Embed Size (px)
Citation preview

Biom. Hum. et Anthropol. 2009, 27, 1-2, p. 0-0. Dambricourt A., Etude au scanner des canaux semi-circulaires d’un anencéphale…
ETUDE AU SCANNER DES CANAUX SEMI-CIRCULAIRES D’UN A NENCEPHALE DEPOURVU DE FLEXION SPHENO-OCCIPITALE ; IMPLICATION S POUR L’INTERPRETATION PALEO-EMBRYOLOGIQUE DES HOMINIDES FOSSILES.
SEMI-CIRCULAR CANALS CT SCAN STUDY OF AN ANENCEPHAL IC HUMAN FETUS DEVOID OF BASICRANIAL FLEXURE, IMPLICATIONS FOR PAL AEO-EMBRYOLOGICAL INTERPRETATION OF THE FOSSIL HOMINIDS .
ANNE DAMBRICOURT MALASSÉ (1), F ABIENNE LALLOUET (2)
RÉSUMÉ
La différenciation des canaux semi-circulaires est contrôlée par les gènes homéobox [MERLO et al. 2002], cependant certains angles spécifiques au genre Homo seraient liés à l’amplitude de la dynamique de flexion de la base [DELATTRE et FENART, 1960], comme l’importante ouverture angulaire entre le canal semicirculaire latéral et le canal semicirculaire postérieur spécifique au genre Homo [SPOOR et ZONNEVELD, 1994, JEFFERY et SPOOR, 2004]. L’étude biométrique d’un crâne anencéphale dépourvu de flexion basicrânienne, âgé de 5 mois i.u. comparé à un fœtus de gorille et un fœtus humain normal de même âge, montre pour la première fois, l’indépendance entre la formation de l’angle et dynamique de flexion basi-crânienne. L’interprétation de l’origine des différentes postures humaines se déplace vers des considérations paléoembryogénétiques et saltationnistes (équilibres ponctués), au lieu d’une résultante d’adaptations graduelles à un long réchauffement climatique. L’hypothèse d’une évolution de la combinatoire des gènes à homéobox se confirmerait [colloque inter-académique de l’Institut de France, DAMBRICOURT MALASSÉ, 2006a].
Mots clés : canaux semicirculaires, angle sphénoïdal, bipédie permanente, embryogenèse, gènes à homéobox, équilibres ponctués.
ABSTRACT The application of the digital medical imaging in human palaeontology allows a new exploration of the basicranial
tissues. The differentiation of semi-circular canals is under the control of homeobox genes [MERLO et al. 2002] however certain specific angles of Homo genus would be connected to the amplitude of the dynamics flexion of the base [DELATTRE et FENART, 1960, JEFFERY et SPOOR, 2004]. The comparison of an anencephalic human fetus devoid of basicranial flexure, with a normal human foetus and a gorilla foetus, 5-month-old i.u., shows that the angular opening between the lateral semicircular and the posterior semicircular canals, specifically human [SPOOR et ZONNEVELD, 1994], is independent from the basi-cranial flexion and precedes it. The first human postures would result from an punctuated evolution of combinatorial genes expression underlying the rostro-caudal embryonic differentiation as previously proposed [Inter-academic conferences of the Institut de France, DAMBRICOURT MALASSÉ, 2006, DAMBRICOURT MALASSÉ, 1988] and not from gradual adaptations to climate changes.
Key words: semicircular canals, sphenoidal angle, permanent bipedalism, embryogenesis, homeobox genes, punctuated equilibria.
INTRODUCTION
La paléontologie humaine connaît un nouvel essor depuis l’apparition de la nouvelle imagerie médicale numérique, notamment dans l’exploration des structures osseuses basicrâniennes parmi les plus délicates d’accès et les plus complexes à décrire, telle que l’oreille interne. L’imagerie conventionnelle de la téléradiographie en limitait l’approche [DELATTRE et FENART, 1960], cependant le corpus de connaissances acquis grâce aux échanges entre l’étude téléradiographique des déséquilibres occluso-posturaux contemporains, d’une part, et la compréhension de la statique occluso-posturale des hominidés fossiles, d’autre part, a permis de progresser dans la compréhension de la morphogenèse basi-cranio-faciale
UMR 7194 CNRS- Département de Préhistoire du Muséum national d’Histoire naturelle, Institut de Paléontologie Humaine, 1, rue René Panhard, 75013, Paris.
Groupe Hospitalier Diaconesses-La Croix Saint Simon, Service d’Imagerie Médicale, Paris.
Article reçu le 00.00.2008, accepté le 00.00.2009
Biom. Hum. et Anthropol. 2009, 27, 1-2, p. 0-0. Dambricourt A., Etude au scanner des canaux semi-circulaires d’un anencéphale…
[GUDIN et GODARD, 1973, DESHAYES et DAMBRICOURT MALASSÉ, 1991]. L’application de ces nouvelles technologies aux hominidés fossiles, tel le scanner et la reconstitution tridimensionnelle, s’inscrit dans la continuité de ces acquisitions [ZONNEVELD et WIND, 1985, ZONNEVELD et SPOOR, 1997, CONROY et al. 1990], elle s’avère d’autant plus nécessaire que cette région anatomique constitue une importante source d’informations sur les postures bipèdes permanentes apparues avec les Australopithécinés (Australopithecus, Paranthropus) et les plus anciennes espèces du genre Homo.
