Embed Size (px)
Citation preview

Evidence of Spinocerebellar Mossy FiberSegregation in the Juvenile Staggerer
Cerebellum
ZHONGQI JI, QIAN JIN, AND MICHAEL W. VOGEL*Maryland Psychiatric Research Center, University of Maryland Medical School,
Baltimore, Maryland 21228
ABSTRACTDevelopmental and experimental studies of climbing fiber andmossy fiber connectivity in
the cerebellum have suggested that Purkinje cells are the critical organizing elements forconnectivity patterns. This hypothesis is supported by evidence that spinocerebellar mossyfiber projections are abnormally diffuse in P25 sg/sgmutant mice in which the differentiationof a reduced number of sg/sg Purkinje cells is blocked due to a cell autonomous defect.However, mossy fiber distribution may be disrupted in sg/sg mutants not because of thePurkinje cell deficits, but because of the death of virtually all granule cells following the 4thpostnatal week. To test this hypothesis, we have analyzed the distribution of wheat germagglutinin-horseradish peroxidase (WGA-HRP)–labeled spinocerebellar mossy fiber termi-nals in sg/sg mutants at the end of the period of granule cell genesis (postnatal day[P]12–P13) and before massive granule cell death (P16). Two percent WGA-HRP was injectedinto the lower thoracic/upper lumbar region of the spinal cord of eight homozygous sg/sgmutants (P12–P16) and five controls (1/sg and 1/1). We have found that spinocerebellarmossy fibers segregate into distinct terminal fields in the anterior cerebellar lobules of P12to P16 sg/sg mutants, although the medial-lateral distribution of spinocerebellar mossyfiber projections is different from controls. The results from this study and previous analy-sis of sg/sg mutants support the hypothesis that topographic cues are expressed in theearly postnatal staggerer mutant, but mossy fiber terminals become disorganized or retractas granule cells die in the older staggerer mutant. J. Comp. Neurol. 378:354–362,1997. r 1997 Wiley-Liss, Inc.
Indexing terms: pattern formation; Purkinje cells; granule cells; compartments; mouse mutant
The neurons of the cerebellar cortex are innervated by adiverse network of afferents that are segregated intodistinct parasagittal compartments within specific lobules(Ito, 1984). The major output neurons of the cerebellarcortex, the Purkinje cells, are innervated by inferiorolivary neuron climbing fibers. The major class of cerebel-lar interneurons, granule cells, are innervated by mossyfibers from diverse sources throughout the central nervoussystem, including neurons in the spinal cord. Duringdevelopment and in adults, Purkinje cells differentiallyexpress a variety of molecules (e.g., Zebrin, P-path, calbin-din) that delineate parasagittal compartments within thecerebellum (Hawkes et al., 1993; Oberdick, 1994; Soteloand Wassef, 1991). The terminal distribution of climbingfibers and mossy fibers are often organized within theborders of the Purkinje cell compartments (Gravel et al.,1987; Gravel and Hawkes, 1990; Hawkes et al., 1992;Sotelo and Wassef, 1991), although there are exceptions(Akintunde and Eisenman, 1995; Gravel and Hawkes,
1990; Ji and Hawkes, 1994; Matsushita and Yaginuma,1989). The congruence of Purkinje cell compartments andafferent projection maps has suggested that Purkinje cellsprovide molecular labels for afferent maps of the cerebel-lum (Hawkes et al., 1993; Sotelo and Wassef, 1991).Staggerer (sg) is an autosomal recessive mutation that
causes a cell-autonomous, lineage-specific block in thedevelopment of the Purkinje cell (Herrup and Mullen,1979a,b, 1981; Messer et al., 1991; Sidman et al., 1962). Acandidate gene for staggerer, RORa, was recently identi-fied (Hamilton et al., 1996). The RORa gene is selectively
Contract grant sponsor: N.I.H.; Contract grant number: NS29277; Con-tract grant sponsor: University of Maryland Medical School, SRIS grant.*Correspondence to: Michael W. Vogel, Maryland Psychiatric Research
Center, University of Maryland Medical School, Maple and Locust Streets,Spring Grove Hospital Grounds, P.O. Box 21247, Baltimore, MD 21228.Received 11 June 1996; Revised 12 September 1996; Accepted 25 October
1996
THE JOURNAL OF COMPARATIVE NEUROLOGY 378:354–362 (1997)
r 1997 WILEY-LISS, INC.

expressed in Purkinje cells from at least embyronic day (E)14, and the gene product may interact with a thyroid-hormone signaling pathway involved in Purkinje cellmaturation (Hamilton et al., 1996). By birth, sg/sg Pur-kinje cells are atrophic and ectopically positioned withinthe cerebellar cortex. Their numbers are reduced by 75%at birth either due to agenesis or death of postmitoticneurons during embryogenesis (Herrup andMullen, 1981).Virtually 100% of the granule cells and 60% of the olivaryneurons die in the staggerermutant secondary to Purkinjecell defects (Herrup, 1983; Zanjani et al., 1990). Manygranule cells in the staggerer mutant die after they havemigrated into the internal granule cell layer (IGL): Theperiod of maximum granule cell degeneration in thismutant is from P16 to 21 days. Virtually all granule cellsare dead by 33 days postnatal (Landis and Sidman, 1978).The role of Purkinje cells as organizers for cerebellar
afferents is supported by studies of mossy fiber distribu-tion in rodents with granule cell or Purkinje cell deficits(Arsenio-Nunes et al., 1988). Spinocerebellar mossy fiberterminals in P25 staggerer mutants with intrinsic defectsin Purkinje cell differentiation are homogeneously distrib-uted throughout the vermis (Arsenio-Nunes et al., 1988).The failure to form normal mossy fiber projections in thestaggerer mutant is thought to be due to the inability ofstaggerer Purkinje cells to provide positional cues becauseof their intrinsic developmental block (Sotelo and Wassef,1991). The implication is that sg/sg Purkinje cells are notdifferentiated into biochemically distinct compartments.In contrast, in rodents with granule cell deficits—X-irradiated rats and weaver mutant mice—mossy fiberterminals form segregated terminal fields. Arsenio-Nuneset al. (1988) have argued that granule cells do not contrib-ute to pattern formation on the basis of the relativelynormal mossy fiber projection patterns in the X-irradiatedrats andweavermutants.In their study of spinocerebellar mossy fiber afferents in
the staggerermutant, however,Arsenio-Nunes et al. (1988)analyzed mossy fiber terminal distribution at P25, whichis aftermost granule cells have degenerated. The degenera-tion of granule cells in the staggerer mutant raises thepossibility that mossy fiber terminals may have segre-gated normally, but then become disorganized because ofthe loss of their target neurons. Developmental studies ofmossy fiber projections suggest that mossy fiber afferentsmay initially contact Purkinje cells and sort out accordingto Purkinje cell topographic cues (Ji and Hawkes, 1995).However, the mossy fibers eventually form synapses ongranule cells, and there is experimental evidence thatgranule cells play a role in the maintenance of topographicmossy fiber projections. Mossy fiber distribution is dis-rupted in the agranular anterior lobules of the meandertail mutant (Eisenman and Arlinghaus, 1991), and recentchimera studies suggest that granule cells, but not Pur-kinje cells, are a primary site of gene action in this mutant(Hamre and Goldowitz, 1995). Disruption of granule celldevelopment with the antimitotic agent methylazoxy-methanol acetate (MAM) results in a partial desegregationin the pattern of mossy fiber afferents (Ji and Hawkes,1996). Finally, the segregation of spinocerebellar mossyfiber afferents is maintained in the 1/Lc mutant in whichvirtually all Purkinje cells degenerate after the 1st week ofdevelopment, but 10% of the granule cells survive (Caddyand Biscoe, 1979; Vogel and Prittie, 1994).
