Embed Size (px)
Citation preview

Vienna KuhnDr. Marshall-Walker
11/15/15Biology 600/1
Evaluating effects of early- and late-onset rasgap-2 mutations on C. elegans’
memory
Abstract
SynGAP, a newly identified rare neurological disease, is one of many new
diseases that have been identified thanks to genomic sequencing of undiagnosed disease
cases. With the disease’s many ties to autism and epilepsy, the mutation in the
SYNGAP1 gene could be more prevalent than previously expected. The mutation, like
many others that cause intellectual disability, affects neurons and their synaptic
transmissions. The exact connection of this mutation to the symptoms of intellectual
disability, such as short-term memory impairment, are as of yet unknown, but a homolog
to the SynGAP gene that has been identified in C. elegans may provide some clues. An
experiment testing worms that have had the gene eliminated will seek to confirm whether
these worms have impaired short-term memory in comparison to worms that have a
functioning copy of the gene. This information might be extrapolated to how the gene
mutation in humans may be, on a molecular level, causing the physical and mental
symptoms that manifest themselves in patients.
Introduction
According to Global Genes, there are 7,000 rare diseases in the world, and only
5% of these have even one FDA-approved drug treatment.1 Considering the fact that
these diseases collectively affect more than 300 million individuals, it is imperative that
research on rare diseases becomes a priority so that they too have treatment options.1
Contributions to a rare disease may individually only assist a handful of patients, but
collectively will have a massive impact.
The advent of genome and full exome sequencing has demystified many
undiagnosed cases of disease; many can now be attributed to a mutation in a specific
gene. New diseases are being identified as more individuals with mutations are
discovered, and new patient communities are formed every day. One of the biggest
impediments to research conducted on these cases of rare disease is the oft-high

variability of factors within a patient community, such as severity of symptoms and age
of onset. Even basic details such as the range of phenotypic effects of a genetic mutation
on the body could be different for each patient and are only beginning to be explored on
the cellular level.
One such mutation is present in the SYNGAP1 gene in some individuals, causing
a disease called SynGAP. SynGAP is typically caused by a de novo mutation, one that
occurs spontaneously and randomly during gamete formation or meiosis, which results in
the truncation of the SYNGAP1 gene, and subsequently haploinsufficiency within the
organism.2 Haploinsufficiency means that the mutated copy of the gene cannot produce a
functioning version of the protein that corresponds to the gene.3 A single complete copy
of the gene is not sufficient to maintain the level of protein production needed for a
healthy organism, causing disability.3
A properly functioning SynGAP protein is required for the negative regulation
(limitation) of Ras protein activity at silent (excitatory) synapses.4 Ras proteins are
responsible for delivering messages about growth rates and nutrient levels between cells,
using the transformation of GDP to GTP to strengthen messages.5 Silent synapses are
called such because they do not trigger a synaptic current when the postsynaptic neuron is
at rest.6 This type of synapse is especially predominant at the beginning of postnatal
development, but as the organism progresses, it is essential that SynGAP negatively
regulate these neurons so that other types of long-term cognitive neural currents can be
developed.2, 6 SynGAP, in concert with Unc51.1, is also essential for monitoring the
growth of axons during postnatal development to negatively regulate when axons should
complete the growth process. 7
The loss of function of the SynGAP protein due to a mutation has drastic effects
on both the molecular and organismal levels of an individual. Research has shown that
mutations in a SYNGAP1 mouse model caused the premature development of dendritic
spine synapses, which was then linked to the manifestation of developmental
irregularities associated with conditions such as intellectual disability and autism
spectrum disorder.2 Both of the aforementioned conditions are ones for which SynGAP
mutations are often misdiagnosed, indicating the possibility that the protein is tied to
intellectual development in some way.8