LA PROBLEMATIQUE
Le paradigme en vigueur pour expliquer l’origine de la verticalité axiale spécifique aux Australopithécinés et au genre Homo (du post-sphénoïde au sacrum) est hypothético-déductif. Il suppose une acquisition lente, graduelle de petites modifications anatomiques isolées du reste de l’organisme, parvenant aux postures érigées permanentes. Il en découle ce qui aurait du être sélectionné graduellement pendant 7 millions d’années pour aboutir à l’unité anatomophysiologique de l’espèce sapiens, rien de moins que l’identité évolutive du genre Homo [DAMBRICOURT MALASSÉ, 2009a,b]. Selon cette démarche hypothético-déductive, la posture érigée résulterait d’une succession d’adaptations locomotrices sélectionnées en raison d’un long réchauffement climatique produisant un buissonnement d’espèces anthropoïdes, sans qu’il soit possible de distinguer une filiation entre les fossiles exhumés [HARCOURT-SMITH et AIELLO, 2004]. L’organisme se serait redressé en abandonnant progressivement l’arboricolisme. La majorité des paléontologues s’inscrivent dans le paradigme gradualiste, cependant bon nombre d’entre eux ont abandonné l’hypothèse paléo-environnementale d’une déforestation, en Afrique du Sud comme en Afrique de l’Est, avec Mac Kee [1999], Tobias : «The evidence for this change came from fossil plants and animals that were contemporaries of the early hominids. The Savanna Hypothesis was no longer tenable.» [TOBIAS, 2004] où les milieux naturels se sont avérés humides et aussi diversifiés et que les biotopes des grands anthropoïdes miocènes: « le milieu d’Orrorin, vieux de 6 millions d’années s’avère bien boisé […]. La découverte a obligé les paléontologues à modifier leurs idées sur le tempo des événements évolutifs et le type d’environnement dans lequel la transition a eu lieu » [SENUT et PICKFORD, 2005]. Les espèces d’Australopithécinés étaient donc arboricoles [DELOISON, 1999] alors que leur squelette axial était déjà verticalisé à la naissance [DAMBRICOURT MALASSÉ, 1988, 1993]. La plus ancienne trace fossile de cette verticalité axiale ne date pas plus de 4,4 M.a avec un fragment isolé du basicrâne (basi-occipital et rocher) découvert en milieu forestier [WOLDGABRIEL et al. 1994]. L’espèce était en outre sympatrique de grands anthropoïdes dont la denture récoltée à un kilomètre de distance, correspond au type d’occlusion des Paninés actuels [WHITE et al. 1994]. Or les recherches en orthopédie maxillo-faciale appliquées à la croissance des trois genres Pan, Gorilla et Pongo ont montré le caractère univoque de leur équilibre occluso-postural irréductible à tout équilibre craniosacral verticalisé [DESHAYES, 1991, DESHAYES et DAMBRICOURT MALASSÉ, 1990, DAMBRICOURT MALASSÉ, 2006]. L’acquisition de la verticalité axiale n’a nécessité ni l’abandon d’une locomotion arboricole ni remplacé un mode de locomotion inadapté à son environnement.
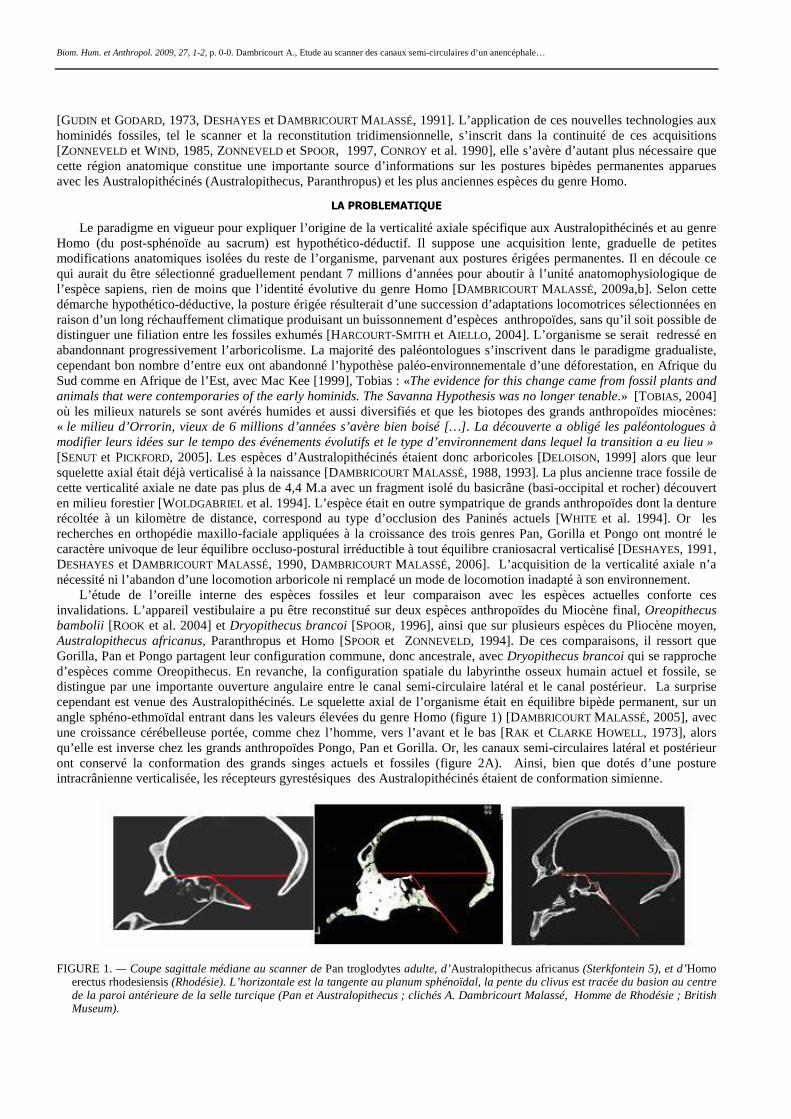
L’étude de l’oreille interne des espèces fossiles et leur comparaison avec les espèces actuelles conforte ces invalidations. L’appareil vestibulaire a pu être reconstitué sur deux espèces anthropoïdes du Miocène final, Oreopithecus bambolii [ROOK et al. 2004] et Dryopithecus brancoi [SPOOR, 1996], ainsi que sur plusieurs espèces du Pliocène moyen, Australopithecus africanus, Paranthropus et Homo [SPOOR et ZONNEVELD, 1994]. De ces comparaisons, il ressort que Gorilla, Pan et Pongo partagent leur configuration commune, donc ancestrale, avec Dryopithecus brancoi qui se rapproche d’espèces comme Oreopithecus. En revanche, la configuration spatiale du labyrinthe osseux humain actuel et fossile, se distingue par une importante ouverture angulaire entre le canal semi-circulaire latéral et le canal postérieur. La surprise cependant est venue des Australopithécinés. Le squelette axial de l’organisme était en équilibre bipède permanent, sur un angle sphéno-ethmoïdal entrant dans les valeurs élevées du genre Homo (figure 1) [DAMBRICOURT MALASSÉ, 2005], avec une croissance cérébelleuse portée, comme chez l’homme, vers l’avant et le bas [RAK et CLARKE HOWELL, 1973], alors qu’elle est inverse chez les grands anthropoïdes Pongo, Pan et Gorilla. Or, les canaux semi-circulaires latéral et postérieur ont conservé la conformation des grands singes actuels et fossiles (figure 2A). Ainsi, bien que dotés d’une posture intracrânienne verticalisée, les récepteurs gyrestésiques des Australopithécinés étaient de conformation simienne.
