Embed Size (px)
Citation preview

Proc. Nail. Acad. Sci. USAVol. 81, pp. 1322-1326, March 1984Biochemistry
Estradiol feedback effects on the a-subunit mRNA in the sheeppituitary gland: Correlation with serum and pituitaryluteinizing hormone concentrations
(gonadotropin/steroid hormone/radioimmunoassay)
THOMAS LANDEFELD*t, JADWIGA KEPA*, AND FRED KARSCHtDepartments of *Pharmacology, tPathology, and WPhysiology, Reproductive Endocrinology Program, The University of Michigan, Ann Arbor, MI 48109-0010
Communicated by M. J. Coon, November 10, 1983
ABSTRACT The effects of estradiol feedback on pituitaryluteinizing hormone (LH) content, serum LH concentration,and in vitro-translated a subunit was examined in the ewe.Three animal models were used representing positive, nega-tive, and no estradiol feedback. Two experiments were carriedout: (i) anestrous ewes were treated acutely with five Silasticestradiol implants to induce a LH surge (positive feedback)and (it) ovariectomized ewes were treated chronically with anestradiol implant (negative feedback) or were not treated (nofeedback). Pituitary RNA was prepared and translated in acell-free system; the a subunit was identified by immuno-precipitation and NaDodSO4/PAGE. cpm/pug of RNA andimmunoprecipitated growth hormone and prolactin were usedto evaluate possible differences in RNA translational efficien-cies among the treatment groups. In experiment 1, significant-ly higher amounts of the a subunit were observed in animalsexhibiting an estradiol-induced LH surge than in normal anes-trous ewes (P < 0.001). Examination of values from individualanimals suggested a correlation between the stage of the LHsurge, pituitary LH, and translated a subunit. In experiment2, the amount of a subunit observed in animals exposed tochronic estradiol negative feedback was significantly less (P <0.005) than that in the untreated ovariectomized animals (nofeedback) and no different from that in intact anestrous ewes.These results suggest that both the negative and the positivefeedback effects of estradiol include regulation of the amountof a-subunit mRNA.
Despite extensive investigation of the events associated withthe positive and negative feedback effects of estradiol on go-nadotropin secretion, the contribution of gonadotropin sub-unit biosynthesis to these reproductive phenomena has notbeen established. For example, it is not known whether thegonadotropin surge produced by an acute increase in estradi-ol is due primarily to an increase in release of newly synthe-sized gonadotropin, an increase in the release of stored hor-mone, or a combination of both processes. In addition, if anincrease in synthesis does occur, it is not known whether itinvolves both subunits or only one. As for the negative feed-back effects of estradiol, the same questions need to be ad-dressed relative to decreased secretion of gonadotropins. Al-though there are recent studies of the effects of negative es-tradiol feedback on a- and ,-subunit mRNAs in castratedsheep (1, 2), similar studies relative to positive feedback ef-fects of estradiol have not been reported.To investigate the events associated with the stimulatory
and inhibitory effects of estradiol, we have evaluated serumluteinizing hormone (LH), pituitary LH, and in vitro synthe-sis of a subunit in anestrous ewes during conditions of pro-
longed negative feedback, acute positive feedback, and nofeedback of estradiol.
MATERIALS AND METHODSAnimals. Two separate experiments were carried out on
sexually mature ewes maintained outdoors during the anes-trous season (June-July). In experiment 1, 11 intact eweswere used to examine the acute positive feedback effect ofestradiol in relation to physiological negative feedback pro-vided by the ovaries. These animals were divided into twogroups: (i) control-6 ewes receiving no treatment, repre-senting normal anestrous animals with physiological nega-tive feedback and (ii) positive feedback-5 ewes receivingfive Silastic estradiol implants each. These implants [3-cmpacked steroid, constructed as described (3)] were insertedsubcutaneously; five such implants have been shown tomaintain serum estradiol concentrations of -20 pg/ml in theovariectomized ewe (4). All animals were bled at two timepoints (1 and 6 hr) prior to implantation. After insertion ofthe implants into the experimental ewes, both groups werebled at 3-hr intervals for the first 12 hr and then at 2-hr inter-vals until slaughter. Ewes treated with estradiol were killed16 hr after insertion of the implants (expected time of the LHsurge), while those ewes without implants were sacrificed 2hr later.