As of yet, there is a small community of 32 known patients with the SynGAP
mutation that span a range of severity of intellectual disability, autism spectrum
disorders, and epilepsy.8 This small community of those with the mutation, however,
could be on track to expand rapidly, as one scientific paper indicates that early prevalence
data suggest the frequency of this mutation could be greater than Fragile X Syndrome and
could already affect over one million individuals. While there are many similarities in
symptoms between the existing patients of SynGAP that could indicate an identifiable
disease, the gene itself is implicated in a number of other intellectual disability-associated
conditions, disorders that fall under the autism spectrum umbrella, as well as epilepsy.2, 9
While SYNGAP1 is known for its molecular association with neural synapses and
organismal association with intellectual disability, the precise outcome of the increased
neural excitement and premature axon growth that subsequently causes intellectual
disability is as of yet unknown. A paper published in October 2015 sought to research
this by observing genes similar to those in the SYNGAP family in Caenorhabditis
elegans nematodes.2, 10 The study revealed the specific effects of genes in the rasgap
family by testing dsRNA knockdown strains for genes within the rasgap family on short-
and long-term memory and olfactory learning.10 In order to do this, bacteria -- producing
dsRNA coding for the gene to be knocked down -- was fed to the nematodes. This
activated a process known as RNA interference, in which the cell, triggered by foreign
copies of double-stranded RNA, activates a process that will successfully thwart all
attempts to produce a protein from that RNA.11 (See Figure 1) The process does not
discriminate between native dsRNA and foreign dsRNA, so it is helpful in experiments to
observe the effects of completely eliminating a gene from an organism.12
Of note is rasgap-2, a homolog (a similar gene in a different organism) of human
SYNGAP1. The study determined that gap-2 was essential for short- and long-term
associative memory.10 Continuing to research rasgap-2 mutants in C. elegans will provide
a small-scale model for preliminary research on SynGAP in higher eukaryotes.
C. elegans nematodes are regularly used in biological research for many reasons.
They are small organisms, made up of only around 1000 somatic cells at full maturity,
which makes them easy to manipulate.13 Their life cycle lasts approximately three days,
so they are rapidly accessible as an experimental model.14 Only around 55 hours are

needed for the nematodes to reach full maturity and to gain the ability to reproduce.15 The
nematodes are also hermaphroditic and each can produce upwards of 300 offspring, so
they are ideal for large-scale experiments.12 What makes C. elegans most applicable to
genetic experiments, however, is that its entire genome sequence is known and many
homologs for genes found in humans have been located.13 One such example was the
aforementioned rasgap-2, which when eliminated, could have important implications for
its homolog’s purpose and effect in a human model.
The previously referenced study focused on a single time period (L3) within the
nematodes’ lifespan at which to begin feeding the worms on a dsRNA-transfected
bacteria strain in order to knock down the rasgap-2 gene (See Figure 2 for C. elegans life
cycle).10 While this created a standard platform from which to compare memory and
olfactory learning experiments across the entirety of the rasgap gene family, the study left
questions unanswered about whether the results of the experiment would have improved
or deteriorated based on if the gene had been knocked down using RNA interference at
earlier and later intervals in the nematode’s lifespan.
This experiment seeks to address those questions and determine the implications
the results might have for the effects of different stages of onset (during postnatal
development) for SynGAP mutations in humans. Three RNAi-sensitive strains of C.
elegans will be used – one as a control, fed on unaltered bacteria, one fed from egg
development on rasgap-2 knockdown dsRNA bacteria, and one fed from young
adulthood on the same strain of rasgap-2 knockdown dsRNA bacteria. An identical
short-term associative memory assay to the assay in the aforementioned paper will be
used to evaluate whether the change in time frame of rasgap knockdown through
feeding will have an effect on the severity of the short-term memory impairment.
Materials and Methods
The reagents were obtained from Sigma-Aldrich in St. Louis, MO, unless otherwise
indicated.
I. Worm Protocol
Both control and RNAi worms were of the eri-(mg366);lin-15b(n744) knockdown C.
elegans strain. This strain was used for RNAi because it enhances neuron sensitivity to

