeedingregimesthatenhancerecombinant13antibodyproduction.Biotechnol.Bioeng.108,3025–3031.14doi:10.1002/bit.2326915
Shaw,G.,Morse,S.,Ararat,M.,Graham,F.L.,2002.Preferentialtransformationof16humanneuronalcellsbyhumanadenovirusesandtheoriginofHEK29317cells.FASEBJ.16,869–871.doi:10.1096/fj.01-0995fje18
Shin,T.H.,Brynczka,C.,Dayyani,F.,Rivera,M.N.,Sweetser,D.A.,2016.TLE419regulationofwnt-mediatedinflammationunderliesitsroleasatumor20suppressorinmyeloidleukemia.Leuk.Res.48,46–56.21doi:10.1016/j.leukres.2016.07.00222
Soneson,C.,Love,M.I.,Robinson,M.D.,2015.DifferentialanalysesforRNA-seq:23transcript-levelestimatesimprovegene-levelinferences.F1000Research4,241521.doi:10.12688/f1000research.7563.125
Stankic,M.,Pavlovic,S.,Chin,Y.,Brogi,E.,Padua,D.,Norton,L.,Massagué,J.,26Benezra,R.,2013.TGF-β-Id1signalingopposestwist1andpromotes27metastaticcolonizationviaamesenchymal-to-epithelialtransition.CellRep.285,1228–1242.doi:10.1016/j.celrep.2013.11.01429
Stepanenko,A.A.,Dmitrenko,V.V.,2015.HEK293incellbiologyandcancer30research:Phenotype,karyotype,tumorigenicity,andstress-induced31genome-phenotypeevolution.Gene.doi:10.1016/j.gene.2015.05.06532
Stepanenko,A.A.,Dmitrenko,V.V.,2015.HEK293incellbiologyandcancer33research:phenotype,karyotype,tumorigenicity,andstress-induced34genome-phenotypeevolution.Gene569,182–190.35doi:10.1016/j.gene.2015.05.06536
Swirski,R.A.,VanDenBerg,D.,Murphy,A.J.M.,Lambert,C.M.,Friedberg,E.C.,37Schimke,R.T.,1992.ImprovementsintheEpstein-Barr-basedshuttlevector38systemfordirectcloninginhumantissueculturecells.Methods4,133–142.39doi:10.1016/1046-2023(92)90045-A40
Tan,R.,Lee,Y.J.,Chen,X.,2012.Id-1playsakeyroleincelladhesioninneural41stemcellsthroughthepreservationofRAP1signaling.CellAdhes.Migr.42doi:10.4161/cam.1980943
Tang,Y.,Liu,Z.,Zhao,L.,Clemens,T.L.,Cao,X.,2008.Smad7stabilizesβ-catenin44bindingtoE-cadherincomplexandpromotescell-celladhesion.J.Biol.45Chem.283,23956–23963.doi:10.1074/jbc.M80035120046
Uhlen,M.,Zhang,C.,Lee,S.,Sjöstedt,E.,Fagerberg,L.,Bidkhori,G.,Benfeitas,R.,47Arif,M.,Liu,Z.,Edfors,F.,Sanli,K.,vonFeilitzen,K.,Oksvold,P.,Lundberg,E.,48Hober,S.,Nilsson,P.,Mattsson,J.,Schwenk,J.M.,Brunnström,H.,Glimelius,49
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

38
B.,Sjöblom,T.,Edqvist,P.-H.,Djureinovic,D.,Micke,P.,Lindskog,C.,1Mardinoglu,A.,Ponten,F.,2017.Apathologyatlasofthehumancancer2transcriptome.Science357,eaan2507.doi:10.1126/science.aan25073
Väremo,L.,Gatto,F.,Nielsen,J.,2014.Kiwi:Atoolforintegrationand4visualizationofnetworktopologyandgene-setanalysis.BMCBioinformatics515.doi:10.1186/s12859-014-0408-96
Väremo,L.,Nielsen,J.,Nookaew,I.,2013.Enrichingthegenesetanalysisof7genome-widedatabyincorporatingdirectionalityofgeneexpressionand8combiningstatisticalhypothesesandmethods.NucleicAcidsRes.41,4378–94391.doi:10.1093/nar/gkt11110