In experiment 2, the effect of chronic estradiol negativefeedback was examined. Twelve animals were ovariecto-mized and divided into two groups of 6 ewes each. Onegroup consisted of ovariectomized ewes receiving no addi-tional treatment (no feedback). The other group (negativefeedback) was treated with a single 3-cm estradiol implant,inserted 7 days after ovariectomy. The concentration of se-rum estradiol produced by this implant has been shown to be-4 pg/ml in the ovariectomized ewe (5). The implant re-mained in place for 14 days; both groups were killed 21 daysafter ovariectomy. Blood samples were taken from bothgroups of animals three times per week from ovariectomyuntil slaughter.RIAs. Blood samples of 8 ml were collected by jugular ve-
nipuncture and serum was stored at -20'C. LH was assayedin duplicate in 20- to 200-1.d aliquots of serum using a modifi-cation (6) of an RIA described previously (7). The averagesensitivity for the 200-,ul samples was 0.24 ng of NIH-LH-S12 per ml. For assay of pituitary LH, the 15,000 x g super-natant from the initial homogenization of the tissue was di-luted 1:10,000 and duplicate determinations were made using50-200 tkl of supernatant. Values are expressed as pkg ofNIH-LH-S12 per g of pituitary tissue.RNA Isolation, Translation, and Identification. Pituitaries
were removed, and RNA was isolated (8) and then translated
Abbreviations: LH, luteinizing hormone; PRL, prolactin; GnRH,gonadotropin-releasing hormone.
1322
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
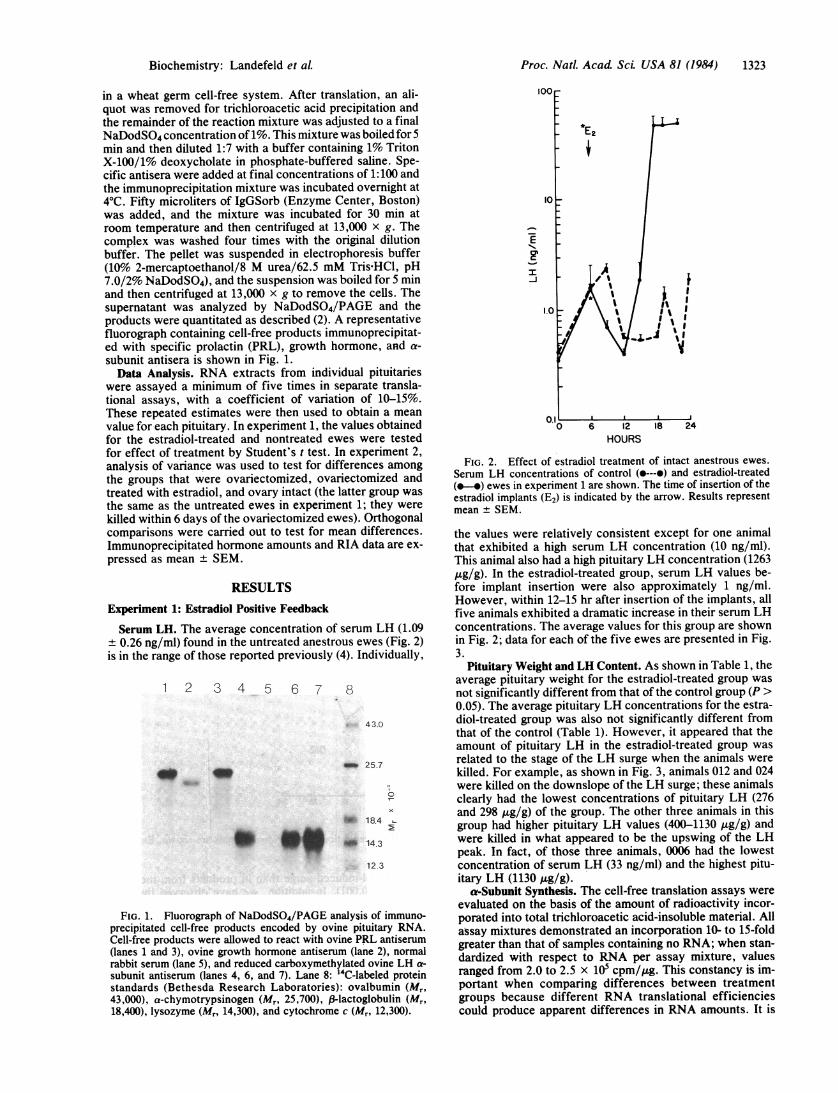
Proc. Natl. Acad Sci USA 81 (1984) 1323
in a wheat germ cell-free system. After translation, an ali-quot was removed for trichloroacetic acid precipitation andthe remainder of the reaction mixture was adjusted to a finalNaDodSO4 concentration of 1%. This mixture was boiled for5min and then diluted 1:7 with a buffer containing 1% TritonX-100/1% deoxycholate in phosphate-buffered saline. Spe-cific antisera were added at final concentrations of 1:100 andthe immunoprecipitation mixture was incubated overnight at40C. Fifty microliters of IgGSorb (Enzyme Center, Boston)was added, and the mixture was incubated for 30 min atroom temperature and then centrifuged at 13,000 x g. Thecomplex was washed four times with the original dilutionbuffer. The pellet was suspended in electrophoresis buffer(10% 2-mercaptoethanol/8 