RNAi.16 The nematodes were incubated at 17 degrees Celsius and were maintained
following standard worm protocol. They were grown on standard 6cm NGM plates of
separate formulas: the control worms on worm plates based on standard protocol, and
RNAi on worm plates whose formula is specifically designed for its support of RNAi.
II. Bacteria
Rasgap-2 dsRNA-expressing bacterial strain (generous gift from Dr. Attila Stetak,
Semmelweis University, Budapest, Hungary) was seeded onto light-sensitive RNAi
plates and fed to a portion of worms. Control worms, also of the eri-lin strain, were
raised on OP50 bacteria that previously existed in the laboratory.
III. RNA Interference (RNAi)
Nematodes of the eri-lin strain from a wide range of locations in their lifespan were
transferred onto light-sensitive RNAi formula plates (NaCl, Bactotryptone, agar, 5
mg/mL cholesterol, 100 mg/mL Ampicillin, 1M IPTG) seeded with an HT115(DE3) E.
coli strain that produces dsRNA against rasgap-2. Plates were incubated at 17 degrees
Celsius.
IV. C. Elegans Memory Assay – OP50-fed worms
Worms were washed from plate containing OP50 using approximately 1500 µl of M9
buffer, which was then transferred to an Erlenmeyer flask. They were centrifuged at 1
RPM for two minutes. M9 was removed down to approximately 50µl and then new M9
was added. The worms were spun twice, and after the second spin, M9 was again
removed to approximately the 50µl mark. This time, the 50µl was transferred to a CTX
plate (standard 6cm worm plate but made up of KH2PO4, K2HPO4, CaCl2, MgSO4, and
agar in distilled water), about 2 centimeters from the edge of the plate. On the lid of
the plate, a 10 µl drop of 1:100 diluted diacetyl was placed and the plate was then
closed and parafilmed. This served as the nematodes’ pre-exposure to diacetyl and
association with starvation. After an hour, the worms were rinsed from the plate using
M9 buffer and again spun down, but only once. They were then moved to the center of
a second CTX plate, which had 10 µl spots of both attractant (diacetyl) and a counter-
spot (200-proof ethanol) 1 cm from opposing ends of the plate. Worms were left at
room temperature for 4 hours and 45 minutes and then counted using a Leica 2000
microscope based on their proximity (within 1 cm) to the attractant and counter-spot.

V. C. Elegans Memory Assay – RNAi-fed worms
This experiment followed the same procedure as the OP50 experiment, but the worms
remained on the second CTX plate approximately 3 hours for counting. One day after
initial transfer of worms from OP50 to RNAi bacteria, all adult and L4 worms were
removed so that the plate only contained worms that would reach maturity on the
RNAi bacteria strain (See Figure 2 for details on C. elegans life span).
Results
For the OP50-fed nematode memory assay outlined within Materials and Methods, 81
worms were counted in the attractant region, 50 in the counter-attractant region, and
161 not in either of the other regions (see Figure 3 for table). The RNAi bacteria-fed
nematode memory assay resulted in 411 worms in the attractant region, 173 in the
counter-attractant region, and an indeterminable amount not in the region due to the
high volume of worms (see Figure 4 for table). As seen by the difference in number of
worms in both assays, the ratio of worms that preferred the attractant was considerably
higher for those that had the rasgap-2 gene knocked down through RNA interference
(See Figure 5 for graph).
Discussion
The original intention of this experiment was to see whether altering the time frame of
when nematodes were exposed to the RNAi process would change the degree of
aversion or attraction to the attractant, namely diacetyl. This would be done by testing
a control fed on OP50, a strain fed on rasgap-2 knockdown dsRNA bacteria from egg
development, and a strain fed from young adulthood on the same strain of rasgap-2
knockdown dsRNA bacteria. This did not prove feasible because I was unable to
collect a sufficient number of individual worms of the right age from a plate on which
to conduct an experiment. Instead, I decided to place worms at various stages of life
onto RNAi-inducing bacteria and let the worms reproduce freely. I could guarantee
they had all had rasgap-2 eliminated by waiting a span of 11 days, at which point
multiple generations had fed on the RNAi bacteria. What I found aligned with the
results of a previous experiment conducted by individuals at Semmelweis University

in Hungary. When RNAi was used to knock down rasgap-2 in C. elegans nematodes,
the hour of pre-conditioning that would associate diacetyl with starvation prompted an
aversion to the previously palatable attractant in OP50 control worms when they were
moved to the second test plate. As evidenced by their graph, which compares OP50 to
gap-2 knockdowns, the gap-2 knockdowns experienced a lower rate of aversion (See
Figure 5). Similarly, in this experiment, while OP50 worms still showed an attraction
to diacetyl after conditioning, the difference between worms in the attractant region
and worms in the counterspot region was significantly lower than the difference for
RNAi-manipulated worms. The larger difference for RNAi-manipulated worms means
less worms retained the negative conditioning that occurred before the experiment. If
this is the case, the results suggest that gap-2, when knocked down, does in fact impair
the worm’s ability to retain short-term memory.
If this conclusion can be drawn, the experiment opens up many more avenues
of questioning and the possibility of extending the results to human patients. The more
that is known about the human gene and its homologs in other organisms, the more
likely it is that similar diseases can be identified, locating possible treatments for the
amelioration of existing symptoms and hopefully an eventual cure.

