Embed Size (px)
Citation preview

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Jan. 2011, p. 281–290 Vol. 77, No. 10099-2240/11/$12.00 doi:10.1128/AEM.01403-10Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Ethanolamine Utilization Contributes to Proliferation of Salmonella entericaSerovar Typhimurium in Food and in Nematodes�
Shabarinath Srikumar and Thilo M. Fuchs*Zentralinstitut fur Ernahrungs- und Lebensmittelforschung (ZIEL), Abteilung Mikrobiologie,
Technische Universitat Munchen, Weihenstephaner Berg 3, D-85354 Freising, Germany
Received 11 June 2010/Accepted 24 October 2010
Only three pathogenic bacterial species, Salmonella enterica, Clostridium perfringens, and Listeria monocyto-genes, are able to utilize both ethanolamine and 1,2-propanediol as a sole carbon source. Degradation of thesesubstrates, abundant in food and the gut, depends on cobalamin, which is synthesized de novo only underanaerobic conditions. Although the eut, pdu, and cob-cbi gene clusters comprise 40 kb, the conditions underwhich they confer a selection advantage on these food-borne pathogens remain largely unknown. Here we usedthe luciferase reporter system to determine the response of the Salmonella enterica serovar Typhimuriumpromoters PeutS, PpocR, PpduF, and PpduA to a set of carbon sources, to egg yolk, to whole milk, and to milkprotein or fat fractions. Depending on the supplements, specific inductions up to 3 orders of magnitude wereobserved for PeutS and PpduA, which drive the expression of most eut and pdu genes. To correlate thesesignificant expression data with growth properties, nonpolar deletions of pocR, regulating the pdu and cob-cbigenes, and of eutR, involved in eut gene activation, were constructed in S. Typhimurium strain 14028. Duringexponential growth of the mutants 14028�pocR and 14028�eutR, 2- to 3-fold-reduced proliferation in milk andegg yolk was observed. Using the Caenorhabditis elegans infection model, we could also demonstrate that theproliferation of S. Typhimurium in the nematode is supported by an active ethanolamine degradation pathway.Taking these findings together, this study quantifies the differential expression of eut and pdu genes underdistinct conditions and provides experimental evidence that the ethanolamine utilization pathway allowssalmonellae to occupy specific metabolic niches within food environments and within their host organisms.
An estimated 26% of all food-borne infections in the UnitedStates are caused by Salmonella enterica (39). Consumption ofcontaminated foods, such as poultry, eggs, raw milk, seafood,and fresh produce, has been the major cause of salmonellosis(18). Eggs and egg products are the food vehicles most oftenidentified in S. enterica outbreaks (10). Dairy farms are well-known reservoirs of S. enterica (21, 41). Between 1992 and2000, 52% of the food-borne outbreaks in England and asimilar proportion in France were attributed to raw milk (16,21). In the United States, the sale of contaminated raw milk tothe public has led to the outbreak of multidrug-resistant sal-monellosis in California and Washington (45). Several studiesreport that S. enterica is the most predominant organism in rawbulk tank milk; Salmonella enterica serovar Typhimurium andSalmonella enterica serovar Newport are the major serotypesidentified here (23, 24, 41).
A variety of gut and environmental genera, including Sal-monella, Escherichia, Enterococcus, and Arthrobacter, can uti-lize ethanolamine as the sole source of carbon, nitrogen, andenergy (6, 12, 17, 48; reviewed in reference 19). The precursormolecule is phosphatidylethanolamine, an abundant phospho-lipid in bacterial and mammalian cell membranes that is bro-ken down to glycerol and ethanolamine by phophodiesterases(33, 44, 48). The enzymes responsible for the degradation of
ethanolamine are encoded mainly by 17 clustered genes whosefunctions have been characterized in great detail (29, 55). Theethanolamine lyase EutBC degrades ethanolamine to acetal-dehyde and ammonia within a multiprotein complex termed acarboxysome (48, 49). This microcompartment is assumed toprevent the loss of acetaldehyde and to protect the cell from itspotential toxic effects (11, 42). The ammonia serves as thecellular supply of reduced nitrogen, while acetaldehyde is con-verted to acetyl coenzyme A (acetyl-CoA) by the eutE-encodedenzyme aldehyde oxidoreductase (48, 49). Acetyl-CoA is sub-sequently absorbed in various metabolic cycles, such as thetricarboxylic acid (TCA) cycle, the glyoxylate cycle, or lipidbiosynthesis. Under aerobic conditions, the activity of EutBCdepends on the exogenous supply of cobalamin (vitamin B12),which is synthesized de novo in the presence of the cob-cbioperon under anaerobic conditions only (25, 49, 50). In manygenomes, the cob-cbi operon is clustered with the pdu genes,which are responsible for cobalamin-dependent propanedioldegradation, and both operons are transcriptionally activatedby the common regulator pocR when Salmonella grows on poorcarbon sources (7, 8, 13, 46, 61). The electron acceptor tetra-thionate is essential for the anaerobic growth of Salmonella onethanolamine and 1,2-propanediol (43). Products of the pduoperon have been found in high abundance by proteome anal-ysis of intracellularly replicating S. Typhimurium strain 14028,and its genes are upregulated under in vivo-mimicking condi-tions (2, 52). Three pathogenic bacteria are able to use bothethanolamine and 1,2-propanediol: the food-borne pathogensS. enterica, Clostridium perfringens, and Listeria monocytogenes(30). Recently, we were able to demonstrate that cobalaminsynthesis and propanediol degradation are required for the
* Corresponding author. Mailing address: Zentralinstitut fur Er-nahrungs- und Lebensmittelforschung (ZIEL), Abteilung Mikrobiolo-gie, Technische Universitat Munchen, Weihenstephaner Berg 3,D-85354 Freising, Germany. Phone: 49-8161-713859. Fax: 49-8161-714492. E-mail: [email protected].
� Published ahead of print on 29 October 2010.
281
on June 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

intracellular replication of S. Typhimurium, as suggested pre-viously with in vivo expression techniques (22, 28). Evidencehas also been provided that pdu mutations are responsible forattenuation of the virulence of Salmonella (14) and that theglobal Salmonella virulence regulators CsrA and Fis influencethe expression of the eut gene (27, 34). Moreover, a lack ofEutBC activity in L. monocytogenes results in attenuated pro-liferation of this food-borne pathogen in epithelial cells (26),and all listerial eut genes are upregulated in the intestine in amouse infection model (58).
However, much less is known about nonmammalian envi-ronments in which the eut and pdu genes provide a selectionadvantage for salmonellae. Using luciferase-based reporter as-says, we investigated the expression of the eut and pdu operonsin minimal medium supplemented with a set of carbon sources,as well as in milk and egg yolk. eutR and pocR deletion mutantsof S. Typhimurium were tested for their fitness under the sameconditions and during proliferation in Caenorhabditis elegans.Our data suggest that ethanolamine degradation constitutes aneglected but crucial metabolic determinant of Salmonella-associated food poisoning and that it also contributes to S.Typhimurium proliferation in vivo.
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions. The bacterial strains andplasmids used in this study are listed in Table 1. S. enterica strains were grown inLuria-Bertani (LB) broth (10 g/liter tryptone, 5 g/liter yeast extract, 5 g/literNaCl) or in minimal medium (MM or M9) supplemented with 2 mM MgSO4, 0.1mM CaCl2 and 27.8 mM (0.5% [wt/vol]) glucose. If appropriate, glucose wasreplaced by 66 mM (0.5% [wt/vol]) 1,2-propanediol or 82 mM (0.5% [wt/vol])ethanolamine. To produce growth on the latter two substrates under aerobicgrowth conditions, the media contained cyanocobalamin (Sigma-Aldrich,Taufkirchen, Germany) at a final concentration of 200 nM. For solid media,1.5% agar (wt/vol) was added. If appropriate, sodium tetrathionate was added asthe final electron acceptor to a final concentration of 40 mM. MacConkey agar(Becton Dickinson, Heidelberg, Germany) with ethanolamine was also used asan indicator medium for acid production. Where necessary, the medium con-
tained the antibiotic ampicillin (150 �g/ml), kanamycin (50 �g/ml), or chloram-phenicol (25 �g/ml). For all growth and promoter probe experiments, bacterialstrains were grown in LB medium overnight at 37°C, washed twice with phos-phate-buffered saline (PBS), and then adjusted to an optical density at 600 nm(OD600) of 0.001 in the desired liquid growth medium or in MM. Growth curveswere obtained from bacterial cultures incubated at 37°C in sterile 25-ml bottleswith 25 ml of MM containing the appropriate carbon source. The OD600 wasmeasured at appropriate time intervals.