To determine whether the disorganization of mossyfibers in the staggerer mutant is a primary defect due toPurkinje cell deficiencies independent of the depletion ofgranule cells, we have analyzed the distribution of spinocer-ebellar mossy fiber terminals in juvenile staggerer pups atP12 to P16, after the period of granule cell birth and beforethe bulk of granule cell death. The results of the studyindicate that positional cues are expressed in the develop-ing sg/sg mutant cerebellum, and suggest that granulecells play a role in the stabilization of mossy fiber afferentprojections.
MATERIALS AND METHODS
Animals
Control (1/1 and 1/sg) and homozygous staggerer (sg/sg) juvenile pups were obtained from two sources in thisstudy. Staggerer (sg/sg) pups were bred either from cross-ing C57BL/6J-d se 1/1 1 sg heterozygotes or crossingB6D1-1/sg heterozygotes. To generate B6D1-1/sg hetero-zygotes, C57BL/6J-d se 1/1 1 sg heterozygotes weremated with DBA/1J mice. B6D1-1/sg progeny were identi-fied by their black coat color and they were mated with oneanother to generate sg/sg and control pups. The day ofbirth was considered P0. The B6D1-1/sg breeding pairswere produced in order to take advantage of a simplesequence length polymorphism (SSLP) closely linked tothe staggerer locus that can be used to genetically identifysg/sg mutants before their phenotype is apparent. How-ever, because P12 to P16 pups were analyzed in this study,sg/sg and control (1/1 or 1/sg) pups could be identified bytheir behavior and cerebellar morphology. We did notdetect any differences in the extent of cerebellar pathologybetween sg/sg mice on the different genetic backgrounds.All mice were maintained at animal care facilities at theMaryland Psychiatric Research Center.
WGA-HRP labeling
The distribution of spinocerebellar mossy fiber termi-nals was analyzed by orthograde labeling with wheat germagglutinin–horseradish peroxidase (WGA-HRP: Sigma) asdescribed previously (Vogel and Prittie, 1994). Control andsg/sg pups at P12 and P15 were anesthetized by inhala-tion with Metophane, and two to three segments of thelower thoracic-upper lumbar vertebral column were ex-posed by a dorsal laminectomy. Then 0.2 to 0.5 µl ofWGA-HRP (2% in distilled water) was pressure injectedbilaterally in the lower thoracic/upper lumbar region witha pulled glass micropipette connected to a 10-µl Hamiltonsyringe. The skin over the laminectomy was closed withsutures and the mouse allowed to recover. Injected micewere killed 18–24 hours after surgery by cardiac perfusionwhile under deep Avertin anesthesia. Each mouse wasperfused with 30–50 ml of warm saline (0.9% NaCl)followed by 50–80ml of 4% cold paraformaldehyde in 0.1Mphosphate buffer (PBS, pH 7.2). The fixed cerebella wereremoved from the skull and cryoprotected overnight in20% sucrose in PBS. Within the next 1–2 days, cerebellawere quick frozen in optimal cutting temperature (OCT),sectioned coronally at 32 µm, and collected in cold 0.1 MPBS. Labeling of WGA-HRP filled terminals was demon-strated by using tetramethyl benzidine (TMB) as de-scribed previously (DeOlmos et al., 1978). In brief, floatingsections were preincubated in 5% nickel ammonium sul-fate for 5 minutes before transferring the sections to the
MOSSY FIBER DISTRIBUTION 355

incubation solution for 1 hour (0.00675% TMB, 0.19%sodium nitroprusside, and 0.006% H2O2 (pH 4.3). Addi-tional H2O2 was added at 20-minute intervals. Stainedsections were mounted on subbed slides, allowed to air dry,and then counterstained with cresyl violet.Over 30 P12 to P17 sg/sg and control pups were injected
with WGA-HRP. Of these, eight homozygous staggerermutants (one P12, four P13, and three P16) and five1/1 or1/sg controls (two P13, two P16, and one P17) survived thesurgery, and, when processed, their cerebellum had accept-able levels of WGA-HRP labeling. Two of the controlcerebella (one P13 and one P16) with WGA-HRP mossyfiber labelingwere also processed for calbindin immunohis-tochemistry. Two of the sg/sg mutant cerebella (P16) withWGA-HRP mossy fiber labeling were processed for calbin-din immunohistochemistry in addition to two sg/sg (P12and P16) mutant cerebella in which the mossy fiberlabeling had not succeeded.
Calbindin immunohistochemistry
Antibody staining for calbindin was used as a specificmarker for Purkinje cells in the control and sg/sg cer-ebella. Alternate 32-µm sections were incubated overnightin calbindin primary monoclonal antibody (Sigma) diluted1/5,000 in TCT (0.1% Triton X-100, 0.6% Carrageenan,0.02% NaN3 in PBS). Sections were then rinsed threetimes (10 minutes each) in PBS before incubating thesections for 2 hours at room temperature in rabbit anti-mouse secondary antibodies diluted 1/100 in TCT. Label-ing of the secondary antibodies was performed using anavidin biotin complex (ABC) kit from Vector Laboratories,Inc. Sections were rinsed 3 times in PBS before stainingwith 0.05% diaminobenzidine (DAB) and 0.009% H2O2.Calbindin-labeled sections were dehydrated in alcohol,mounted, and coverslipped.
