Embed Size (px)
Citation preview
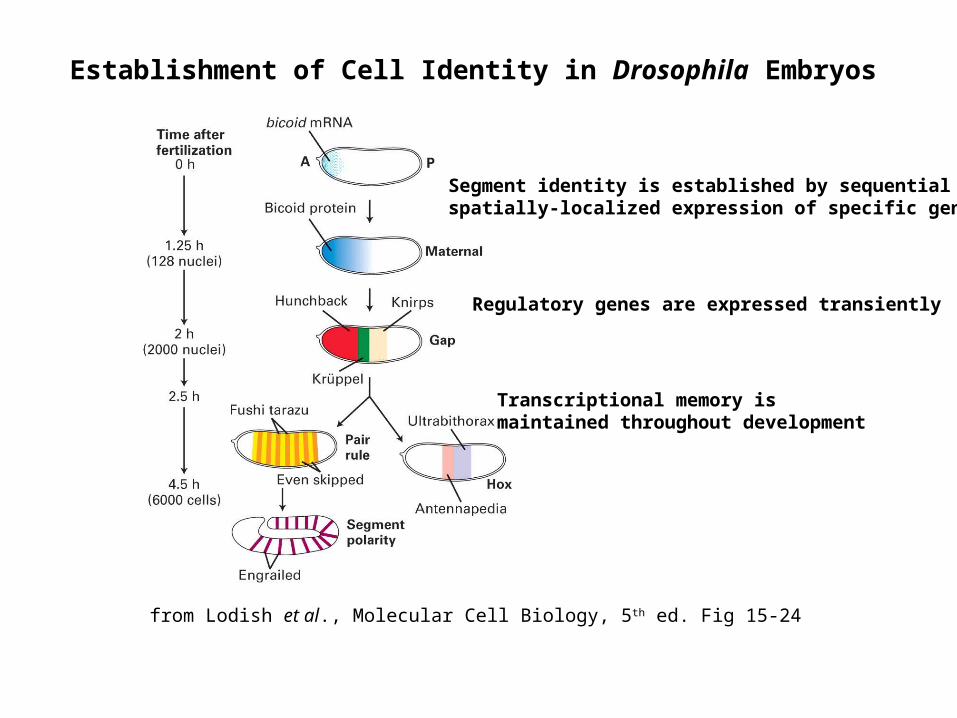
Establishment of Cell Identity in Drosophila Embryos
from Lodish et al., Molecular Cell Biology, 5th ed. Fig 15-24
Segment identity is established by sequential spatially-localized expression of specific genes
Regulatory genes are expressed transiently
Transcriptional memory is maintained throughout development

Misexpression of Homeotic Genes Lead to Morphological Abominations
from Lodish et al., Molecular Cell Biology, 5th ed. Fig 15-25

Polycomb-group Proteins
Maintains a silenced state
Trithorax-group Proteins
Maintains an active state
Counteracts the action of PcG proteins
Memory system composed of PcG and trxG complexes is linked to the histone code
Prevents changes in cell identity by preserving transcription patterns
Polycomb and Trithorax Complexes
Chromatin is altered in a heritable manner
Prevents chromatin remodelling
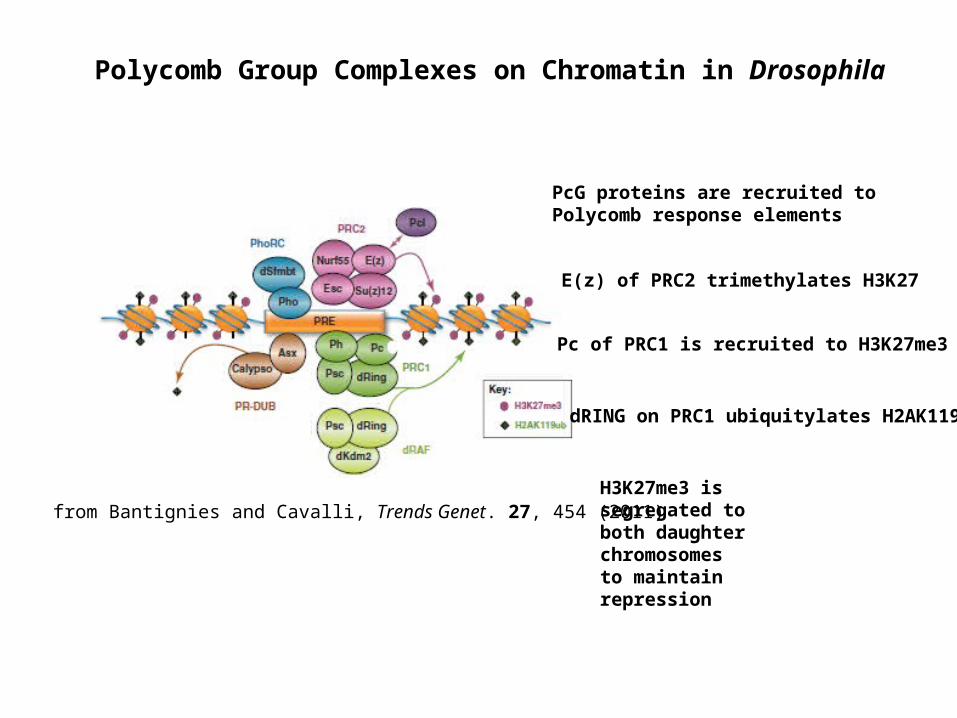
Polycomb Group Complexes on Chromatin in Drosophila
from Bantignies and Cavalli, Trends Genet. 27, 454 (2011)
PcG proteins are recruited to Polycomb response elements
E(z) of PRC2 trimethylates H3K27
Pc of PRC1 is recruited to H3K27me3
dRING on PRC1 ubiquitylates H2AK119
H3K27me3 is segregated to both daughter chromosomes to maintain repression
