Embed Size (px)
Citation preview

Epigenetics, nutrition and bowel cancer risk
John Mathers Human Nutrition Research Centre
UK

h"p://www.wcrf-‐uk.org/research_science/expert_report.lasso
• WCRF/ AICR Report
• Comprehensive and systemaEc assessment of epidemiological evidence
• 2nd ediEon November 2007
Diet and bowel cancer risk .1
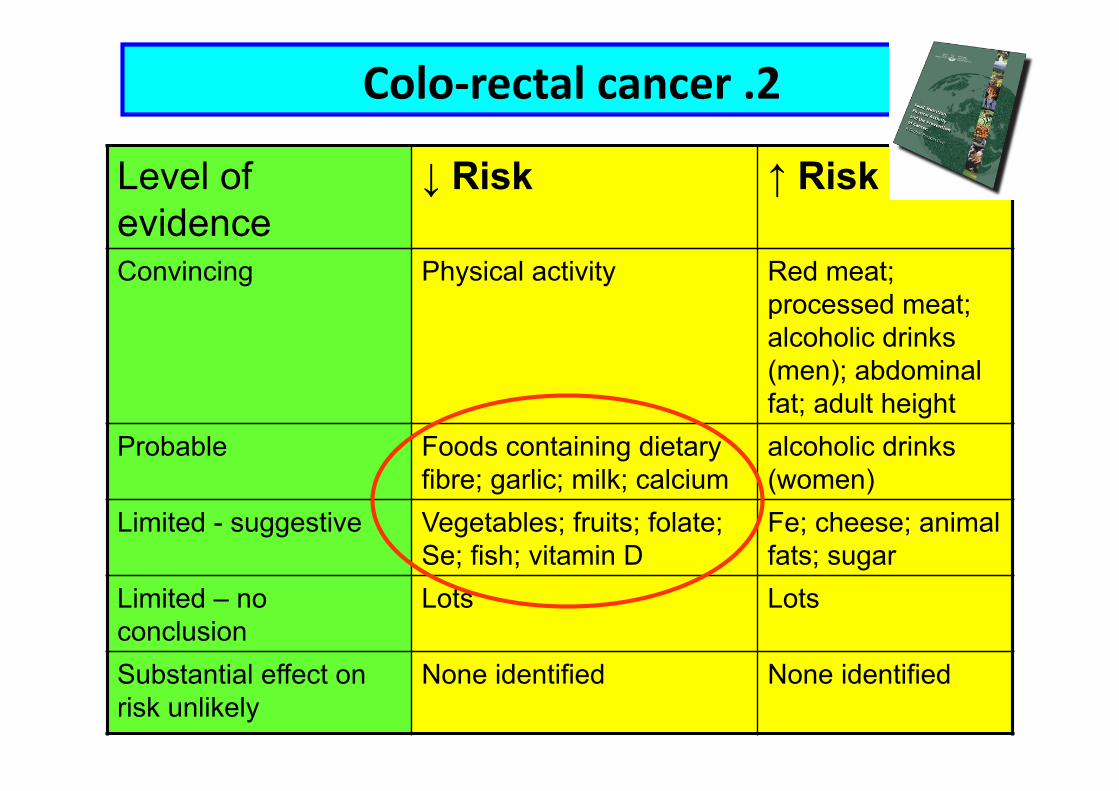
Level of evidence