Standard procedures. DNA manipulations and isolation of chromosomal orplasmid DNA were performed according to standard protocols (51) by followingthe manufacturers’ instructions. GeneRuler DNA Ladder Mix (Fermentas, St.Leon-Rot, Germany) was used as a marker for DNA analysis. Plasmid DNA wastransformed via electroporation by using a Bio-Rad Gene pulser II as recom-mended by the manufacturer and as described previously (28). PCRs werecarried out with Taq polymerase (Fermentas). As a template for PCR, chromo-somal DNA, plasmid DNA, or an aliquot of a single colony resuspended in 100�l H2O was used. The oligonucleotides used for PCRs are listed in Table 2. S.Typhimurium gene numbers refer to LT2 annotation (NC 003197). The homepages for NCBI and Microbes Online were used to determinate the distributionof the pdu and eut clusters in different serotypes of S. enterica. Promoter se-quences located upstream of the genes identified were predicted with BPROM(Softberry).
Construction of deletion mutants. STM2036 (pocR), STM2454 (eutR), andSTM4048-STM4049 (rhaS-rhaR) were deleted in frame in S. Typhimurium strain14028 by the one-step deletion protocol based on the � Red recombinase system(15). Briefly, PCR products comprising the kanamycin resistance cassette ofplasmid pKD4, including the FLP recombination target (FRT) sites, were gen-erated using pairs of 70-nucleotide primers that included 20-nucleotide primingsequences for pKD4 as template DNA (Table 2). Homology extensions of 50 bpoverlapped 18 nucleotides of the 5� end and 36 nucleotides of the 3� end of thetarget gene (38). The fragment DNA was transferred to 14028 cells harboringplasmid pKD46. Allelic replacement of the kanamycin resistance gene cassettewas confirmed by PCR, and nonpolar deletion mutants were obtained followingtransformation of pCP20. Gene deletions were verified by PCR analysis andDNA sequencing with the primers listed in Table 2.
Cloning of promoter fusions. The promoter regions of approximately 250 bpupstream of the start codons of STM2036 (pocR), STM2037 (pduF), STM2038(pduA), and STM2470 (eutS) were amplified from the chromosomal DNA of14028 by PCR using the primers listed in Table 2. The fragments were thencloned via EcoRI and BamHI into pDEW201. After transformation into Esch-erichia coli DH5�, plasmids were verified by PCR analysis and sequencing. Thecorrect plasmids were transformed into strain 14028 cells by electroporation.
TABLE 1. Bacterial strains and plasmids used in this study
Strain or plasmid Description Source
StrainsDH5� Escherichia coli DH5�MCR Invitrogen, Carlsbad, CA14028 Salmonella enterica subsp. enterica serovar Typhimurium ATCCParatyphi B S. enterica subsp. enterica serovar Paratyphi B ATCCNewport S. enterica subsp. enterica serovar Newport ATCCVirchow S. enterica subsp. enterica serovar Virchow ATCCGallinarum S. enterica subsp. enterica serovar Gallinarum 287/91 57Choleraesuis S. enterica subsp. enterica serovar Choleraesuis ATCCArizonae S. enterica subsp. arizonae ATCC14028�pocR In-frame pocR (STM2036) deletion mutant This study14028�eutR In-frame eutR (STM2454) deletion mutant This study14028�rhaSR In-frame rhaSR (STM4048-STM4049) deletion mutant This study
PlasmidspKD4 Kanr; pir dependent, FRT sites CGSC, YalepKD46 Lambda red helper plasmid; Ampr CGSC, YalepCP20 FLP recombinase plasmid; Cmr Ampr CGSC, YalepDEW201 Promoter probe vector; Ampr; luxCDABE 60pDEW-PpduA pDEW201 with 250 bp upstream of pduA (STM2038) This studypDEW-PpocR PDEW201 with 250 bp upstream of pocR (STM2036) This studypDEW-PpduF PDEW201 with 250 bp upstream of pduF (STM2037) This studypDEW-PeutS PDEW201 with 250 bp upstream of eutS (STM2470) This studypDEW-PargS PDEW201 with 244 bp upstream of argS 32pDEW-�STM0047� pDEW201 with 350-bp intragenic fragment of STM0047 32
282 SRIKUMAR AND FUCHS APPL. ENVIRON. MICROBIOL.
on June 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

Preparation of milk and food fractions. Pasteurized milk and poultry eggsused in this assay were obtained from the local markets and were used foranalysis under sterile conditions. Milk was taken directly for analysis. The yolkwas separated from the poultry egg and was mixed with a 1% NaCl solution toa final concentration of 45%.
Quantification of promoter activities. Raw milk and egg yolk preparationswere taken directly for the assay. The milk fractions were prepared to a concen-tration of 1% in MM, stirred constantly for dissolution, and then used for theassay. The ampicillin-resistant reporter strains carrying the corresponding plas-mid were grown overnight in LB broth, washed twice in phosphate-bufferedsaline, and inoculated into the corresponding food material to an OD600 of 0.001.Bioluminescence measurements were performed in microtiter plates. At appro-priate time points, 200 �l of each sample was transferred to a 96-well plate, andthe OD600 and the emitted bioluminescence, measured as relative light units(RLU), were recorded in a Wallac Victor3 1420 multilabel counter (Perkin-Elmer Life Sciences, Turku, Finland). At the same time points, aliquots wereplated onto LB plates containing the antibiotic ampicillin. The resulting bacterialcounts were used to determine the corresponding OD600 values from a curve ofthe OD600 value versus the CFU per milliliter, prepared earlier from a Salmo-nella culture in LB with ampicillin. These OD600 values were used to calculatethe respective RLU/OD600 values.
Growth assays of strain 14028 and mutant 14028�eutR in food. Strains 14028and 14028�eutR were initially starved by growing the bacterium overnight in MMwith glucose. Bacteria were washed twice in PBS, inoculated into 50 ml of theparticular food sample to an OD600 of 0.001, and incubated at 37°C withoutshaking. Aliquots of the spiked food samples were taken at regular intervals andwere plated onto LB broth-based agar plates at appropriate dilutions for deter-mination of the counts.
C. elegans infection and quantification of intestinal bacterial cells. Mainte-nance of wild-type C. elegans N2 (var. Bristol), including feeding, transfer, andsynchronization, was performed according to standard procedures (54). Briefly,an E. coli OP50 overnight culture was seeded on nematode growth medium(NGM) agar plates containing 3.0 g NaCl, 2.5 g peptone, 1.0 ml of 1 M CaCl2,1.0 ml cholesterol (5-mg/ml stock prepared in 95% ethanol), 25.0 ml of 1 MKPO4 buffer (108.3 g/liter KH2PO4 and 35.6 g/liter K2HPO4; pH 6.0), 1.0 ml of1 M MgSO4, and 17.0 g high-strength Bacto agar per liter (36). Plates with E. coliOP50 were incubated overnight at room temperature and were stored at 4°C. N2worms were cultivated at 22°C and were transferred to new plates every 2 to 3days. A 50-�l volume of an overnight culture of bacterial strains was spread onto
NGM agar plates (diameter, 8.5 cm), which were then incubated overnight at theappropriate temperature. Plates were equilibrated to room temperature (22°C)before use. C. elegans larvae at larval stage 4 (L4) were individually transferredto the bacterial lawn. The infection assay was performed at 22°C for 5 h. Theinfected worms were then transferred back to NGM with E. coli OP50 until theend of the experiment.
