Embed Size (px)
Citation preview

doi:10.1016/j.jmb.2006.08.068 J. Mol. Biol. (2006) 363, 878–890
Evidence that Creation of Invasion Sites Determinesthe Rate of Strand Transfer Mediated by HIV-1Reverse Transcriptase
Mark Nils Hanson1, Mini Balakrishnan1, Bernard P. Roques2
and Robert A. Bambara1,3⁎
1Department of Biochemistryand Biophysics, Center,University of RochesterMedical Center, Rochester,NY 14642, USA2Departement dePharmacochimie Moleculaireet Structurale, U266 INSERM,URA D1500 CNRS,UER des SciencesPharmaceutiques et Biologiques,4 Avenue de l'Observatoire,75270 Paris Cedex 06, France3Cancer Center, University ofRochester Medical Center,Rochester, NY 14642, USAAbbreviations used: HIV-1, humavirus; RT, reverse transcriptase; NC,FL, full length; TP, transfer product;E-mail address of the correspondi
0022-2836/$ - see front matter © 2006 E
Strand transfer during reverse transcription can produce genetic recombi-nation in human immunodeficiency virus type 1 (HIV-1) when two genomicRNAs, that are not identical, are co-packaged in the virus. Strand transferwas measured in vitro, in reactions involving primer switching from a donorto acceptor RNA template. The transfer product appeared with muchslower kinetics than full-length synthesis on the donor template. The goal ofthis study was to learn more about the transfer mechanism by defining thesteps that limit its rate. We previously proposed transfer to include the stepsof acceptor invasion, hybrid propagation, terminus transfer, and re-initiation of synthesis on the acceptor template. Unexpectedly, with ourtemplates increasing acceptor concentration increased the transfer efficiencybut had no effect on the rate of transfer. Templates with a short region ofhomology limiting hybrid propagation exhibited a slow accumulation oftransfer products, suggesting that for tested long homology templateshybrid propagation was not rate limiting. Substituting a DNA acceptor andadding Klenow polymerase accelerated re-initiation and extension exclu-sively on the DNA acceptor. This lead to a small rate increase due to fasterextension on the acceptor, suggesting re-initiation of synthesis on the testedRNA acceptors was not rate limiting. A substrate was designed in which the5′ end of the primer was single stranded, and complimentary to theacceptor, i.e. having a pre-made invasion site. With this substrate, increasingconcentrations of acceptor increased the rate of transfer. Together these datasuggest that RNase H cleavage, and dissociation of RNA fragments creatingan invasion site was rate limiting on most tested templates. When anaccessible invasion site was present, acceptor interaction at that siteinfluence the rate.
© 2006 Elsevier Ltd. All rights reserved.
Keywords: HIV-1; reverse transcriptase; strand transfer; rate-limiting;template switching
*Corresponding authorIntroduction
Replication of human immunodeficiency virustype 1 (HIV-1) requires the conversion of its plusstrand RNA genome to double-stranded DNA byreverse transcriptase (RT).1 This process includestwo obligatory DNA primer strand transfer events
n immunodeficiencynucleocapsid protein;TE, transfer efficiency.ng author:u
lsevier Ltd. All rights reserve
at each end of the template, called minus strong stopand plus strand transfer.2 Strand transfer occurswhen the elongating cDNA leaves the initialtemplate, the donor, and begins elongation onanother template, the acceptor. The virus packagestwo copies of the RNA genome, allowing for thepossibility that additional strand transfers can occurat any time during primer elongation. Such transfersresult in recombination that can produce a geneticshift3–5 in the cDNA. This shift is a mechanism ofviral evolution leading to the development of multi-drug resistant strains,6–8 viral evasion of the hostimmune system,9,10 and dispersion of mutationsintroduced by the low-fidelity RT.11–15
d.

879Rate-limiting Step of HIV-1 RT Strand Transfer
HIV-1 is a highly recombinogenic retrovirus withstudies revealing three transfer events per genomeperreplication cycle.16,17 Recombination is dispersedwith markers positioned 1 kb apart having nearlyrandom segregation,18 andmarkers only 103 bp aparthaving approximately a 12% recombination fre-quency.19,20 Studies have shown recombination tooccur primarily duringminus strand synthesis.16,21–23
The viral nucleocapsid protein (NC) is an RNAchaperone that aids in the formation of the mostthermodynamically stable RNA conformation.24,25
NC has also been demonstrated to increase theefficiency of strand transfer26–28 by assisting in theannealing of two nucleic acid strands,29–35 andincreasing RNase H activity leading to an increasedconcentration of cleavages at pause sites.36–38
Analyses of the mechanism of strand transfer fromthe ends of donor templates in vitro have consis-tently shown that transfer products appear at amuch slower rate than full-length extension on thedonor template.39–41 On templates with a shortregion of homology this has been attributed to slowRNase H cleavage of the blunt 5′ end fragment of thedonor RNA–cDNA hybrid, needed to make theDNA terminus available for transfer. It was surpris-ing, however, that templates that only allow internaltransfers, which must occur before synthesis to theend of the donor, also showed slow appearance ofthe transfer product.42–44 This suggests that thetransfer reaction involves unique slow steps that donot apply to the extension of most primers on thedonor template.Our previous work suggests that transfers can
occur by a multi-step invasion mechanism. Themechanism includes RNase H cleavage of the donorhybrid, acceptor interaction with the cleared cDNA(invasion), hybrid propagation of this interaction,and finally primer terminus transfer.36,42,45 Here, weattempted to discern which steps in this mechanismcontribute to the slow kinetics of transfer. Resultsrevealed slow steps that support the proposedinvasion mechanism, and suggest why strandtransfer occurs at a distinctly slower rate than theextension of most primers.
Results
Strand transfer reactions measured in vitro show adistinct increase in transfer efficiency with time,indicative of slow steps unique to the transferreaction.42–44 Our approach has been to accelerateor eliminate individual proposed steps in thetransfer reaction, to attempt to determine whichstep is limiting the rate of appearance of the finaltransfer product. All the templates used in this studywere designed so that once synthesis using thedonor template is completed the full-length DNAproduct cannot contribute to transfer. In this mannerwe specifically assessed the mechanism of internaltransfers in vitro. This analysis provides insight intothe mechanism of strand transfer recombinationduring synthesis of the HIV-1 genome.
Strand transfer is described here using severalparameters. The first is transfer efficiency, which isdefined as the percentage of complete primerextensions that finished on the acceptor templateand is calculated using the formula TE=(TP/TP+FL)×100 where TP is the transfer product and FL isthe full-length donor extension product. The secondis relative transfer efficiency, which is the transferefficiency at any time expressed as a percentage of thevalue at 64 min. The values are graphed as a fractionof the 100% value versus time. If transfers hadoccurred very rapidly with respect to synthesis, thepoints would be at 100% throughout the time course.Because the rate of transfer is slow compared tosynthesis, relative transfer efficiencies increase withtime. Relative transfer efficiency values for reactionsin which the transfer rate is faster approach 100% atearlier times, compared to reactions in which thetransfer rate is slower. The third is relative transferrate, which at any time, for example 16 min as usedthroughout this work, can be defined as the quotientof relative transfer efficiency values at that time.Relative transfer rate can be used to comparedifferent templates or conditions. When this valuedeviates from 1, it signifies differences in the rate ofthe transfer reaction between two template sets.