Appendix
Figure 1: RNA Interference
The natural process of the cell to silence genes is taken advantage of using non-native dsRNA.17

Figure 2: C. elegans Life Cycle
Life cycle of a C. elegans nematode from egg to adult. Worms become fertile at approximately 55 days.18

Figure 3: OP50-fed Memory Assay Results
Worms in Attractant
RegionNot in Either Region
Worms in Counter-attractant
Region Total
81 161 50 292
Worms were counted within a 2x1 cm rectangle around point where attractant and
counter-attractant were added to the plate. Many worms did not move from their point of
origin. Bacteria remained on the CTX plate that was transferred from the original NGM
plate.

Figure 4: RNAi-fed Memory Assay Results
Worms in Attractant
RegionNot in Either Region
Worms in Counter-attractant
Region Total
411 indeterminate 173 unknown
Worms were counted within a 2x1 cm rectangle around point where attractant and
counter-attractant were added to the plate. Again, many worms did not move from their
point of origin. Based on the density of worms, I surmised there were upwards of 1000 on
the test plate. Bacteria also remained on this CTX plate, again transferred from the
original NGM plate.

Figure 5: Comparison of worm counts in regions of attractant and counterspot for different feeding groups of worms
0
50
100
150
200
250
300
350
400
450
Control versus RNAi gap-2 worms: at-traction to diacetyl
OP50-fed wormsRNAi-fed worms
Worms in attractant region Worms in counterspot region
Num
ber
of w
orm
s
OP50 experimental size was significantly smaller, which accommodates for the initial
difference between OP50 and RNAi experiments.
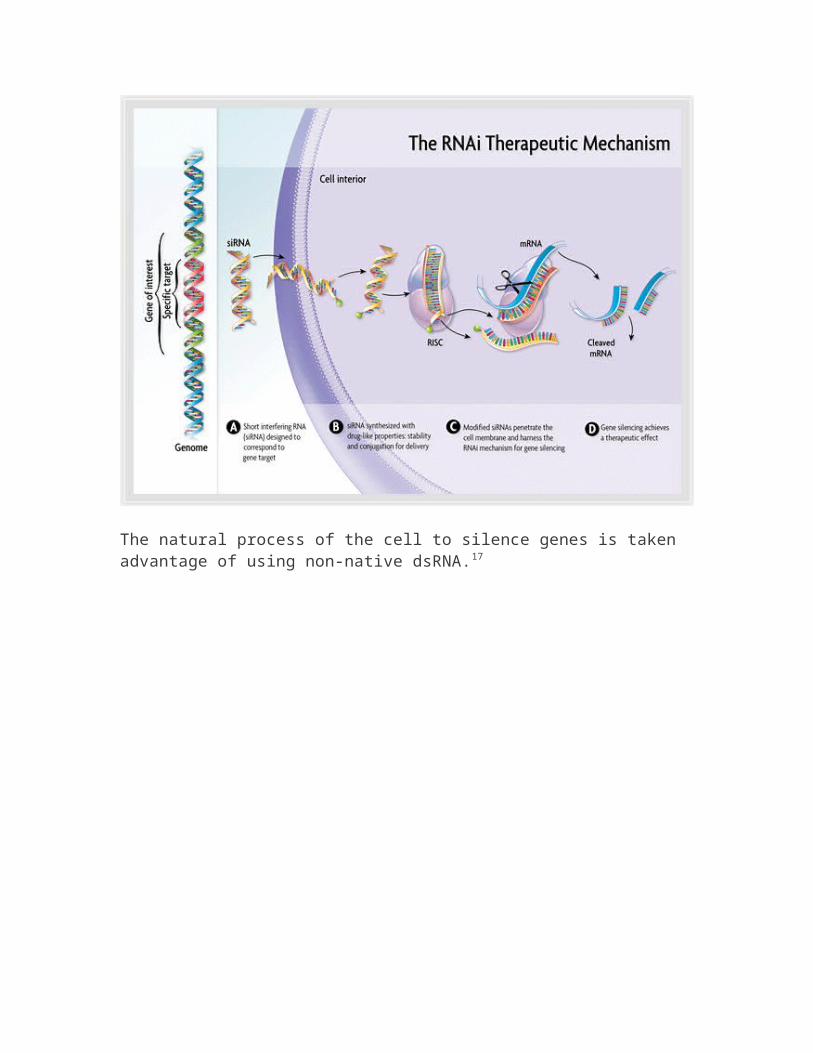
Figure 6: Graph of results of RNAi on memory (from outside experiment)
“Negative conditioning assays were combined with RNA interference experiments
using eri-1(mg366);lin-15B(n744) RNA sensitized worms, fed against (A) gap-1 (n = 3),
(B) gap-2 (n = 13, p = 2.72 × 10−7), and (C) gap-3 (n = 9, p = 3.75 × 10−5) dsRNA
carrying bacteria. Dark grey represents the same strain fed with bacteria carrying an
empty GFP marker dsRNA as reference. N: naïve, C: conditioned, R: recovered animals
(see Materials and Methods for details). Error bars indicate SD and asterisks indicate
significant differences (***P < 0.001).”10