M urea/62.5 mM Tris-HCl, pH7.0/2% NaDodSO4), and the suspension was boiled for 5 minand then centrifuged at 13,000 x g to remove the cells. Thesupernatant was analyzed by NaDodSO4/PAGE and theproducts were quantitated as described (2). A representativefluorograph containing cell-free products immunoprecipitat-ed with specific prolactin (PRL), growth hormone, and a-subunit antisera is shown in Fig. 1.Data Analysis. RNA extracts from individual pituitaries
were assayed a minimum of five times in separate transla-tional assays, with a coefficient of variation of 10-15%.These repeated estimates were then used to obtain a meanvalue for each pituitary. In experiment 1, the values obtainedfor the estradiol-treated and nontreated ewes were testedfor effect of treatment by Student's t test. In experiment 2,analysis of variance was used to test for differences amongthe groups that were ovariectomized, ovariectomized andtreated with estradiol, and ovary intact (the latter group wasthe same as the untreated ewes in experiment 1; they werekilled within 6 days of the ovariectomized ewes). Orthogonalcomparisons were carried out to test for mean differences.Immunoprecipitated hormone amounts and RIA data are ex-pressed as mean ± SEM.
RESULTSExperiment 1: Estradiol Positive Feedback
Serum LH. The average concentration of serum LH (1.09± 0.26 ng/ml) found in the untreated anestrous ewes (Fig. 2)is in the range of those reported previously (4). Individually,
1 2 3 4 5 6 7 8
43.0
_0 tom..-..
OD
_i 25.7
aW. 18.4 -
_I 14.3
12 3
FIG. 1. Fluorograph of NaDodSO4/PAGE analysis of immuno-precipitated cell-free products encoded by ovine pituitary RNA.Cell-free products were allowed to react with ovine PRL antiserum(lanes 1 and 3), ovine growth hormone antiserum (lane 2), normalrabbit serum (lane 5), and reduced carboxymethylated ovine LH a-
subunit antiserum (lanes 4, 6, and 7). Lane 8: '4C-labeled proteinstandards (Bethesda Research Laboratories): ovalbumin (Mr,43,000), a-chymotrypsinogen (Mr, 25,700), (-lactoglobulin (Mr,18,400), lysozyme (Mr, 14,300), and cytochrome c (Mr, 12,300).
1.0 9
0 6 12 18 24HOURS
FIG. 2. Effect of estradiol treatment of intact anestrous ewes.Serum LH concentrations of control (e---e) and estradiol-treated(0*-) ewes in experiment 1 are shown. The time of insertion of theestradiol implants (E2) is indicated by the arrow. Results representmean ± SEM.