RESULTS
Cerebellar size and cytoarchitecturein the juvenile sg/sgmutant
The cell autonomous defect in Purkinje cell maturationin the sg/sg mutant causes profound changes in the sizeand cytoarchitecture of the developing cerebellum. Asshown in Figure 1A,B the sg/sg cerebellum is abnormallysmall by the end of the period of granule cell genesis, due toextensive Purkinje and granule cell loss. In control mice atP12 to P16, Purkinje cells line up in a continuous mono-layer between the granule cell and molecular layers. Insg/sg mutants at P12 to P16, the surviving Purkinje cellsare distributed in a diffuse layer throughout the cortex ofthe cerebellar hemispheres and vermis as shown by anti-body labeling with the Purkinje cell marker, calbindin(Fig. 1B,C). Irregular clusters of Purkinje cells alignedalong the edge of the cerebellar cortex are indicated byarrows in Figure 1C. There is invariably an elongatedcluster of Purkinje cells spanning the midline in thevermal region (Figs. 1C, 4C). Although virtually all gran-ule cells eventually degenerate in the sg/sg mutant, atP12 to P16 there are still appreciable numbers of granulecells that form an internal granule cell layer (Figs. 2, 3).The sg/sg granule cells that migrate to the internalgranule cell layer appear to differentiate normally, includ-ing extending long parallel fibers (Vogel et al., 1993).
Medial to lateral distributionof spinocerebellar mossy fiber terminals
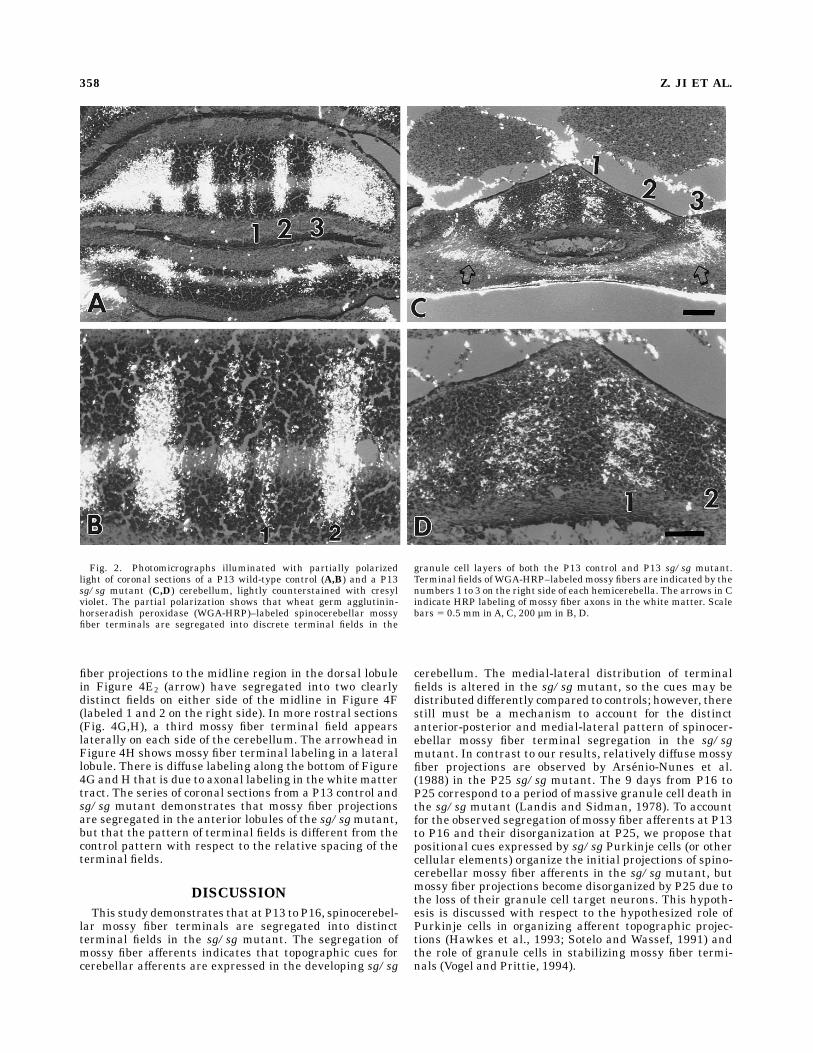
The terminals of spinocerebellar mossy fibers originat-ing from the lumbar region of the spinal cord werevisualized by orthograde labeling with WGA-HRP. Incontrol mice, spinocerebellar mossy fibers project primar-ily to anterior vermal lobules I–V, with a smaller projectionto vermal posterior lobules VIII and IX (Vogel and Prittie,1994). Within each lobule, mossy fiber terminals aresegregated into discrete terminal fields. There are gener-ally three terminal fields on each side of the midline inanterior lobules, a narrow field aligned close to the midlineand two additional fields lateral to the midline band(labeled 1 to 3 in Figs. 2A,B and 4A–D). Whereas ArsenioNunes et al. (1988) report that spinocerebellar mossy fiberterminals in the P25 sg/sg mutant are diffusely distrib-uted in the cerebellar vermis, we find that in P13 and P16sg/sgmutant cerebella, spinocerebellar mossy fiber termi-nals in anterior and posterior regions of the cerebellarvermis are segregated into discrete terminal fields (Figs.2–4). Although there were variations among differentcerebella in the strength and density of labeling, thedistribution of labeled mossy fiber terminals was consis-tent in all of the sg/sg and control cerebella. Becausemossy fiber terminal labeling was generally weaker inposterior lobules, we have focused our analysis on WGA-HRP labeling patterns in the anterior half of the sg/sgmutant and control cerebellum.The spinocerebellar mossy fiber terminals in the ante-
rior lobules of the juvenile sg/sgmutant are segregated inup to three discrete terminal fields on each side of thecerebellum (Fig. 2C,D; the terminal fields on the right sideof the cerebellum are labeled from 1 to 3 starting from themidline). For comparison of mossy fiber terminal distribu-tions, the photomicrographs of control sections (Fig. 2A,B)show mossy fiber terminal labeling in anterior lobule II.The photomicrographs of the sg/sgmutant (Fig. 2C,D) arealso taken from the anterior half of the cerebellum, butbecause of the severe shrinkage and loss of foliation in thesg/sg cerebellum, it is difficult to identify the preciselobule. HRP-labeled mossy fiber terminals in each half ofthe control lobule are segregated in a narrow mossy fiberterminal field along the midline (designated number 1 inFig. 2A,B) and two bilaterally symmetrical fields lateral tothe midline band (designated number 2 and 3). The samenumber of mossy fiber terminal fields are present in thesg/sgmutant, but the distribution of the terminal fields isdifferent. At the rostrocaudal level of the cerebellumshown in Figure 2C,D, the most striking change is that thefirst and second terminal fields from the midline appear tohave moved laterally and become thicker. In Figure 2C,the third lateral band (labeled number 3) is difficult todiscern because it blends in with heavy axonal labeling inthe white matter. Diffuse HRP labeling of axonal tracts inthe white matter is indicated by arrows in Figure 2C. Asshown by the higher-magnification photomicrograph inFigure 2D, the WGA-HRP–labeled mossy fiber terminalsin the sg/sg mutant are distributed within the internalgranule cell layer, and the terminal fields are clearlysegregated into distinct columns.The change in mossy fiber projection patterns in the
sg/sg mutant is not simply the result of a more diffusedistribution of mossy fiber terminals. The terminal fieldsformed by spinocerebellar mossy fiber afferents in the