1References RARE diseases: Facts and statistics [Fact sheet]. (n.d.). Retrieved from
https://globalgenes.org/rare-diseases-facts-statistics/
2 Clement, J. P., Aceti, M., Creson, T. K., Ozkan, E. D., Shi, Y., Reish, N. J., . . . Rumbaugh,
G. (2012). Pathogenic SYNGAP1 mutations impair cognitive development by disrupting
maturation of dendritic spine synapses [PDF]. Cell, (151), 709-723.
http://dx.doi.org/10.1016/j.cell.2012.08.045
3 Ibid.4 Kim, J. H., Liao, D., Lau, L.-F., & Huganir, R. L. (1998). SynGAP: a synaptic rasGAP that
associates with the PSD-95/SAP90 protein family [PDF]. Neuron, 20, 683-691.
http://dx.doi.org/10.1016/S0896-6273(00)81008-9
5 Goodsell, D. (2012, April). Ras protein. Retrieved from RCSB Protein Data Bank database.
(Accession No. 148)
6 Feldman, D. E. (n.d.). Activity-dependent maturation of glutamate receptor content. In L. R.
Squire (Ed.), Encyclopedia of neuroscience. Retrieved from http://ScienceDirect
7 Jankowski, S. A. (2013, September 24). Synaptic RAS-GTPase-activating protein 1;
SYNGAP1 (C. L. Kniffin, A. Hamosh, P. A. Hartz, & P. J. Converse, Ed.) [Fact sheet].
Retrieved from Online Mendelian Inheritance of Man database. (Accession No. 603384)
8 Unpublished research, July 2015.9 Berryer, M. H., Hamdan, F. F., Klitten, L. L., Møller, R. S., Carmant, L., Schwartzentruber,
J., . . . Di Cristo, G. (2013). Mutations in SYNGAP1 cause intellectual disability, autism,
and a specific form of epilepsy by inducing haploinsufficiency. Human Mutation, 34(2),
385-394. http://dx.doi.org/10.1002/humu.22248

10 Gyurkó, M. D., Csermely, P., S?ti, C., & Steták, A. (2015). Distinct roles of the RasGAP
family proteins in C. elegans associative learning and memory. Nature, (5).
http://dx.doi.org/10.1038/srep15084
11 Timmons, L., & Fire, A. (1998). Specific interference by ingested dsRNA. Nature, 395, 854.
12 Lau, N. C., & Bartel, D. P. (2003, August). Censors of the genome. Scientific American, 34-
41.
13 Alberts B, Johnson A, Lewis J, et al. Molecular Biology of the Cell. 4th edition. New York:
Garland Science; 2002. Caenorhabditis Elegans: Development from the Perspective of the
Individual Cell. Available from: http://www.ncbi.nlm.nih.gov/books/NBK26861/
14 Toxicol. Sci. (2008) 106 (1): 5-28.doi: 10.1093/toxsci/kfn121
15 Introduction to C. elegans anatomy. (n.d.). Retrieved from Worm Atlas website:
http://www.wormatlas.org/ver1/handbook/anatomyintro/anatomyintro.htm
16 http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3820814/
17 RNA interference. (n.d.). Retrieved from Alnylam Pharmaceuticals website:
http://www.alnylam.com/rnai_primer/rna-interference-pg5.htm
18 Life cycle [Photograph]. (n.d.). Retrieved from http://www.wormatlas.org/ver1/
handbook/anatomyintro/anatomyintro.htm





