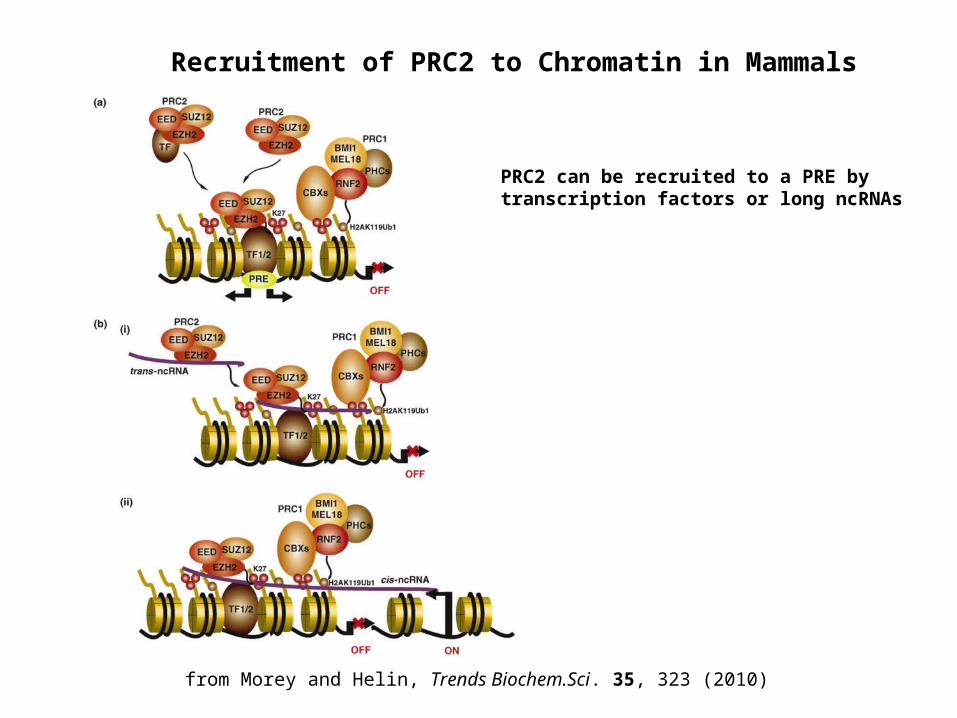
Recruitment of PRC2 to Chromatin in Mammals
PRC2 can be recruited to a PRE by transcription factors or long ncRNAs
from Morey and Helin, Trends Biochem.Sci. 35, 323 (2010)
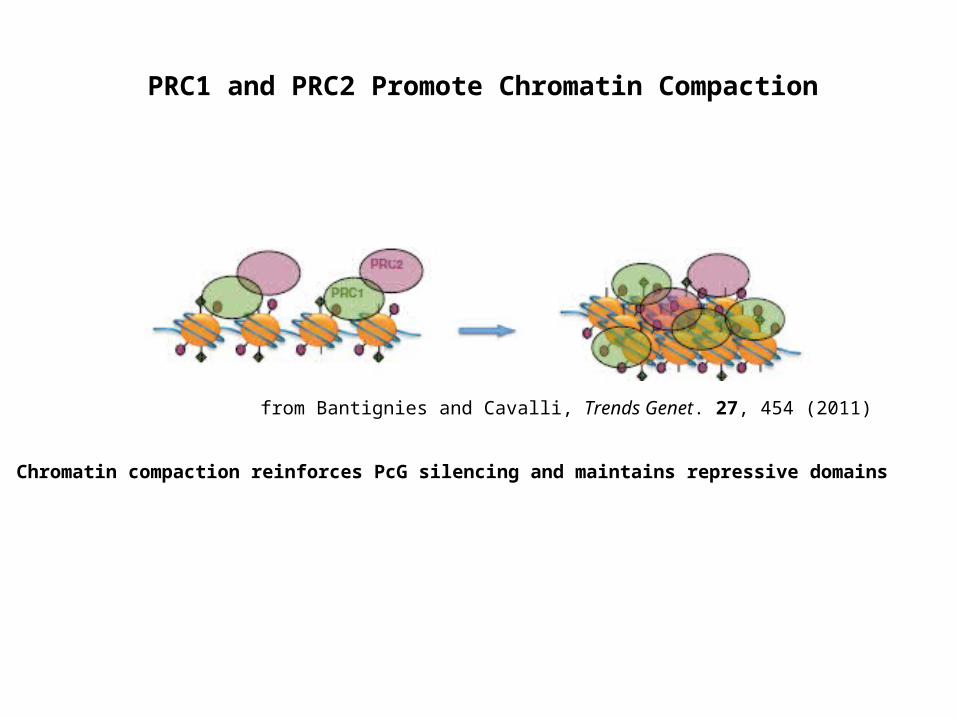
from Bantignies and Cavalli, Trends Genet. 27, 454 (2011)
Chromatin compaction reinforces PcG silencing and maintains repressive domains
PRC1 and PRC2 Promote Chromatin Compaction
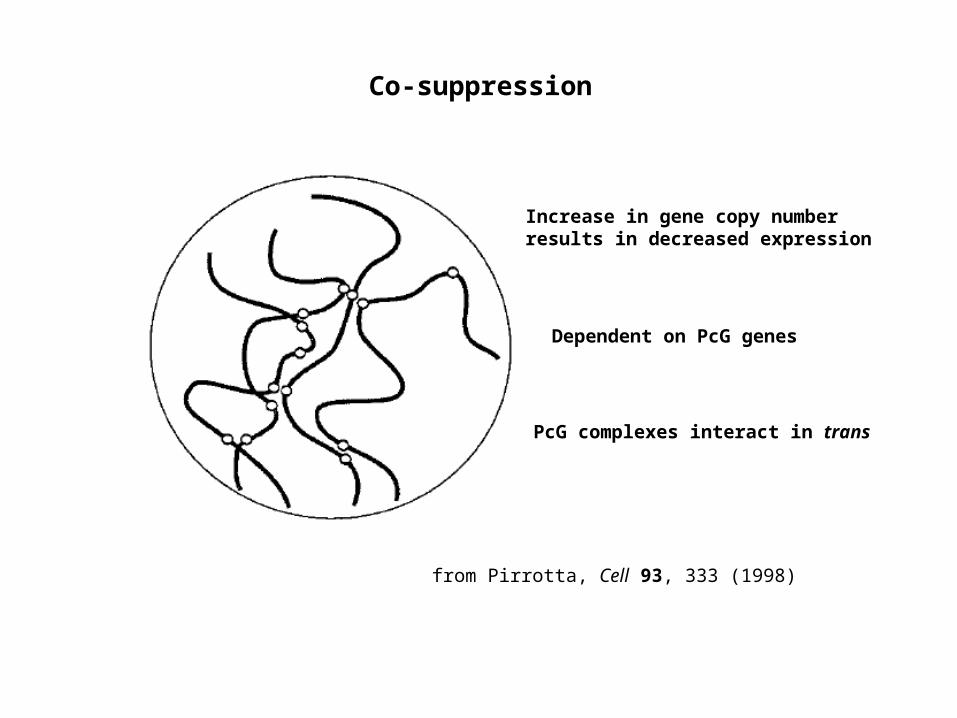
Co-suppression
PcG complexes interact in trans
Increase in gene copy number results in decreased expression
Dependent on PcG genes
from Pirrotta, Cell 93, 333 (1998)
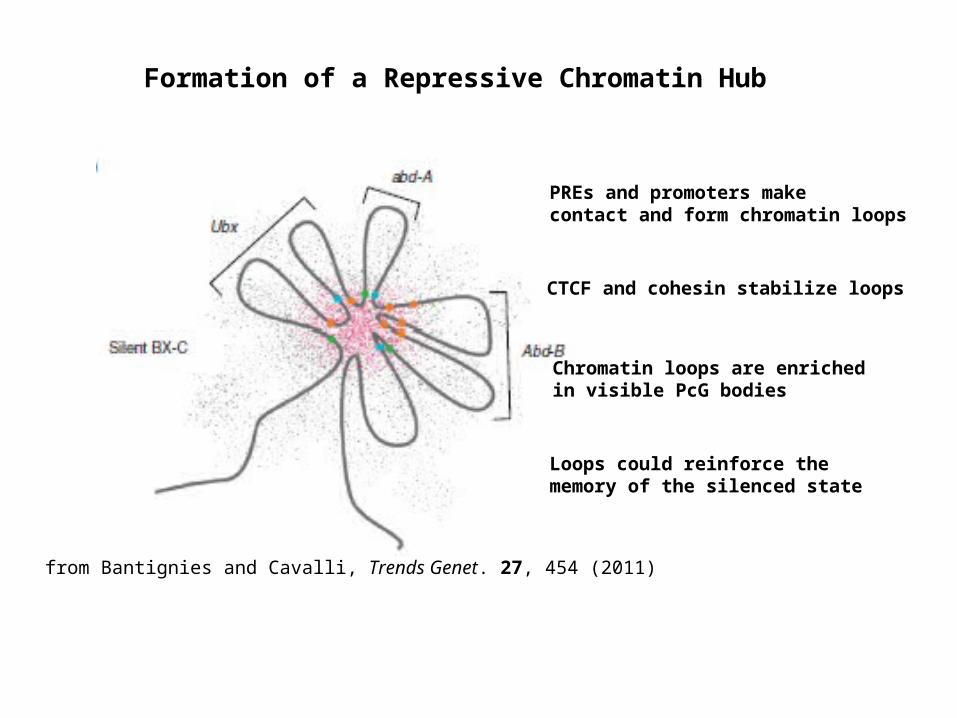
Formation of a Repressive Chromatin Hub
PREs and promoters make contact and form chromatin loops
CTCF and cohesin stabilize loops
Chromatin loops are enriched in visible PcG bodies
Loops could reinforce the memory of the silenced state
from Bantignies and Cavalli, Trends Genet. 27, 454 (2011)

Chromosome Kissing
PcG proteins mediate long-range chromatin contacts
Distant complexes of chromosome loops can interact with eath other
from Bantignies and Cavalli, Trends Genet. 27, 454 (2011)
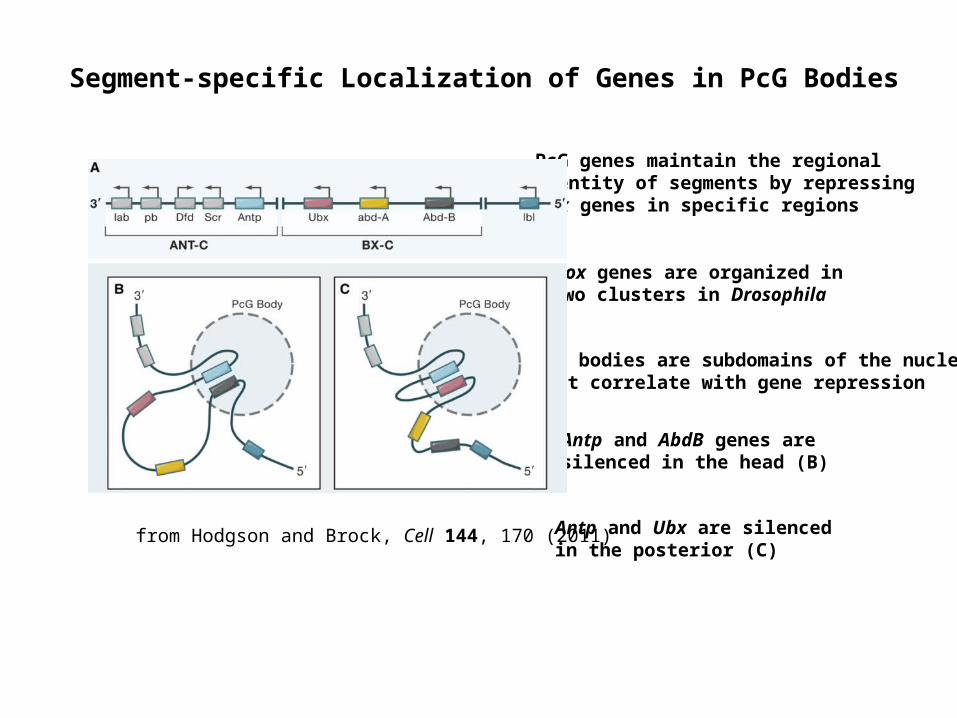
Segment-specific Localization of Genes in PcG Bodies
Hox genes are organized in two clusters in Drosophila