FIGURE 1. — Coupe sagittale médiane au scanner de Pan troglodytes adulte, d’Australopithecus africanus (Sterkfontein 5), et d’Homo
erectus rhodesiensis (Rhodésie). L’horizontale est la tangente au planum sphénoïdal, la pente du clivus est tracée du basion au centre de la paroi antérieure de la selle turcique (Pan et Australopithecus ; clichés A. Dambricourt Malassé, Homme de Rhodésie ; British Museum).

Biom. Hum. et Anthropol. 2009, 27, 1-2, p. 0-0. Dambricourt A., Etude au scanner des canaux semi-circulaires d’un anencéphale…
FIGURE 2A. — schéma de l’appareil vestibulaire en vue supérieure de Pan, Homo et Australopithecus, montrant la communauté angulaire du Paniné et de l’Australopithéciné et l’acquisition chez l’homme d’un angle plus ouvert entre le canal postérieur et le canal latéral (d’après SPOOR et al. 1994).
Le labyrinthe humain se démarque donc de toute espèce de primate quel que soit leur équilibre bipède au sol,
occasionnel ou exclusif (les gibbons par exemple). L’acquisition du contrôle de l’équilibre humain, ou l’équilibre humain proprement dit, n’a donc pas été graduelle, quant à l’acquisition de l’équilibre bipède permanent en soi (tous les homininés), elle ne fut pas la substitution de l’arboricolisme par suite d’une lente adaptation à un réchauffement climatique. L’observation des origines embryonnaires de la posture avait déjà permis de le constater ; pour l’homme comme pour les Australopithécinés, l’acquisition de la verticalité axiale est de nature saltationniste, c’est un équilibre ponctué qui concerne l’organogenèse céphalo-caudale de l’embryon (absence de transformation morphogénétique entre deux longues périodes géologiques) [DAMBRICOURT MALASSÉ, 1988, 1993, 2006a, 2009a,b, CHALINE et al. 1998, 1998, CARO, 2004]. Ce type de mutation concerne les gènes du développement (homéogènes) [LACOMBE, 2000]. L’angle sphénoïdal des grands singes fossiles n’est pas connu directement, il est cependant possible de situer la loge cérébelleuse sur une trajectoire morphogénétique. Ainsi, l’inion de Sahelanthropus tchadensis (TM 266) se situe sur la trajectoire commune aux paninés et ponginés et non sur celle des homininés, une dichotomie radicale. La région iniaque ne s’est pas développée vers le bas et l’avant comme cela aurait du être le cas si la loge cérébelleuse s’était abaissé davantage au cours du développement intra-utérin [DAMBRICOURT MALASSÉ, 2006b, 2009a,b]. Ce constat infirme l’interprétation d’un équilibre bipède hominien [ZÖLLIKOFER et al. 2005] et confirme l’identité de grand singe anthropoïde [WOLPOFF et al. 2002, 2006, SENUT et PICKFORD, 2004] en accord avec un âge géologique voisinant 10 M.a. [BEAUVILAIN , 2008]. L’espèce Sahelanthropus tchadensis n’était pas plus bipède qu’un grande singe au stade infantile de sa croissance. Il importe donc de distinguer l’acquisition de la verticalité axiale saltationniste car d’origine embryonnaire, de l’équilibre bipède occasionnel observé chez les espèces de primates appartenant à des embryogenèses classées dans des niveaux hiérarchiques taxonomiques plus anciens et de moindre flexion embryonnaire. Il importe également d’intégrer la morphogenèse dans la détermination taxonomique d’un fossile.