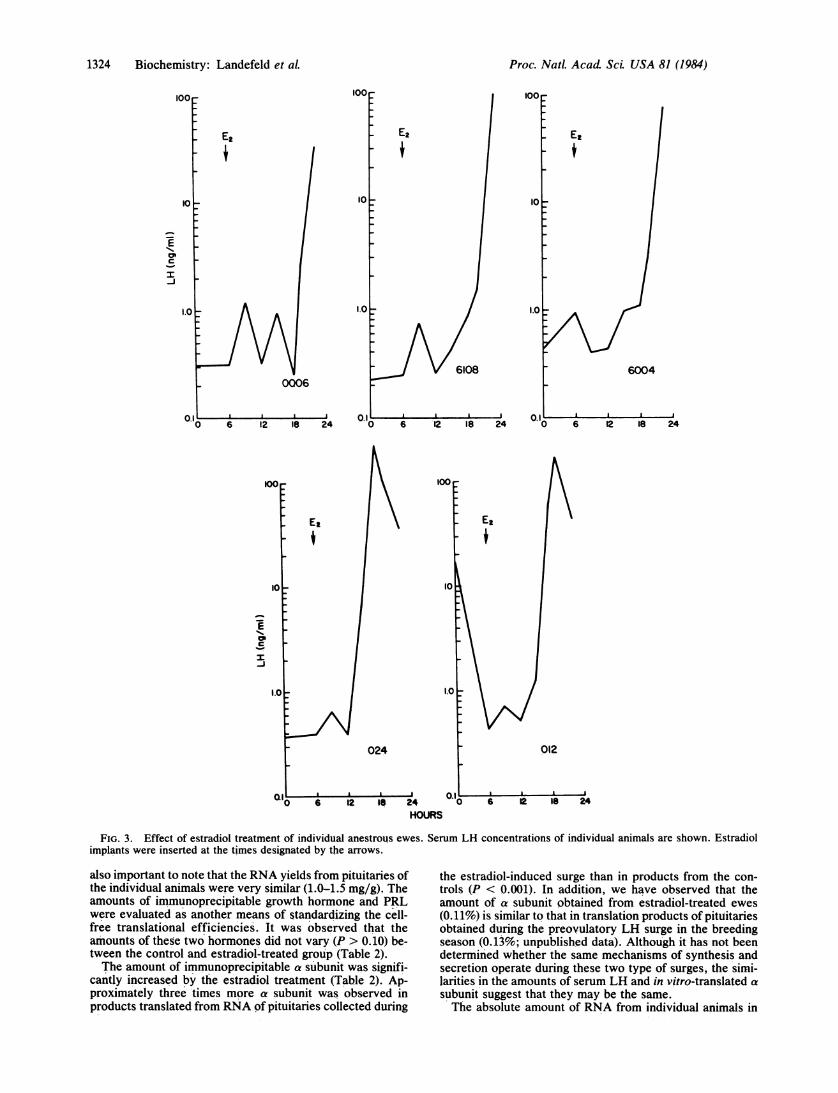
the values were relatively consistent except for one animalthat exhibited a high serum LH concentration (10 ng/ml).This animal also had a high pituitary LH concentration (1263ttg/g). In the estradiol-treated group, serum LH values be-fore implant insertion were also approximately 1 ng/ml.However, within 12-15 hr after insertion of the implants, allfive animals exhibited a dramatic increase in their serum LHconcentrations. The average values for this group are shownin Fig. 2; data for each of the five ewes are presented in Fig.3.
Pituitary Weight and LH Content. As shown in Table 1, theaverage pituitary weight for the estradiol-treated group wasnot significantly different from that of the control group (P >0.05). The average pituitary LH concentrations for the estra-diol-treated group was also not significantly different fromthat of the control (Table 1). However, it appeared that theamount of pituitary LH in the estradiol-treated group wasrelated to the stage of the LH surge when the animals werekilled. For example, as shown in Fig. 3, animals 012 and 024were killed on the downslope of the LH surge; these animalsclearly had the lowest concentrations of pituitary LH (276and 298 ug/g) of the group. The other three animals in thisgroup had higher pituitary LH values (400-1130 pkg/g) andwere killed in what appeared to be the upswing of the LHpeak. In fact, of those three animals, 0006 had the lowestconcentration of serum LH (33 ng/ml) and the highest pitu-itary LH (1130 ,ug/g).