356 Z. JI ET AL.

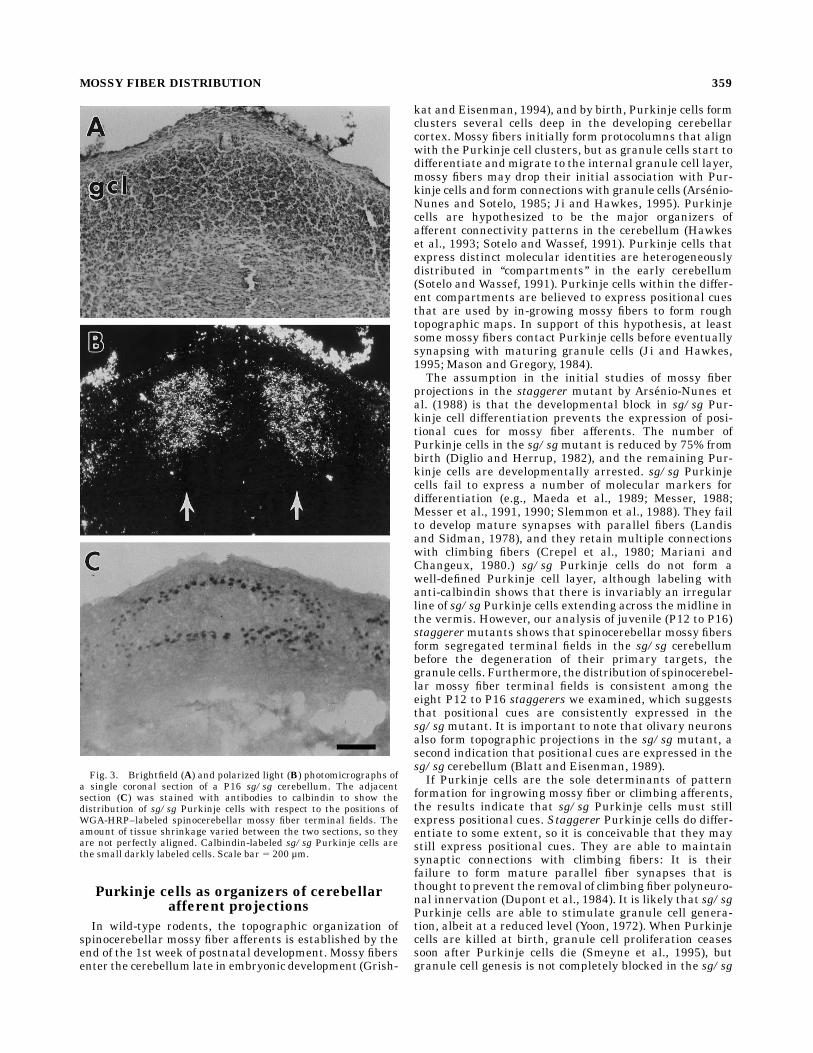
anterior lobules of the cerebellum have well-defined bor-ders and are positioned within the granule cell layer. Asshown in an example from a P16 sg/sg mutant, there arestill an appreciable number of surviving granule cells thatform an easily recognizable granule cell layer at this age(Fig. 3A). Spinocerebellar mossy fiber afferents project tothe granule cell layer, and in this example, the terminalsfill a column that extends throughout the granule celllayer (compare Fig. 3A,B). There does not appear to be anysimple relationship between the distribution of sg/sgPurkinje cells and the distribution of labeled mossy fiberterminals at this age. As shown in Figure 3C, there is anirregular line of sg/sg Purkinje cells that extends forseveral hundred microns across the midline. This line ofPurkinje cells is coincident with both amossy fiber free gapacross the midline as well as the two medial-most mossyfiber terminal fields.
Rostral to caudal distributionof spinocerebellar mossy fiber terminals
The terminal fields of spinocerebellar mossy fiber affer-ents extend throughout the anterior lobules in both con-trols and sg/sg mutants (Fig. 4). In the anterior lobules of
control cerebella, mossy fiber terminal fields are clearlysegregated into distinct terminal fields throughout indi-vidual lobules. The distribution of HRP-labeled mossyfiber terminals is shown through lobules III (Fig. 4A) andII (Fig. 4B–D) for the control cerebellum. In the sg/sgmutant, mossy fiber terminal labeling appears to bediffusely distributed near the center of the cerebellum(Fig. 4E), but in more rostral sections the mossy fiberterminals become segregated into distinct terminal fields.The first section shown for the sg/sg cerebellum (Fig. 4 E1,E2) was taken from approximately the middle of thecerebellum. The photomicrograph in Figure 4E1 was takenwith completely polarized light to show that mossy fiberprojections appear to project primarily to a large mass intothe middle of the vermis. This projection pattern closelyresembles the pattern described for P25 staggerers (Arse-nio-Nunes et al., 1988). However, a photomicrograph of thesame section taken with partially polarized light (Fig. 4E2)shows that most of theWGA-HRP labeling in Figure 4E1 isin the white matter (black double arrows). The segregationof mossy fiber terminals into distinct fields in the juvenilesg/sg mutant becomes apparent in more rostral sectionsthrough the anterior cerebellum (Fig. 4F–H). The mossy
Fig. 1. Comparison of cerebellar size and the distribution ofPurkinje cells labeled with antibodies to calbindin in coronal sectionsof a P13 wild-type cerebellum (A) and a P12 sg/sg mutant (B). Amontage of the P12 sg/sg mutant is shown at a higher magnification(C) to illustrate the ectopic distribution of calbindin-labeled sg/sg
Purkinje cells. Purkinje cells are irregularly distributed throughoutthe margins of the cerebellar cortex (arrows). There is often a linearcluster of Purkinje cells that spans the midline (double arrows). Scalebars 5 2 mm inA, B, 0.5 mm in C.
MOSSY FIBER DISTRIBUTION 357
fiber projections to the midline region in the dorsal lobulein Figure 4E2 (arrow) have segregated into two clearlydistinct fields on either side of the midline in Figure 4F(labeled 1 and 2 on the right side). In more rostral sections(Fig. 4G,H), a third mossy fiber terminal field appearslaterally on each side of the cerebellum. The arrowhead inFigure 4H shows mossy fiber terminal labeling in a laterallobule. There is diffuse labeling along the bottom of Figure4G andH that is due to axonal labeling in the white mattertract. The series of coronal sections from a P13 control andsg/sg mutant demonstrates that mossy fiber projectionsare segregated in the anterior lobules of the sg/sgmutant,but that the pattern of terminal fields is different from thecontrol pattern with respect to the relative spacing of theterminal fields.