LA FLEXION SPHENO-OCCIPITALE ET L’ORIENTATION DES CANAUX SEMI-CIRCULAIRES
Delattre et Fenart (1960) ont initié la recherche de liens possibles entre la bascule de la loge cérébelleuse et celle de l’appareil vestibulaire entraîné par la rotation du rocher sur son axe (figure 2B). Zonneveld et Wind (1985) ont repris cette recherche en l’appliquant à une série de fossiles Australopithecus et Homo (Taung, Sts 5, Stw 53, MLD 37/38, SK 48 et TM 1517), tenant compte de la pente endocrânienne du clivus. L’étude fut ensuite élargie à un plus grand nombre d’hominidés pour être comparée aux Paninés [SPOOR et ZONNEVELD, 1994, 1997]. La pente du clivus des Australopithèques forme un angle avec le canal latéral variant entre 25° et 30°, au lieu de 53° chez un Homo sapiens. La différence tient à sa plus forte déclivité (78° en moyenne) chez Homo sapiens au lieu de 56° sur Sts 5 (Australopithecus africanus). Les paléoprimatologues abordent la région cérébelleuse sous l’angle d’une biomécanique locomotrice, tandis qu’en paléontologie humaine celle-ci est étudiée en fonction de l’enroulement fœtal du télencéphale: « the human labyrinth differs markedly in shape, following a pattern that seems to reflect the characteristic architecture of the human basicranium. Indeed, it is found that labyrinthine and basicranial shape are interspecifically correlated […]. Differences in brain growth and development are proposed as the predominant factor underlying both the unique shap” [HARTCOURT et al. 2004]. Jeffery et Spoor (2002) ont étudié une série de fœtus humain en IRM dans cette perspective, mais les coefficients de corrélations sont avérés négatifs ; en effet en période fœtale la base est en rétro-flexion. Jeffery et Spoor (2004) ont alors poursuivi leur investigation sur la croissance fœtale de l’oreille interne depuis la neuvième semaine, soit au terme de l’organogenèse. Les deux auteurs observent bien une légère migration coronale du canal latéral concomitante de la frontalisation du rocher (alors que l’angle sphénoïdal s’ouvre en moyenne), mais l’écart angulaire entre les canaux semi-circulaire latéral et postérieur est déjà visible à 9 semaines. Son origine n’est donc pas une conséquence mécanique d’une croissance cérébrale différentielle.

Biom. Hum. et Anthropol. 2009, 27, 1-2, p. 0-0. Dambricourt A., Etude au scanner des canaux semi-circulaires d’un anencéphale…
FIGURE 2B. — schéma de l’appareil vestibulaire en vue latérale gauche d’un renard et d’un enfant Homo sapiens, montrant par des vecteurs, les modifications angulaires des trois canaux semi-circulaires et de la cochlée entre le quadrupède et l’homme [DELATTRE et FENART, 1960).
La conformation basi-crânienne embryonnaire d’origine est plane, elle correspond à la forme mammalienne ancestrale.
La plicature apparaît entre les sixième et huitième semaines en relation avec l’orpercularisation de la gouttière neurale sus-jacente, elle résulte d’une rotation du planum basal autour de ses synchondroses intrasphénoïdale et sphéno-occipitale [DAMBRICOURT MALASSE, 1988, 2006a]. Au stade 16-17 de la Carnegie (37–42 jours), la base est formée, entre autres, par le post-sphénoïde et les futurs rochers constitués des ébauches de la cochlée et des capsules otiques. Celles-ci contiennent l’ébauche des canaux semi-circulaires latéral et postérieur qui se forment par apoptose [NICHIKORI et al. 1999], le canal semi-circulaire supérieur n’est pas encore visible [YASUDA et al. 2007]. Au stade 17-18 (42-44 jours) la base est toujours plane, mais elle se développe dans le sens transversal, les phénomènes d’apoptose s’intensifient. Au stade 19 (environ 46 jours), le basi-sphénoïde tourne sur son axe transversal et l’extrémité caudale du basi-occipital montre une légère plicature, cependant celui-ci ne bascule pas encore. Le canal semi-circulaire supérieur est formé [ARNOLD et LANG, 2001]. Au stade 23 (environ 56 jours), le basi-occipital entame sa rotation et entraîne les capsules otiques dans sa course vers l’avant et le bas. La flexion de la base est acquise au terme de la huitième semaine. L’absence de neurulation provoque l’agénésie des arcs vertébraux (rachischisis ou spina bifida), ainsi que des cranio-rachischisis, elle se traduit également par l’absence de flexion de la base. L’étude du cranio-rachischisis permet donc de vérifier si l’écart angulaire entre le canal latéral et le canal postérieur visible à neuf semaines est une adaptation aux contraintes de la flexion sphénoidale, ou si sa formation est d’origine génétique indépendante de celle-ci.
MATERIELS ET METHODES
Les collections d’anthropologie du Musée de l’Homme (Muséum national d’Histoire naturelle) possèdent un spécimen unique de cranio-rachischisis (référencé 24 673). Son planum basale est plan. Ce type de malformation neurologique est rare, de l’ordre de 1‰ en France [ALLOUCHE et al. 1996]. En revanche les données statistiques sur la croissance fœtale humaine normale sont accessibles dans la littérature [JEFFERY et SPOOR, 2004]. Il n’en existe pas sur les paninés, le fœtus de gorille étudié est l’unique spécimen connu des collections nationales du Département de Préhistoire et le premier étudié pour l’oreille interne. De sexe féminin, son âge est estimé au cinquième mois gestationnel [LALLOUET, 2002].
L’âge au décès du fœtus humain anencéphale n’étant pas connu, celui-ci a été estimé par les dimensions longitudinales basion-nasion et le degré de développement des alvéoles maxillaires en comparaison avec un fœtus ne présentant pas d’anomalies (référence 822-1 du Musée de l’Homme). L’anencéphale serait décédé au cours du cinquième mois de gestation (31 semaines). Les trois spécimens ont donc le même âge. Les crânes ont été scannés au Service de Radiologie du Dr. Jacquenod, à l’Hôpital de la Croix Saint Simon (Groupe Hospitalier Diaconesses-Croix Saint Simon de Paris).