a-Subunit Synthesis. The cell-free translation assays wereevaluated on the basis of the amount of radioactivity incor-porated into total trichloroacetic acid-insoluble material. Allassay mixtures demonstrated an incorporation 10- to 15-foldgreater than that of samples containing no RNA; when stan-dardized with respect to RNA per assay mixture, valuesranged from 2.0 to 2.5 x 10 cpm/pg. This constancy is im-portant when comparing differences between treatmentgroups because different RNA translational efficienciescould produce apparent differences in RNA amounts. It is
Biochemistry: Landefeld et aL
1324 Biochemistry: Landefeld et al.
10
c
I-J
E2
6004
0 6 12 i8 24 "0 6 2 18 24 0 6 12 18 24
100 ~~~~~~~~100
Et Ez
10 10
CPI
1.0 1.0
024 012
al . . | | o~~~~~~~~~or .0 6 12 la 24 0 6 2 la 24
HOURS
FIG. 3. Effect of estradiol treatment of individual anestrous ewes. Serum LH concentrations of individual animals are shown. Estradiolimplants were inserted at the times designated by the arrows.
also important to note that the RNA yields from pituitaries ofthe individual animals were very similar (1.0-1.5 mg/g). Theamounts of immunoprecipitable growth hormone and PRLwere evaluated as another means of standardizing the cell-free translational efficiencies. It was observed that theamounts of these two hormones did not vary (P > 0.10) be-tween the control and estradiol-treated group (Table 2).The amount of immunoprecipitable a subunit was signifi-
cantly increased by the estradiol treatment (Table 2). Ap-proximately three'times more a subunit was observed inproducts translated from RNA of pituitaries collected during
the estradiol-induced surge than in products from the con-trols (P < 0.001). In addition, we have observed that theamount of a subunit obtained from estradiol-treated ewes(0.11%) is similar to that in translation products of pituitariesobtained during the preovulatory LH surge in the breedingseason (0.13%; unpublished data). Although it has not beendetermined whether the same mechanisms of synthesis andsecretion operate during these two type of surges, the simi-larities in the amounts of serum LH and in vitro-translated asubunit suggest that they may be the same.The absolute amount of RNA from individual animals in
Proc. Natl. Acad Sci. USA 81 (1984)
Proc. Natl. Acad ScL USA 81 (1984) 1325
Table 1. Average pituitary weight and LH concentration inexperiment 1 (positive feedback of estradiol)
Treatment group Pituitary weight, g Pituitary LH, Mg/gControl 1.12 ± 0.11 633 ± 134Estradiol-treated 1.57 ± 0.17 569 ± 162
Values are mean ± SEM. None of the values are different (P >0.05, Student's t test).
the estradiol-treated group appeared to be related to thestage of the LH surge. Although valid statistical compari-sons are not possible because of the small number of ani-mals, there appeared to be a trend for higher amounts (0.14± 0.01%) of a subunit in RNA products from ewes on thedownslope of their surge (animals 012 and 024) than thosevalues (0.08 ± 0.01%) from animals on the incline of the peak(animals 6004, 6108, and 0006).