↓ Risk ↑ Risk
Convincing Physical activity Red meat; processed meat; alcoholic drinks (men); abdominal fat; adult height
Probable Foods containing dietary fibre; garlic; milk; calcium
alcoholic drinks (women)
Limited - suggestive Vegetables; fruits; folate; Se; fish; vitamin D
Fe; cheese; animal fats; sugar
Limited – no conclusion
Lots Lots
Substantial effect on risk unlikely
None identified None identified
Colo-‐rectal cancer .2

Overview of lecture
Overview of epigenetic mechanisms
Epigenetic events in bowel cancer
Towards novel diet-related DNA methylation biomarkers of bowel cancer risk
MicroRNA, diet and development of bowel cancer
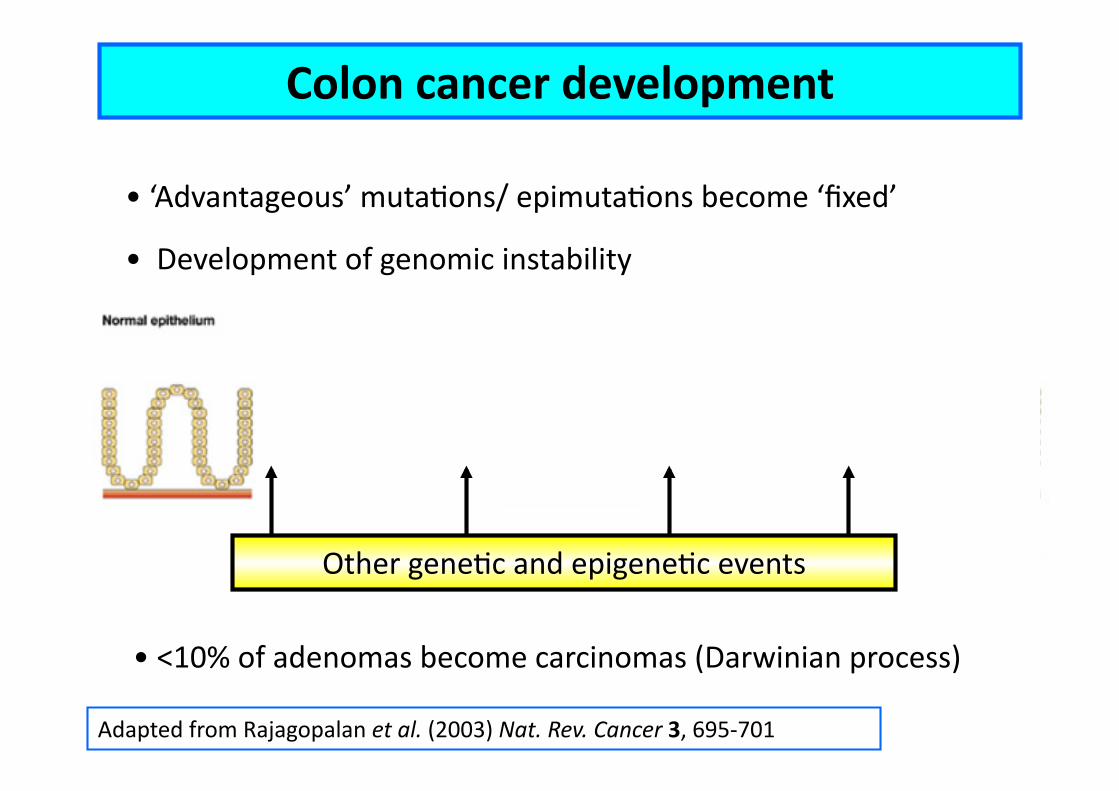
Adapted from Rajagopalan et al. (2003) Nat. Rev. Cancer 3, 695-‐701