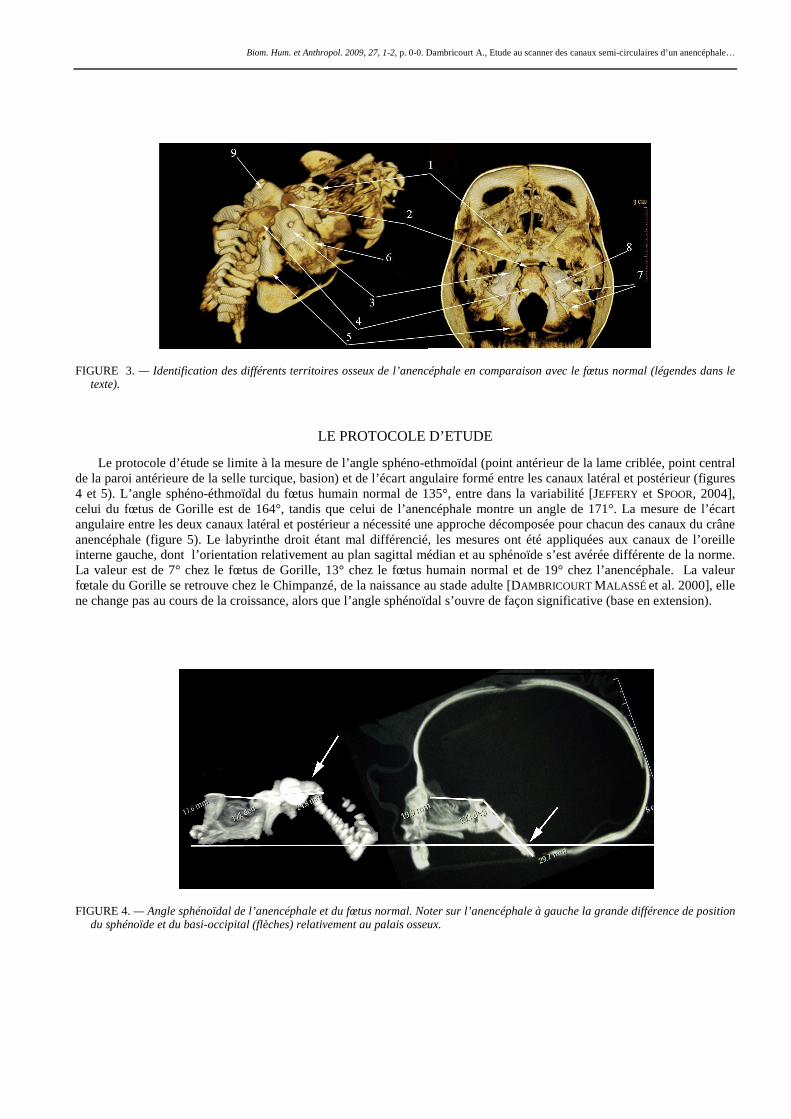
Le fœtus anencéphale est atteint de spina bifida et ne présente pas de voûte (figure 3), les seuls tissus de membranes différenciés sont les écailles du temporal déversées latéralement, et l’ébauche des grandes ailes du sphénoïde. Le maxillaire est bien différencié mais le palais en connexion avec le sphénoïde est étroit en raison d’une croissance transversale neurocrânienne stoppée par l’anencéphalie. De la base du crâne, on identifie l’ébauche des petites ailes du sphénoïde (1), l’espace correspondant à la synchondrose sphéno-occipitale bordé par une ébauche des apophyses clinoïdes postérieures (2), l’extrémité antérieure du rocher (3) et le méat auditif interne en position apicale (9), donc non basculée contrairement au foetus normal (8), le basi-occipital plan et sa partie caudale coudée (4) à hauteur de l’articulation avec chacun des deux exo-occipitaux (5). En revanche l’extrémité distale des rochers bien visible sur le fœtus normal avec les canaux antérieur et postérieur discernables (7) n’apparaît pas, une masse osseuse s’observe dans une position plus antérieure et plus basse (6).
Biom. Hum. et Anthropol. 2009, 27, 1-2, p. 0-0. Dambricourt A., Etude au scanner des canaux semi-circulaires d’un anencéphale…
FIGURE 3. — Identification des différents territoires osseux de l’anencéphale en comparaison avec le fœtus normal (légendes dans le texte).