For quantification, nematodes were transferred at appropriate time points to600 �l ice-cold lysis buffer (M9 buffer with 0.1% Triton X-100) and were shakenfor 10 min at 400 rpm to release bacteria from the cuticula (53). Nematodes weresedimented by centrifugation for 2 min at 430 � g, washed twice with M9 buffer,and resuspended in 1 ml M9 buffer. After the addition of 500 �l of 1.0-mm-diameter zirconia silica beads (BioSpec, Bartlesville, OK), nematodes were dis-rupted in a FastPrep-24 instrument (MP Biomedicals, Solon, OH) for 20 s atmaximal speed, and the suspension was placed on ice. This step was repeatedtwice. Dilutions were made in LB medium, and aliquots of the suspensions wereplated on MacConkey agar containing 1% lactose. The colorless colonies ofSalmonella were then enumerated.
RESULTS
Distribution of the cob, pdu, and eut gene clusters in salmo-nellae. The genetic organization and distribution of the cob,pdu, and eut operons of the S. Typhimurium LT2 genome werecompared with those of different Salmonella serotypes usingamino acid homology BLAST programs (4). Within the S.Typhimurium LT2 genome, 20 genes responsible for cobal-amin biosynthesis (cob operon) and 23 genes responsible for1,2-propanediol utilization (pdu operon) are clustered in agenomic island from STM2016 to STM2058 (GEI2016/2058)(Fig. 1). The eut operon comprises 17 genes encoding proteinscontributing to ethanolamine degradation. In Salmonella en-terica subsp. enterica serovar Paratyphi B strain SPB7, Salmo-nella enterica subsp. enterica serovar Choleraesuis strain SC-B67, S. enterica subsp. enterica serovar Paratyphi strainAKU_12601, S. enterica subsp. enterica serovar Agona strain
TABLE 2. Primers used in this study
Primer name Sequence Target
PpocR FP TGAATTCGCTCATCTGACGTTAATC pocR promoter regionPpocR RP TGGATCCATAGCAATATTTTGCTATTGCPpduF FP TGAATTCTGAGTTCTGAGTTCAGAG pduF promoter regionPpduF RP TGGATCCGGAAATGCGAACCAGATTCACPpduA FP TGAATTCGCGTCTTTCGCCATATC pduA promoter regionPpduA RP TGGATCCAAAGACCTCGCATGGAGPeutS FP TGAATTCATTTTTATCAATGGTAGTAATAC eutS promoter regionPeutS RP TGGATCCTGCTCACCGTCTCCACTest pocR FP ATGATTTCTGCGAGCGC pocR flanking regionTest pocR RP TCATAACGATGGAGGATGTest eutR FP ATGAAAAAGACCCGTACA eutR flanking regionTest eutR RP TCAAGCCCATTGCCGCTest rhaSR FP ATGACCGTACTGCATAG rhaSR flanking regionTest rhaSR RP TTAATCGCTTTGATTACTGDel pocR FPa ATCAAAAATCGGCAATAGCAAAATATTGCTATATGATTTCTGCGAGCGCTGT
GTAGGCTGGAGCTGCTTCpocR
Del pocR RPa TGAGGGGTTTTATCTCATAACGATGGAGGATGAGAATTCTCATTTATCTGCATATGAATATCCTCCTTA
Del eutR FPa GGCGTCCTGGCGGGCGTCCGGCATAGAAGATCATGAAAAAGACCCGTACAGTGTAGGCTGGAGCTGCTTC
eutR
Del eutR RPa CCTCCCCATCAATATCAAGCCCATTGCCGCATCCGTTGATGCAGCGTCAACATATGAATATCCTCCTTA
Del rhaSR FPa TTCGGACATGTTGAATTCAGGAGGAAGCAACCATGACCGTACTGCATAGCGTGTAGGCTGGAGCTGCTTC
rhaSR
Del rhaSR RPa TTAGGCATGGCCAGTTAATCGCTTTGATTACTGAGATGACGCCATTGGCTCATATGAATATCCTCCTTA
a Used for nonpolar gene deletion.
VOL. 77, 2011 ETHANOLAMINE UTILIZATION BY SALMONELLA ENTERICA 283
on June 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

SL483, S. enterica serovar Typhi strain CT18, S. enterica subsp.enterica serovar Paratyphi A strain ATCC 9150, S. entericasubsp. enterica serovar Weltevreden strain HI_NO5-537, S.enterica subsp. enterica serovar Virchow strain SL491, S. en-terica subsp. enterica serovar Newport strain SL317, and S.enterica subsp. arizonae strain RSK2980, the genetic organiza-tion of the eut, pdu, and cob gene cluster is essentially similarto that of the clusters in the reference strain S. TyphimuriumLT2, and the encoded amino acid sequences are homologouswith at least 95% identity to the LT2 counterparts. However,minor aberrations among those serovars with respect to ab-sence and replacements of single genes within the sequencedclusters were observed. S. Choleraesuis strain SC-B67 containsa hypothetical protein between pduT and pduU. S. Newportstrain SL317 lacks pduP, encoding a propionaldehyde dehy-drogenase. Within the S. Newport strain SL317 genome, pduG,encoding a 1,2-propanediol dehydratase reactivation protein,pduU, encoding a polyhedral body protein, and pduW, encod-ing a propionate kinase, are replaced by a glycerol dehydroge-nase reactivation protein, a putative ethanolamine utilizationprotein, and an acetate kinase, respectively. The two latterreplacements were also found in the S. Virchow strain SL491genome. Here, pduP, encoding a propionaldehyde dehydroge-nase, is replaced with eutE, encoding a putative aldehyde oxi-doreductase. A major island reduction was observed in theSalmonella enterica subsp. enterica serovar Gallinarum strain287/91 genome, which lacks pocR, pduO, pduG, cbiO, cbiK,cbiD, and cbiC (Fig. 1). The eut operon is conserved within allthe tested Salmonella serovars mentioned above except S.Choleraesuis strain SC-B67, S. Newport strain SL317, and S.Virchow strain SL491. S. Choleraesuis strain SC-B67 lackseutK, eutC, and eutA (Fig. 1); S. Virchow strain SL491 lackseutM and eutN; and S. Newport strain SL317 lacks eutM.Lack of EutM and EutN in Salmonella Typhimurium does not
inhibit the bacterium from utilizing ethanolamine as the car-bon source (11, 29).