DISCUSSION
This study demonstrates that at P13 to P16, spinocerebel-lar mossy fiber terminals are segregated into distinctterminal fields in the sg/sg mutant. The segregation ofmossy fiber afferents indicates that topographic cues forcerebellar afferents are expressed in the developing sg/sg
cerebellum. The medial-lateral distribution of terminalfields is altered in the sg/sg mutant, so the cues may bedistributed differently compared to controls; however, therestill must be a mechanism to account for the distinctanterior-posterior and medial-lateral pattern of spinocer-ebellar mossy fiber terminal segregation in the sg/sgmutant. In contrast to our results, relatively diffuse mossyfiber projections are observed by Arsenio-Nunes et al.(1988) in the P25 sg/sg mutant. The 9 days from P16 toP25 correspond to a period of massive granule cell death inthe sg/sg mutant (Landis and Sidman, 1978). To accountfor the observed segregation of mossy fiber afferents at P13to P16 and their disorganization at P25, we propose thatpositional cues expressed by sg/sg Purkinje cells (or othercellular elements) organize the initial projections of spino-cerebellar mossy fiber afferents in the sg/sg mutant, butmossy fiber projections become disorganized by P25 due tothe loss of their granule cell target neurons. This hypoth-esis is discussed with respect to the hypothesized role ofPurkinje cells in organizing afferent topographic projec-tions (Hawkes et al., 1993; Sotelo and Wassef, 1991) andthe role of granule cells in stabilizing mossy fiber termi-nals (Vogel and Prittie, 1994).
Fig. 2. Photomicrographs illuminated with partially polarizedlight of coronal sections of a P13 wild-type control (A,B) and a P13sg/sg mutant (C,D) cerebellum, lightly counterstained with cresylviolet. The partial polarization shows that wheat germ agglutinin-horseradish peroxidase (WGA-HRP)–labeled spinocerebellar mossyfiber terminals are segregated into discrete terminal fields in the
granule cell layers of both the P13 control and P13 sg/sg mutant.Terminal fields ofWGA-HRP–labeledmossy fibers are indicated by thenumbers 1 to 3 on the right side of each hemicerebella. The arrows in Cindicate HRP labeling of mossy fiber axons in the white matter. Scalebars 5 0.5 mm inA, C, 200 µm in B, D.
358 Z. JI ET AL.
Purkinje cells as organizers of cerebellarafferent projections
In wild-type rodents, the topographic organization ofspinocerebellar mossy fiber afferents is established by theend of the 1st week of postnatal development. Mossy fibersenter the cerebellum late in embryonic development (Grish-
kat and Eisenman, 1994), and by birth, Purkinje cells formclusters several cells deep in the developing cerebellarcortex. Mossy fibers initially form protocolumns that alignwith the Purkinje cell clusters, but as granule cells start todifferentiate and migrate to the internal granule cell layer,mossy fibers may drop their initial association with Pur-kinje cells and form connectionswith granule cells (Arsenio-Nunes and Sotelo, 1985; Ji and Hawkes, 1995). Purkinjecells are hypothesized to be the major organizers ofafferent connectivity patterns in the cerebellum (Hawkeset al., 1993; Sotelo and Wassef, 1991). Purkinje cells thatexpress distinct molecular identities are heterogeneouslydistributed in ‘‘compartments’’ in the early cerebellum(Sotelo and Wassef, 1991). Purkinje cells within the differ-ent compartments are believed to express positional cuesthat are used by in-growing mossy fibers to form roughtopographic maps. In support of this hypothesis, at leastsome mossy fibers contact Purkinje cells before eventuallysynapsing with maturing granule cells (Ji and Hawkes,1995; Mason and Gregory, 1984).The assumption in the initial studies of mossy fiber
projections in the staggerer mutant by Arsenio-Nunes etal. (1988) is that the developmental block in sg/sg Pur-kinje cell differentiation prevents the expression of posi-tional cues for mossy fiber afferents. The number ofPurkinje cells in the sg/sgmutant is reduced by 75% frombirth (Diglio and Herrup, 1982), and the remaining Pur-kinje cells are developmentally arrested. sg/sg Purkinjecells fail to express a number of molecular markers fordifferentiation (e.g., Maeda et al., 1989; Messer, 1988;Messer et al., 1991, 1990; Slemmon et al., 1988). They failto develop mature synapses with parallel fibers (Landisand Sidman, 1978), and they retain multiple connectionswith climbing fibers (Crepel et al., 1980; Mariani andChangeux, 1980.) sg/sg Purkinje cells do not form awell-defined Purkinje cell layer, although labeling withanti-calbindin shows that there is invariably an irregularline of sg/sg Purkinje cells extending across the midline inthe vermis. However, our analysis of juvenile (P12 to P16)staggerermutants shows that spinocerebellar mossy fibersform segregated terminal fields in the sg/sg cerebellumbefore the degeneration of their primary targets, thegranule cells. Furthermore, the distribution of spinocerebel-lar mossy fiber terminal fields is consistent among theeight P12 to P16 staggerers we examined, which suggeststhat positional cues are consistently expressed in thesg/sgmutant. It is important to note that olivary neuronsalso form topographic projections in the sg/sg mutant, asecond indication that positional cues are expressed in thesg/sg cerebellum (Blatt and Eisenman, 1989).If Purkinje cells are the sole determinants of pattern
formation for ingrowing mossy fiber or climbing afferents,the results indicate that sg/sg Purkinje cells must stillexpress positional cues. Staggerer Purkinje cells do differ-entiate to some extent, so it is conceivable that they maystill express positional cues. They are able to maintainsynaptic connections with climbing fibers: It is theirfailure to form mature parallel fiber synapses that isthought to prevent the removal of climbing fiber polyneuro-nal innervation (Dupont et al., 1984). It is likely that sg/sgPurkinje cells are able to stimulate granule cell genera-tion, albeit at a reduced level (Yoon, 1972). When Purkinjecells are killed at birth, granule cell proliferation ceasessoon after Purkinje cells die (Smeyne et al., 1995), butgranule cell genesis is not completely blocked in the sg/sg
Fig. 3. Brightfield (A) and polarized light (B) photomicrographs ofa single coronal section of a P16 sg/sg cerebellum. The adjacentsection (C) was stained with antibodies to calbindin to show thedistribution of sg/sg Purkinje cells with respect to the positions ofWGA-HRP–labeled spinocerebellar mossy fiber terminal fields. Theamount of tissue shrinkage varied between the two sections, so theyare not perfectly aligned. Calbindin-labeled sg/sg Purkinje cells arethe small darkly labeled cells. Scale bar 5 200 µm.