Experiment 2: Estradiol Negative Feedback
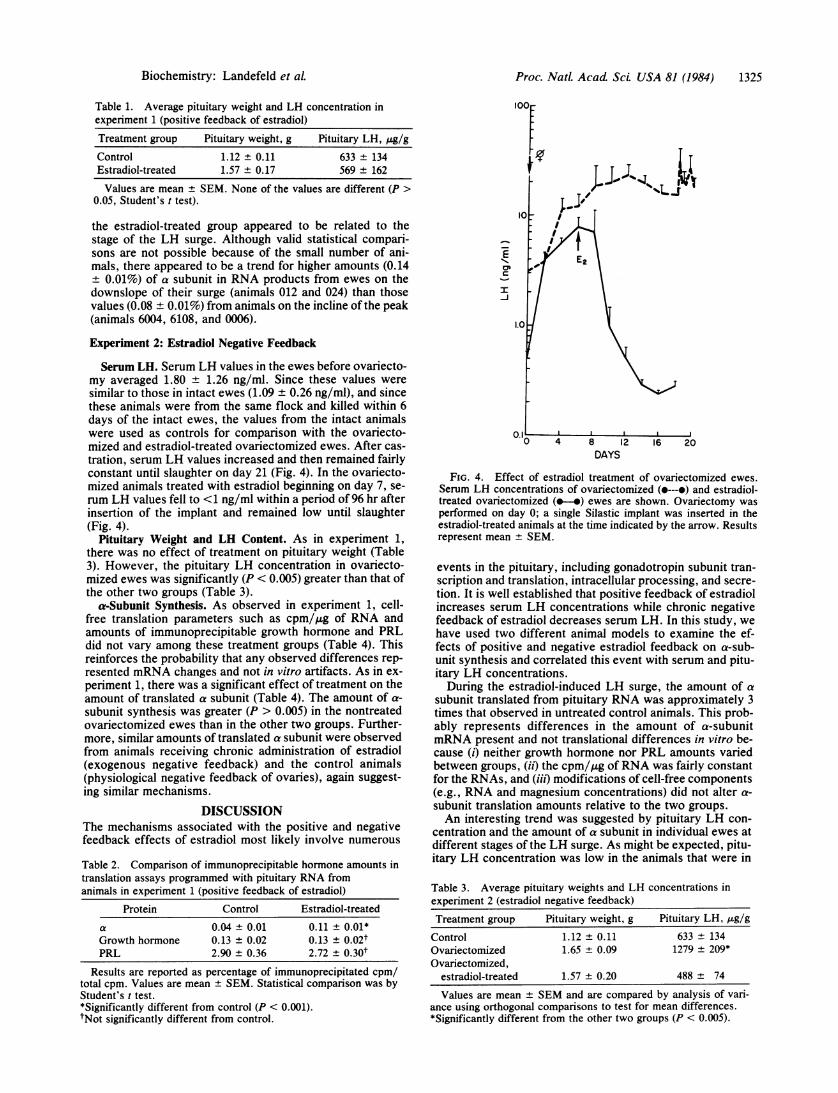
Serum LH. Serum LH values in the ewes before ovariecto-my averaged 1.80 + 1.26 ng/ml. Since these values weresimilar to those in intact ewes (1.09 ± 0.26 ng/ml), and sincethese animals were from the same flock and killed within 6days of the intact ewes, the values from the intact animalswere used as controls for comparison with the ovariecto-mized and estradiol-treated ovariectomized ewes. After cas-tration, serum LH values increased and then remained fairlyconstant until slaughter on day 21 (Fig. 4). In the ovariecto-mized animals treated with estradiol beginning on day 7, se-rum LH values fell to <1 ng/ml within a period of 96 hr afterinsertion of the implant and remained low until slaughter(Fig. 4).
Pituitary Weight and LH Content. As in experiment 1,there was no effect of treatment on pituitary weight (Table3). However, the pituitary LH concentration in ovariecto-mized ewes was significantly (P < 0.005) greater than that ofthe other two groups (Table 3).
a-Subunit Synthesis. As observed in experiment 1, cell-free translation parameters such as cpm/,ug of RNA andamounts of immunoprecipitable growth hormone and PRLdid not vary among these treatment groups (Table 4). Thisreinforces the probability that any observed differences rep-resented mRNA changes and not in vitro artifacts. As in ex-periment 1, there was a significant effect of treatment on theamount of translated a subunit (Table 4). The amount of a-subunit synthesis was greater (P > 0.005) in the nontreatedovariectomized ewes than in the other two groups. Further-more, similar amounts of translated a subunit were observedfrom animals receiving chronic administration of estradiol(exogenous negative feedback) and the control animals(physiological negative feedback of ovaries), again suggest-ing similar mechanisms.
DISCUSSIONThe mechanisms associated with the positive and negativefeedback effects of estradiol most likely involve numerous
Table 2. Comparison of immunoprecipitable hormone amounts intranslation assays programmed with pituitary RNA fromanimals in experiment 1 (positive feedback of estradiol)
Protein Control Estradiol-treated
a 0.04 ± 0.01 0.11 ± 0.01*Growth hormone 0.13 ± 0.02 0.13 ± 0.02tPRL 2.90 ± 0.36 2.72 ± 0.30t
Results are reported as percentage of immunoprecipitated cpm/total cpm. Values are mean ± SEM. Statistical comparison was byStudent's t test.*Significantly different from control (P < 0.001).tNot significantly different from control.
T
I /-JI
1.0
0.10 4 8 12 16 20
DAYS
FIG. 4. Effect of estradiol treatment of ovariectomized ewes.