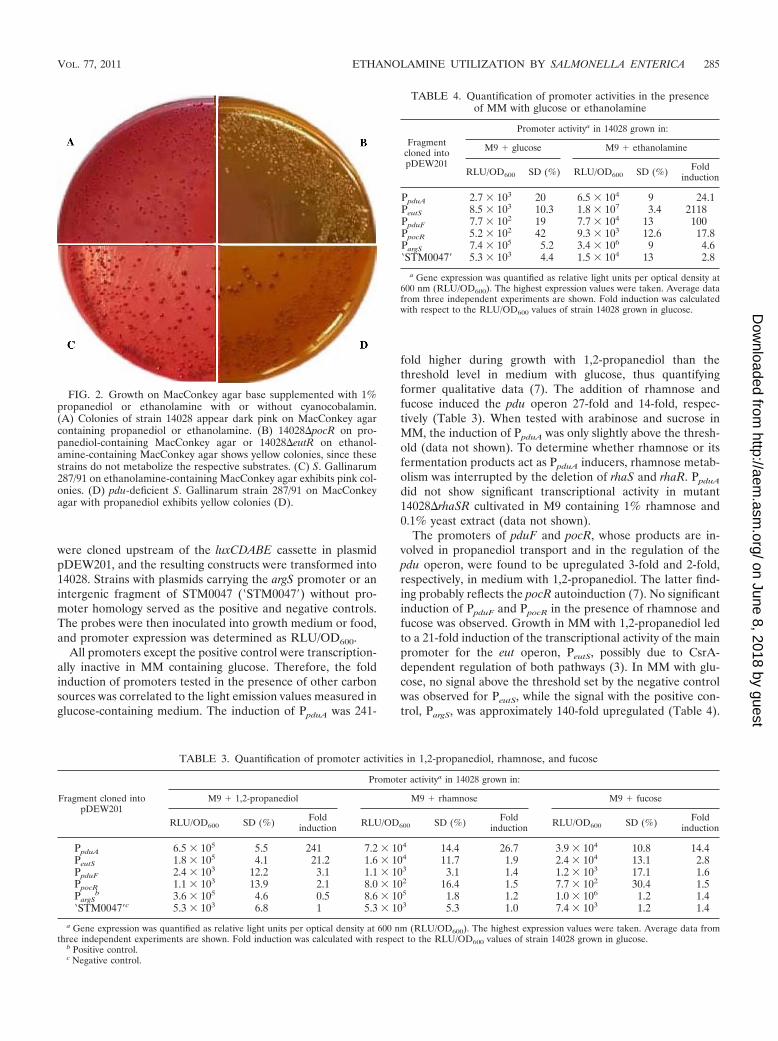
Correlation of genotype and growth properties. For thestudy of genotypes and growth properties, we deleted pocR,encoding the pdu transcriptional regulator (7), and eutR, en-coding the positive regulator of the eut operon, resulting in thenonpolar deletion mutants 14028�pocR and 14028�eutR. Bothgenes appeared to be essential for acid production on Mac-Conkey agar base or MM supplemented with 1,2-propanediolor ethanolamine (Fig. 2A and B), thus confirming the litera-ture (29, 47, 49). The S. enterica subsp. arizonae and S. entericaserovar Paratyphi B, Choleraesuis, Newport, Virchow, andGallinarum strains were streaked onto MacConkey agar platescontaining 1,2-propanediol or ethanolamine and cyanocobal-amin as the sole carbon source (Table 1). All but one serovartested showed pink colonies due to a decrease in the pH upon1,2-propanediol or ethanolamine utilization (35). S. Gallina-rum strain 287/91 utilizes ethanolamine, but not propanediol,irrespective of the presence or absence of cyanocobalamin, dueto an incomplete pdu operon (Fig. 1 and 2C and D). While S.Choleraesuis strain SC-B67 lacks the eutK, eutC, and eutA,these genes are present in the ATCC strain used here, asdemonstrated by PCR (data not shown). The phenotypes of allserovars and mutants mentioned above were further confirmedin growth assays using MM supplemented with cyanocobal-amin and 1,2-propanediol or ethanolamine (data not shown).
Differential expression of genes involved in propanediol andethanolamine utilization. According to the literature, the pduoperon has a single transcription start site upstream of pduA,and pduF and pocR are controlled by their own promoters (13);the eut operon has a main promoter located upstream of eutSand a minor promoter upstream of eutR (29) (Fig. 1). Frag-ments 250 bp upstream of pduA, pduF, pocR, and eutS, whichwere computationally confirmed to possess promoter regions,
FIG. 1. Genetic organization of the eut, pdu, and cob-cbi genes in S. Typhimurium LT2. The genomes of the S. enterica serovars tested (Table1) carry intact operons with most of the genes homologous to each other (filled arrows). The cob and pdu operons in the genome of S. Gallinarumstrain 287/91 lack the cbiOKDC and pocR genes (open arrows). The organization of the eut operon within most Salmonella serovars is also shown.An exception is the eut operon of S. Choleraesuis strain SC-B67, lacking eutACK. Shaded arrows indicate gene replacement with respect to theLT2 genome.
284 SRIKUMAR AND FUCHS APPL. ENVIRON. MICROBIOL.
on June 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
were cloned upstream of the luxCDABE cassette in plasmidpDEW201, and the resulting constructs were transformed into14028. Strains with plasmids carrying the argS promoter or anintergenic fragment of STM0047 (�STM0047�) without pro-moter homology served as the positive and negative controls.The probes were then inoculated into growth medium or food,and promoter expression was determined as RLU/OD600.
All promoters except the positive control were transcription-ally inactive in MM containing glucose. Therefore, the foldinduction of promoters tested in the presence of other carbonsources was correlated to the light emission values measured inglucose-containing medium. The induction of PpduA was 241-
fold higher during growth with 1,2-propanediol than thethreshold level in medium with glucose, thus quantifyingformer qualitative data (7). The addition of rhamnose andfucose induced the pdu operon 27-fold and 14-fold, respec-tively (Table 3). When tested with arabinose and sucrose inMM, the induction of PpduA was only slightly above the thresh-old (data not shown). To determine whether rhamnose or itsfermentation products act as PpduA inducers, rhamnose metab-olism was interrupted by the deletion of rhaS and rhaR. PpduA
did not show significant transcriptional activity in mutant14028�rhaSR cultivated in M9 containing 1% rhamnose and0.1% yeast extract (data not shown).
The promoters of pduF and pocR, whose products are in-volved in propanediol transport and in the regulation of thepdu operon, were found to be upregulated 3-fold and 2-fold,respectively, in medium with 1,2-propanediol. The latter find-ing probably reflects the pocR autoinduction (7). No significantinduction of PpduF and PpocR in the presence of rhamnose andfucose was observed. Growth in MM with 1,2-propanediol ledto a 21-fold induction of the transcriptional activity of the mainpromoter for the eut operon, PeutS, possibly due to CsrA-dependent regulation of both pathways (3). In MM with glu-cose, no signal above the threshold set by the negative controlwas observed for PeutS, while the signal with the positive con-trol, PargS, was approximately 140-fold upregulated (Table 4).
FIG. 2. Growth on MacConkey agar base supplemented with 1%propanediol or ethanolamine with or without cyanocobalamin.(A) Colonies of strain 14028 appear dark pink on MacConkey agarcontaining propanediol or ethanolamine. (B) 14028�pocR on pro-panediol-containing MacConkey agar or 14028�eutR on ethanol-amine-containing MacConkey agar shows yellow colonies, since thesestrains do not metabolize the respective substrates. (C) S. Gallinarum287/91 on ethanolamine-containing MacConkey agar exhibits pink col-onies. (D) pdu-deficient S. Gallinarum strain 287/91 on MacConkeyagar with propanediol exhibits yellow colonies (D).
TABLE 3. Quantification of promoter activities in 1,2-propanediol, rhamnose, and fucose
Fragment cloned intopDEW201
Promoter activitya in 14028 grown in:
M9 � 1,2-propanediol M9 � rhamnose M9 � fucose
RLU/OD600 SD (%) Foldinduction RLU/OD600 SD (%) Fold
induction RLU/OD600 SD (%) Foldinduction
PpduA 6.5 � 105 5.5 241 7.2 � 104 14.4 26.7 3.9 � 104 10.8 14.4PeutS 1.8 � 105 4.1 21.2 1.6 � 104 11.7 1.9 2.4 � 104 13.1 2.8PpduF 2.4 � 103 12.2 3.1 1.1 � 103 3.1 1.4 1.2 � 103 17.1 1.6PpocR 1.1 � 103 13.9 2.1 8.0 � 102 16.4 1.5 7.7 � 102 30.4 1.5PargS
b 3.6 � 105 4.6 0.5 8.6 � 105 1.8 1.2 1.0 � 106 1.2 1.4�STM0047�c 5.3 � 103 6.8 1 5.3 � 103 5.3 1.0 7.4 � 103 1.2 1.4
a Gene expression was quantified as relative light units per optical density at 600 nm (RLU/OD600). The highest expression values were taken. Average data fromthree independent experiments are shown. Fold induction was calculated with respect to the RLU/OD600 values of strain 14028 grown in glucose.
b Positive control.c Negative control.