MOSSY FIBER DISTRIBUTION 359
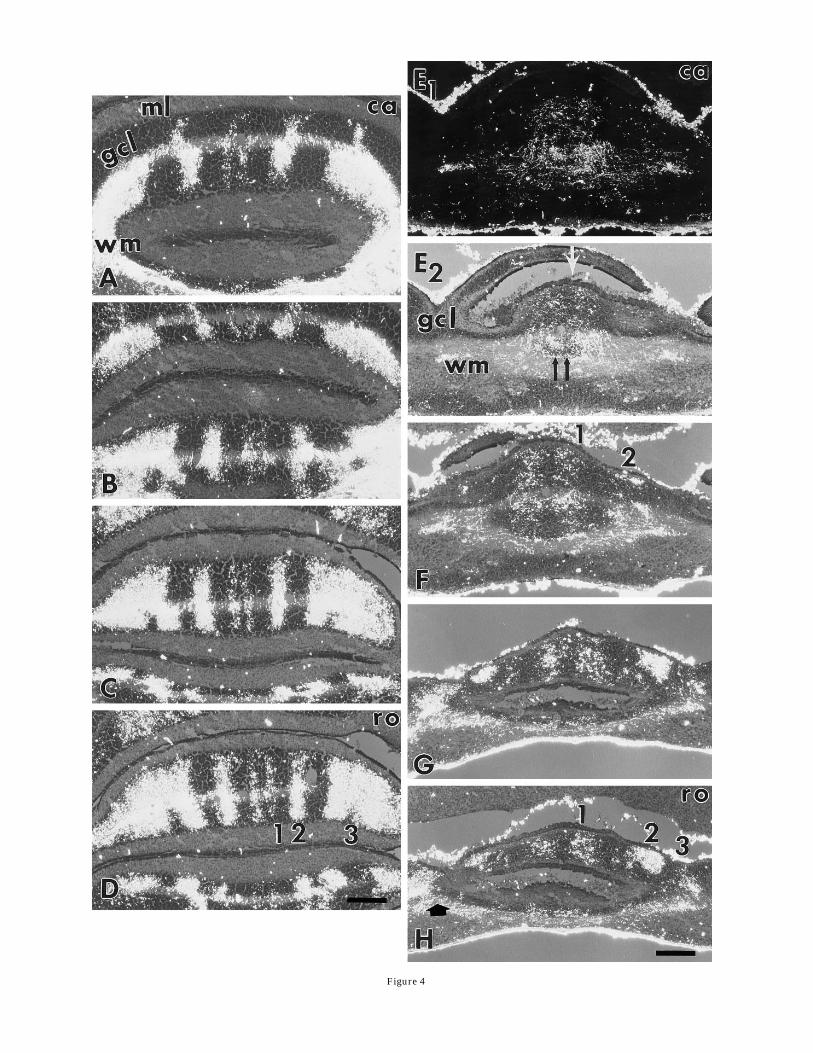
Figure 4

mutant. Finally, sg/sg Purkinje cells express calbindin,which is one of the markers that distinguishes Purkinjecell heterogeneities during embryonic development (Wassefet al., 1985). Sg/sg Purkinje cells do not appear to berandomly distributed throughout the mutant cerebellum;in particular, sg/sg Purkinje cells are arrayed in a roughline across the midline in the vermis. This suggests thatsg/sg Purkinje cells could still provide positional cues forspinocerebellar mossy fiber afferents based on their posi-tion during early development. Although spinocerebellarmossy fiber terminals in the P13 to P16 sg/sg mutant aredistributed into six distinct terminal fields in the granulecell layer of anterior lobules as in controls, there aredistinct differences in the anterior-posterior and medial-lateral distribution of terminal fields. Mossy fiber projec-tions appear relatively disorganized in the center of thecerebellum, and more rostrally the two terminal fieldsadjacent to the midline are broader and are spaced furtherapart than the corresponding terminal fields in controls.The lateral terminal fields in the sg/sgmutant also appearto have been translocated laterally. One possible explana-tion for the observed results is that sg/sg Purkinje cellsprovide positional cues for afferents, but the changes inmossy fiber projections in the sg/sgmutant are due to thedramatic reduction in sg/sg Purkinje cell numbers andtheir altered lobular distribution.An alternative possibility is that there are cues other
than Purkinje cells that can be used by mossy fiberafferents to attain their appropriate positions. The embry-onic projections of climbing and mossy fibers are topo-graphically organized as they enter the cerebellum, sug-gesting that pathway guidance cues could play a role inestablishing topographic projections (Grishkat and Eisen-man, 1994; Paradies and Eisenman, 1993; Sotelo et al.,1984). Cerebellar nuclear neurons could also provide posi-tional cues to afferent projections (see discussion by Leclercet al., 1988), and neuronal activity may play a role inforcing or maintaining the segregation of mossy fiberterminals among their granule cell targets (Tolbert et al.,1994). We are currently attempting to distinguish betweenthe roles of Purkinje cells and other cues in establishingmossy fiber projection patterns by examining the distribu-tions of WGA-HRP–labeled mossy fiber afferents and
sg/sg Purkinje cells during early postnatal development.If Purkinje cells provide positional cues through interac-tions between Purkinje cell protocolumns and ingrowingafferents in the young neonate (Arsenio-Nunes and Sotelo,1985; Ji and Hawkes, 1995), then spinocerebellar mossyfiber afferents should codistribute with sg/sg Purkinjecells in the sg/sg neonate.
Granule cells and the stabilizationof mossy fiber afferents
The segregation of mossy fiber afferents in the P13–P16staggerer mutants indicates that positional cues are ex-pressed in the staggerer cerebellum. Yet, at P25 Arsenio-Nunes et al. (1988) report that spinocerebellar mossy fiberafferents are diffusely distributed in the cerebellar vermis.Because we have not yet analyzed sg/sgmutants at P25 orolder we cannot rule out the possibility that the differencesbetween the two studies may be due to differences instaggerer mutant genetic backgrounds or methodologicaldetails. However, a more interesting alternative hypoth-esis that we are currently investigating is that the spinocer-ebellarmossy fiber projections become disorganized follow-ing the degeneration of most granule cells after P16. Thereis increasing evidence that granule cells play a role instabilizingmossy fiber projections. Vogel and Prittie (1994)demonstrated recently that spinocerebellar mossy fiberafferents project to the appropriate lobules and segregateinto relatively normal parasagittal bands in the Lurchermutant, despite the degeneration of virtually all Purkinjecells following the 1st week of postnatal development. Thisresult indicates that granule cells may stabilize mossyfiber terminals following the death of virtually all Purkinjecells. If granule cell migration and foliation patterns aredisturbed by partial ablation of the external granule celllayer with low-doseMAM injections, mossy fiber terminalsbecome more homogeneously distributed within the ver-mal lobules (Ji and Hawkes, 1996). The distribution ofspinocerebellar mossy fiber terminals is also disrupted inthe agranular anterior lobes of the meander tail cerebel-lum (Eisenman and Arlinghaus, 1991). Evidence wasrecently presented that granule cells, but not Purkinjecells, are intrinsically defective in meander tail (Hamreand Goldowitz, 1995), which indicates that the aberrantprojection patterns in the anterior lobules are due togranule cell defects. The stabilization of mossy fiber-granule cell synapses many involve neuronal activity;mossy fiber terminals aremore diffusely distributed follow-ing blockade of synaptic activity with N-methyl-D-aspar-tate and g-aminobutyric acid receptor antagonists (Tolbertet al., 1994). To investigate the role of granule cells instabilizing mossy fiber projections, in future experimentsthe distribution of spinocerebellar mossy fiber afferents inthe staggerer mutant will be analyzed in older mutants asthe number of granule cells declines. If granule cell targetneurons are necessary for the maintenance of mossy fiberprojections, we expect that mossy fiber projections willbecome more diffuse, and possibly less numerous, asgranule cells degenerate.