LE PROTOCOLE D’ETUDE
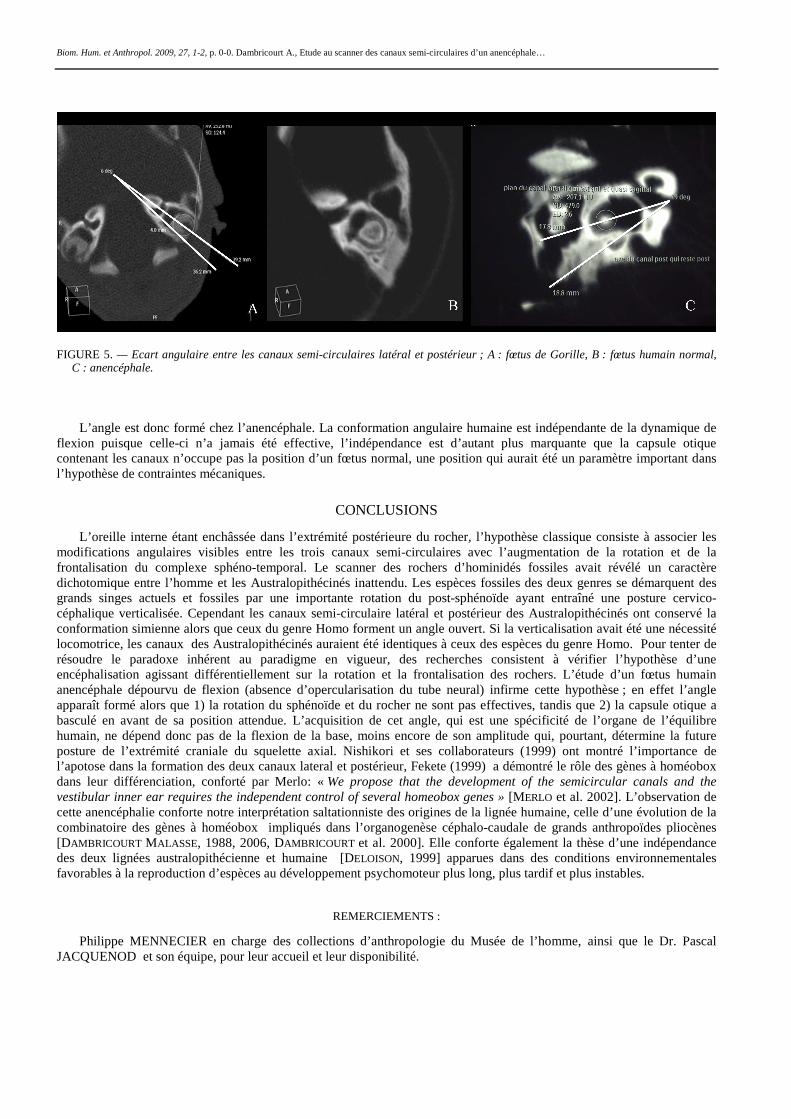
Le protocole d’étude se limite à la mesure de l’angle sphéno-ethmoïdal (point antérieur de la lame criblée, point central de la paroi antérieure de la selle turcique, basion) et de l’écart angulaire formé entre les canaux latéral et postérieur (figures 4 et 5). L’angle sphéno-éthmoïdal du fœtus humain normal de 135°, entre dans la variabilité [JEFFERY et SPOOR, 2004], celui du fœtus de Gorille est de 164°, tandis que celui de l’anencéphale montre un angle de 171°. La mesure de l’écart angulaire entre les deux canaux latéral et postérieur a nécessité une approche décomposée pour chacun des canaux du crâne anencéphale (figure 5). Le labyrinthe droit étant mal différencié, les mesures ont été appliquées aux canaux de l’oreille interne gauche, dont l’orientation relativement au plan sagittal médian et au sphénoïde s’est avérée différente de la norme. La valeur est de 7° chez le fœtus de Gorille, 13° chez le fœtus humain normal et de 19° chez l’anencéphale. La valeur fœtale du Gorille se retrouve chez le Chimpanzé, de la naissance au stade adulte [DAMBRICOURT MALASSÉ et al. 2000], elle ne change pas au cours de la croissance, alors que l’angle sphénoïdal s’ouvre de façon significative (base en extension).
FIGURE 4. — Angle sphénoïdal de l’anencéphale et du fœtus normal. Noter sur l’anencéphale à gauche la grande différence de position du sphénoïde et du basi-occipital (flèches) relativement au palais osseux.
Biom. Hum. et Anthropol. 2009, 27, 1-2, p. 0-0. Dambricourt A., Etude au scanner des canaux semi-circulaires d’un anencéphale…
FIGURE 5. — Ecart angulaire entre les canaux semi-circulaires latéral et postérieur ; A : fœtus de Gorille, B : fœtus humain normal, C : anencéphale.
L’angle est donc formé chez l’anencéphale. La conformation angulaire humaine est indépendante de la dynamique de
flexion puisque celle-ci n’a jamais été effective, l’indépendance est d’autant plus marquante que la capsule otique contenant les canaux n’occupe pas la position d’un fœtus normal, une position qui aurait été un paramètre important dans l’hypothèse de contraintes mécaniques.
CONCLUSIONS
L’oreille interne étant enchâssée dans l’extrémité postérieure du rocher, l’hypothèse classique consiste à associer les modifications angulaires visibles entre les trois canaux semi-circulaires avec l’augmentation de la rotation et de la frontalisation du complexe sphéno-temporal. Le scanner des rochers d’hominidés fossiles avait révélé un caractère dichotomique entre l’homme et les Australopithécinés inattendu. Les espèces fossiles des deux genres se démarquent des grands singes actuels et fossiles par une importante rotation du post-sphénoïde ayant entraîné une posture cervico-céphalique verticalisée. Cependant les canaux semi-circulaire latéral et postérieur des Australopithécinés ont conservé la conformation simienne alors que ceux du genre Homo forment un angle ouvert. Si la verticalisation avait été une nécessité locomotrice, les canaux des Australopithécinés auraient été identiques à ceux des espèces du genre Homo. Pour tenter de résoudre le paradoxe inhérent au paradigme en vigueur, des recherches consistent à vérifier l’hypothèse d’une encéphalisation agissant différentiellement sur la rotation et la frontalisation des rochers. L’étude d’un fœtus humain anencéphale dépourvu de flexion (absence d’opercularisation du tube neural) infirme cette hypothèse ; en effet l’angle apparaît formé alors que 1) la rotation du sphénoïde et du rocher ne sont pas effectives, tandis que 2) la capsule otique a basculé en avant de sa position attendue. L’acquisition de cet angle, qui est une spécificité de l’organe de l’équilibre humain, ne dépend donc pas de la flexion de la base, moins encore de son amplitude qui, pourtant, détermine la future posture de l’extrémité craniale du squelette axial. Nishikori et ses collaborateurs (1999) ont montré l’importance de l’apotose dans la formation des deux canaux lateral et postérieur, Fekete (1999) a démontré le rôle des gènes à homéobox dans leur différenciation, conforté par Merlo: « We propose that the development of the semicircular canals and the vestibular inner ear requires the independent control of several homeobox genes » [MERLO et al. 2002]. L’observation de cette anencéphalie conforte notre interprétation saltationniste des origines de la lignée humaine, celle d’une évolution de la combinatoire des gènes à homéobox impliqués dans l’organogenèse céphalo-caudale de grands anthropoïdes pliocènes [DAMBRICOURT MALASSE, 1988, 2006, DAMBRICOURT et al. 2000]. Elle conforte également la thèse d’une indépendance des deux lignées australopithécienne et humaine [DELOISON, 1999] apparues dans des conditions environnementales favorables à la reproduction d’espèces au développement psychomoteur plus long, plus tardif et plus instables.