TABLE 4. Quantification of promoter activities in the presenceof MM with glucose or ethanolamine
Fragmentcloned intopDEW201
Promoter activitya in 14028 grown in:
M9 � glucose M9 � ethanolamine
RLU/OD600 SD (%) RLU/OD600 SD (%) Foldinduction
PpduA 2.7 � 103 20 6.5 � 104 9 24.1PeutS 8.5 � 103 10.3 1.8 � 107 3.4 2118PpduF 7.7 � 102 19 7.7 � 104 13 100PpocR 5.2 � 102 42 9.3 � 103 12.6 17.8PargS 7.4 � 105 5.2 3.4 � 106 9 4.6�STM0047� 5.3 � 103 4.4 1.5 � 104 13 2.8
a Gene expression was quantified as relative light units per optical density at600 nm (RLU/OD600). The highest expression values were taken. Average datafrom three independent experiments are shown. Fold induction was calculatedwith respect to the RLU/OD600 values of strain 14028 grown in glucose.
VOL. 77, 2011 ETHANOLAMINE UTILIZATION BY SALMONELLA ENTERICA 285
on June 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

In MM containing ethanolamine, high transcriptional activityof PeutS during the growth of strain 14028 was observed in thelate-logarithmic phase in comparison to that for the controlfragment �STM0047� and PargS. Standardization to the data ofthe control experiment with glucose revealed that these valuescorrespond to a 2,118-fold induction of PeutS, but only to 4.6-fold- and 2.4-fold-enhanced transcriptional activities of PargS
and �STM0047�, respectively (Table 4).14028�eutR neither transcriptionally activates the eut
operon and nor grows with ethanolamine. As a control, plas-mid pDEW-PeutS was therefore transformed into 14028�eutRand investigated for promoter induction in ethanolamine-con-taining MM. The RLU/OD600 values of 6.4 � 102 were similarto the light emission by the negative control, thus confirmingthe role of EutR as an essential inductor of the eut operon.
The data presented above show the specific induction of thecloned promoters in response to carbon sources added to themedium. They also quantify the response of PpduA and PeutS tothe presence of 1,2-propanediol or ethanolamine in the ab-sence of glucose, resulting in an increase in pdu or eut genetranscription by as much as 2 to 3 orders of magnitude at theend of the logarithmic phase.
Induction of the eut operon in milk and egg yolk. The im-pacts of raw milk and egg on the expression of the eut operonwere then tested using plasmids pDEW-PeutS, pDEW-PargS,and pDEW-�STM0047�. To this end, the strains harboring thepromoter fusions were directly inoculated into raw milk, andthe bioluminescence of the three constructs was measured.During the growth of S. Typhimurium in milk, the activity ofPeutS reached a maximum of 1.5 � 106 RLU/OD600 (Fig. 3A).This is approximately two times higher than the biolumines-cence observed with PargS controlling a housekeeping gene and428 times higher than the background expression from thenegative-control sequence without promoter homology(�STM0047�). To determine which factor in milk specificallyinduced the operon, the constructs were further tested in thepresence of three milk fractions, namely a lipid-rich milk fatglobule fraction, a protein-rich casein macropeptide (CMP)fraction, and an ultrafiltrate permeate fraction rich in carbo-hydrates. We observed that the lipid-rich milk fat fraction (1%in MM) upregulated the eut operon to 11-fold-higher activityof PeutS in comparison to �STM0047� (Table 5). In contrast,only a background emission level of PeutS, similar to that of thenegative control, was detected when S. Typhimurium wasgrown in MM with 1% of a protein-rich CMP fraction, or withan ultrafiltrate permeate fraction lacking CMP and the milk fatglobule membrane material (data not shown). The strain withpDEW-PeutS was also inoculated into milk with different con-centrations of fat. While the milk rich in fat (3.5%) induced theeut operon 428-fold with respect to the negative control, themilk with less fat—0.1 or 1.5%—activated this operon by afactor of 62 or 46, respectively (Table 5).
This induction of the eut operon in the presence of lipid-richfood fractions was further investigated using pure egg yolkcontaining 80% lipids, of which as much as 30% is phospho-lipids (31). Under these conditions, a 139-fold induction ofPeutS with respect to the control fragment �STM0047� wasdetermined (Fig. 3B). During growth in both milk and eggyolk, the induction of the eut operon started approximately 4 hafter the inoculation of 14028 into the food medium, and the
promoter reached its maximal activity approximately 5 h later(Fig. 3A and B). The pdu operon did not show any upregula-tion in the different milk fractions. The specific induction ofPeutS during S. Typhimurium growth in milk and egg yolk wasalso monitored with a Xenogen IVIS Lumina station (Fig. 3C).
Growth deficiency of mutant 14028�eutR in milk and eggyolk. The high and specific induction of PeutS in the presence ofmilk and egg yolk suggests that the eut operon contributes tothe replication of S. Typhimurium under these conditions. Wetherefore constructed the mutant 14028�eutR, which is unableto grow in a minimal medium with ethanolamine as the onlysource of carbon and energy (data not shown).
The bacterial proliferation of strains 14028 and 14028�eutRspiked into sterile milk was monitored over a period of 24 h at37°C by enumerating the bacteria at several time points. Fourand 6 h after inoculation, 2 � 106 and 3.7 � 107 CFU/ml ofmutant 14028�eutR, but 6.6 � 106 and 1.1 � 108 CFU/ml ofstrain 14028, were counted (Fig. 4A). A significantly reducedcell number of mutant 14028�eutR was still observed after 8 h,but not after 10 h or later, when cells reached the stationaryphase.
A correlation between ethanolamine operon induction andthe proliferation of S. Typhimurium in food was further exam-ined using egg yolk as the growth medium. The data obtainedwere similar to those from the milk experiment. Four and 8 hafter inoculation, mutant 14028�eutR exhibited 2.1 � 107 and3.5 � 108 CFU/ml, in comparison to 4.7 � 107 and 6.5 � 108
CFU/ml obtained with strain 14028 (Fig. 4B). This 2-fold re-duction in the mutant cell counts obliterated gradually to anequal cell number of the two strains 10 h postinfection. Thereduced proliferation of 14028�eutR both in milk and in eggyolk fits well with the pattern of PeutS expression, showing thatthe ethanolamine operon is induced 4 to 6 h after the entry ofthe bacterium into the food and that this induction lasts ap-proximately 10 h. These results, in combination with the ex-pression data, demonstrate that the ethanolamine utilizationpathway not only is stimulated during the initial hours of pro-liferation within milk and egg yolk but also contributes to theproliferation of S. Typhimurium in food.
Impact of ethanolamine utilization for proliferation of S.Typhimurium within C. elegans. The relevance of PocR andEutR, and thus of ethanolamine and propanediol utilization, invivo was tested by comparing the bacterial proliferation of therespective mutants with that of strain 14028 following C. el-egans infection. Both mutants 14028�eutR and 14028�pocRexhibited a compromised ability to proliferate within nema-todes compared with that of strain 14028 (Fig. 5). Twelve hourspostinfection, 1.9 � 103 CFU/worm of strain 14028, but only1.1 � 103 CFU/worm and 1.2 � 103 CFU/worm of 14028�eutRand 14928�pocR, respectively, were obtained, correspondingto a 1.7- and 1.6-fold reduction in the 14028�eutR and14028�pocR counts in comparison with those of strain 14028.This growth attenuation increased considerably during the nexthours of infection. Twenty-four hours after C. elegans wormswere fed on Salmonella strains, a 2.4-fold-lower number ofviable 14028�eutR cells than of viable strain 14028 cells wasdetermined. 14028�pocR even showed 8.6-fold-weaker growththan strain 14028. The measurements after 36 and 48 h re-vealed 1.8-fold and 2.3-fold reductions of 14028�eutR and14028�pocR cell numbers from those of the control. Thus,
286 SRIKUMAR AND FUCHS APPL. ENVIRON. MICROBIOL.
on June 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

FIG. 3. Luciferase-based expression profiling of the eut and pdu operons in milk and egg yolk. (A) The transcriptional activity of PeutS in milkwas 428-fold higher than the background expression of the negative control (�STM0047�) and 2-fold higher than that of the positive control (PargS).Bars represent numbers of viable cells of 14028/pDEW-�STM0047�, expressed in CFU/ml. (B) Expression profile of PeutS in egg yolk. During thegrowth of 14028 in egg yolk, the operon was upregulated to a level 139-fold higher than that for �STM0047�. PargS served as the positive control.(C) PeutS expression in pasteurized milk and egg yolk. PargS and �STM0047� served as positive and negative controls, respectively. PeutS is highlyexpressed in pasteurized milk and egg yolk due to the high content of phospholipids. Photographs were made using an IVIS Lumina system witha charge-coupled device (CCD) camera (Xenogen, Alameda, CA).