Colon cancer development
• ‘Advantageous’ mutaEons/ epimutaEons become ‘fixed’
• Development of genomic instability
• <10% of adenomas become carcinomas (Darwinian process)
Other geneEc and epigeneEc events
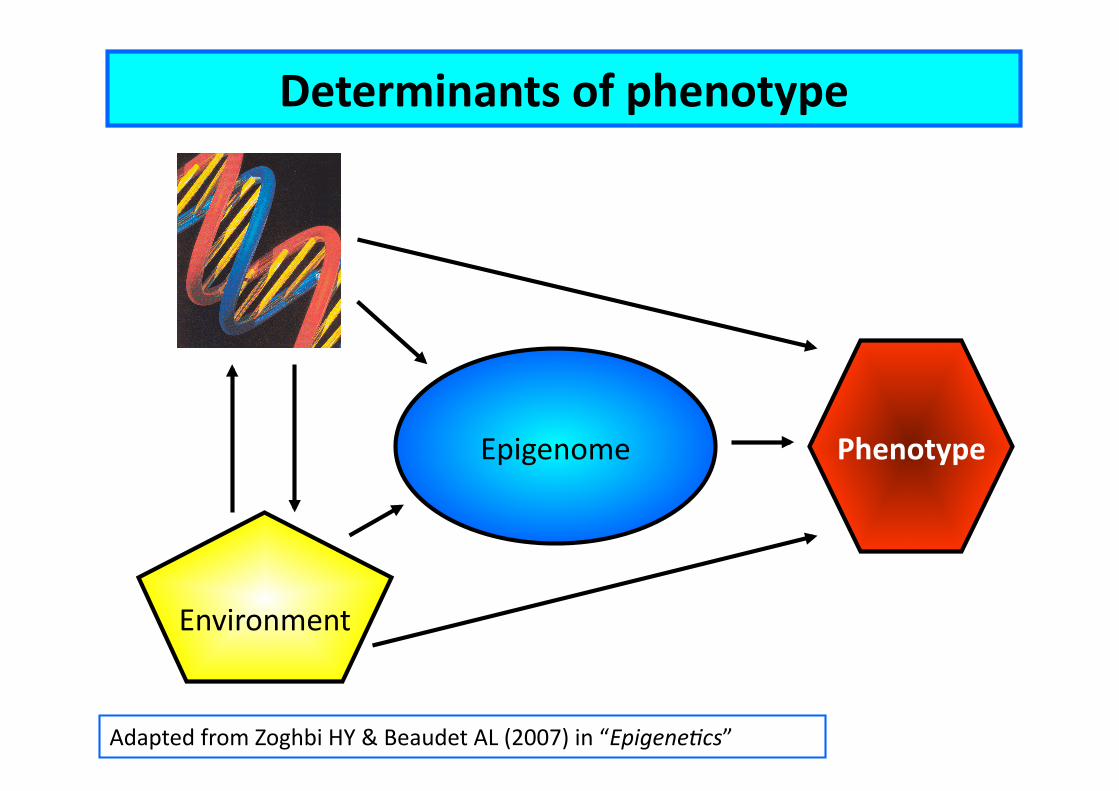
Environment
Phenotype Epigenome
Adapted from Zoghbi HY & Beaudet AL (2007) in “Epigene2cs”
Determinants of phenotype
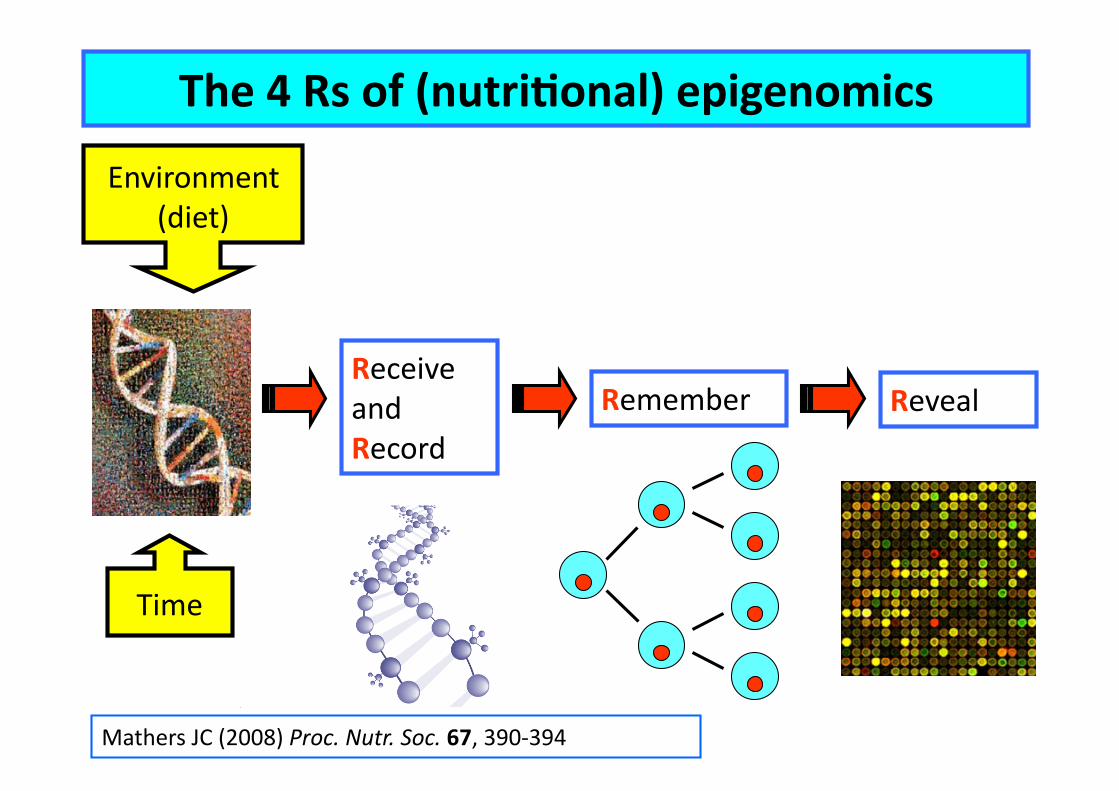