REMERCIEMENTS :
Philippe MENNECIER en charge des collections d’anthropologie du Musée de l’homme, ainsi que le Dr. Pascal JACQUENOD et son équipe, pour leur accueil et leur disponibilité.

Biom. Hum. et Anthropol. 2009, 27, 1-2, p. 0-0. Dambricourt A., Etude au scanner des canaux semi-circulaires d’un anencéphale…
RÉFÉRENCES
ALLOUCHE C., BOUDLALAL A., ELHAIK S. (1996) - Cranio-rachischisis et cœlosomie totale : A propos d’un cas. Revue française de gynécologie et d’obstétrique, 91, 1-2: 39-41.
ARNOLD W. H., LANG T. (2001) - Development of the membranous labyrinth of human embryos and fetuses using computer aided 3D-reconstruction. Annals of Anatomy, Anatomischer Anzeiger. 183, 1 : 61-66.
BEAUVILAIN, A. (2008) - The contexts of discovery of Australopithecus bahrelghazali (Abel) and of Sahelanthropus tchadensis (Toumaï): unearthed, embedded in sandstone, or surface collected? South African Journal of Science 104(5/6): 165-168.
CARO P. (2004) - Développement embryonnaire et évolution. Entretien avec Anne Dambricourt Malassé. La Lettre de l’Académie des Sciences. Automne: 14-15.
CHALINE J., A. DAMBRICOURT MALASSÉ, B. DAVID, B., F. MAGNIEZ-JANNIN, D. MARCHAND, COURANT F., MILLET J.J. (1998) - Quantification de l'évolution morphologique du crâne des hominidés et hétérochronies. Comptes Rendus de l'Académie des Sciences de la Terre et des Planètes, 326: 291-298.
CONROY C., VANNIER M.W., TOBIAS P.V. (1990) - Endocranial Features of Australopithecus africanus by 2- and 3-D Computed Tomography. Science vol. 247: 838-841.
DAMBRICOURT MALASSÉ A. (1988) - Hominisation et foetalisation. C. R. Académie des Sciences, 307, II: 199-204.
DAMBRICOURT MALASSÉ A. (1993) - Continuity and discontinuity during modalities of hominization. Quaternary International, 19: 85-100.
DAMBRICOURT MALASSÉ A., MARTIN J.P., de KERVILER E. (2000) - Test des corrélations angulaires au cours de l’ontogenèse entre la flexion sphénoïdale et les canaux semi-circulaires de Pan troglodytes. Implications en paléontologie humaine. In : L’identité humaine en question. Nouvelles problématiques et nouvelles technologies en paléontologie humaine et en paléoanthropologie biologique. (Ed) P. Andrieux, D. Hadjouis, A Dambricourt Malassé, Artcom : 24-42.
DAMBRICOURT MALASSÉ A. (2005) - Un nouvel angle de la base pour une meilleure compréhension de la morphogenèse basicrânienne des hominidés et des modalités évolutives. In: « Biométrie et anthropologie de la tête, de la face et du cou », Biométrie Humaine et Anthropologie, 23: 1-2:53-62.
DAMBRICOURT MALASSÉ A. (2006a) - Evolution du chondrocrâne et de la face des grands anthropoïdes miocènes jusqu’à Homo sapiens, continuités et discontinuités. Comptes Rendus de l’Académie des Sciences, Paléontologie systématique. Paléovol, 5 :109-117.
DAMBRICOURT MALASSÉ A. (2006b) - La question des origines de la lignée humaine : les modèles évolutionnistes confrontés aux fossiles, implications en orthopédie et posturologie. Les Cahiers du C.E.O.P.S., Toulouse, 4: 38-57.
DAMBRICOURT MALASSÉ A. (2009a) - Embryogenic cranial evidences in fossil hominids, taxonomical and phylogenetical implications in human palaeontology. Actes du 16th World Congress of the IUAES, 27-31 July 2009, Kunming, China.
DAMBRICOURT MALASSÉ A. (2009b) - The human lineage: macro-evolutionary process acting during embryogeny with emergent macro-evolutionary implications. Actes du Colloque L’évolution biologique: les faits et les théories. Une évaluation critique 150 ans après ‘L'origine des espèces’, 3-7 mars 2009, Rome.
DELATTRE A., FENART R. (1960) - L’hominisation du crâne. Ed. CNRS.
DELOISON Y. (1999) - L’Homme ne descend pas d’un Primate arboricole. Biom. Hum. et Anthropol. 17 : 147-150.
DESHAYES M.J. (1986) - Biodynamique Cranio-faciale et Orthodontie. Masson.
DESHAYES M.J. (1991) - Reconsidération de la croissance cranio-faciale au cours du développement et de l’évolution. Conséquences en orthopédie dento-maxillo-faciale - Rev Orthop Dento-faciale: 25, 3 : 353-365.
DESHAYES M.J., DAMBRICOURT MALASSÉ A. (1990) - Analyse des différents types architecturaux cranio-faciaux par l’approche ontogénique de l’hominisation., Rev. Stom. Chir.Max.Fac., 4 : 249-258.