Template features altered the transfer rate
We first determined whether the template struc-ture could alter the transfer rate. We comparedtransfer rates on templates that have differentsecondary structures, as judged by the pausingpatterns of synthesis and Mfold46 predictions,despite only minor differences in sequence. Thesetemplates have been shown to display differenttransfer efficiencies.47 The initial donor (D0) andacceptor (A0) templates are shown in Figure 1(a).Based on the pausing pattern during reverse
transcription, template D0 has minimal structure,whereas D1, D2 and D3 have structures that inducepausing at different locations. When these fourdonor templates, all with the same acceptor A0,which has a region exactly homologous to D0, werecompared, there was a significant difference amongsome of the transfer rates (Figure 1(b)). Thedifference in transfer rates is clearly evident atintermediate time points. For this reason the relativetransfer efficiency at 16 min was used to comparetransfer rates. The relative transfer efficiencies at16 min were 45%, 65%, 54% and 41% for D0 throughD3, respectively. If D0 is used as a standard D3shows little change with a relative transfer rate of 0.9(41%/45%), while D1 and D2 show a modestincrease to 1.4 (65%/45%) and 1.2 (54%/45%),respectively. A1, A2, and A3 are acceptor templatesthat include the same sequence changes as D1, D2,and D3, respectively. The acceptor sequence changeswere also found to generally alter the rate of strandtransfer (Figure 1(c) to (e)). When D1 was used withA1 instead of A0 the relative transfer rate decreasedto 0.6 (39%/65%), and when D2 was used with A2instead of A0 the relative transfer rate increased to
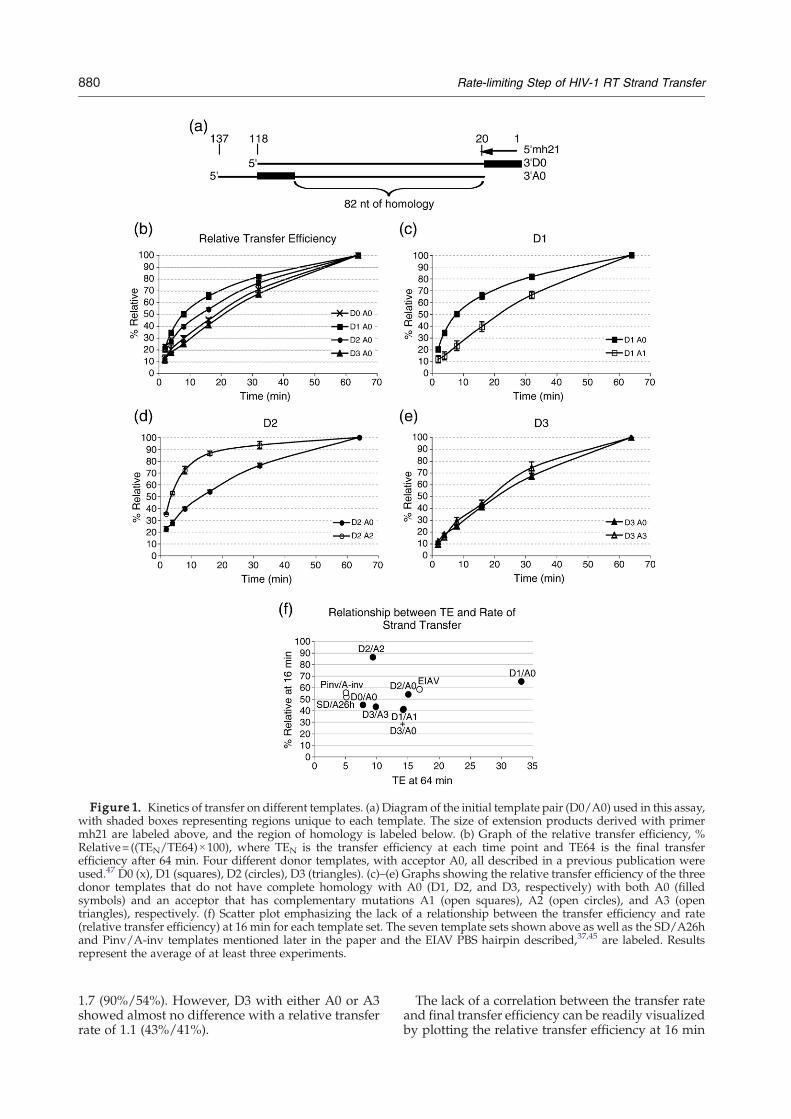
Figure 1. Kinetics of transfer on different templates. (a) Diagram of the initial template pair (D0/A0) used in this assay,with shaded boxes representing regions unique to each template. The size of extension products derived with primermh21 are labeled above, and the region of homology is labeled below. (b) Graph of the relative transfer efficiency, %Relative=((TEN/TE64)×100), where TEN is the transfer efficiency at each time point and TE64 is the final transferefficiency after 64 min. Four different donor templates, with acceptor A0, all described in a previous publication wereused.47 D0 (x), D1 (squares), D2 (circles), D3 (triangles). (c)–(e) Graphs showing the relative transfer efficiency of the threedonor templates that do not have complete homology with A0 (D1, D2, and D3, respectively) with both A0 (filledsymbols) and an acceptor that has complementary mutations A1 (open squares), A2 (open circles), and A3 (opentriangles), respectively. (f) Scatter plot emphasizing the lack of a relationship between the transfer efficiency and rate(relative transfer efficiency) at 16 min for each template set. The seven template sets shown above as well as the SD/A26hand Pinv/A-inv templates mentioned later in the paper and the EIAV PBS hairpin described,37,45 are labeled. Resultsrepresent the average of at least three experiments.
880 Rate-limiting Step of HIV-1 RT Strand Transfer
1.7 (90%/54%). However, D3 with either A0 or A3showed almost no difference with a relative transferrate of 1.1 (43%/41%).
The lack of a correlation between the transfer rateand final transfer efficiency can be readily visualizedby plotting the relative transfer efficiency at 16 min
881Rate-limiting Step of HIV-1 RT Strand Transfer
against the transfer efficiency at 64 min (Figure 1(f)).This was done for the template pairs in Figure 1(b) to(e), along with two other template sets used here(SD/A26h, Pinv/A-inv) and the EIAV PBS hairpintemplate that was used to first identify the two-stepmechanism of strand transfer.37,45 Consider, forexample, D2/A2, which had a low transfer effi-ciency and the highest relative transfer efficiency at16 min while six templates with higher transferefficiency values showed significantly lower relativetransfer efficiencies at 16 min.
NC increased the rate of the transfer reaction buttransfer remains a slow process
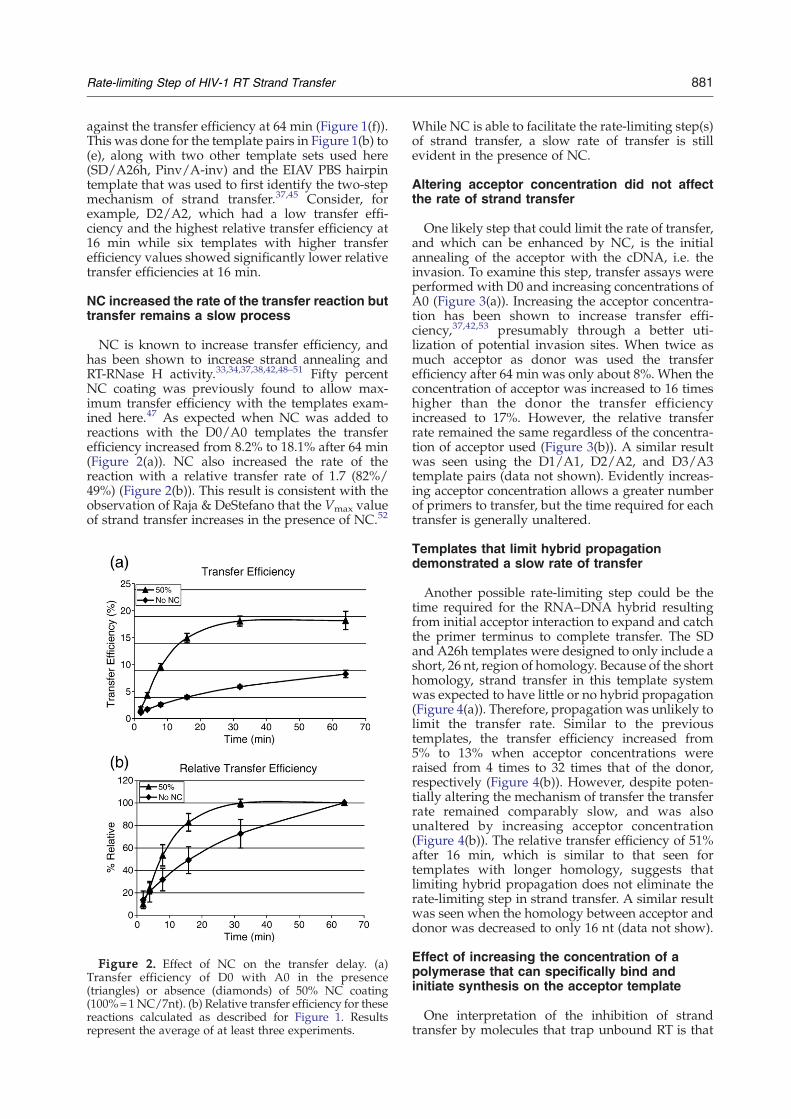
NC is known to increase transfer efficiency, andhas been shown to increase strand annealing andRT-RNase H activity.33,34,37,38,42,48–51 Fifty percentNC coating was previously found to allow max-imum transfer efficiency with the templates exam-ined here.47 As expected when NC was added toreactions with the D0/A0 templates the transferefficiency increased from 8.2% to 18.1% after 64 min(Figure 2(a)). NC also increased the rate of thereaction with a relative transfer rate of 1.7 (82%/49%) (Figure 2(b)). This result is consistent with theobservation of Raja & DeStefano that the Vmax valueof strand transfer increases in the presence of NC.52
Figure 2. Effect of NC on the transfer delay. (a)Transfer efficiency of D0 with A0 in the presence(triangles) or absence (diamonds) of 50% NC coating(100%=1NC/7nt). (b) Relative transfer efficiency for thesereactions calculated as described for Figure 1. Resultsrepresent the average of at least three experiments.
While NC is able to facilitate the rate-limiting step(s)of strand transfer, a slow rate of transfer is stillevident in the presence of NC.
Altering acceptor concentration did not affectthe rate of strand transfer
One likely step that could limit the rate of transfer,and which can be enhanced by NC, is the initialannealing of the acceptor with the cDNA, i.e. theinvasion. To examine this step, transfer assays wereperformed with D0 and increasing concentrations ofA0 (Figure 3(a)). Increasing the acceptor concentra-tion has been shown to increase transfer effi-ciency,37,42,53 presumably through a better uti-lization of potential invasion sites. When twice asmuch acceptor as donor was used the transferefficiency after 64 min was only about 8%. When theconcentration of acceptor was increased to 16 timeshigher than the donor the transfer efficiencyincreased to 17%. However, the relative transferrate remained the same regardless of the concentra-tion of acceptor used (Figure 3(b)). A similar resultwas seen using the D1/A1, D2/A2, and D3/A3template pairs (data not shown). Evidently increas-ing acceptor concentration allows a greater numberof primers to transfer, but the time required for eachtransfer is generally unaltered.