ACKNOWLEDGMENTS
This work was supported by grants to M.W.V. from theNIH (NS 29277) and a SRIS grant from the University ofMaryland Medical School. The authors thank Drs. Ro-salinda Roberts and Douglas Frost for their expert advice
Fig. 4. Polarized light photomicrographs of four coronal sections ofthe anterior lobules of a P13 wild-type (A–D) and a P13 sg/sgmutant(E–H) cerebella show that WGA-HRP–labeled spinocerebellar mossyfiber terminals form segregated fields. The sections extend from closeto the middle of the cerebellum (A,E) rostrally to the anterior end ofthe cerebellum (D,H). The photomicrograph in E1 is taken withcompletely polarized light, whereas in all of the other photomicro-graphs, the illumination was only partially polarized so that thecytoarchitecture of the cerebellum could be seen along with theWGA-HRP labeling. The positions of the terminal fields are indicatedby numbers on the right side of the control (D) and mutant hemicer-ebella (F,H). E2: The mass of heavy WGA-HRP labeling in the whitematter is centered around a small slice of a lobule that was just barelyincluded in this section (black double arrows). In addition, there is alighter mossy fiber projection to the granule cell layer that is clusteredaround the midline of the lobule indicated with a white arrow. There isa narrow gap in the terminal fields at the midline, suggesting that theprojections form two separate fields. F: The small slice of lobuleobscured by mossy fiber labeling in E2 has emerged as a separatelobule, and it is innervated by mossy fibers that are segregated intotwo distinct terminal fields across the midline. G,H: Three mossy fiberterminal fields are evident in more rostral sections. ca, caudal; ro,rostral; ml, molecular layer; gcl, granule cell layer; wm, white matter.Scale bars 5 200 µm.
MOSSY FIBER DISTRIBUTION 361

and criticisms and Ms. Sharon Candeloro for her secre-tarial assistance.
LITERATURE CITED
Akintunde, A., and L.M. Eisenman (1995) External cuneocerebellar projec-tion and Purkinje cell zebrin II bands—a direct comparison of parasag-ittal banding in the mouse cerebellum. J. Chem. Neuroanat. 7:75–86.
Arsenio Nunes, M.L., and C. Sotelo (1985) Development of the spinocerebel-lar system in the postnatal rat. J. Comp. Neurol. 237:291–306.
Arsenio-Nunes, M.L., C. Sotelo, and R. Wehrle (1988) Organization ofspinocerebellar projection map in three types of agranular cerebellum:Purkinje cells vs. granule cells as organizer element. J. Comp. Neurol.273:120–136.
Blatt, G.J., and L.M. Eisenman (1989) Regional and topographic organiza-tion of the olivocerebellar projection in homozygous staggerer (sg/sg)mutant mice: an anterograde and retrograde tracing study. Neurosci-ence 30:703–715.
Caddy, K.W.T., and T.J. Biscoe (1979) Structural and quantitative studieson the normal C3H and Lurcher mutant mouse. Philos. Trans. R. Soc.Lond. [Biol.] 287:67–201.
Crepel, F., N. Delhaye-Bouchaud, J.M. Gaustavino, and I. Sampaio (1980)Multiple innervation of cerebellar Purkinje cells by climbing fibers instaggerermutant mouse. Nature 283:483–484.
DeOlmos, J., H. Hardy, and L. Heimer (1978) The afferent connections ofthe main and the accesory olfactory bulb formations in the rat: anexperimental HRP-study. J. Comp. Neurol. 181:213–244.
Diglio, T., and K. Herrup (1982)A significant fraction of the adult number ofmature cerebellar Purkinje cells first appears between postnatal days16 and 30. Soc. Neurosci. Abstr. 8:636.
Dupont, J.L., E. Fournier, R. Gardette, and F. Crepel (1984) Effect ofexcitatory amino acids on Purkinje cell dendrites in cerebellar slicesfrom normal and staggerermice. Neuroscience 12:613–619.
Eisenman, L.M., and L.E. Arlinghaus (1991) Spinocerebellar projection inthe meander tail mutant mouse: organization in the granular posteriorlobe and the agranular anterior lobe. Brain Res. 558:149–152.
Gravel, C., and R. Hawkes (1990) Parasagittal organization of the ratcerebellar cortex: direct comparison of Purkinje cell compartments andthe organization of the spinocerebellar projection. J. Comp. Neurol.291:79–102.
Gravel, C., L.M. Eisenman, R. Sasseville, and R. Hawkes (1987) Parasagit-tal organization of the rat cerebellar cortex: direct correlation betweenantigenic Purkinje cells bands realed by mabQ113 and the organizationof the olivocerebellar projection. J. Comp. Neurol. 265:294–310.
Grishkat, H.L., and L.M. Eisenman (1994)Anterograde labeling of spinocer-ebellar fibers in fetal mouse cerebellum. Soc. Neurosci. Abstr. 20:1749.
Hamilton, B.A.,W.N. Frankel,A.W. Kerrebrock, T.L. Hawkins,W. FitzHugh,K. Kusumi, L.B. Russell, K.L. Mueller, V.v. Berkel, B.W. Birren, L.Kruglyak, and E.S. Lander (1996) Disruption of the nuclear hormonereceptor RORa in staggerermice. Nature 379:736–739.
Hamre, K.M., and D. Goldowitz (1995). The effects of the murine mutation,meander tail, on cerebellar Purkinje and Golgi epithelial cells aresecondary to the loss of granule cells. Soc. Neurosci. Abstr. 21:1042.