Serum LH concentrations of ovariectomized (e---o) and estradiol-treated ovariectomized (e-o) ewes are shown. Ovariectomy was
performed on day 0; a single Silastic implant was inserted in theestradiol-treated animals at the time indicated by the arrow. Resultsrepresent mean + SEM.
events in the pituitary, including gonadotropin subunit tran-scription and translation, intracellular processing, and secre-
tion. It is well established that positive feedback of estradiolincreases serum LH concentrations while chronic negativefeedback of estradiol decreases serum LH. In this study, wehave used two different animal models to examine the ef-fects of positive and negative estradiol feedback on a-sub-unit synthesis and correlated this event with serum and pitu-itary LH concentrations.During the estradiol-induced LH surge, the amount of a
subunit translated from pituitary RNA was approximately 3times that observed in untreated control animals. This prob-ably represents differences in the amount of a-subunitmRNA present and not translational differences in vitro be-cause (i) neither growth hormone nor PRL amounts variedbetween groups, (ii) the cpm/,ug ofRNA was fairly constantfor the RNAs, and (iii) modifications of cell-free components(e.g., RNA and magnesium concentrations) did not alter a-
subunit translation amounts relative to the two groups.An interesting trend was suggested by pituitary LH con-
centration and the amount of a subunit in individual ewes atdifferent stages of the LH surge. As might be expected, pitu-itary LH concentration was low in the animals that were in
Table 3. Average pituitary weights and LH concentrations inexperiment 2 (estradiol negative feedback)
Treatment group Pituitary weight, g Pituitary LH, ,ug/gControl 1.12 ± 0.11 633 + 134Ovariectomized 1.65 ± 0.09 1279 ± 209*Ovariectomized,
estradiol-treated 1.57 ± 0.20 488 ± 74
Values are mean + SEM and are compared by analysis of vari-ance using orthogonal comparisons to test for mean differences.*Significantly different from the other two groups (P < 0.005).
Biochemistry: Landefeld et aL

1326 Biochemistry: Landefeld et al.
Table 4. Comparison of immunoprecipitable hormone amounts intranslation assays programmed with pituitary RNA from animalsin experiment 2 (estradiol negative feedback)
Ovariectomized,Protein Control Ovariectomized estradiol-treated
a 0.04 + 0.01 0.18 ± 0.03* 0.04 ± 0.01Growthhormone 0.13 ± 0.02 0.11 ± 0.02 0.12 ± 0.01
PRL 2.90 ± 0.36 2.55 ± 0.17 2.53 ± 0.11
Results are reported as percentage of immunoprecipitated cpm/total cpm. The values are expressed as mean ± SEM and were com-pared by analysis of variance using orthogonal comparisons to testfor mean differences.*Significantly different from the other two groups (P < 0.005).
the final stage of their surge (nos. 024 and 012) while thoseanimals on the upswing had a higher level of pituitary LH.With regard to a-subunit synthesis, however, the results sug-gest that there may be an increase near the completion of thepeak, as LH content becomes depleted, rather than on theupswing when LH concentration is the greatest. Furtherstudies with additional animals are needed to verify thesetrends.With regard to the negative feedback effects of estradiol, it
was observed that ovariectomy resulted in a 3- to 4-fold in-crease in the amount of translated a subunit. Further, chron-ic treatment of castrated ewes with a physiological dose ofestradiol reduced the amount of translated a subunit to thatin the normal anestrous ewes (O0.05% of total products).These results are similar to those reported (2) in which cas-trated ewes during the breeding season were treated with es-tradiol and progesterone. Furthermore, these studies agreewith results of others who have demonstrated an effect ofhormonal manipulation on the amount of a-subunit mRNA(9-12), supporting differential regulation of the individualsubunit mRNAs.Although it has been established that estradiol exerts both
a positive and a negative effect on serum LH concentrationsduring the estrous cycle, the mechanisms underlying theseactions are not clear. For example, Moss et al. (13) demon-strated an increase in gonadotropin-releasing hormone(GnRH) receptors in the ovine pituitary after estradiol treat-ment while Goodman and Karsch (14) observed a suppres-sion of the response of the pituitary to GnRH after estradioladministration. Those results, in addition to others (15-17),suggest effects on the pituitary response to GnRH. Althoughthe results of this study cannot delineate the actual mecha-nisms involved, it is interesting that the levels of a-subunitmRNA are affected in both positive and negative feedbackstates. Thus, it is conceivable that these actions could be atthe level of transcription of the subunit mRNAs, similar toeffects of estradiol on uteroglobin (18) and on ovalbumin andconalbumin (19) and the effects of glucocorticoids on pro-opiomelanocortin (20). In addition, recent results suggestingthe presence of GnRH receptors on the nucleus (21) supportan alternative possibility of GnRH action on RNA synthesis/degradation. Thus, estradiol could be affecting mRNAamounts directly or through GnRH, and both actions may
occur under physiologic conditions. Further studies usingcDNA probes and transcription assays are needed to investi-gate the effects on transcription.