Templates that limit hybrid propagationdemonstrated a slow rate of transfer
Another possible rate-limiting step could be thetime required for the RNA–DNA hybrid resultingfrom initial acceptor interaction to expand and catchthe primer terminus to complete transfer. The SDand A26h templates were designed to only include ashort, 26 nt, region of homology. Because of the shorthomology, strand transfer in this template systemwas expected to have little or no hybrid propagation(Figure 4(a)). Therefore, propagation was unlikely tolimit the transfer rate. Similar to the previoustemplates, the transfer efficiency increased from5% to 13% when acceptor concentrations wereraised from 4 times to 32 times that of the donor,respectively (Figure 4(b)). However, despite poten-tially altering the mechanism of transfer the transferrate remained comparably slow, and was alsounaltered by increasing acceptor concentration(Figure 4(b)). The relative transfer efficiency of 51%after 16 min, which is similar to that seen fortemplates with longer homology, suggests thatlimiting hybrid propagation does not eliminate therate-limiting step in strand transfer. A similar resultwas seen when the homology between acceptor anddonor was decreased to only 16 nt (data not show).
Effect of increasing the concentration of apolymerase that can specifically bind andinitiate synthesis on the acceptor template
One interpretation of the inhibition of strandtransfer by molecules that trap unbound RT is that
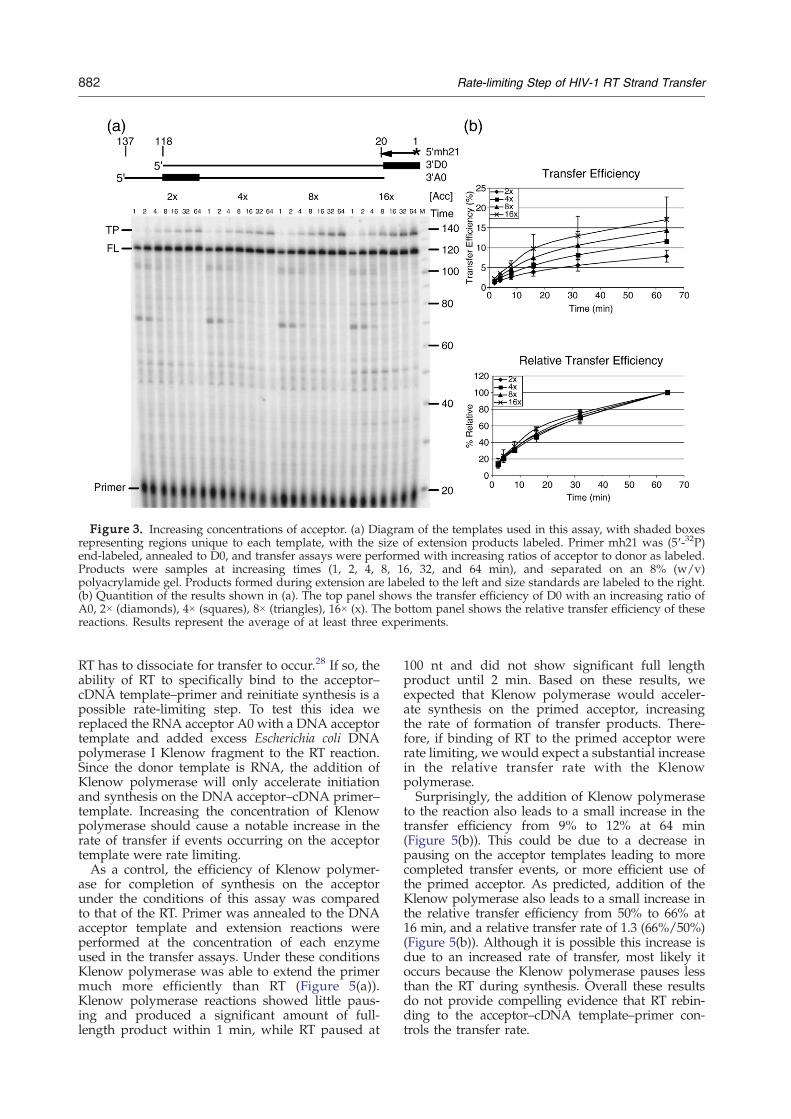
Figure 3. Increasing concentrations of acceptor. (a) Diagram of the templates used in this assay, with shaded boxesrepresenting regions unique to each template, with the size of extension products labeled. Primer mh21 was (5′-32P)end-labeled, annealed to D0, and transfer assays were performed with increasing ratios of acceptor to donor as labeled.Products were samples at increasing times (1, 2, 4, 8, 16, 32, and 64 min), and separated on an 8% (w/v)polyacrylamide gel. Products formed during extension are labeled to the left and size standards are labeled to the right.(b) Quantition of the results shown in (a). The top panel shows the transfer efficiency of D0 with an increasing ratio ofA0, 2× (diamonds), 4× (squares), 8× (triangles), 16× (x). The bottom panel shows the relative transfer efficiency of thesereactions. Results represent the average of at least three experiments.
882 Rate-limiting Step of HIV-1 RT Strand Transfer
RT has to dissociate for transfer to occur.28 If so, theability of RT to specifically bind to the acceptor–cDNA template–primer and reinitiate synthesis is apossible rate-limiting step. To test this idea wereplaced the RNA acceptor A0 with a DNA acceptortemplate and added excess Escherichia coli DNApolymerase I Klenow fragment to the RT reaction.Since the donor template is RNA, the addition ofKlenow polymerase will only accelerate initiationand synthesis on the DNA acceptor–cDNA primer–template. Increasing the concentration of Klenowpolymerase should cause a notable increase in therate of transfer if events occurring on the acceptortemplate were rate limiting.As a control, the efficiency of Klenow polymer-
ase for completion of synthesis on the acceptorunder the conditions of this assay was comparedto that of the RT. Primer was annealed to the DNAacceptor template and extension reactions wereperformed at the concentration of each enzymeused in the transfer assays. Under these conditionsKlenow polymerase was able to extend the primermuch more efficiently than RT (Figure 5(a)).Klenow polymerase reactions showed little paus-ing and produced a significant amount of full-length product within 1 min, while RT paused at
100 nt and did not show significant full lengthproduct until 2 min. Based on these results, weexpected that Klenow polymerase would acceler-ate synthesis on the primed acceptor, increasingthe rate of formation of transfer products. There-fore, if binding of RT to the primed acceptor wererate limiting, we would expect a substantial increasein the relative transfer rate with the Klenowpolymerase.Surprisingly, the addition of Klenow polymerase
to the reaction also leads to a small increase in thetransfer efficiency from 9% to 12% at 64 min(Figure 5(b)). This could be due to a decrease inpausing on the acceptor templates leading to morecompleted transfer events, or more efficient use ofthe primed acceptor. As predicted, addition of theKlenow polymerase also leads to a small increase inthe relative transfer efficiency from 50% to 66% at16 min, and a relative transfer rate of 1.3 (66%/50%)(Figure 5(b)). Although it is possible this increase isdue to an increased rate of transfer, most likely itoccurs because the Klenow polymerase pauses lessthan the RT during synthesis. Overall these resultsdo not provide compelling evidence that RT rebin-ding to the acceptor–cDNA template–primer con-trols the transfer rate.