PcG bodies are subdomains of the nucleus that correlate with gene repression
Antp and AbdB genes are silenced in the head (B)
Antp and Ubx are silenced in the posterior (C)
PcG genes maintain the regional identity of segments by repressing Hox genes in specific regions
from Hodgson and Brock, Cell 144, 170 (2011)
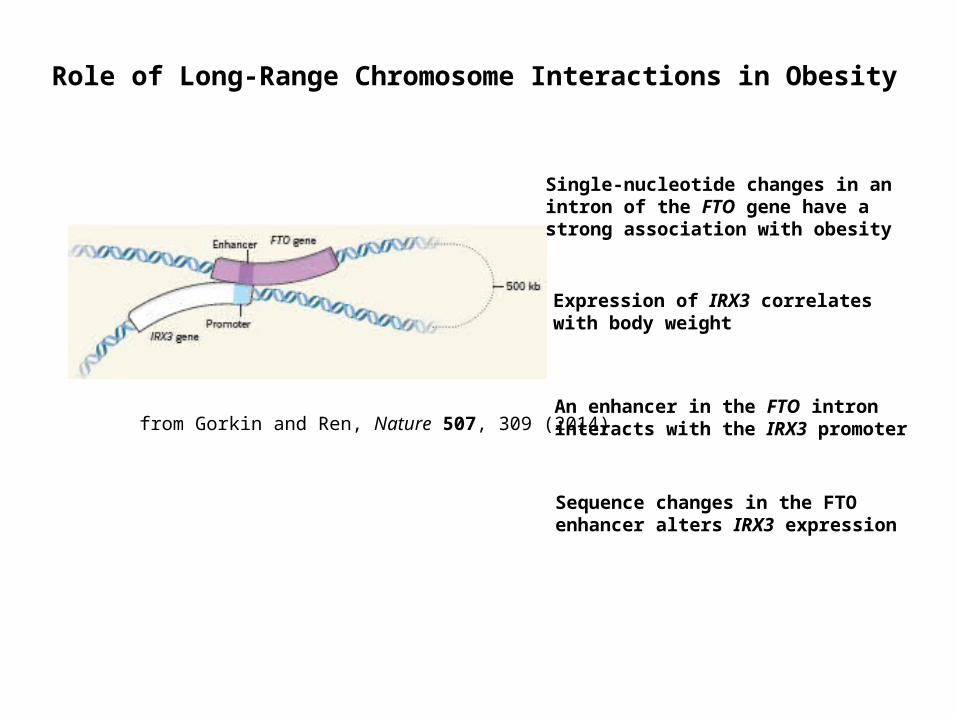
from Gorkin and Ren, Nature 507, 309 (2014)
Role of Long-Range Chromosome Interactions in Obesity
Single-nucleotide changes in an intron of the FTO gene have a strong association with obesity
Expression of IRX3 correlates with body weight
Sequence changes in the FTO enhancer alters IRX3 expression
An enhancer in the FTO intron interacts with the IRX3 promoter
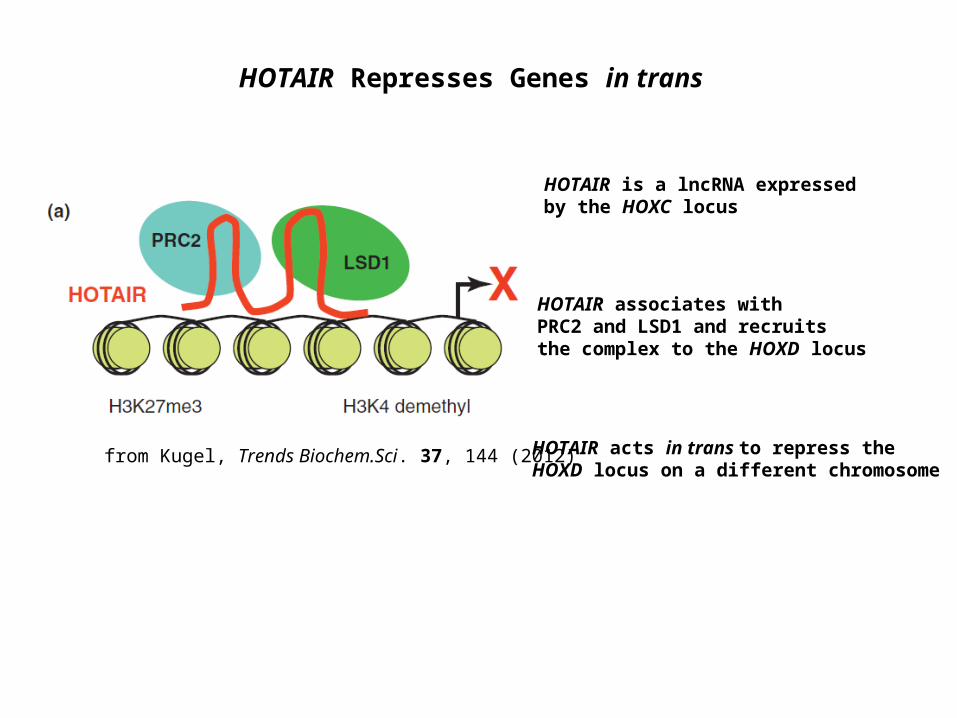
HOTAIR Represses Genes in trans
HOTAIR is a lncRNA expressed by the HOXC locus
HOTAIR acts in trans to repress the HOXD locus on a different chromosome
HOTAIR associates with PRC2 and LSD1 and recruits the complex to the HOXD locus
from Kugel, Trends Biochem.Sci. 37, 144 (2012)
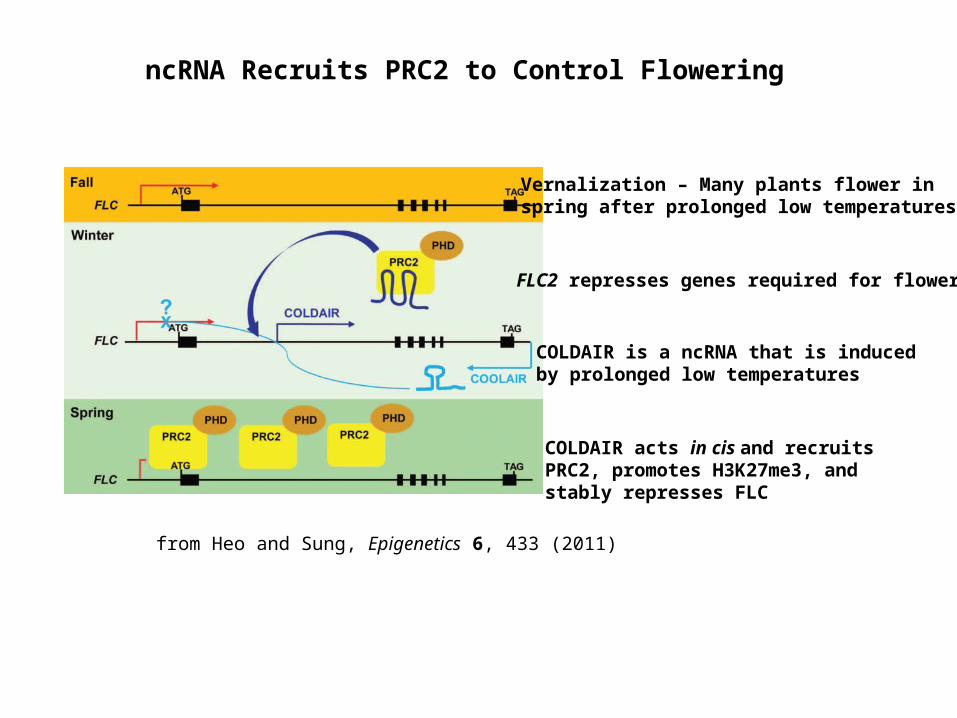
ncRNA Recruits PRC2 to Control Flowering
from Heo and Sung, Epigenetics 6, 433 (2011)
Vernalization – Many plants flower in spring after prolonged low temperatures
FLC2 represses genes required for flowering
COLDAIR is a ncRNA that is induced by prolonged low temperatures