VOL. 77, 2011 ETHANOLAMINE UTILIZATION BY SALMONELLA ENTERICA 287
on June 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

both PocR and EutR are required for wild-type-like growth ofS. Typhimurium during infection of C. elegans.
DISCUSSION
S. enterica is widely known to utilize 1,2-propanediol andethanolamine as the sole sources of carbon and energy, butvery little is known about the impact of these metabolic path-ways on the replication of the pathogen under natural condi-tions. 1,2-Propanediol and, indirectly, sugars such as rhamnoseand fucose, known as sources of 1,2-propanediol (5), weredemonstrated here to upregulate the pdu operon significantlyand specifically. The highest induction, by more than 3 ordersof magnitude, was measured for PeutS in the presence of eth-anolamine.
About 95% of cases of human salmonellosis are attributed
TABLE 5. Quantification of promoter activities in the presence of MM with CMP or the fat fraction of milk
Fragmentcloned intopDEW201
Promoter activitya in 14028 grown in:
M9 � milk CMP fraction (1%) M9 � milk fat fraction (1%) Pasteurized milk with 0.1%/1.5% fat Pasteurized milk with 3.5% fat
RLU/OD600 SD (%) Foldinduction RLU/OD600 SD (%) Fold
induction RLU/OD600 SD (%) Foldinduction RLU/OD600 SD (%) Fold
induction
PpduA BT BT 1.8 � 102 28 0.2PeutS 1.65 � 102 13 1.0 8.4 � 104 7.3 11 2.2 � 105/1.6 � 105 5/5 62/46 1.5 � 106 3.6 428PpduF BT BT 1.4 � 102 22 0.2PpocR BT BT BT BTPargS
b 5.7 � 103 0.6 36 8.2 � 104 0.9 103�STM0047�c 1.6 � 102 20 1 8.0 � 102 7 1
a Fold induction was calculated with respect to the RLU/OD600 values of negative-control strain pDEW-�STM0047�. CMP, casein macropeptide. BT, below thethreshold set by the medium control.
b Positive control.c Negative control.
FIG. 4. Contribution of ethanolamine degradation to S. Typhi-murium proliferation in food. (A) Replication of strains 14028 and14028�eutR in milk. 14028�eutR exhibited a 3-fold lower cell numberthan 14028 after 4 to 6 h, a pattern that parallels the strongest etha-nolamine expression. (B) 14028�eutR showed a 2-fold lower cell num-ber than 14028 during growth in egg yolk. Asterisks indicate timepoints at which the difference between the values for 14028 and14028�eutR was statistically significant as determined by Student’s ttest.
FIG. 5. Bacterial proliferation of strains 14028, 14028�pocR, and14028�eutR in C. elegans. Strain 14028 showed 2.4-fold-increased pro-liferation relative to that of 14028�eutR and a cell number 8.6-foldhigher than that of 14028�pocR. Average data from three independentreactions are shown. Asterisks indicate time points at which statisti-cally significant differences (according to Student’s t test) between theexperiments with strain 14028 and its mutants were revealed. A total of20 nematodes were used for each data point.
288 SRIKUMAR AND FUCHS APPL. ENVIRON. MICROBIOL.
on June 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

to contaminated food products such as meat, poultry, eggs, andmilk (18). Interestingly, raw milk and egg yolk were shown hereto strongly upregulate the eut operon, but not the pdu operon.On average, bovine milk contains about 33 g of total lipid perliter. This includes a variety of phospholipids derived from themammary plasma membrane surrounding each milk fat glob-ule (20). Phosphatidylethanolamine is the second most abun-dant phospholipid in mammalian cell membranes, constituting20% to 50% of the total phospholipids (59). Further investi-gation of eut induction in a casein macropepetide (CMP) frac-tion and the fat fraction of milk revealed that the fat fraction,with the higher phospholipid content, but not the CMP frac-tion, induced the eut operon. A 139-fold induction in egg yolkconfirms that eut operon induction in Salmonella is specificallyhigh in food materials rich in lipids. Indeed, in both raw milkand egg yolk, the proliferation of the mutant 14028�eutR,which is unable to utilize ethanolamine, was significantly re-tarded up to 10 h after inoculation. This is in line with theobservation that cells preadapted by overnight growth in MMwith ethanolamine exhibited PeutS induction as early as 2 hpostinoculation (data not shown). This provides evidence thatthe utilization of phosphatidylethanolamine from phospholip-ids plays a neglected role during the proliferation of S. Typhi-murium in foods moderately rich in fat.
C. elegans infection is a feasible model for the study ofSalmonella pathogenesis (56). Different strains of S. Typhi-murium, including SL1344 and 14028, had similar prolifer-ation patterns within C. elegans, and the bacterium killed theworms with a TD50 (time for 50% of the nematodes to die)of 5.1 � 0.7 days (1). The C. elegans model was used here toevaluate the importance of ethanolamine and propanediolutilization during the proliferation of Salmonella within thenematode. Our experiments demonstrated that 14028�eutRand 14028�pocR are significantly growth deficient within theworm compared with strain 14028. The lysis of E. coli OP50cells by the nematode probably results in the accumulationof bacterial phospholipids and polysaccharides in the C.elegans intestine. These substrates and their degradationproducts might induce the ethanolamine and/or propanediolpathways of S. enterica strains that have successfully colo-nized the nematode, allowing this pathogen to proliferatefurther within the worm. Mutants incapable of utilizingthose substrates have a significant growth disadvantage dur-ing proliferation within nematodes. Interestingly, an in-creasing number of recent reports provide further evidencethat the functionally coupled eut, pdu, and cob operonscontribute to the replication of pathogenic bacteria in eu-karyotic hosts, as reviewed recently (19). Examples, in ad-dition to data from mouse infection or the cell culture assaysmentioned above, include the induction of the Photorhabdusluminescens eut genes in insects (40) and the observationthat an eut mutant of Enterococcus faecalis is attenuated inthe killing of C. elegans (19). Moreover, tetrathionate, whichacts as a terminal electron acceptor during the anaerobicdegradation of 1,2-propanediol, is formed in the inflamedgut upon infection (62).
Since the cob and pdu operons constitute a single reguloncontrolled by PocR, the possibility cannot be excluded that theproliferation phenotype of 14028�pocR is caused by the lack ofde novo cobalamin synthesis, thus reflecting the attenuated
growth of 14028�eutR, which is unable to use ethanolamine ina cobalamin-dependent manner. However, nematode growthmedium (NGM) harbors traces of vitamin B12. Furthermore, ithas been shown previously that pdu genes are essential for thereplication of Salmonella in macrophages (28), suggesting arole for ethanolamine and propanediol degradation in vivo.
Taken together, these findings point to an overlooked con-tribution of ethanolamine and propanediol utilization to theproliferation of S. Typhimurium in soil organisms, in food, andin different mammalian compartments. Thus, both metabolictraits provide a selection advantage for the food-borne patho-gen not only in a single step of infection, but over all stages ofthe infection chain, and might therefore constitute a promisingnew set of possible targets for food formulas that suppressbacterial growth (9, 37).
ACKNOWLEDGMENTS
We thank Siegfried Scherer for financial support. The BayerischeForschungsstiftung is gratefully acknowledged for a postdoctoral sti-pendium to S.S.
We thank Ulrich Kulozik for helpful discussions, Nadja Siegert forproviding CMP and milk fat fractions, Patrick Schiwek for technicalassistance, and Nicholas Thomson for supplying S. Gallinarum strain287/91.