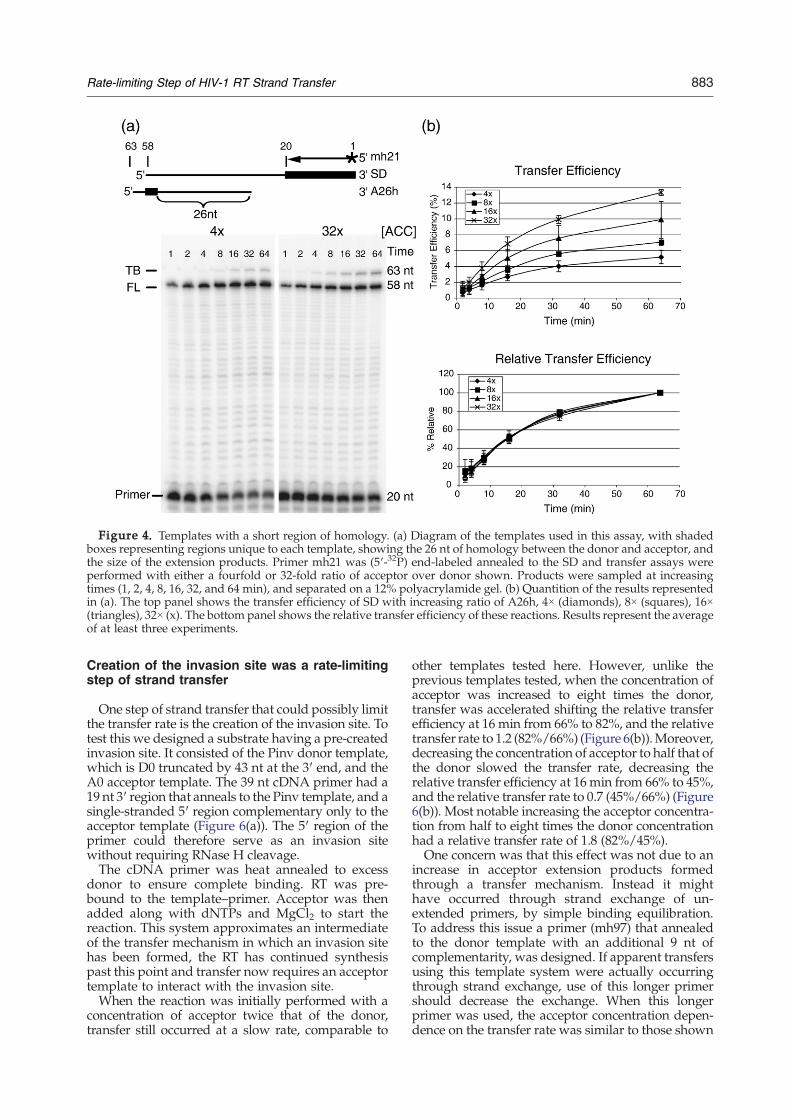
Figure 4. Templates with a short region of homology. (a) Diagram of the templates used in this assay, with shadedboxes representing regions unique to each template, showing the 26 nt of homology between the donor and acceptor, andthe size of the extension products. Primer mh21 was (5′-32P) end-labeled annealed to the SD and transfer assays wereperformed with either a fourfold or 32-fold ratio of acceptor over donor shown. Products were sampled at increasingtimes (1, 2, 4, 8, 16, 32, and 64 min), and separated on a 12% polyacrylamide gel. (b) Quantition of the results representedin (a). The top panel shows the transfer efficiency of SD with increasing ratio of A26h, 4× (diamonds), 8× (squares), 16×(triangles), 32× (x). The bottom panel shows the relative transfer efficiency of these reactions. Results represent the averageof at least three experiments.
883Rate-limiting Step of HIV-1 RT Strand Transfer
Creation of the invasion site was a rate-limitingstep of strand transfer
One step of strand transfer that could possibly limitthe transfer rate is the creation of the invasion site. Totest this we designed a substrate having a pre-createdinvasion site. It consisted of the Pinv donor template,which is D0 truncated by 43 nt at the 3′ end, and theA0 acceptor template. The 39 nt cDNA primer had a19 nt 3′ region that anneals to the Pinv template, and asingle-stranded 5′ region complementary only to theacceptor template (Figure 6(a)). The 5′ region of theprimer could therefore serve as an invasion sitewithout requiring RNase H cleavage.The cDNA primer was heat annealed to excess
donor to ensure complete binding. RT was pre-bound to the template–primer. Acceptor was thenadded along with dNTPs and MgCl2 to start thereaction. This system approximates an intermediateof the transfer mechanism in which an invasion sitehas been formed, the RT has continued synthesispast this point and transfer now requires an acceptortemplate to interact with the invasion site.When the reaction was initially performed with a
concentration of acceptor twice that of the donor,transfer still occurred at a slow rate, comparable to
other templates tested here. However, unlike theprevious templates tested, when the concentration ofacceptor was increased to eight times the donor,transfer was accelerated shifting the relative transferefficiency at 16 min from 66% to 82%, and the relativetransfer rate to 1.2 (82%/66%) (Figure 6(b)).Moreover,decreasing the concentration of acceptor to half that ofthe donor slowed the transfer rate, decreasing therelative transfer efficiency at 16min from 66% to 45%,and the relative transfer rate to 0.7 (45%/66%) (Figure6(b)). Most notable increasing the acceptor concentra-tion from half to eight times the donor concentrationhad a relative transfer rate of 1.8 (82%/45%).One concern was that this effect was not due to an
increase in acceptor extension products formedthrough a transfer mechanism. Instead it mighthave occurred through strand exchange of un-extended primers, by simple binding equilibration.To address this issue a primer (mh97) that annealedto the donor template with an additional 9 nt ofcomplementarity, was designed. If apparent transfersusing this template system were actually occurringthrough strand exchange, use of this longer primershould decrease the exchange. When this longerprimer was used, the acceptor concentration depen-dence on the transfer rate was similar to those shown
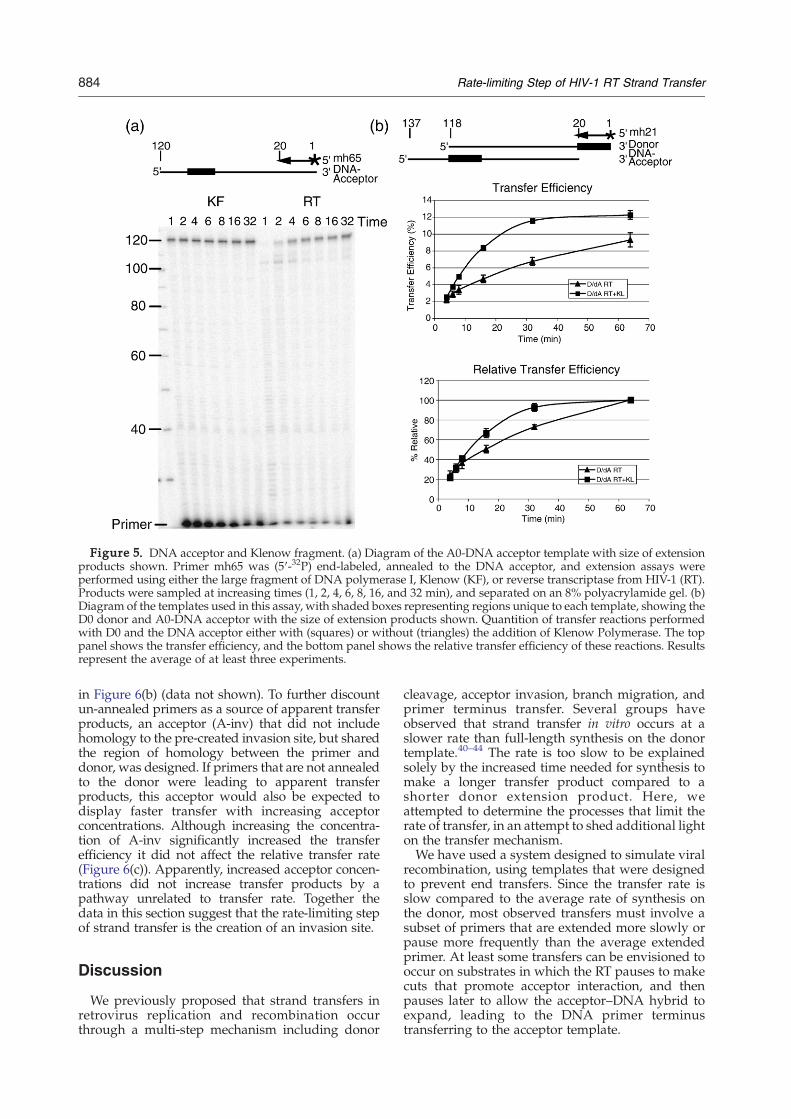
Figure 5. DNA acceptor and Klenow fragment. (a) Diagram of the A0-DNA acceptor template with size of extensionproducts shown. Primer mh65 was (5′-32P) end-labeled, annealed to the DNA acceptor, and extension assays wereperformed using either the large fragment of DNA polymerase I, Klenow (KF), or reverse transcriptase from HIV-1 (RT).Products were sampled at increasing times (1, 2, 4, 6, 8, 16, and 32 min), and separated on an 8% polyacrylamide gel. (b)Diagram of the templates used in this assay, with shaded boxes representing regions unique to each template, showing theD0 donor and A0-DNA acceptor with the size of extension products shown. Quantition of transfer reactions performedwith D0 and the DNA acceptor either with (squares) or without (triangles) the addition of Klenow Polymerase. The toppanel shows the transfer efficiency, and the bottom panel shows the relative transfer efficiency of these reactions. Resultsrepresent the average of at least three experiments.
884 Rate-limiting Step of HIV-1 RT Strand Transfer
in Figure 6(b) (data not shown). To further discountun-annealed primers as a source of apparent transferproducts, an acceptor (A-inv) that did not includehomology to the pre-created invasion site, but sharedthe region of homology between the primer anddonor, was designed. If primers that are not annealedto the donor were leading to apparent transferproducts, this acceptor would also be expected todisplay faster transfer with increasing acceptorconcentrations. Although increasing the concentra-tion of A-inv significantly increased the transferefficiency it did not affect the relative transfer rate(Figure 6(c)). Apparently, increased acceptor concen-trations did not increase transfer products by apathway unrelated to transfer rate. Together thedata in this section suggest that the rate-limiting stepof strand transfer is the creation of an invasion site.