Hawkes, R., N. Leclerc, G.A. Schwarting, K. Herrup, and M. Yamamoto(1992) Compartmentation in mammalian cerebellum: Zebrin II andP-path antibodies define three classes of sagittally organized bands ofPurkinje cells. Proc. Natl. Acad. Sci. U.S.A. 89:5006–5010.
Hawkes, R., S. Blyth, V. Chockkan, D. Tano, Z. Ji, and C. Mascher (1993)Structural and molecular compartmentation in the cerebellum. Can. J.Neurol. Sci. 20(Suppl. 3):S29–S35.
Herrup, K. (1983) Role of staggerer gene in determining cell number incerebellar cortex. I. Granule cell death is an indirect consequence ofstaggerer gene action. Dev. Brain Res. 11:267–274.
Herrup, K., and R.J. Mullen (1979a) Regional variation and absence oflarge neurons in the cerebellum of the staggerer mouse. Brain Res.172:1–12.
Herrup, K., and R.J. Mullen (1979b) Staggerer chimeras: intrinsic nature ofPurkinje cell defects and implications for normal cerebellar develop-ment. Brain Res. 178:443–457.
Herrup, K., and R.J. Mullen (1981) Role of the staggerer gene in determin-ing Purkinje cell number in the cerebellar cortex of mouse chimeras.Dev. Brain Res. 1:475–485.
Ito, M. (1984) The Cerebellum and Neural Control. NewYork: Raven Press.Ji, Z., and R. Hawkes (1994) Topography of Purkinje cell compartments and
mossy fiber terminal fields in lobules II and III of the rat cerebellar
cortex: spinocerebellar and cuneocerebellar projections. Neuroscience61:935–954.
Ji, Z., and R. Hawkes (1995) Developing mossy fiber terminal fields in therat cerebellar cortexmay segregate because of Purkinje cell comparmen-tation and not competition. J. Comp. Neurol. 359:197–212.
Ji, J., and R. Hawkes (1996) Partial ablation of the neonatal externalgranular layer disrupts mossy fiber topography in adult the ratcerebellum. J. Comp. Neurol. 371:578–588.
Landis, D.M.D., and R.L. Sidman (1978) Electron microscopic analysis ofpostnatal histogenesis in the cerebellar cortex of staggerer mutantmice. J. Comp. Neuron. 179:831–864.
Leclerc, N., C. Gravel, and R. Hawkes (1988) Development of parasagittalzonation in the rat cerebellar cortex: MabQ113 antigenic bands arecreated postnatally by the suppression of antigen expression in a subsetof Purkinje cells. J. Comp. Neurol. 273:399–420.
Maeda, N., M. Niinobe, Y. Inoue, and K. Mikoshiba (1989) Developmentalexpression and intercellular location of P400 protein characteristic ofPurkinje cells in the mouse cerebellum. Dev. Biol. 133:67–76.
Mariani, J., and J.P. Changeux (1980) Multiple innervation of Purkinjecells by climbing fibers in the cerebellum of the adult staggerer mutantmouse. J. Neurobiol. 11:41–50.
Mason, C.A., and E. Gregory (1984) Postnatal maturation of cerebellarmossy and climbing fibers: transient expression of dual features onsingle axons. J. Neurosci. 4:1715–1735.
Matsushita, M., and H. Yaginuma (1989) Spinocerebellar projections fromspinal border cells in the cat as studied by anterograde transport ofwheat germ agglutinin-horseradish peroxidase. J. Comp. Neurol. 288:19–38.
Messer, A. (1988) Thyroxine injections do not cause premature induction ofthymidine kinase in sg/sgmice. J. Neurochem. 51:888–891.
Messer, A., J. Plummer-Siegard, and B. Eisenberg (1990) Staggerer mutantmouse Purkinje cells do not contain detectable calmodulin mRNA. J.Neurochem. 55:293–302.
Messer, A., B. Eisenberg, and J. Plummer (1991) The lurcher cerebellarmutant phenotype is not expressed on a staggerer mutant background.J. Neurosci. 11:2295–2302.
Oberdick, J. (1994) Evidence for a genetically encoded map of functionaldevelopment in the cerebellum. Histochemistry 102:1–14.
Paradies, M.A., and L.M. Eisenman (1993) Evidence of early topographicorganization in the embryonic olivocerebellar projection: a modelsystem for the study of pattern formation processes in the centralnervous system. Dev. Dyn. 197:125–145.
Sidman, R.L., P.W. Lane, and M.M. Dickie (1962) Staggerer: a newmutation in the mouse affecting the cerebellum. Science 137:610–612.
Slemmon, J.R., D. Goldowitz, R. Blacher, and J.I. Morgan (1988) Evidencefor the transneuronal regulation of cerebellin biosynthesis in develop-ing Purkinje cells. J. Neurosci. 8:4603–4611.
Smeyne, R.J., T. Chu,A. Lewin, F. Bian, S.S. Crisman, C. Kunsch, S.A. Lira,and J. Oberdick (1995) Local control of granule cell generation bycerebellar Purkinje cells. Mech. Cell. Neurosci. 6:230–251.
Sotelo, C., andM.Wassef (1991) Cerebellar development: afferent organiza-tion and Purkinje cell heterogeneity. Philos. Trans. R. Soc. Lond. [Biol.]331:307–313.
Sotelo, C., F. Bourrat, and A. Triller (1984) Postnatal development of theinferior olivary complex in the rat. II. Topographic organization of theimmature olivocerebellar projections. J. Comp. Neurol. 222:177–199.
Tolbert, D.L., T. Pittman, J.M.Alisky, and B.R. Clark (1994) Chronic NMDAreceptor blockade or muscimol inhibition of cerebellar cortical neuronalactivity alters the development of spinocerebellar afferent topography.Dev. Brain Res. 80:268–274.
Vogel, M.W., and J. Prittie (1994) Topographic spinocerebellar mossy fiberprojections are maintained in the Lurcher mutant. J. Comp. Neurol.343:341–351.
Vogel, M.W., J. Soha, J. Prittie, and J.E. Crandall (1993)Analysis of parallelfiber elongation in the wild type and mutant mouse cerebellum. Soc.Neurosci. Abstr. 19:1089.
Wassef, M., J.P. Zanetta, A. Brehier, and C. Sotelo (1985) Transientbiochemical compartmentalization of Purkinje cells during early cerebel-lar development. Dev. Biol. 111:129–137.
Yoon, C.H. (1972) Developmental mechanism for changes in cerebellum of‘‘staggerer’’ mouse, a neurological mutant of genetic origin. Neurology22:743–754.
Zanjani, H.S., J. Mariani, and K. Herrup (1990) Cell loss in the inferiorolive of the staggerer mutant mouse is an indirect effect of the gene. J.Neurogenet. 6:229–241.
362 Z. JI ET AL.