REFERENCES
1. Aballay, A., P. Yorgey, and F. M. Ausubel. 2000. Salmonella typhimuriumproliferates and establishes a persistent infection in the intestine of Caeno-rhabditis elegans. Curr. Biol. 10:1539–1542.
2. Adkins, J. N., H. M. Mottaz, A. D. Norbeck, J. K. Gustin, J. Rue, T. R.Clauss, S. O. Purvine, K. D. Rodland, F. Heffron, and R. D. Smith. 2006.Analysis of the Salmonella typhimurium proteome through environmentalresponse toward infectious conditions. Mol. Cell. Proteomics 5:1450–1461.
3. Ailion, M., T. A. Bobik, and J. R. Roth. 1993. Two global regulatory systems(Crp and Arc) control the cobalamin/propanediol regulon of Salmonellatyphimurium. J. Bacteriol. 175:7200–7208.
4. Altschul, S. F., T. L. Madden, A. A. Schaffer, J. Zhang, Z. Zhang, W. Miller,and D. J. Lipman. 1997. Gapped BLAST and PSI-BLAST: a new generationof protein database search programs. Nucleic Acids Res. 25:3389–3402.
5. Badia, J., J. Ros, and J. Aguilar. 1985. Fermentation mechanism of fucoseand rhamnose in Salmonella typhimurium and Klebsiella pneumoniae. J. Bac-teriol. 161:435–437.
6. Blackwell, C. M., F. A. Scarlett, and J. M. Turner. 1976. Ethanolaminecatabolism by bacteria, including Escherichia coli. Biochem. Soc. Trans.4:495–497.
7. Bobik, T. A., M. Ailion, and J. R. Roth. 1992. A single regulatory geneintegrates control of vitamin B12 synthesis and propanediol degradation. J.Bacteriol. 174:2253–2266.
8. Bobik, T. A., Y. Xu, R. M. Jeter, K. E. Otto, and J. R. Roth. 1997. Pro-panediol utilization genes (pdu) of Salmonella typhimurium: three genes forthe propanediol dehydratase. J. Bacteriol. 179:6633–6639.
9. Boigegrain, R. A., J. P. Liautard, and S. Kohler. 2005. Targeting of thevirulence factor acetohydroxyacid synthase by sulfonylureas results in inhi-bition of intramacrophagic multiplication of Brucella suis. Antimicrob.Agents Chemother. 49:3922–3925.
10. Braden, C. R. 2006. Salmonella enterica serotype Enteritidis and eggs: anational epidemic in the United States. Clin. Infect. Dis. 43:512–517.
11. Brinsmade, S. R., T. Paldon, and J. C. Escalante-Semerena. 2005. Minimalfunctions and physiological conditions required for growth of Salmonellaenterica on ethanolamine in the absence of the metabolosome. J. Bacteriol.187:8039–8046.
12. Chang, G. W., and J. T. Chang. 1975. Evidence for the B12-dependentenzyme ethanolamine deaminase in Salmonella. Nature 254:150–151.
13. Chen, P., M. Ailion, T. Bobik, G. Stormo, and J. Roth. 1995. Five promotersintegrate control of the cob/pdu regulon in Salmonella typhimurium. J. Bac-teriol. 177:5401–5410.
14. Conner, C. P., D. M. Heithoff, S. M. Julio, R. L. Sinsheimer, and M. J.Mahan. 1998. Differential patterns of acquired virulence genes distinguishSalmonella strains. Proc. Natl. Acad. Sci. U. S. A. 95:4641–4645.
15. Datsenko, K. A., and B. L. Wanner. 2000. One-step inactivation of chromo-somal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad.Sci. U. S. A. 97:6640–6645.
16. De Buyser, M. L., B. Dufour, M. Maire, and V. Lafarge. 2001. Implication ofmilk and milk products in food-borne diseases in France and in differentindustrialised countries. Int. J. Food Microbiol. 67:1–17.
VOL. 77, 2011 ETHANOLAMINE UTILIZATION BY SALMONELLA ENTERICA 289
on June 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

17. Del Papa, M. F., and M. Perego. 2008. Ethanolamine activates a sensorhistidine kinase regulating its utilization in Enterococcus faecalis. J. Bacteriol.190:7147–7156.
18. Foley, S. L., A. M. Lynne, and R. Nayak. 2009. Molecular typing methodol-ogies for microbial source tracking and epidemiological investigations ofGram-negative bacterial foodborne pathogens. Infect. Genet. Evol. 9:430–440.
19. Garsin, D. A. 2010. Ethanolamine utilization in bacterial pathogens: rolesand regulation. Nat. Rev. Microbiol. 8:290–295.
20. German, J. B., R. A. Gibson, R. M. Krauss, P. Nestel, B. Lamarche, W. A.van Staveren, J. M. Steijns, L. C. de Groot, A. L. Lock, and F. Destaillats.2009. A reappraisal of the impact of dairy foods and milk fat on cardiovas-cular disease risk. Eur. J. Nutr. 48:191–203.
21. Gillespie, I. A., G. K. Adak, S. J. O’Brien, and F. J. Bolton. 2003. Milkbornegeneral outbreaks of infectious intestinal disease, England and Wales, 1992–2000. Epidemiol. Infect. 130:461–468.
22. Heithoff, D. M., C. P. Conner, U. Hentschel, F. Govantes, P. C. Hanna, andM. J. Mahan. 1999. Coordinate intracellular expression of Salmonella genesinduced during infection. J. Bacteriol. 181:799–807.
23. Jayarao, B. M., S. C. Donaldson, B. A. Straley, A. A. Sawant, N. V. Hegde,and J. L. Brown. 2006. A survey of foodborne pathogens in bulk tank milkand raw milk consumption among farm families in Pennsylvania. J. Dairy Sci.89:2451–2458.
24. Jayarao, B. M., and D. R. Henning. 2001. Prevalence of foodborne patho-gens in bulk tank milk. J. Dairy Sci. 84:2157–2162.
25. Jeter, R. M. 1990. Cobalamin-dependent 1,2-propanediol utilization by Sal-monella typhimurium. J. Gen. Microbiol. 136:887–896.
26. Joseph, B., K. Przybilla, C. Stuhler, K. Schauer, J. Slaghuis, T. M. Fuchs,and W. Goebel. 2006. Identification of Listeria monocytogenes genes contrib-uting to intracellular replication by expression profiling and mutant screen-ing. J. Bacteriol. 188:556–568.
27. Kelly, A., M. D. Goldberg, R. K. Carroll, V. Danino, J. C. Hinton, and C. J.Dorman. 2004. A global role for Fis in the transcriptional control of metab-olism and type III secretion in Salmonella enterica serovar Typhimurium.Microbiology 150:2037–2053.
28. Klumpp, J., and T. M. Fuchs. 2007. Identification of novel genes in genomicislands that contribute to Salmonella typhimurium replication in macro-phages. Microbiology 153:1207–1220.
29. Kofoid, E., C. Rappleye, I. Stojiljkovic, and J. Roth. 1999. The 17-geneethanolamine (eut) operon of Salmonella typhimurium encodes five homo-logues of carboxysome shell proteins. J. Bacteriol. 181:5317–5329.
30. Korbel, J. O., T. Doerks, L. J. Jensen, C. Perez-Iratxeta, S. Kaczanowski,S. D. Hooper, M. A. Andrade, and P. Bork. 2005. Systematic association ofgenes to phenotypes by genome and literature mining. PLoS Biol. 3:e134.
31. Kovacs-Nolan, J., M. Phillips, and Y. Mine. 2005. Advances in the value ofeggs and egg components for human health. J. Agric. Food Chem. 53:8421–8431.
32. Kroger, C., and T. M. Fuchs. 2009. Characterization of the myo-inositolutilization island of Salmonella enterica serovar Typhimurium. J. Bacteriol.191:545–554.