COLDAIR acts in cis and recruits PRC2, promotes H3K27me3, and stably represses FLC
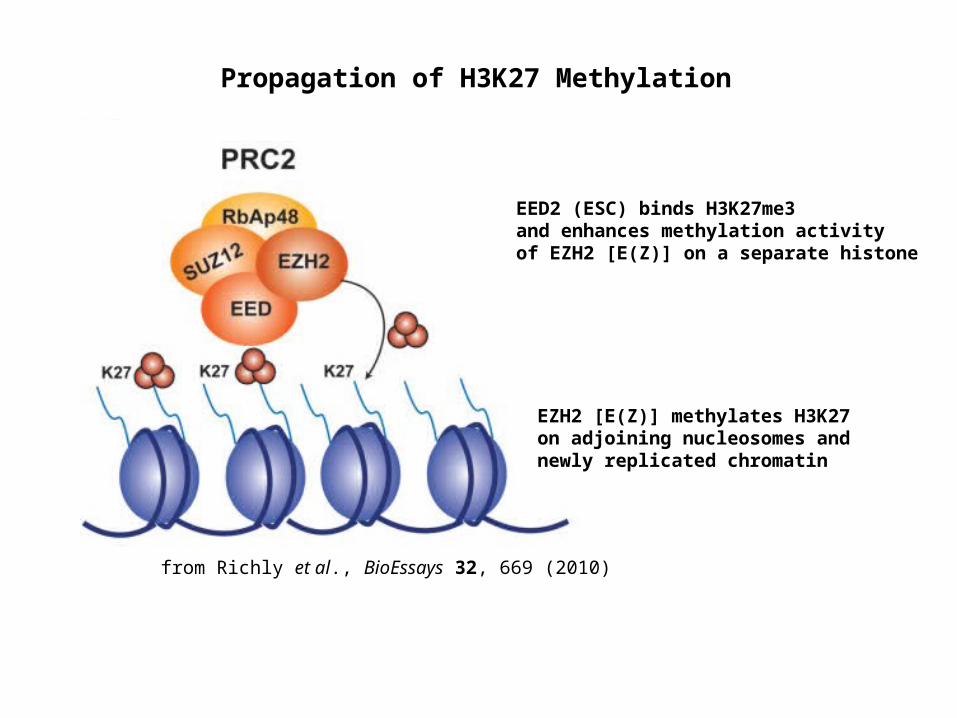
from Richly et al., BioEssays 32, 669 (2010)
Propagation of H3K27 Methylation
EED2 (ESC) binds H3K27me3 and enhances methylation activity of EZH2 [E(Z)] on a separate histone
EZH2 [E(Z)] methylates H3K27 on adjoining nucleosomes and newly replicated chromatin
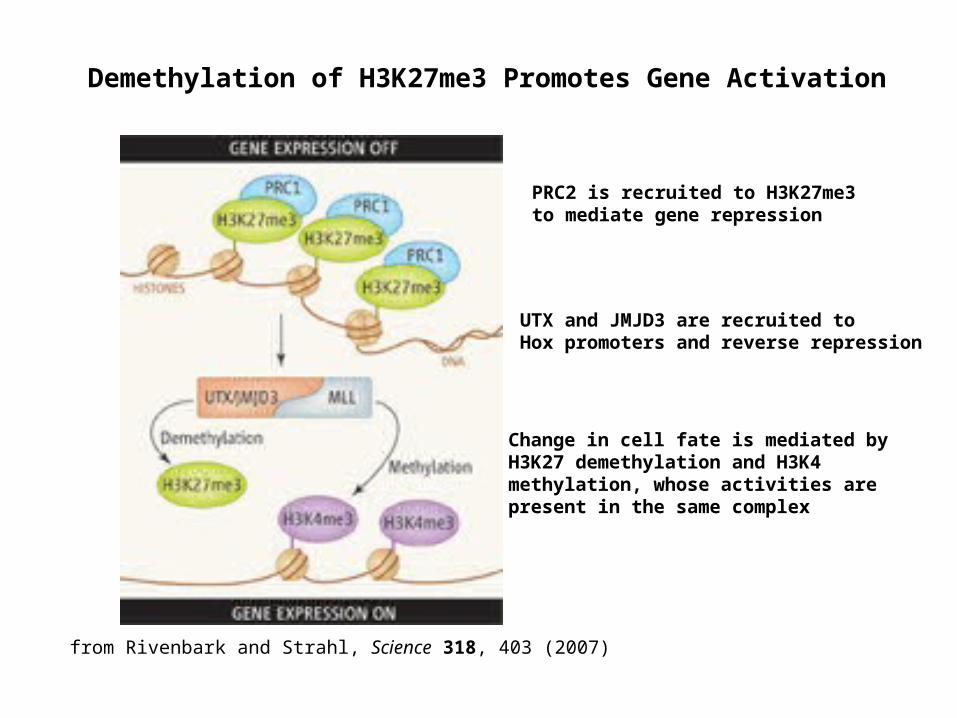
Demethylation of H3K27me3 Promotes Gene Activation
PRC2 is recruited to H3K27me3 to mediate gene repression
UTX and JMJD3 are recruited to Hox promoters and reverse repression
Change in cell fate is mediated by H3K27 demethylation and H3K4 methylation, whose activities are present in the same complex
from Rivenbark and Strahl, Science 318, 403 (2007)

Trithorax-group Protein Mechanism of Action
TrxG proteins maintain an active transcriptional state
TrxG proteins modify histones, remodel chromatin, and oppose PcG-mediated gene silencing
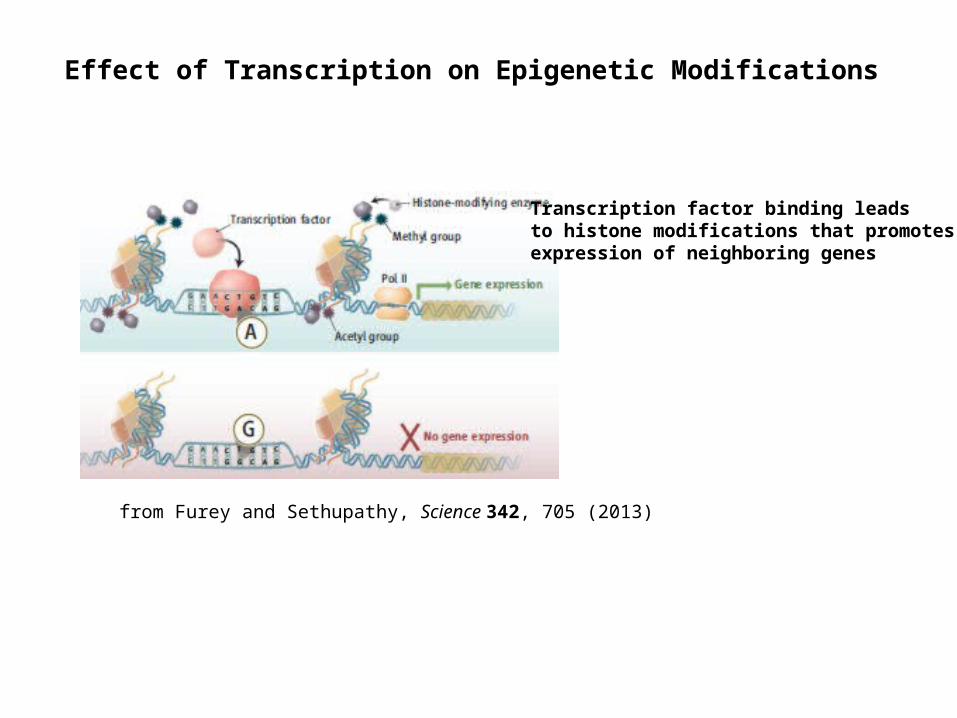
from Furey and Sethupathy, Science 342, 705 (2013)
Effect of Transcription on Epigenetic Modifications
Transcription factor binding leads to histone modifications that promotes expression of neighboring genes
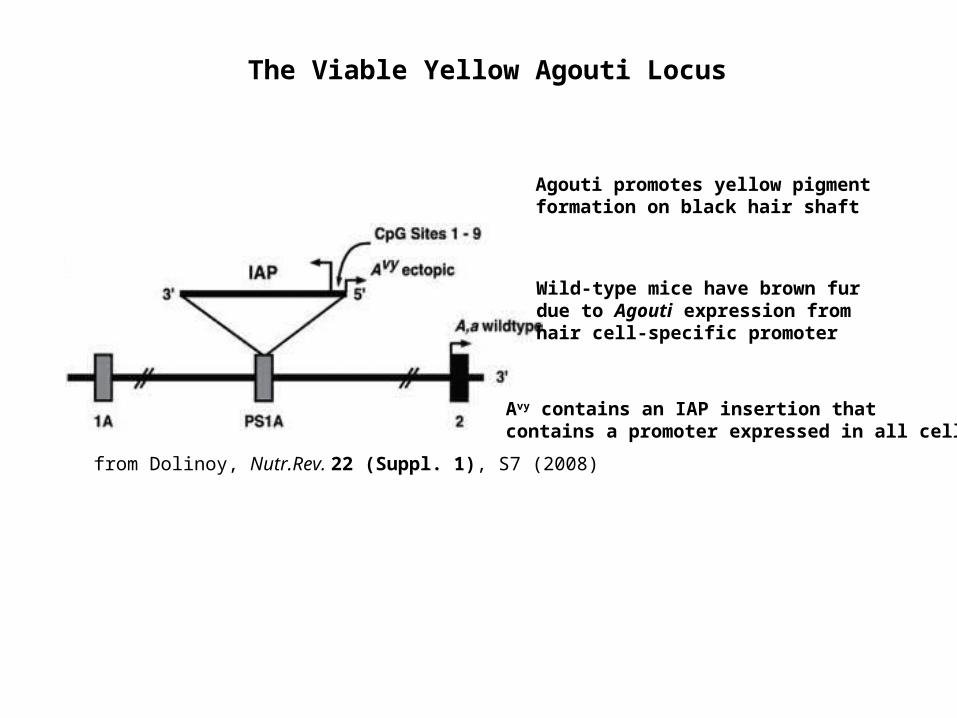
from Dolinoy, Nutr.Rev. 22 (Suppl. 1), S7 (2008)