Discussion
We previously proposed that strand transfers inretrovirus replication and recombination occurthrough a multi-step mechanism including donor
cleavage, acceptor invasion, branch migration, andprimer terminus transfer. Several groups haveobserved that strand transfer in vitro occurs at aslower rate than full-length synthesis on the donortemplate.40–44 The rate is too slow to be explainedsolely by the increased time needed for synthesis tomake a longer transfer product compared to ashorter donor extension product. Here, weattempted to determine the processes that limit therate of transfer, in an attempt to shed additional lighton the transfer mechanism.We have used a system designed to simulate viral
recombination, using templates that were designedto prevent end transfers. Since the transfer rate isslow compared to the average rate of synthesis onthe donor, most observed transfers must involve asubset of primers that are extended more slowly orpause more frequently than the average extendedprimer. At least some transfers can be envisioned tooccur on substrates in which the RT pauses to makecuts that promote acceptor interaction, and thenpauses later to allow the acceptor–DNA hybrid toexpand, leading to the DNA primer terminustransferring to the acceptor template.
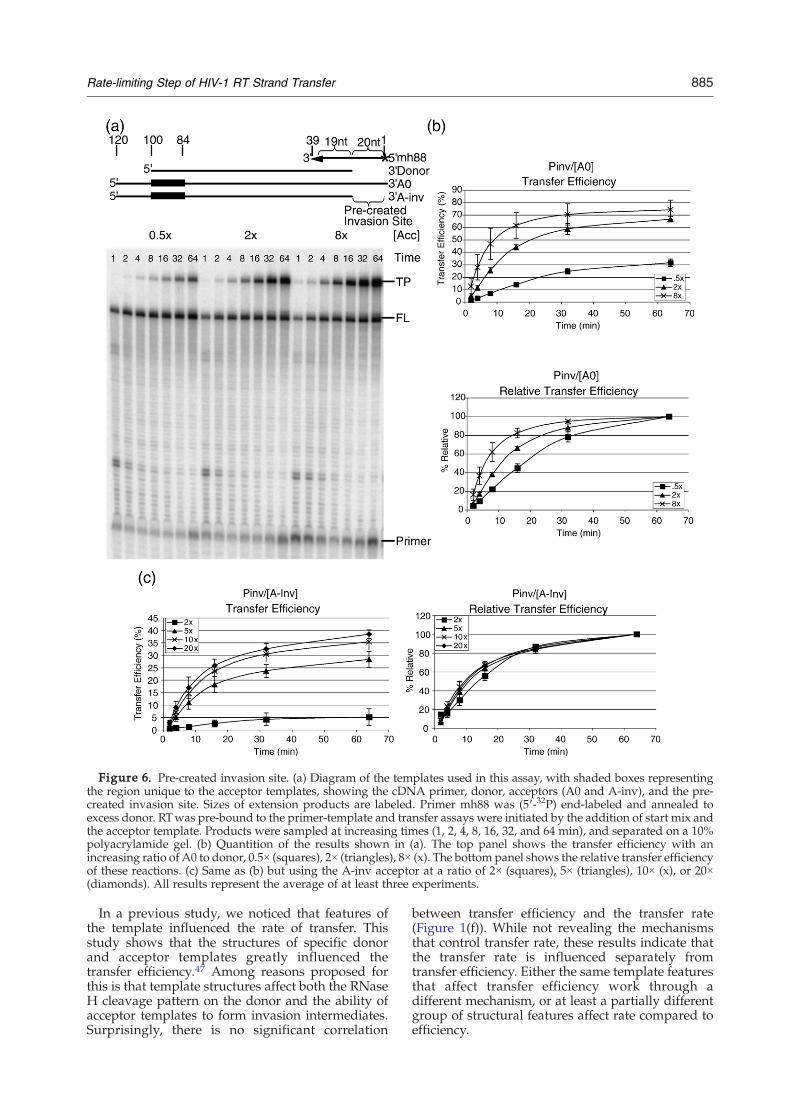
Figure 6. Pre-created invasion site. (a) Diagram of the templates used in this assay, with shaded boxes representingthe region unique to the acceptor templates, showing the cDNA primer, donor, acceptors (A0 and A-inv), and the pre-created invasion site. Sizes of extension products are labeled. Primer mh88 was (5′-32P) end-labeled and annealed toexcess donor. RTwas pre-bound to the primer-template and transfer assays were initiated by the addition of start mix andthe acceptor template. Products were sampled at increasing times (1, 2, 4, 8, 16, 32, and 64 min), and separated on a 10%polyacrylamide gel. (b) Quantition of the results shown in (a). The top panel shows the transfer efficiency with anincreasing ratio of A0 to donor, 0.5× (squares), 2× (triangles), 8× (x). The bottom panel shows the relative transfer efficiencyof these reactions. (c) Same as (b) but using the A-inv acceptor at a ratio of 2× (squares), 5× (triangles), 10× (x), or 20×(diamonds). All results represent the average of at least three experiments.
885Rate-limiting Step of HIV-1 RT Strand Transfer
In a previous study, we noticed that features ofthe template influenced the rate of transfer. Thisstudy shows that the structures of specific donorand acceptor templates greatly influenced thetransfer efficiency.47 Among reasons proposed forthis is that template structures affect both the RNaseH cleavage pattern on the donor and the ability ofacceptor templates to form invasion intermediates.Surprisingly, there is no significant correlation
between transfer efficiency and the transfer rate(Figure 1(f)). While not revealing the mechanismsthat control transfer rate, these results indicate thatthe transfer rate is influenced separately fromtransfer efficiency. Either the same template featuresthat affect transfer efficiency work through adifferent mechanism, or at least a partially differentgroup of structural features affect rate compared toefficiency.

886 Rate-limiting Step of HIV-1 RT Strand Transfer
Our general approach to identify the rate-limitingstep for transfer was to accelerate or eliminatedifferent proposed transfer steps and then measurewhether the rate of transfer increased. These stepsincluded the creation of a site on the primed donorcomplex for invasion by the acceptor, initial acceptorRNA interaction with the complex, propagation ofthe acceptor–DNA hybrid, transfer of the DNAterminus, and re-initiation and completion ofsynthesis on the acceptor template. Experimentsdesigned to probe acceptor invasion, propagation,and re-initiation showed little or no effect on the rateof strand transfer.Promoting acceptor interactions with the donor–
primer complex by leaving a region of the primerclear of RNA approximated a highly accessibleinvasion site. Significantly, increasing acceptor con-centration with this substrate lead to a faster rate oftransfer (Figure 6). This acceptor concentration effectwas not seen using conventional substrates withwhich an increase in acceptor concentrationincreased transfer efficiency but not rate (Figure 3).This demonstrated that creation of a site for efficientacceptor interaction with the cDNA, or invasion, is arate-limiting step of strand transfer on our conven-tional templates. We further interpret these results tomean that once the invasion site is created, acceptorinteraction becomes rate limiting. Acceptor interac-tion would then be the second slowest process in thetransfer reaction. However, since creation of theinvasion site and the invasion of the acceptor occurat similar rates we expect that with differenttemplates or reaction conditions acceptor annealingcould become rate limiting. Since creation of aninteraction site and then acceptor interaction aresubsequent steps, it is clear that the major determi-nant of the transfer rate is the ability of the acceptorto interact with the cDNA. However, as mentionedwhen a conventional substrate was used to measuretransfer, the transfer rate was not affected by anincrease in acceptor concentration (Figure 3). Wesuggest that the higher acceptor concentration doesnot alter the rate of formation of potential invasionsites, but allows for the use of a higher percentage ofinvasion-competent sites, accounting for theincrease in transfer efficiency.We note that the transfers seen with the Pinv/
A-inv template most likely initiated from invasionsites created during synthesis, at sites of RNase Hcleavage of the donor template, and not from thepre-created invasion site as this acceptor has nohomology with this region. Transfers by thismechanism probably occur with the Pinv/A0template pair and likely account for some of thedelay seen with this template pair.What is likely to constitute creation of the invasion
site? Our experiments suggest that a region on thecDNA that has been cleared of template RNA allowsthe substrate to experience a rapid rate of transfer. Inthe virus, template RNA cleavage is thought to occurboth during synthesis,54 and after the RT haspassed.55,56 Later cleavages would involve revisitsby one of the approximately 50 RTs present in the
HIV-1 virion. This cleavage would most likely be car-ried out by both internal cleavages57 and the 5′ RNAend-directed mechanism described previously.55,56
DeStefano investigated the transfer rate in vitrousing a similar substrate system to the oneemployed here.44 He showed some acceleration oftransfer with increased acceptor template, con-cluding that acceptor interaction was significant indetermining rate. The major basis of the slowreaction, however, remained elusive. He also pre-sented evidence that the 3′ end region of a pausedprimer, which is chased to transfer product, iscleared of donor RNA at a substantially faster ratethan appearance of transfer product. He interpretedthis to mean that primers became quickly availablefor transfer. This work predated the proposedinvasion mechanism for transfer, in which the 3′end region of the primer is not necessarily the firstposition of acceptor interaction. It is possible thatstructures in the donor–primer intermediate of hissystem, important for transfer, formed at a slow rate.The ability of NC to increase the rate of strand
transfer is also consistent with the interpretationthat RNase H cleavage and dissociation of thesmall fragments of RNA is the rate limiting process.NC is able to increase RNase H cleavage of thedonor template often increasing the number ofdistinct cleavages within regions involved with in-vasion.36–38,58 NC has also been shown to decreasethe stability of weak hybrids,59–62 leading to anincrease in strand exchange of the acceptor tem-plate with small fragments of cleaved donor.33–35,63
Both of these activities could explain how NC isable to increase the rate of strand transfer byincreasing the rate of creation of an invasion site.These interpretations also suggest why there are