33. Larson, T. J., M. Ehrmann, and W. Boos. 1983. Periplasmic glycerophos-phodiester phosphodiesterase of Escherichia coli, a new enzyme of the glpregulon. J. Biol. Chem. 258:5428–5432.
34. Lawhon, S. D., J. G. Frye, M. Suyemoto, S. Porwollik, M. McClelland, andC. Altier. 2003. Global regulation by CsrA in Salmonella typhimurium. Mol.Microbiol. 48:1633–1645.
35. Lawrence, J. G., and J. R. Roth. 1996. Evolution of coenzyme B12 synthesisamong enteric bacteria: evidence for loss and reacquisition of a multigenecomplex. Genetics 142:11–24.
36. Lewis, J. A., and J. T. Fleming. 1995. Basic culture methods, p. 3–29. In H. F.Epstein and D. C. Shakes (ed.), Caenorhabditis elegans: modern biologicalanalysis of an organism. Academic Press, New York, NY.
37. Liautard, J. P., V. Jubier-Maurin, R. A. Boigegrain, and S. Kohler. 2006.Antimicrobials: targeting virulence genes necessary for intracellular multi-plication. Trends Microbiol. 14:109–113.
38. Link, A. J., D. Phillips, and G. M. Church. 1997. Methods for generatingprecise deletions and insertions in the genome of wild-type Escherichia coli:application to open reading frame characterization. J. Bacteriol. 179:6228–6237.
39. Mead, P. S., L. Slutsker, V. Dietz, L. F. McCaig, J. S. Bresee, C. Shapiro,P. M. Griffin, and R. V. Tauxe. 1999. Food-related illness and death in theUnited States. Emerg. Infect. Dis. 5:607–625.
40. Munch, A., L. Stingl, K. Jung, and R. Heermann. 2008. Photorhabdus lumi-nescens genes induced upon insect infection. BMC Genomics 9:229.
41. Oliver, S. P., B. M. Jayarao, and R. A. Almeida. 2005. Foodborne pathogens
in milk and the dairy farm environment: food safety and public healthimplications. Foodborne Pathog. Dis. 2:115–129.
42. Penrod, J. T., and J. R. Roth. 2006. Conserving a volatile metabolite: a rolefor carboxysome-like organelles in Salmonella enterica. J. Bacteriol. 188:2865–2874.
43. Price-Carter, M., J. Tingey, T. A. Bobik, and J. R. Roth. 2001. The alterna-tive electron acceptor tetrathionate supports B12-dependent anaerobicgrowth of Salmonella enterica serovar Typhimurium on ethanolamine or1,2-propanediol. J. Bacteriol. 183:2463–2475.
44. Proulx, P., and C. K. Fung. 1969. Metabolism of phosphoglycerides in E. coli.IV. The positional specificity and properties of phospholipase A. Can. J. Bio-chem. 47:1125–1128.
45. Reed, B. A., and L. E. Grivetti. 2000. Controlling on-farm inventories ofbulk-tank raw milk: an opportunity to protect public health. J. Dairy Sci.83:2988–2991.
46. Rondon, M. R., and J. C. Escalante-Semerena. 1996. In vitro analysis of theinteractions between the PocR regulatory protein and the promoter regionof the cobalamin biosynthetic (cob) operon of Salmonella typhimurium LT2.J. Bacteriol. 178:2196–2203.
47. Roof, D. M., and J. R. Roth. 1992. Autogenous regulation of ethanolamineutilization by a transcriptional activator of the eut operon in Salmonellatyphimurium. J. Bacteriol. 174:6634–6643.
48. Roof, D. M., and J. R. Roth. 1988. Ethanolamine utilization in Salmonellatyphimurium. J. Bacteriol. 170:3855–3863.
49. Roof, D. M., and J. R. Roth. 1989. Functions required for vitamin B12-dependent ethanolamine utilization in Salmonella typhimurium. J. Bacteriol.171:3316–3323.
50. Roth, J. R., J. G. Lawrence, and T. A. Bobik. 1996. Cobalamin (coenzymeB12): synthesis and biological significance. Annu. Rev. Microbiol. 50:137–181.
51. Sambrook, J., and D. W. Russell. 2001. Molecular cloning: a laboratorymanual, 3rd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
52. Sonck, K. A., G. Kint, G. Schoofs, C. Vander Wauven, J. Vanderleyden, andS. C. De Keersmaecker. 2009. The proteome of Salmonella Typhimuriumgrown under in vivo-mimicking conditions. Proteomics 9:565–579.
53. Spanier, B., M. Starke, F. Higel, S. Scherer, and T. M. Fuchs. 2010. Yersiniaenterocolitica infection and tcaA-dependent killing of Caenorhabditis elegans.Appl. Environ. Microbiol. 76:6277–6285.
54. Stiernagle, T. 11 February 2006. Maintenance of C. elegans. In The C. elegansResearch Community (ed.). WormBook. doi/10.1895/wormbook.1.101.1.http://www.wormbook.org.
55. Stojiljkovic, I., A. J. Baumler, and F. Heffron. 1995. Ethanolamine utilizationin Salmonella typhimurium: nucleotide sequence, protein expression, andmutational analysis of the cchA cchB eutE eutJ eutG eutH gene cluster. J.Bacteriol. 177:1357–1366.
56. Tenor, J. L., B. A. McCormick, F. M. Ausubel, and A. Aballay. 2004. Cae-norhabditis elegans-based screen identifies Salmonella virulence factors re-quired for conserved host-pathogen interactions. Curr. Biol. 14:1018–1024.
57. Thomson, N. R., D. J. Clayton, D. Windhorst, G. Vernikos, S. Davidson, C.Churcher, M. A. Quail, M. Stevens, M. A. Jones, M. Watson, A. Barron, A.Layton, D. Pickard, R. A. Kingsley, A. Bignell, L. Clark, B. Harris, D.Ormond, Z. Abdellah, K. Brooks, I. Cherevach, T. Chillingworth, J. Wood-ward, H. Norberczak, A. Lord, C. Arrowsmith, K. Jagels, S. Moule, K.Mungall, M. Sanders, S. Whitehead, J. A. Chabalgoity, D. Maskell, T.Humphrey, M. Roberts, P. A. Barrow, G. Dougan, and J. Parkhill. 2008.Comparative genome analysis of Salmonella Enteritidis PT4 and SalmonellaGallinarum 287/91 provides insights into evolutionary and host adaptationpathways. Genome Res. 18:1624–1637.
58. Toledo-Arana, A., O. Dussurget, G. Nikitas, N. Sesto, H. Guet-Revillet, D.Balestrino, E. Loh, J. Gripenland, T. Tiensuu, K. Vaitkevicius, M. Bar-thelemy, M. Vergassola, M. A. Nahori, G. Soubigou, B. Regnault, J. Y.Coppee, M. Lecuit, J. Johansson, and P. Cossart. 2009. The Listeria tran-scriptional landscape from saprophytism to virulence. Nature 459:950–956.
59. Vance, J. E. 2008. Phosphatidylserine and phosphatidylethanolamine inmammalian cells: two metabolically related aminophospholipids. J. LipidRes. 49:1377–1387.
60. Van Dyk, T. K., and R. A. Rosson. 1998. Photorhabdus luminescens lux-CDABE promoter probe vectors. Methods Mol. Biol. 102:85–95.
61. Walter, D., M. Ailion, and J. Roth. 1997. Genetic characterization of the pduoperon: use of 1,2-propanediol in Salmonella typhimurium. J. Bacteriol. 179:1013–1022.
62. Winter, S. E., P. Thiennimitr, M. G. Winter, B. P. Butler, D. L. Huseby, R. W.Crawford, J. M. Russell, C. L. Bevins, L. G. Adams, R. M. Tsolis, J. R. Roth,and A. J. Baumler. 2010. Gut inflammation provides a respiratory electronacceptor for Salmonella. Nature 467:426–429.
290 SRIKUMAR AND FUCHS APPL. ENVIRON. MICROBIOL.
on June 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from





