The Viable Yellow Agouti Locus
Agouti promotes yellow pigment formation on black hair shaft
Wild-type mice have brown fur due to Agouti expression from hair cell-specific promoter
Avy contains an IAP insertion that contains a promoter expressed in all cells
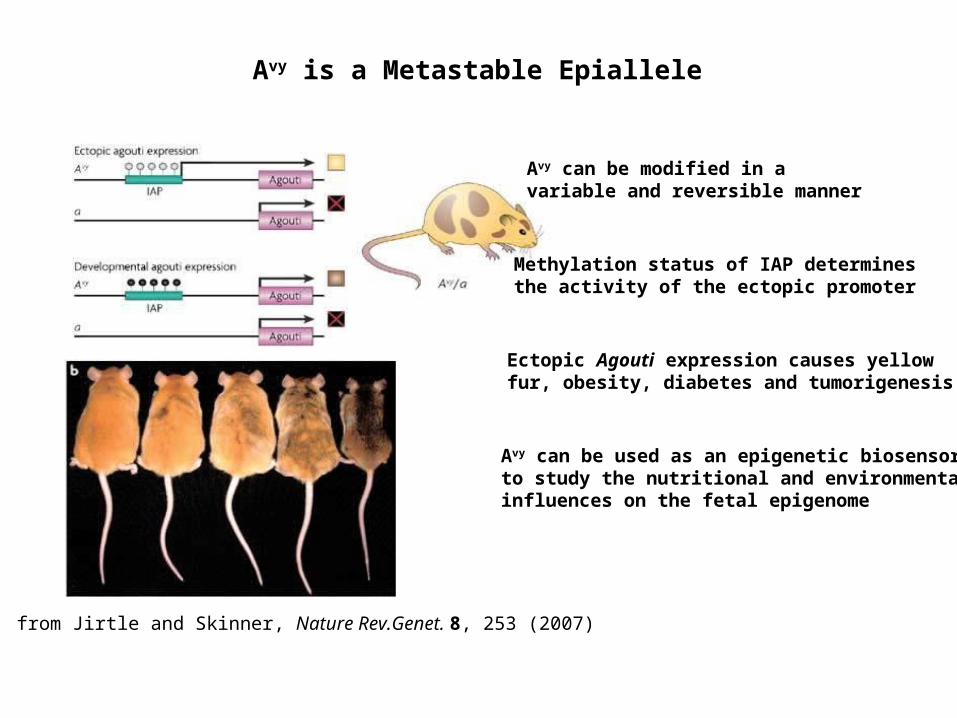
from Jirtle and Skinner, Nature Rev.Genet. 8, 253 (2007)
Avy is a Metastable Epiallele
Avy can be modified in a variable and reversible manner
Methylation status of IAP determines the activity of the ectopic promoter
Avy can be used as an epigenetic biosensor to study the nutritional and environmental influences on the fetal epigenome
Ectopic Agouti expression causes yellow fur, obesity, diabetes and tumorigenesis
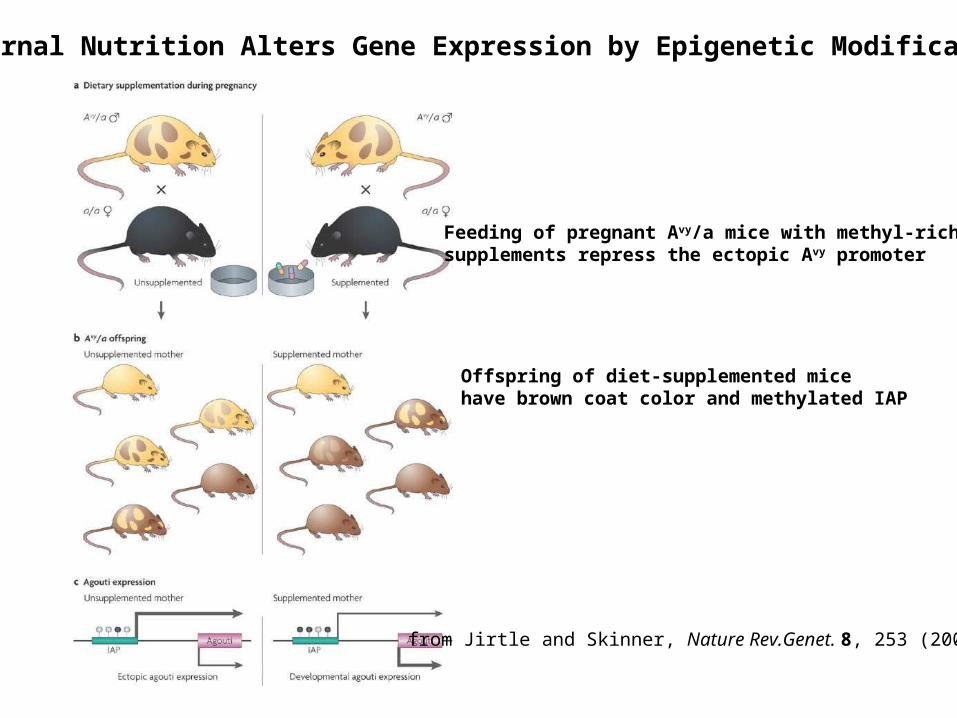
Maternal Nutrition Alters Gene Expression by Epigenetic Modification
from Jirtle and Skinner, Nature Rev.Genet. 8, 253 (2007)
Feeding of pregnant Avy/a mice with methyl-rich supplements repress the ectopic Avy promoter
Offspring of diet-supplemented mice have brown coat color and methylated IAP
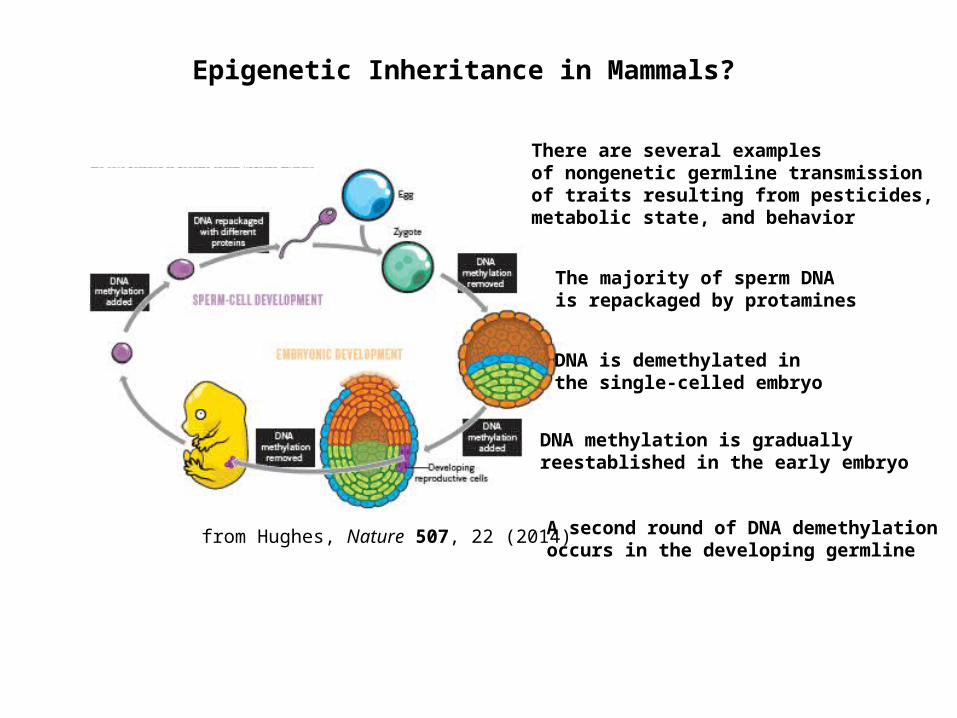
from Hughes, Nature 507, 22 (2014)
The majority of sperm DNA is repackaged by protamines
DNA is demethylated in the single-celled embryo
DNA methylation is gradually reestablished in the early embryo
Epigenetic Inheritance in Mammals?