Environment (diet)
Receive and Record
Time
Reveal
The 4 Rs of (nutriConal) epigenomics
Mathers JC (2008) Proc. Nutr. Soc. 67, 390-‐394
Remember
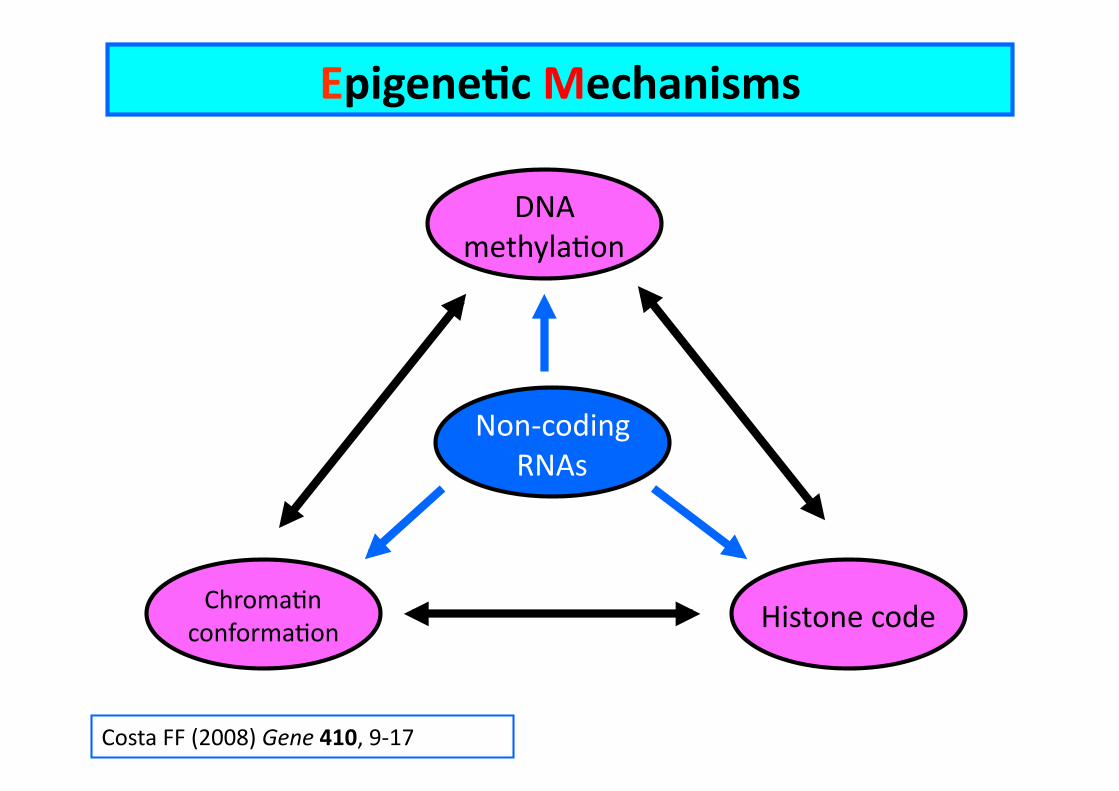
Costa FF (2008) Gene 410, 9-‐17
EpigeneCc Mechanisms
DNA methylaEon
Histone code ChromaEn conformaEon
Non-‐coding RNAs
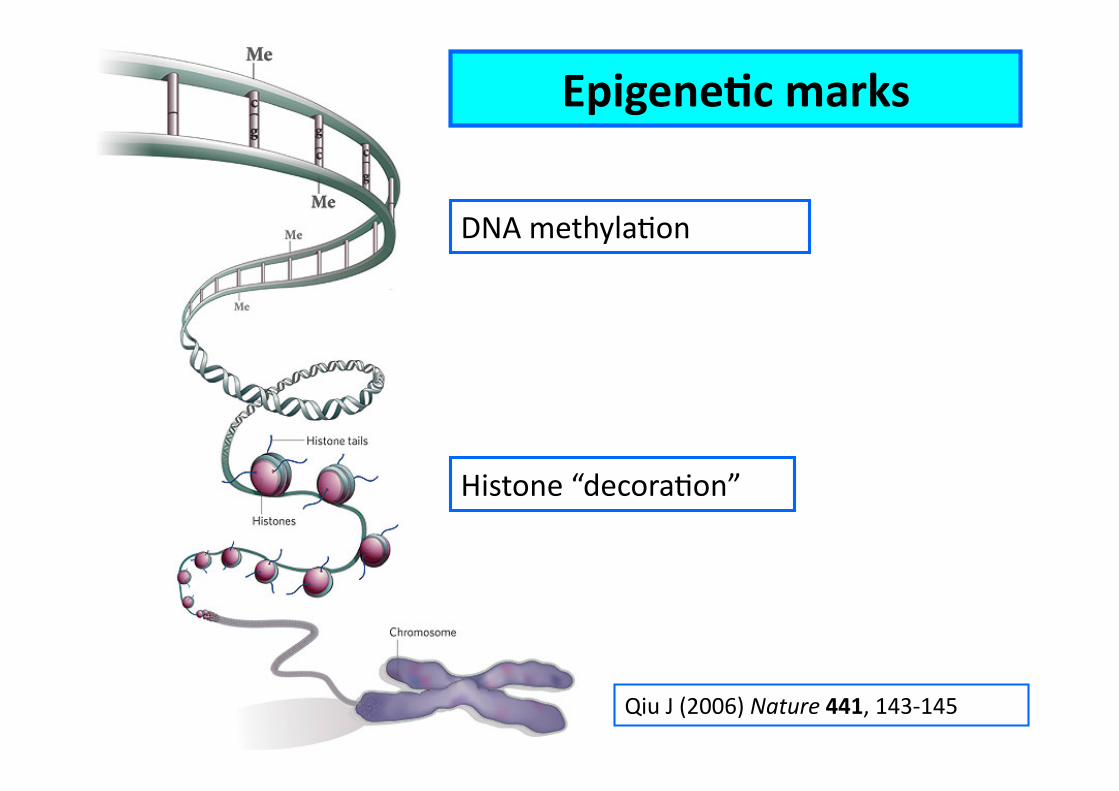
EpigeneCc marks
DNA methylaEon