large differences in transfer rates among templatepairs. Most likely templates have different rates oftransfer because they vary in their ability to createand use invasion sites. The number of templates thatuse invasion sites during a transfer reaction, leadingto completion of synthesis on the acceptor, deter-mines transfer efficiency. The speed at which theyproduce the used sites64 determines the rate oftransfer. Features of donor templates that increasethe speed of invasion site creation, such as pausing,which can lead to increased RNase H cleavage overa short region,36,37,47,54,65–67 and local sequences thatfavor RNase H cleavage and/or a more rapiddissociation of RNA fragments, are likely to increasethe rate of strand transfer. Acceptor templatestructure, which has been shown by several groupsto influence transfer,45,47,64,68–71 is likely to alsoinfluence the rate of strand transfer by determiningwhich potential invasion sites created on the donorare utilized. Since sites are likely to be created atdifferent rates, use of one group of sites over anotherwould affect the overall rate of transfer.The only other modification of the transfer
reaction that altered the rate of transfer wasacceleration of the initiation and completion ofDNA synthesis on the acceptor template. This wasaccomplished by addition of the Klenow fragment

887Rate-limiting Step of HIV-1 RT Strand Transfer
of DNA polymerase I to reactions with a DNAacceptor. The effect is to increase the amount ofpolymerase that can initiate highly efficient synth-esis after the primer has annealed to the acceptortemplate. These reaction conditions produced amoderate increase in the transfer rate (Figure 5(b)).The observed effects are most likely the result ofmore rapid primer extension, and the absence ofpausing on the acceptor template with Klenowpolymerase (Figure 5(a)). This would also suggestthat changing pausing on either the donor oracceptor templates would have predictable effectson the rate of transfer, but this is not the case. Forexample, contrary to expectations, when pausing onthe donor was increased, D3/A0 versus D0/A0, thetransfer rate was slower, but when pausing wasincreased on the acceptor, D2/A0 versus D2/A2, thetransfer rate was increased (Figure 1). Together thesedata suggest that other features of the templates arelikely to have a larger role in determining thekinetics of transfer.In summary we have shown that at least two steps
of strand transfer in our system in vitro, creation ofthe invasion site and interaction of the acceptor withthe cDNA, occur at rates distinctly slower thanprimer extension. The creation of the invasion site isthe step that limits the overall reaction. This step isenvisioned to involve RNase H cleavage of thetemplate during synthesis, movement of the synthe-sizing RT, possible revisits by other RTs and theirdissociation, and dissociation of the short RNAproducts of cleavage. Sequence and structure of thedonor template influence the transfer rate, presum-ably by influencing these steps. Structure of theacceptor template, also influences the transfer rate,presumably by favoring the use of particular sitesfor invasion over others. These results are consistentwith the proposed steps of the invasion mechanismfor strand transfer, and point out the multi-stepnature of the transfer reaction.
Materials and Methods
Reagents
Recombinant heterodimer HIV-1 reverse transcriptasewas purified as described.37,72 Chemically synthesizednucleocapsid protein (1-72) NCp7 was synthesized asdescribed.73 NC was stored at –80 °C in a buffer of 50 mMTris–HCl (pH 7.5) and 5 mM dithiothreitol. Taq polymer-ase was from Invitrogen (San Diego, CA). Radiolabeledcompounds were from Perkin-Elmer Life Sciences (Bos-ton, MA), and Micro Bio-Spin columns were from Bio-Rad(Hercules, CA). Integrated DNA Technologies, Inc. (Cor-alville, IA) synthesized all DNA primers. All otherenzymes were purchased from Roche Molecular Biochem-icals (Indianapolis, IN).
Generation of donor and acceptor templates
D0, D1, D2, D3, A0, A1, A2, and A3 DNA templateswere described.47 Pinv and A-inv DNA templates were
created by PCR amplification of a 5′ fragment of D0 orA0, respectively, using the primer mh80 (5′-TAATAC-GACTCACTATAGGGC), which includes the T7 RNApolymerase recognition sequence, and mh89 (5′-TGGT-AAACATTCTTGAGTCATCG).The SD and A26h DNA templates were purchased from
IDT. The Short D (5′-GACGCATGTGACTGATATCCGGTT-CTATAACGGTATGATGTGAAACATTCTTGAGTCCTA-TAGTGAGTCGTATTA) and Short A-10 (5′-CGGTATGAT-GTGAAACATTCTTGAGTGTATGCCTATAGTGAGTCG-TATTA) were annealed to an equal concentration of T7primer (5′-TAATACGACTCACTATAGG) by heating in TEbuffer at 65 °C for 3 min and cooling on ice.RNA templates were generated in vitro by run-off
transcription from BamHI-linearized plasmids (D0, D1,D2, D3, A0, A1, A2, A3), PCR products (Pinv, A-inv), orannealed primers (SD, A26h) using T7 RNA polymerase,as per the manufacturer's protocol (Roche MolecularBiochemicals). RNA substrates were gel-purified. TheDNA acceptor containing the same sequence as A0 waspurchased from IDT and gel-purified.
Labeling and annealing of substrates
DNA primers were labeled at the 5′ end usingpolynucleotide kinase and [γ-32P]ATP (6000 Ci/mmol).Unincorporated nucleotides were separated using P-30Micro Bio-Spin columns (Bio-Rad). For standard reactionsdonor RNA, acceptor RNA, and labeled primer at a ratioof 1:2:1.5 were brought to a volume of 10 μl in 50 mMTris–HCl (pH 8.0), 80 mM KCl, 1 mM dithiothreitol, and 1 mMEDTA, heated for 5 min at 95 °C, and slow-cooled to roomtemperature. D0, D1, D2, D3, and SD were primed withthe primer MH21 (5′-GACGCATGTGACTGATATCC).Pinv templates were heat annealed to the primer MH88
(5′-CCGGTTCTATAACGGTATGATGGTAAACATTCTT-GAGTC) with 19 nt of homology to the donor template, orthe primer MH98 (5′-CCGGTTCTATAACGGTATGATGG-TAAACATTCTTGAGTCATCGATCTG), which includes28 nt of homology with the donor template at the 3′ end,in the absence of acceptor with a donor/primer ratio of6.25:1 as described above.
Reverse transcriptase assays
Reactions were carried out in a final volume of 12.5 μl.Typical reactions were performed by incubating 32 nM ofHIV-1 reverse transcriptase for 5 min at room temperaturewith 4 nM of annealed duplex template-primer (seeabove), and 8 nM of acceptor template in 50 mM Tris–HCl (pH 8), 80 mM KCl, 1 mM dithiothreitol, and 1 mMEDTA. Reactions were started by adding dNTPs andMgCl2 to a final concentration of 50 μM and 6 mM,respectively. Reactions were incubated at 37 °C andstopped at the appropriate time by adding 1 volume of2× termination buffer (90% (v/v) formamide, 10 mMEDTA (pH 8), and 0.1% (w/v) each of bromophenol blueand xylene cyanole). Reactions with the DNA acceptorand Klenow polymerase were performed the same asdescribed above except that 3 nM of Klenow polymerasewas added immediately after the start mix. For reactionswith the Pinv templates 4 nM of donor template and0.64 nM of primer were used with described amounts ofacceptor added along with the dNTPs and MgCl2.Reaction products were resolved on an 8% (D0), 10%(Pinv), or 12% (SD) polyacrylamide–urea gel, and visua-lized using a PhosphorImager and ImageQuant software(Amersham Biosciences). Transfer efficiency (TE) was

888 Rate-limiting Step of HIV-1 RT Strand Transfer
determined using the formula TE=(TP/(FL+ TP)), whereFL is full-length extension of the donor template, and TP isfull-length extension of transfer products on the acceptortemplate. The transfer efficiencies of different templateswere normalized by comparing the transfer efficiency ateach time point (TEN) with that at 64 min (TE64), andusing the equation % Relative=((TEN/TE64)×100).
Acknowledgements
We thank Dr Vandana Purohit for helpful discus-sions and critical reading of the manuscript, alongwith Lu Gao and Min Song for helpful discussions.This work was supported by National Institutes ofHealth grant GM 049573 (to R.A.B.).
References
1. Telesnitsky, A. & Goff, S. P. (1993). Strong-stopStrand Transfer during Reverse Transcription. InReverse Transcriptase (Skalka, A. M. & Goff, S. P.,eds), pp. 49–83, Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, NY.
2. Telesnitsky, A. & Goff, S. P. (1997). Reverse Transcrip-tase and the Generation of Retroviral DNA. In Retro-viruses (Coffin, J. M., Hughes, S. H. & Varmus, H. E.,eds), pp. 121–160, Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, NY.
3. Clavel, F., Hoggan, M. D., Willey, R. L., Strebel, K.,Martin, M. A. & Repaske, R. (1989). Genetic recombi-nation of human immunodeficiency virus. J. Virol. 63,1455–1459.