In summary, it appears that there is an increase in a-sub-unit mRNA during the estradiol-induced LH surge in the an-estrous ewe (positive feedback), associated with the in-creased LH secretion observed at that time. Also, it is sug-gested that the amount of the mRNA increase may bedependent on the stage of the surge, increasing as the pitu-itary becomes depleted of LH. Conversely, chronic treat-ment of ovariectomized ewes with estradiol (negative feed-back) appears to decrease a-subunit synthesis, which is cor-related with decreased serum LH concentrations.
The technical assistance of Inna Levitan, Natalie Williams, andDoug Doop is gratefully acknowledged. The reagents for the RIAwere provided by Dr. G. D. Niswender. The ovine growth hormoneantiserum was provided by Dr. Al Parlow. This work was supportedby National Institutes of Health Grants HD-12016 and HD-11311.
1. Alexander, D. C. & Miller, W. L. (1982) J. Biol. Chem. 257,2282-2286.
2. Landefeld, T. D., Kepa, J. & Karsch, F. J. (1983) J. Biol.Chem. 258, 2390-2393.
3. Karsch, F. J., Dierschke, D. J., Weick, R. F., Yamaji, T.,Hotchkiss, J. & Knobil, E. (1973) Endocrinology 92, 799-804.
4. Karsch, F. J. & Foster, D. L. (1975) Endocrinology 97, 373-379.
5. Legan, S. J., Karsch, F. J. & Foster, D. L. (1977) Endocrinol-ogy 101, 818-824.
6. Hauger, R. L., Karsch, F. J. & Foster, D. L. (1977) Endocri-nology 101, 807-817.
7. Niswender, G. D., Reichert, L. E., Jr., Midgley, A. R., Jr., &Nalbandov, A. V. (1969) Endocrinology 84, 1166-1173.
8. Landefeld, T. D. (1979) J. Biol. Chem. 254, 3685-3688.9. Counis, R., Corbani, M. & Jutisz, M. (1982) in Pituitary Hor-
mones and Related Peptides, eds. Motta, M., Zanisi, M. &Piva, F. (Academic, New York), Vol. 49, pp. 49-61.
10. Chin, W. W. & Habener, J. F. (1983) Clin. Res. 31, 469(abstr.).
11. Keller, D., Fetherston, J. & Boime, I. (1980) Eur. J. Biochem.108, 367-372.
12. Shupnik, M. A., Chin, W. W., Ross, D. S., Downing, M. F.,Habener, J. F. & Ridgway, E. C. (1983) in J. Biol. Chem. 258,15120-15124.
13. Moss, G. E., Crowder, M. E. & Nett, T. M. (1981) Biol. Re-prod. 25, 938-944.
14. Goodman, R. L. & Karsch, F. J. (1980) Endocrinology 107,1286-1290.
15. Moss, G. E. & Nett, T. M. (1980) Biol. Reprod. 23, 398-403.16. Huang, E. S. R. & Miller, W. L. (1980) Biol. Reprod. 23, 124-
134.17. Liu, T.-C. & Jackson, G. L. (1977) Endocrinology 100, 1294-
1302.18. Shen, X.-Z., Ming-Jer, T., Bullock, D. W. & Woo, S. L. C.
(1983) Endocrinology 112, 871-876.19. Palmiter, R. D., Mulvihill, E. R., Shephard, J. H. & Mc-
Knight, G. S. (1981) J. Biol. Chem. 256, 7910-7916.20. Schachter, B. S., Johnson, L. K., Baxter, J. D. & Roberts,
J. L. (1982) Endocrinology 110, 1442-1444.21. Millar, R., Rosen, H., Badminton, M., Pasqualini, C. & Ker-
delhue, B. (1983) FEBS Lett. 153, 382-386.
Proc. NatL Acad Sci. USA 81 (1984)





