FEKETE D. M. (1999) - Development of the vertebrate ear insight from knockouts and mutants. Tr. Neuroscie. 22: 263–269.
GUDIN R.G, GODARD B. (1973) - Les troubles dyspnéiques des sphères oro-faciales et pharyngées. Leur influence sur la morphogenèse cranio-faciale et leur rapport avec la posture mandibulaire en période de croissance. Revue d’O.D.F., 23, 4: 407- 425.
HARCOURT-SMITH W. E. H., AIELLO L. C. (2004) - Fossils, feet and the evolution of human bipedal locomotion. J Anat. 204, 5: 403–416.
JEFFERY N., SPOOR F. (2002) - Brain size and the human cranial base: a prenatal perspective. Am. J. Phys. Anthrop 118 (4):324-340.
JEFFERY N., SPOOR F. (2004) - Prenatal growth and development of the modern human Labyrinth. J Anat, 2:71-92.
LACOMBE D. (2000) - Maladies des gènes du développement. médecine/sciences. 16 : 354-362.
LALLOUET F. (2002) - Atlas d’anatomie du crâne d’un foetus humain et d’un foetus de gorille de 5 mois sur support CD-ROM et analyse morphologique comparative. Mémoire de DEA, Centre Universitaire des Saints Pères, Université René Descartes.

Biom. Hum. et Anthropol. 2009, 27, 1-2, p. 0-0. Dambricourt A., Etude au scanner des canaux semi-circulaires d’un anencéphale…
MAC KEE J. (1999) - The Autocatalytic Nature of Hominid Evolution in African Plio-Pleistocene Environments. In: African Biogeography, Climate Change and Early Hominid Evolution, Eds. F. Schrenk & T. Bromage. Oxford University Press.
MERLO G.R., L. PALEARI, S. MANTERO, B. ZEREGA, M. ADAMSKA, S. RINKWITZ, BOBER E., LEVI G. (2002) - The Dlx5 Homeobox Gene is Essential for Vestibular Morphogenesis in the Mouse Embryo through a BMP4-Mediated Pathway. Developmental Biology, 248: 157-169.
NISHIKORI T., H. TOSHIHISA, K. HIDEYUKI, HIROKI O. (1999) - Apoptosis during inner ear development in human and mouse embryos: an analysis by computer-assisted three-dimensional reconstruction In: Anatomy and Embryology, 200, 1: 19-26.
RAK Y., CLARK HOWELL F. (1973) - Cranium of a juvenile Australopithecus boisei from the Lower Omo Basin, Ethiopa. Am. J. Phys. Anthr. 48, 3: 345- 364.
ROOK L., L. BONDIOLI, F. CASALI, M. ROSSI, M. KÖHLER, Á S. MOYÁ SOL, MACCHIARELLI R. (2004) - The bony labyrinth of Oreopithecus bambolii, Journal of Human Evolution 46:347-354.
SENUT B., PICKFORD M. (2005) - Comment Orrorin a changé nos conceptions sur les origines des hominidés. Anthropologie, 43, 2-3 : 111-119.
SENUT B., PICKFORD M. (2004) - La dichotomie grands singes–homme revisitée. In : Les premiers hominidés, Comptes Rendus Palevol, 3, 4: 265-276.
SPOOR F. (1996) - The ancestral morphology of the hominid bony labyrinth: the evidence from Dryopithecus. Am. J. phys. Anthrop. 22 Suppl., p. 219.
SPOOR F., ZONNEVELD F. (1994) - The bony labyrinth in Homo erectus: a preliminary report. Cour. Forsch. -Institut Sencenberg, 171: 251-256.
SPOOR F., ZONNEVELD F. (1997) - The bony labyrinth of Sts 5, South African Journal of Science, 93: 180.
TOBIAS P.V. (2004) - African Origins. Innovation for Development DST Magazine, 4: 8-11.
WHITE T.D., SUWA G., ASFAW B. (1994) - Australopithecus ramidus, a new species of early hominid from Aramis, Ethiopia. Nature, 371: 306-312.
WOLDGABRIEL G., WHITE T.D., SUWA G., RENNE G. (1994) - Ecological and temporal placement of early pliocene hominids at Aramis, Ethiopia. Nature, 371: 330-333.
WOLPOFF M. H., SENUT B., PICKFORD M., HAWKS J. (2002) - Sahelanthropus or “Sahelpithecus” ? Nature, 419: 581-582.
WOLPOFF M. H, HAWKS J., SENUT B., PICKFORD M., AHERN J. (2006) - Is the Toumaï cranium TM 266 an hominid ? PaleoAnthropology: 36-50.
YASUDA M., S. YAMADA, C. UWABE, K. SHIOTA, YASUDA Y. (2007) - Three-dimensional analysis of inner ear development in human embryos. Anatomical Science International, 82, 3:156-63.
ZOLLIKOFER C. P. E., PONCE DE LEON M. S., LIEBERMAN D. E., GUY F., PILBEAM D., LIKIUS A., MACKAYE H. T., VIGNAUD P., BRUNET M., (2005) - Virtual cranial reconstruction of Sahelanthropus tchadensis. Nature, vol. 434: 755-759.
ZONNEVELD, F.W., WIND J. (1985) - High-Resolution computed tomography of fossil hominid skulls: a new method and some results. In: Hominid Evolution: Past, Present, Future: 427-436.
ZONNEVELD, F.W., SPOOR C.F. (1997) - The bony labyrinth of Sts 5. In: 50 years of ‘Mrs Ples’ Research Brief. South African Journal of Science, 93: 180.





