A second round of DNA demethylation occurs in the developing germline
There are several examples of nongenetic germline transmission of traits resulting from pesticides, metabolic state, and behavior
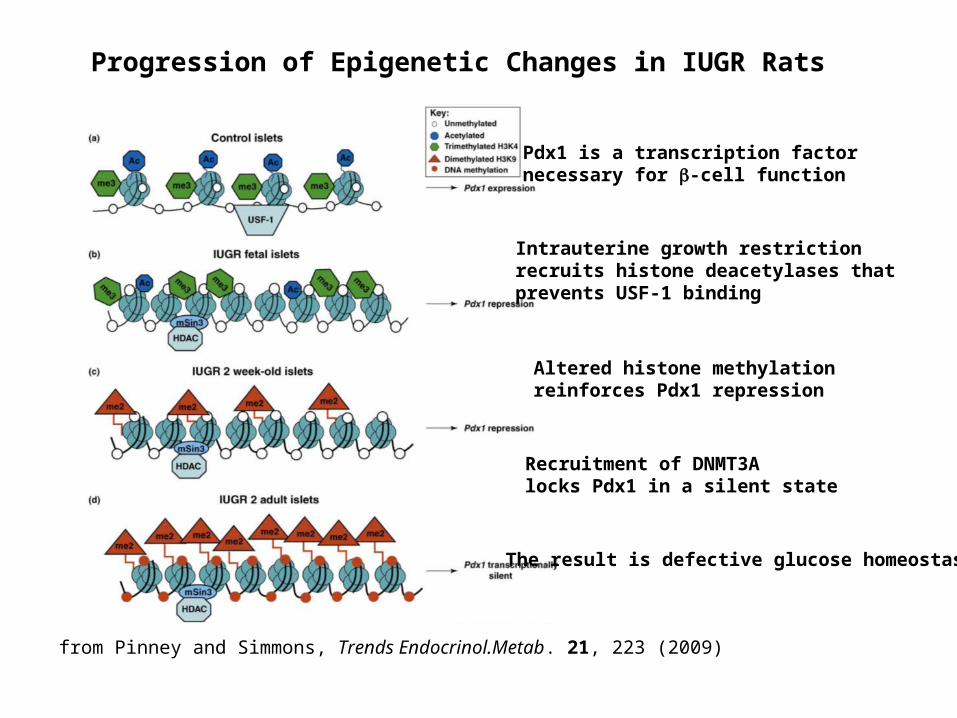
Progression of Epigenetic Changes in IUGR Rats
Pdx1 is a transcription factor necessary for -cell function
from Pinney and Simmons, Trends Endocrinol.Metab. 21, 223 (2009)
Intrauterine growth restriction recruits histone deacetylases that prevents USF-1 binding
Altered histone methylation reinforces Pdx1 repression
Recruitment of DNMT3A locks Pdx1 in a silent state
The result is defective glucose homeostasis
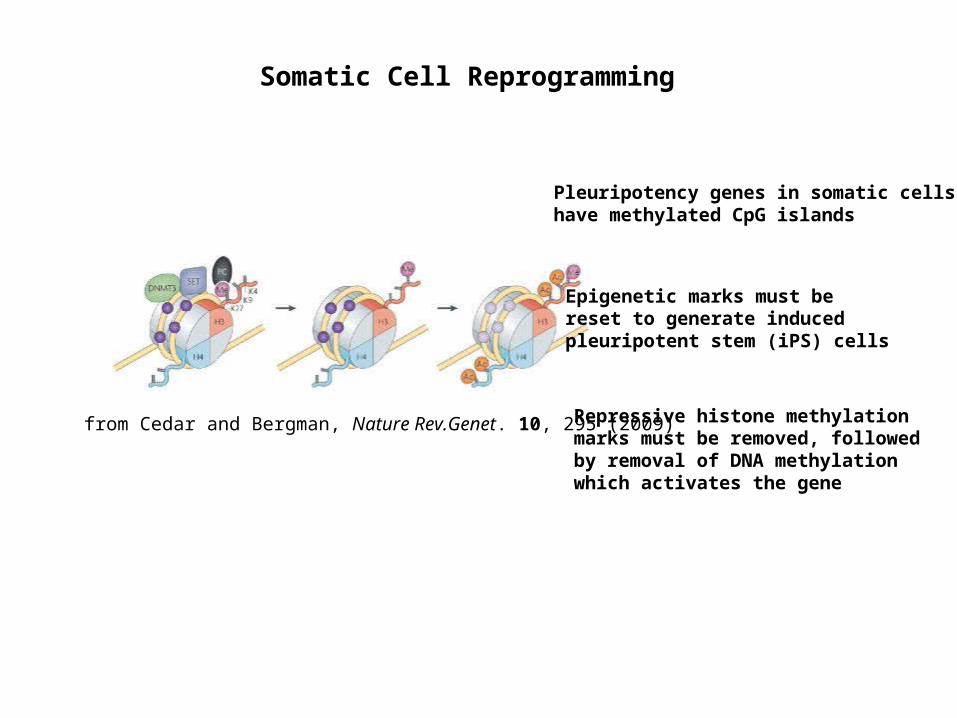
Somatic Cell Reprogramming
from Cedar and Bergman, Nature Rev.Genet. 10, 295 (2009)
Pleuripotency genes in somatic cells have methylated CpG islands
Epigenetic marks must be reset to generate induced pleuripotent stem (iPS) cells
Repressive histone methylation marks must be removed, followed by removal of DNA methylation which activates the gene
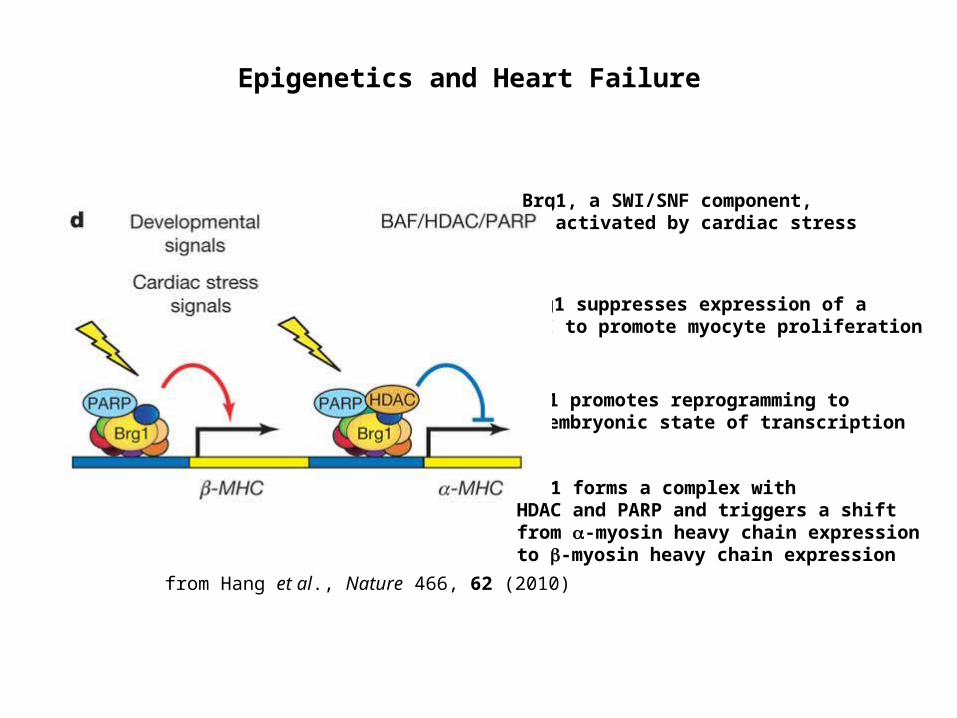
Brg1, a SWI/SNF component, is activated by cardiac stress
Brg1 suppresses expression of a CKI to promote myocyte proliferation
Brg1 forms a complex with HDAC and PARP and triggers a shift from -myosin heavy chain expression to -myosin heavy chain expression
Epigenetics and Heart Failure
from Hang et al., Nature 466, 62 (2010)
Brg1 promotes reprogramming to an embryonic state of transcription
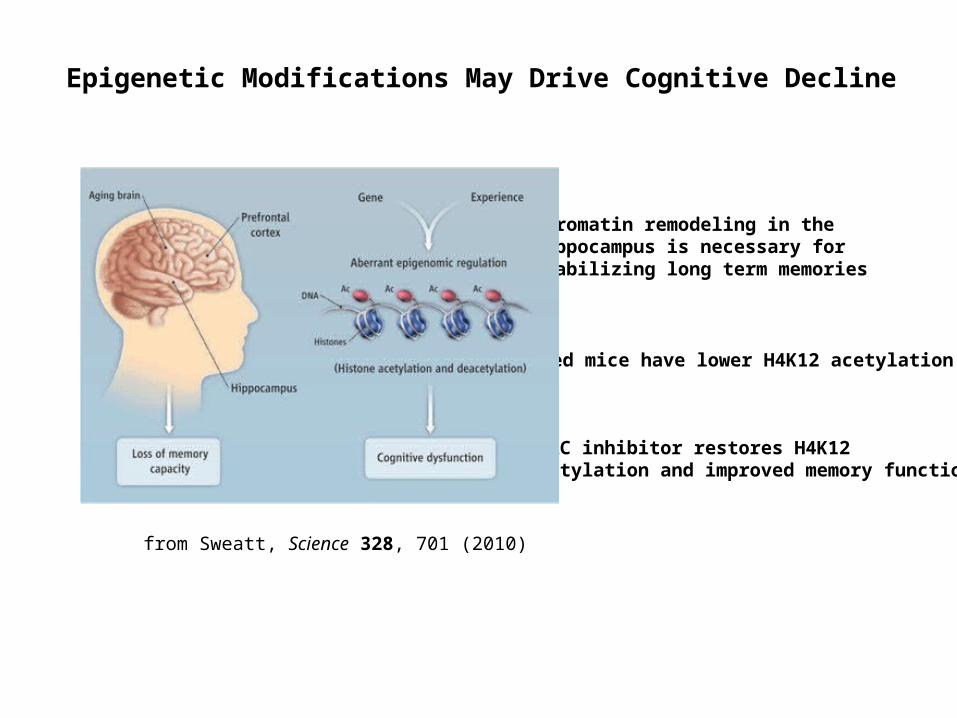
Epigenetic Modifications May Drive Cognitive Decline