Histone “decoraEon”
Qiu J (2006) Nature 441, 143-‐145

Molecular mechanisms linking diet with bowel cancer risk
All cancers arise from (unrepaired) genomic damage so “protecCve” dietary factors “must”:
• ↓ genomic damage
• ↑ genomic repair
• ↑ removal of damaged cells by apoptosis
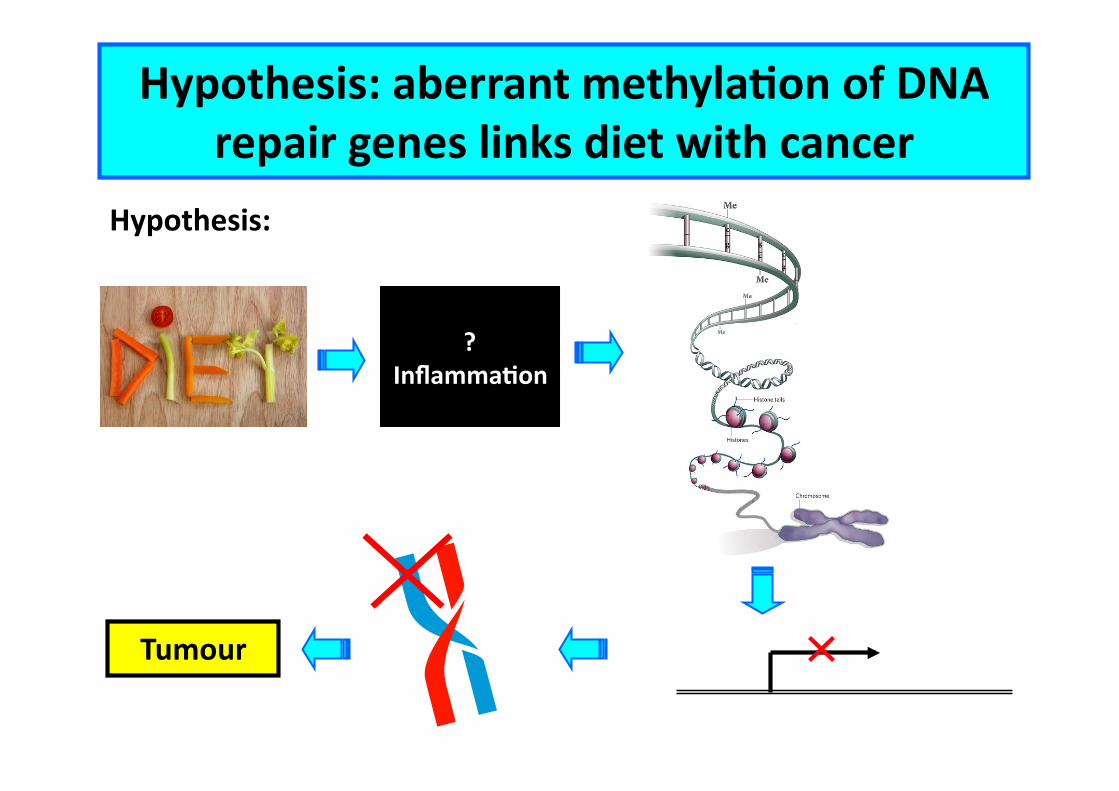
Hypothesis: aberrant methylaCon of DNA repair genes links diet with cancer