4. Goodrich, D. W. & Duesberg, P. H. (1990). Evidencethat retroviral transduction is mediated by DNA notby RNA. Proc. Natl Acad. Sci. USA, 87, 3604–3608.
5. Hu, W. S. & Temin, H. M. (1990). Genetic conse-quences of packaging two RNA genomes in oneretroviral particle: pseudodiploidy and high rate ofgenetic recombination. Proc. Natl Acad. Sci. USA, 87,1556–1560.
6. Gu, Z., Gao, Q., Faust, E. A. &Wainberg, M. A. (1995).Possible involvement of cell fusion and viral recombi-nation in generation of human immunodeficiencyvirus variants that display dual resistance to AZT and3TC. J. Gen. Virol. 76, 2601–2605.
7. Gunthard, H. F., Leigh-Brown, A. J., D'Aquila, R. T.,Johnson, V. A., Kuritzkes, D. R., Richman, D. D. &Wong, J. K. (1999). Higher selection pressure fromantiretroviral drugs in vivo results in increasedevolutionary distance in HIV-1 pol. Virology, 259,154–165.
8. Kellam, P. & Larder, B. A. (1995). Retroviral recombi-nation can lead to linkage of reverse transcriptasemutations that confer increased zidovudine resis-tance. J. Virol. 69, 669–674.
9. Pekrun, K., Shibata, R., Igarashi, T., Reed, M.,Sheppard, L., Patten, P. A. et al. (2002). Evolution ofa human immunodeficiency virus type 1 variant withenhanced replication in pig-tailed macaque cells byDNA shuffling. J. Virol. 76, 2924–2935.
10. Robertson, D. L., Hahn, B. H. & Sharp, P. M. (1995).Recombination in AIDS viruses. J. Mol. Evol. 40,249–259.
11. Boyer, J. C., Bebenek, K. & Kunkel, T. A. (1992).
Unequal human immunodeficiency virus type 1reverse transcriptase error rates with RNA and DNAtemplates. Proc. Natl Acad. Sci. USA, 89, 6919–6923.
12. Ji, J. P. & Loeb, L. A. (1992). Fidelity of HIV-1 reversetranscriptase copying RNA in vitro. Biochemistry, 31,954–958.
13. Preston, B. D., Poiesz, B. J. & Loeb, L. A. (1988).Fidelity of HIV-1 reverse transcriptase. Science, 242,1168–1171.
14. Roberts, J. D., Bebenek, K. & Kunkel, T. A. (1988). Theaccuracy of reverse transcriptase from HIV-1. Science,242, 1171–1173.
15. Weber, J. & Grosse, F. (1989). Fidelity of human im-munodeficiency virus type I reverse transcriptase incopying natural DNA. Nucl. Acids Res. 17, 1379–1393.
16. Jetzt, A. E., Yu, H., Klarmann, G. J., Ron, Y., Preston,B. D. & Dougherty, J. P. (2000). High rate ofrecombination throughout the human immunodefi-ciency virus type 1 genome. J. Virol. 74, 1234–1240.
17. Zhuang, J., Jetzt, A. E., Sun, G., Yu, H., Klarmann, G.,Ron, Y. et al. (2002). Human immunodeficiency virustype 1 recombination: Rate, fidelity, and putative hotspots. J. Virol. 76, 11273–11282.
18. Rhodes, T., Wargo, H. & Hu, W.-S. (2003). High ratesof human immunodeficiency virus type 1 recombina-tion: near-random segregation of markers one kilo-base apart in one round of viral replication. J. Virol. 77,11193–11200.
19. Chen, J., Rhodes, T. D. & Hu, W.-S. (2005). Compar-ison of the genetic recombination rates of humanimmunodeficiency virus type 1 in macrophages and TCells. J. Virol. 79, 9337–9340.
20. Rhodes, T. D., Nikolaitchik, O., Chen, J., Powell, D. &Hu, W.-S. (2005). Genetic recombination of humanimmunodeficiency virus type 1 in one round of viralreplication: Effects of genetic distance, target cells,accessory genes, and lack of high negative interferencein crossover events. J. Virol. 79, 1666–1677.
21. Anderson, J. A., Teufel, R. J., II, Yin, P. D. & Hu, W. S.(1998). Correlated template-switching events duringminus-strand DNA synthesis: a mechanism for highnegative interference during retroviral recombination.J. Virol. 72, 1186–1194.
22. Hu, W. S. & Temin, H. M. (1992). Effect of gammaradiation on retroviral recombination. J. Virol. 66,4457–4463.
23. Zhang, J., Tang, L.-Y., Li, T., Ma, Y. & Sapp, C. M.(2000). Most retroviral recombinations occur duringminus-strand DNA synthesis. J. Virol. 74, 2313–2322.
24. Darlix, J. L., Lapadat-Tapolsky, M., de Rocquigny, H.& Roques, B. P. (1995). First glimpses at structure-function relationships of the nucleocapsid protein ofretroviruses. J. Mol. Biol. 254, 523–537.
25. Rein, A., Henderson, L. E. & Levin, J. G. (1998).Nucleic-acid-chaperone activity of retroviral nucleo-capsid proteins: significance for viral replication.Trends Biochem. Sci. 23, 297–301.
26. Allain, B., Lapadat-Tapolsky, M., Berlioz, C. & Darlix,J. L. (1994). Transactivation of the minus-strand DNAtransfer by nucleocapsid protein during reversetranscription of the retroviral genome. EMBO J. 13,973–981.
27. Darlix, J. L., Vincent, A., Gabus, C., de Rocquigny, H.& Roques, B. (1993). Trans-activation of the 5′ to 3′viral DNA strand transfer by nucleocapsid proteinduring reverse transcription of HIV1 RNA. C.R. Acad.Sci. III, 316, 763–771.
28. Rodriguez-Rodriguez, L., Tsuchihashi, Z., Fuentes,G. M., Bambara, R. A. & Fay, P. J. (1995). Influence

889Rate-limiting Step of HIV-1 RT Strand Transfer
of human immunodeficiency virus nucleocapsidprotein on synthesis and strand transfer by thereverse transcriptase in vitro. J. Biol. Chem. 270,15005–15011.
29. Negroni, M. & Buc, H. (1999). Recombination duringreverse transcription: An evaluation of the role of thenucleocapsid protein. J. Mol. Biol. 286, 15–31.
30. DeStefano, J. J. (1996). Interaction of human immuno-deficiency virus nucleocapsid protein with a structuremimicking a replication intermediate. Effects onstability, reverse transcriptase binding, and strandtransfer. J. Biol. Chem. 271, 16350–16356.
31. Golinelli, M. P. & Hughes, S. H. (2003). Secondarystructure in the nucleic acid affects the rate of HIV-1nucleocapsid-mediated strand annealing. Biochemis-try, 42, 8153–8162.
32. Hsu, M., Rong, L., de Rocquigny, H., Roques, B. P. &Wainberg, M. A. (2000). The effect of mutations in theHIV-1 nucleocapsid protein on strand transfer in cell-free reverse transcription reactions.Nucl. Acids Res. 28,1724–1729.
33. Lapadat-Tapolsky, M., Pernelle, C., Borie, C. & Darlix,J. L. (1995). Analysis of the nucleic acid annealingactivities of nucleocapsid protein from HIV-1. Nucl.Acids Res. 23, 2434–2441.
34. Tsuchihashi, Z. & Brown, P. O. (1994). DNA strandexchange and selective DNA annealing promoted bythe human immunodeficiency virus type 1 nucleo-capsid protein. J. Virol. 68, 5863–5870.
35. You, J. C. & McHenry, C. S. (1994). Human immuno-deficiency virus nucleocapsid protein acceleratesstrand transfer of the terminally redundant sequencesinvolved in reverse transcription. J. Biol. Chem. 269,31491–31495.
36. Chen, Y., Balakrishnan, M., Roques, B. P. & Bambara,R. A. (2003). Steps of the acceptor invasion mechanismfor HIV-1(minus strand strong stop transfer. J. Biol.Chem. 278, 38368–38375.
37. Roda, R. H., Balakrishnan, M., Hanson, M. N., Wohrl,B. M., Le Grice, S. F. J., Roques, B. P. et al. (2003). Roleof the reverse transcriptase, nucleocapsid protein, andtemplate structure in the two-step transfer mechan-ism in retroviral recombination. J. Biol. Chem. 278,31536–31546.
38. Peliska, J. A., Balasubramanian, S., Giedroc, D. P. &Benkovic, S. J. (1994). Recombinant HIV-1 nucleocap-sid protein accelerates HIV-1 reverse transcriptasecatalyzed DNA strand transfer reactions and mod-ulates RNase H activity. Biochemistry, 33, 13817–13823.
39. Chen, Y., Balakrishnan, M., Roques, B. P., Fay, P. J.& Bambara, R. A. (2003). Mechanism of minusstrand strong stop transfer in HIV-1 reverse tran-scription. J. Biol. Chem. 278, 8006–8017.
40. Peliska, J. A. & Benkovic, S. J. (1992). Mechanism ofDNA strand transfer reactions catalyzed by HIV-1reverse transcriptase. Science, 258, 1112–1118.