from Sweatt, Science 328, 701 (2010)
Chromatin remodeling in the hippocampus is necessary for stabilizing long term memories
Aged mice have lower H4K12 acetylation
HDAC inhibitor restores H4K12 acetylation and improved memory function
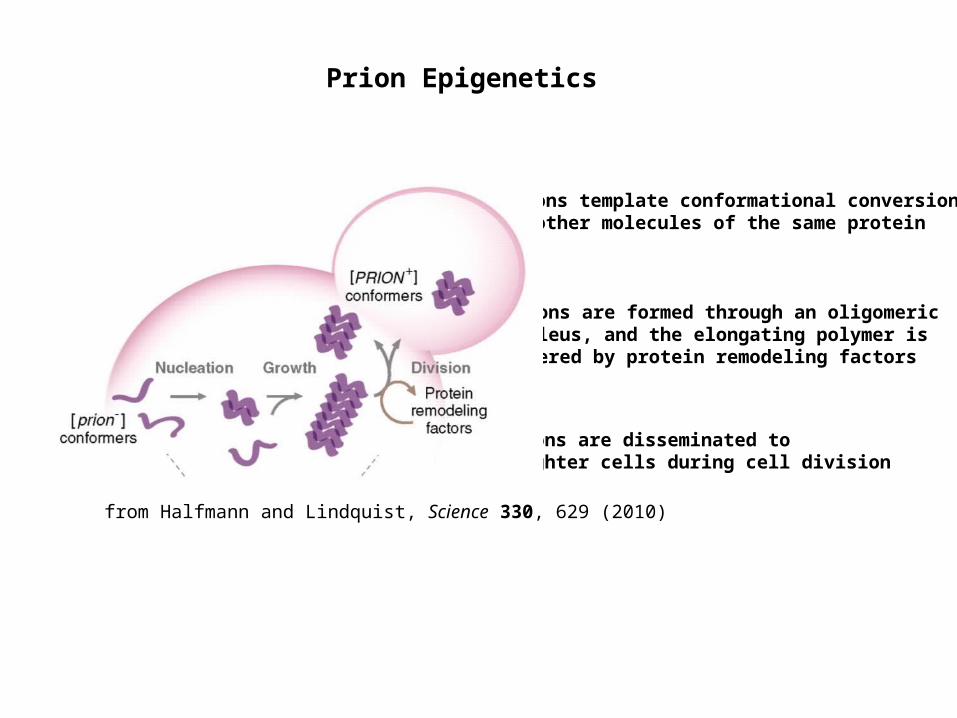
Prion Epigenetics
Prions template conformational conversion of other molecules of the same protein
Prions are disseminated to daughter cells during cell division
Prions are formed through an oligomeric nucleus, and the elongating polymer is severed by protein remodeling factors
from Halfmann and Lindquist, Science 330, 629 (2010)
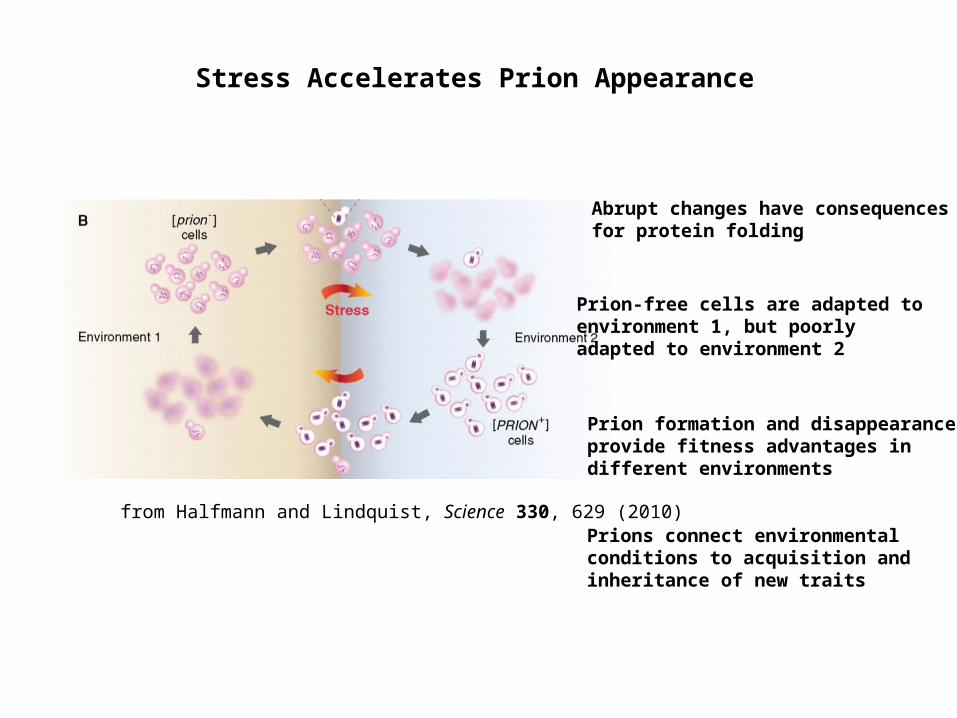
Stress Accelerates Prion Appearance
from Halfmann and Lindquist, Science 330, 629 (2010)
Prion-free cells are adapted to environment 1, but poorly adapted to environment 2
Prion formation and disappearance provide fitness advantages in different environments
Abrupt changes have consequences for protein folding
Prions connect environmental conditions to acquisition and inheritance of new traits

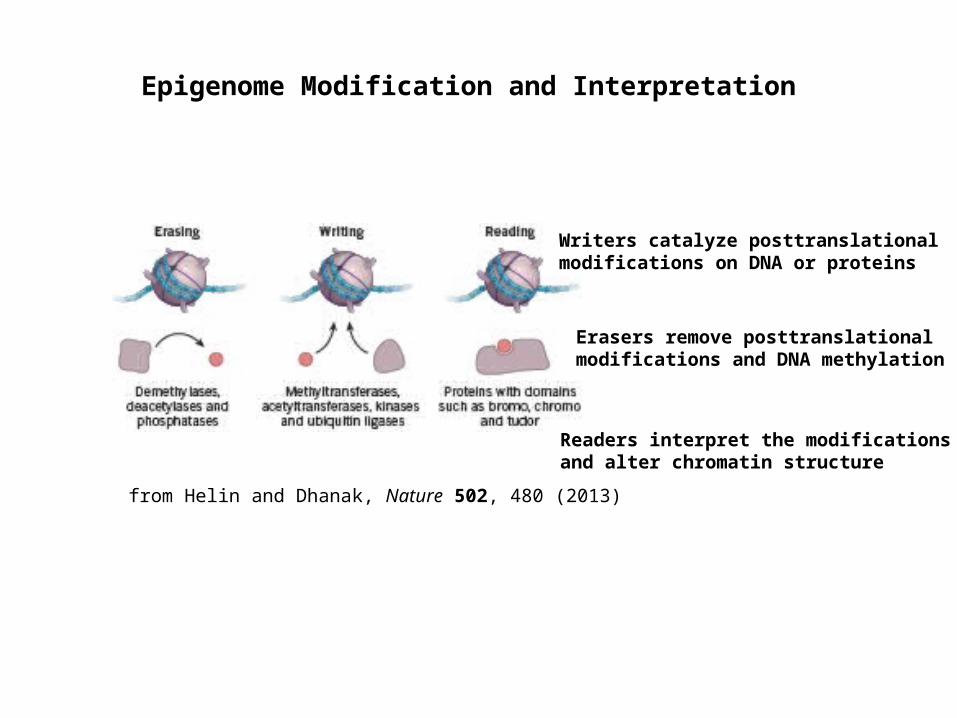
Epigenome Modification and Interpretation
Erasers remove posttranslational modifications and DNA methylation
Writers catalyze posttranslational modifications on DNA or proteins
Readers interpret the modifications and alter chromatin structure
from Helin and Dhanak, Nature 502, 480 (2013)
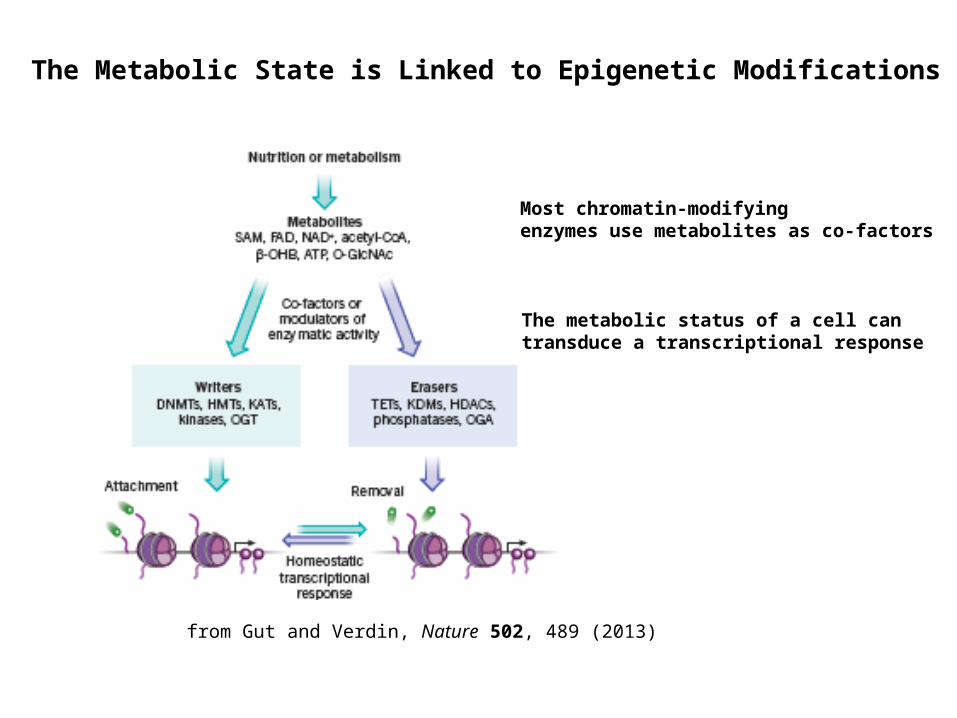
Most chromatin-modifying enzymes use metabolites as co-factors
The metabolic status of a cell can transduce a transcriptional response