Vcelar,S.,Jadhav,V.,Melcher,M.,Auer,N.,Hrdina,A.,Sagmeister,R.,Heffner,K.,11Puklowski,A.,Betenbaugh,M.,Wenger,T.,Leisch,F.,Baumann,M.,Borth,N.,122018.KaryotypevariationofCHOhostcelllinesovertimeinculture13characterizedbychromosomecountingandchromosomepainting.14Biotechnol.Bioeng.115,165–173.doi:10.1002/bit.2645315
Walsh,G.,2014.Biopharmaceuticalbenchmarks2014.Nat.Biotechnol.32,992–161000.doi:10.1038/nbt.304017
Wang,S.-Y.,Gao,K.,Deng,D.-L.,Cai,J.-J.,Xiao,Z.-Y.,He,L.-Q.,Jiao,H.-L.,Ye,Y.-P.,18Yang,R.-W.,Li,T.-T.,Liang,L.,Liao,W.-T.,Ding,Y.-Q.,2016.TLE4promotes19colorectalcancerprogressionthroughactivationofJNK/c-Junsignaling20pathway.Oncotarget7,2878–88.doi:10.18632/oncotarget.669421
Wiggins,T.,Kumar,S.,Markar,S.R.,Antonowicz,S.,Hanna,G.B.,2015.Tyrosine,22phenylalanine,andtryptophaningastroesophagealmalignancy:a23systematicreview.CancerEpidemiol.BiomarkersPrev.24,32–8.24doi:10.1158/1055-9965.EPI-14-098025
Wlaschin,K.F.,Nissom,P.M.,Gatti,M.deL.,Ong,P.F.,Arleen,S.,Tan,K.S.,Rink,A.,26Cham,B.,Wong,K.,Yap,M.,Hu,W.-S.,2005.ESTsequencingforgene27discoveryinChinesehamsterovarycells.Biotechnol.Bioeng.91,592–606.28doi:10.1002/bit.2051129
Wu,X.cai,Xiao,C.cui,Li,H.,Tai,Y.,Zhang,Q.,Yang,Y.,2016.Down-regulationof30Transducin-LikeEnhancerofSplitprotein4inhepatocellularcarcinoma31promotescellproliferationandepithelial-Mesenchymal-Transition.32Biochem.Biophys.Res.Commun.477,161–166.33doi:10.1016/j.bbrc.2016.06.03734
Wurm,F.,2013.CHOQuasispecies—ImplicationsforManufacturingProcesses.35Processes1,296–311.doi:10.3390/pr103029636
Xiao,S.,Shiloach,J.,Betenbaugh,M.J.,2014.Engineeringcellstoimproveprotein37expression.doi:10.1016/j.sbi.2014.03.00538
Xu,X.,Nagarajan,H.,Lewis,N.E.,Pan,S.,Cai,Z.,Liu,X.,Chen,W.,Xie,M.,Wang,W.,39Hammond,S.,Andersen,M.R.,Neff,N.,Passarelli,B.,Koh,W.,Fan,H.C.,Wang,40Jianbin,Gui,Y.,Lee,K.H.,Betenbaugh,M.J.,Quake,S.R.,Famili,I.,Palsson,41B.O.,Wang,Jun,2011.ThegenomicsequenceoftheChinesehamsterovary42(CHO)-K1cellline.Nat.Biotechnol.29,735–741.doi:10.1038/nbt.193243
Yang,J.,Weinberg,R.A.,2008.Epithelial-MesenchymalTransition:Atthe44CrossroadsofDevelopmentandTumorMetastasis.Dev.Cell.45doi:10.1016/j.devcel.2008.05.00946
Zhao,M.,Kong,L.,Liu,Y.,Qu,H.,2015.dbEMT:anepithelial-mesenchymal47transitionassociatedgeneresource.Sci.Reports201555,11459.48doi:10.1038/srep1145949
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint

39
Zhu,Y.,Meng,Q.,Wang,C.,Liu,Q.,Sun,H.,Kaku,T.,Liu,K.,2012.Organicanion1transportersinvolvedintheexcretionofbestatininthekidney.Peptides33,2265–271.doi:10.1016/j.peptides.2012.01.0073
4
preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for thisthis version posted January 30, 2020. ; https://doi.org/10.1101/2020.01.29.924894doi: bioRxiv preprint