41. Luo, G. X. & Taylor, J. (1990). Template switching byreverse transcriptase during DNA synthesis. J. Virol.64, 4321–4328.
42. Balakrishnan, M., Roques, B. P., Fay, P. J. & Bambara,R. A. (2003). Template dimerization promotes anacceptor invasion-induced transfer mechanism duringhuman immunodeficiency virus type 1 minus-strandsynthesis. J. Virol. 77, 4710–4721.
43. DeStefano, J. J., Mallaber, L.M., Rodriguez-Rodriguez,L., Fay, P. J. & Bambara, R. A. (1992). Requirements forstrand transfer between internal regions of hetero-polymer templates by human immunodeficiency virusreverse transcriptase. J. Virol. 66, 6370–6378.
44. DeStefano, J. J. (1994). Kinetic analysis of the catalysisof strand transfer from internal regions of hetero-polymeric RNA templates by human immunodefi-ciency virus reverse transcriptase. J. Mol. Biol. 243,558–567.
45. Roda, R. H., Balakrishnan, M., Kim, J. K., Roques, B. P.,Fay, P. J. & Bambara, R. A. (2002). Strand transferoccurs in retroviruses by a pause-initiated two-stepmechanism. J. Biol. Chem. 277, 46900–46911.
46. Zuker, M. (2003). Mfold web server for nucleic acidfolding and hybridization prediction. Nucl. Acids Res.31, 3406–3415.
47. Hanson, M. N., Balakrishnan, M., Roques, B. P. &Bambara, R. A. (2005). Effects of donor and acceptorRNA structures on the mechanism of strand transferby HIV-1 reverse transcriptase. J. Mol. Biol. 353,772–787.
48. Chen, Y., Balakrishnan, M., Roques, B. P. & Bambara,R. A. (2005). Acceptor RNA cleavage profile supportsan invasion mechanism for HIV-1(minus strandtransfer. J. Biol. Chem. 280, 14443–14452.
49. Guo, J., Wu, T., Anderson, J., Kane, B. F., Johnson,D. G., Gorelick, R. J. et al. (2000). Zinc fingerstructures in the human immunodeficiency virus type1 nucleocapsid protein facilitate efficient minus- andplus-strand transfer. J. Virol. 74, 8980–8988.
50. Levin, J. G., Guo, J., Rouzina, I. & Musier-Forsyth, K.(2005). Nucleic acid chaperone activity of HIV-1nucleocapsid protein: critical role in reverse transcrip-tion and molecular mechanism. Prog. Nucl. Acid Res.Mol. Biol. 80, 217–286.
51. Wisniewski, M., Chen, Y., Balakrishnan, M., Palaniap-pan, C., Roques, B. P., Fay, P. J. & Bambara, R. A.(2002). Substrate requirements for secondary cleavageby HIV-1 Reverse transcriptase RNase H. J. Biol. Chem.277, 28400–28410.
52. Raja, A. & DeStefano, J. J. (1999). Kinetic analysis ofthe effect of HIV nucleocapsid protein (NCp) oninternal strand transfer reactions. Biochemistry, 38,5178–5184.
53. Reynolds, M. A., Arnold, L. J., Jr, Almazan, M. T.,Beck, T. A., Hogrefe, R. I. & Metzler, M. D. (1994). et alTriple-strand-forming methylphosphonate oligodeox-ynucleotides targeted to mRNA efficiently blockprotein synthesis. Proc. Natl Acad. Sci. USA, 91,12433–12437.
54. Purohit, V., Balakrishnan,M., Kim, B. &Bambara, R. A.(2005). Evidence that HIV-1 reverse transcriptaseemploys the DNA 3′ end-directed primary/secondaryRNase H cleavage mechanism during synthesis andstrand transfer. J. Biol. Chem. 280, 40534–40543.
55. Wisniewski, M., Balakrishnan, M., Palaniappan, C.,Fay, P. J. & Bambara, R. A. (2000). Unique progressivecleavage mechanism of HIV reverse transcriptaseRNase H. Proc. Natl Acad. Sci. USA, 97, 11978–11983.
56. Wisniewski, M., Balakrishnan, M., Palaniappan, C.,Fay, P. J. & Bambara, R. A. (2000). The sequentialmechanism of HIV reverse transcriptase RNase H.J. Biol. Chem. 275, 37664–37671.
57. Schultz, S. J., Zhang, M. & Champoux, J. J. (2004).Recognition of internal cleavage sites by retroviralRNases H. J. Mol. Biol. 344, 635–652.
58. Iwatani, Y., Rosen, A. E., Guo, J., Musier-Forsyth, K. &Levin, J. G. (2003). Efficient initiation of HIV-1 reversetranscription i. Requirement for RNA sequencesdownstream of the primer binding site abrogated bynucleocapsid protein-dependent primer-templateinteractions. J. Biol. Chem. 278, 14185–14195.
59. Hargittai, M. R. S., Gorelick, R. J., Rouzina, I. &

890 Rate-limiting Step of HIV-1 RT Strand Transfer
Musier-Forsyth, K. (2004). Mechanistic insights intothe kinetics of HIV-1 nucleocapsid Protein-facilitatedtRNA annealing to the primer binding site. J. Mol. Biol.337, 951–968.
60. Heath, M. J., Derebail, S. S., Gorelick, R. J. &DeStefano, J. J. (2003). Differing roles of the N-terminal and C-terminal zinc fingers in humanimmunodeficiency virus nucleocapsid proteinenhanced nucleic acid annealing. J. Biol. Chem. 278,30755–30763.
61. Hong, M. K., Harbron, E. J., O'Connor, D. B., Guo, J.,Barbara, P. F., Levin, J. G. & Musier-Forsyth, K. (2003).Nucleic acid conformational changes essential forHIV-1 nucleocapsid protein-mediated inhibition ofself-priming in minus-strand transfer. J. Mol. Biol. 325,1–10.
62. Urbaneja, M. A., Wu, M., Casas-Finet, J. R. & Karpel,R. L. (2002). HIV-1 nucleocapsid protein as a nucleicacid chaperone: spectroscopic study of its helix-destabilizing properties, structural binding specificity,and annealing activity. J. Mol. Biol. 318, 749–764.
63. Heath, M. J. & Destefano, J. J. (2005). A complemen-tary single-stranded docking site is required forenhancement of strand exchange by human immuno-deficiency virus nucleocapsid protein on substratesthat model viral recombination. Biochemistry, 44,3915–3925.
64. Berkhout, B., Vastenhouw, N. L., Klasens, B. I. &Huthoff, H. (2001). Structural features in the HIV-1repeat region facilitate strand transfer during reversetranscription. RNA, 7, 1097–1114.
65. DeStefano, J. J., Bambara, R. A. & Fay, P. J. (1994). Themechanism of human immunodeficiency virus reversetranscriptase- catalyzed strand transfer from internalregions of heteropolymeric RNA templates. J. Biol.Chem. 269, 161–168.
66. Suo, Z. & Johnson, K. A. (1997). Effect of RNA secon-dary structure on RNA cleavage catalyzed by HIV-1reverse transcriptase. Biochemistry, 36, 12468–12476.
67. Suo, Z. & Johnson, K. A. (1997). RNA secondarystructure switching during DNA synthesis catalyzedby HIV-1 reverse transcriptase. Biochemistry, 36,14778–14785.
68. Heilman-Miller, S. L., Wu, T. & Levin, J. G. (2004).Alteration of nucleic acid structure and stabilitymodulates the efficiency of minus-strand transfermediated by the HIV-1 nucleocapsid protein. J. Biol.Chem. 279, 44154–44165.
69. Moumen, A., Polomack, L., Unge, T., Veron, M., Buc,H. & Negroni, M. (2003). Evidence for a mechanism ofrecombination during reverse transcription depen-dent on the structure of the acceptor RNA. J. Biol.Chem. 278, 15973–15982.
70. Negroni, M. & Buc, H. (2000). Copy-choice recombi-nation by reverse transcriptases: Reshuffling ofgenetic markers mediated by RNA chaperones. Proc.Natl Acad. Sci. USA, 97, 6385–6390.
71. Topping, R., Demoitie, M. A., Shin, N. H. &Telesnitsky, A. (1998). Cis-acting elements requiredfor strong stop acceptor template selection duringmoloney murine leukemia virus reverse transcription.J. Mol. Biol. 281, 1–15.
72. Pandey, V. N., Kaushik, N., Rege, N., Sarafianos, S. G.,Yadav, P. N. &Modak, M. J. (1996). Role of methionine184 of human immunodeficiency virus type-1 reversetranscriptase in the polymerase function and fidelityof DNA synthesis. Biochemistry, 35, 2168–2179.
73. de Rocquigny, H., Ficheux, D., Gabus, C., Fournie-Zaluski, M. C., Darlix, J. L. & Roques, B. P. (1991). Firstlarge scale chemical synthesis of the 72 amino acidHIV-1 nucleocapsid protein NCp7 in an active form.Biochem. Biophys. Res. Commun. 180, 1010–1018.
Edited by J. Karn
(Received 7 July 2006; received in revised form 11 August 2006; accepted 15 August 2006)Available online 30 August 2006