The Metabolic State is Linked to Epigenetic Modifications
from Gut and Verdin, Nature 502, 489 (2013)
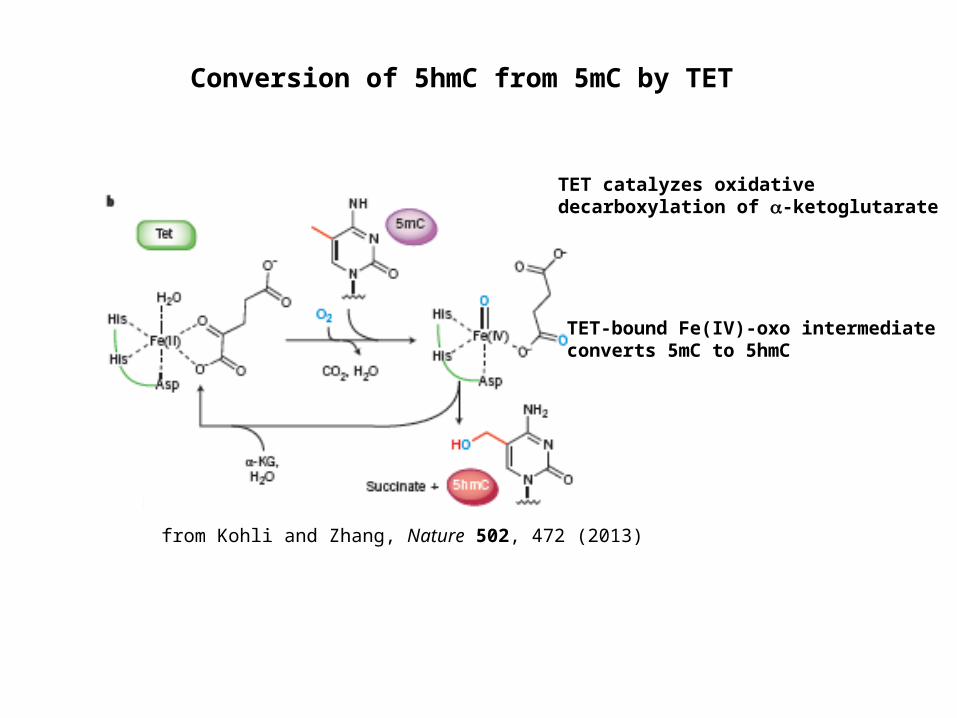
Conversion of 5hmC from 5mC by TET
TET catalyzes oxidative decarboxylation of -ketoglutarate
TET-bound Fe(IV)-oxo intermediate converts 5mC to 5hmC
from Kohli and Zhang, Nature 502, 472 (2013)
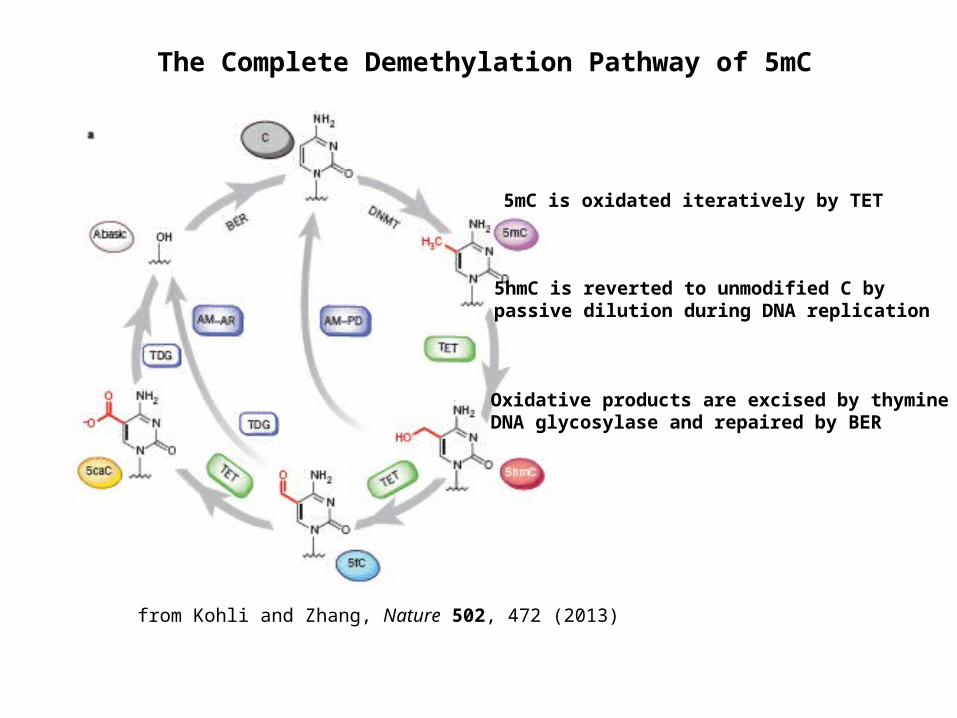
The Complete Demethylation Pathway of 5mC
from Kohli and Zhang, Nature 502, 472 (2013)
5mC is oxidated iteratively by TET
5hmC is reverted to unmodified C by passive dilution during DNA replication
Oxidative products are excised by thymine DNA glycosylase and repaired by BER
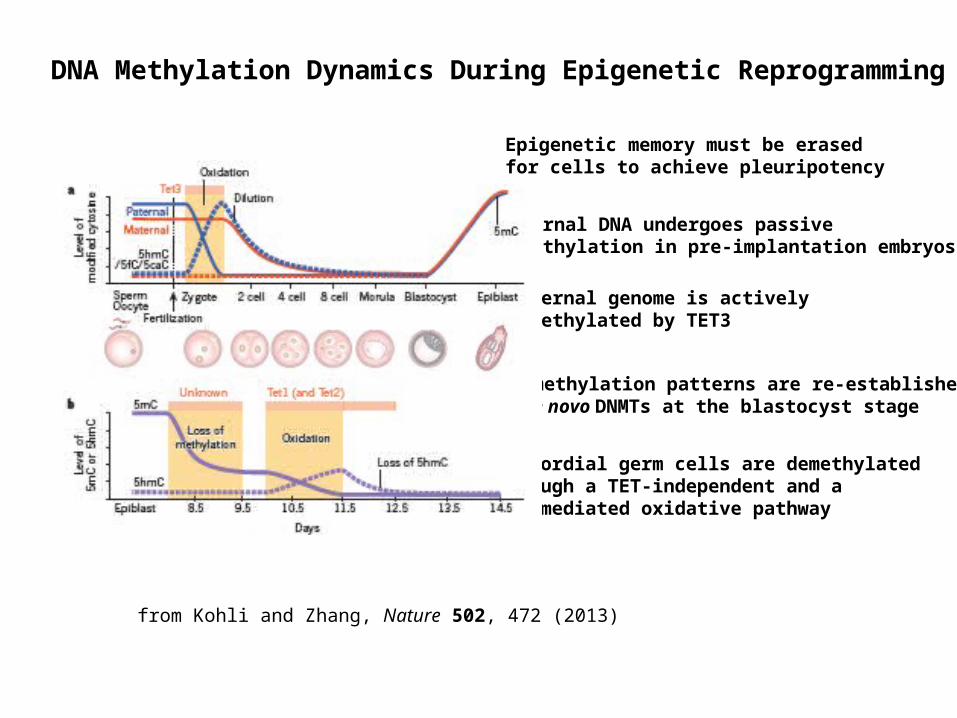
DNA Methylation Dynamics During Epigenetic Reprogramming
Maternal DNA undergoes passive demethylation in pre-implantation embryos
Paternal genome is actively demethylated by TET3
DNA methylation patterns are re-established by de novo DNMTs at the blastocyst stage
Primordial germ cells are demethylated through a TET-independent and a TET-mediated oxidative pathway
from Kohli and Zhang, Nature 502, 472 (2013)
Epigenetic memory must be erased for cells to achieve pleuripotency
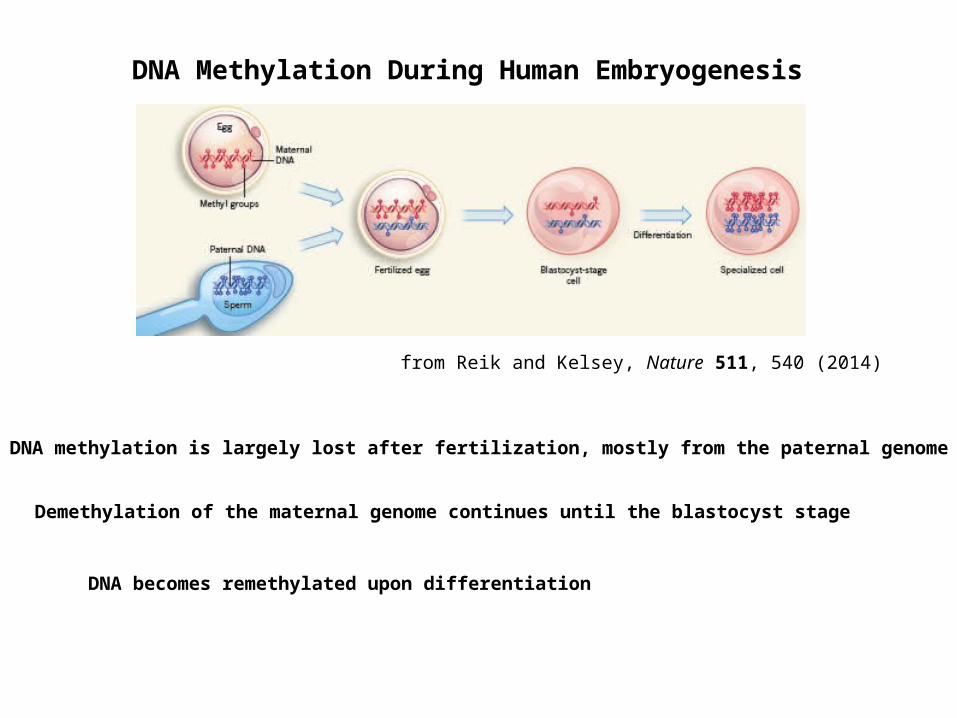
DNA Methylation During Human Embryogenesis
from Reik and Kelsey, Nature 511, 540 (2014)
DNA methylation is largely lost after fertilization, mostly from the paternal genome
Demethylation of the maternal genome continues until the blastocyst stage
DNA becomes remethylated upon differentiation





