Hypothesis:
? InflammaCon
Tumour
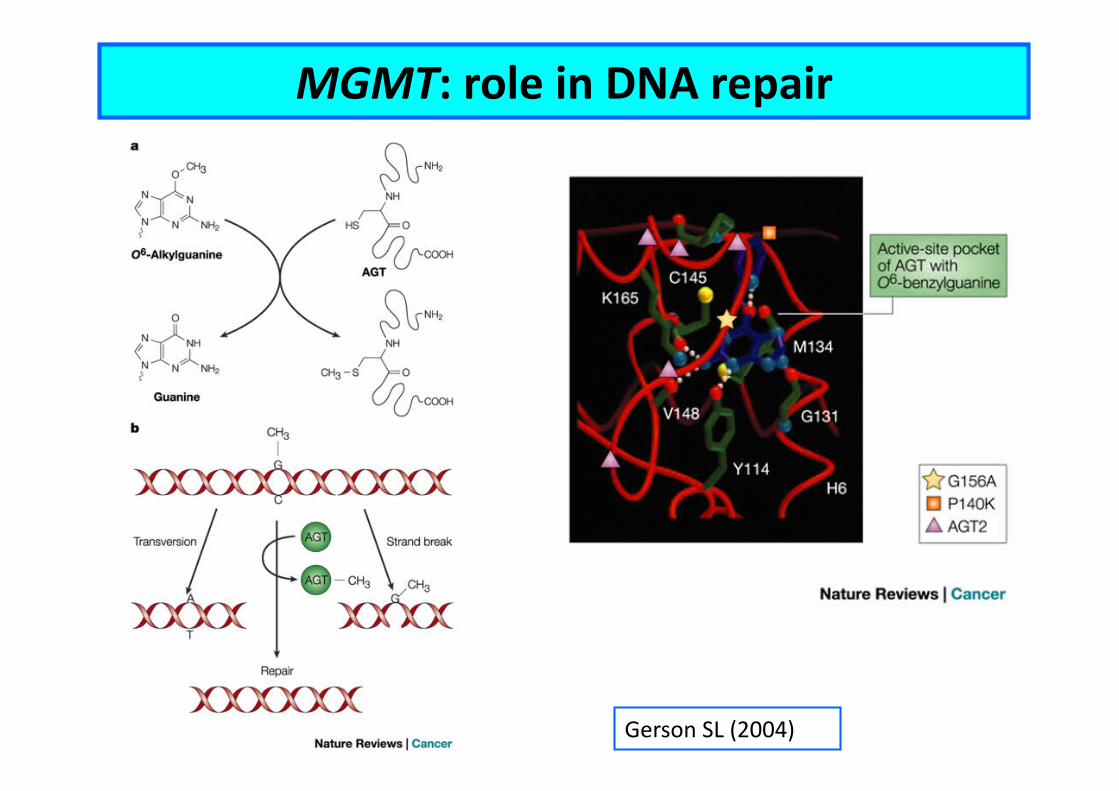
Gerson SL (2004)
MGMT: role in DNA repair

Jones PA & Baylin SB (2002)
EpigeneCc events in cancer development – silencing of DNA repair genes
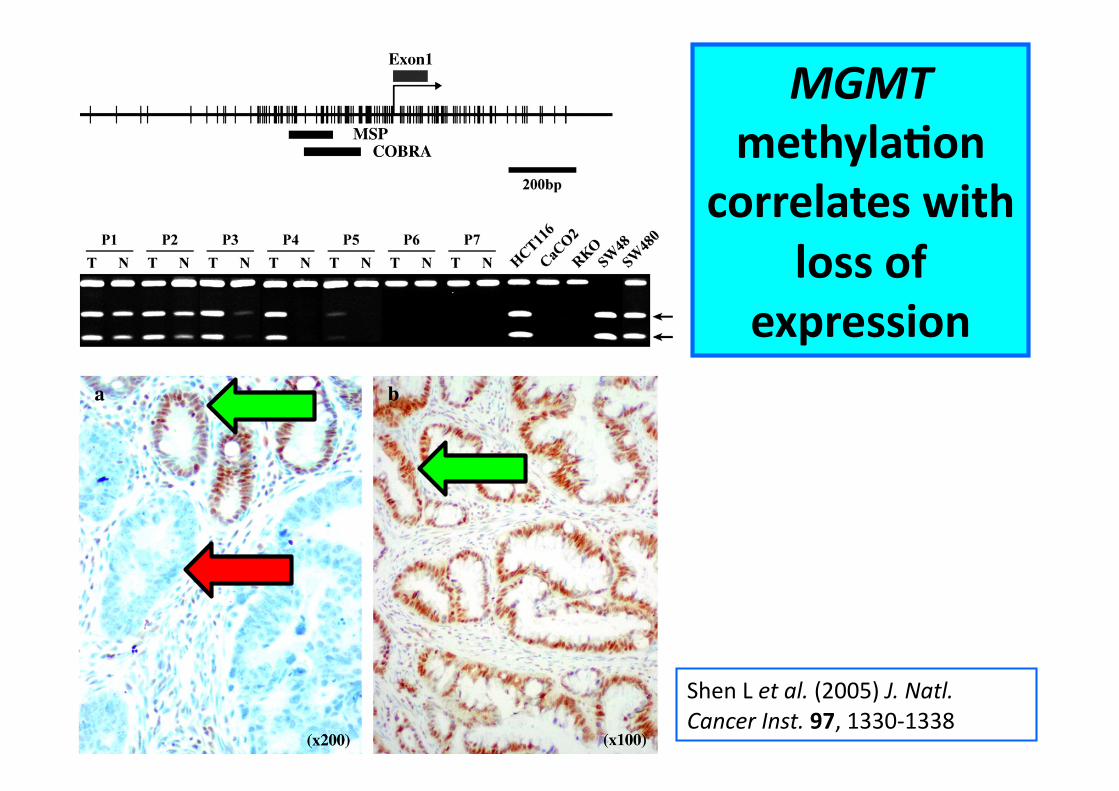
Shen L et al. (2005) J. Natl. Cancer Inst. 97, 1330-‐1338
MGMT methylaCon
correlates with loss of
expression
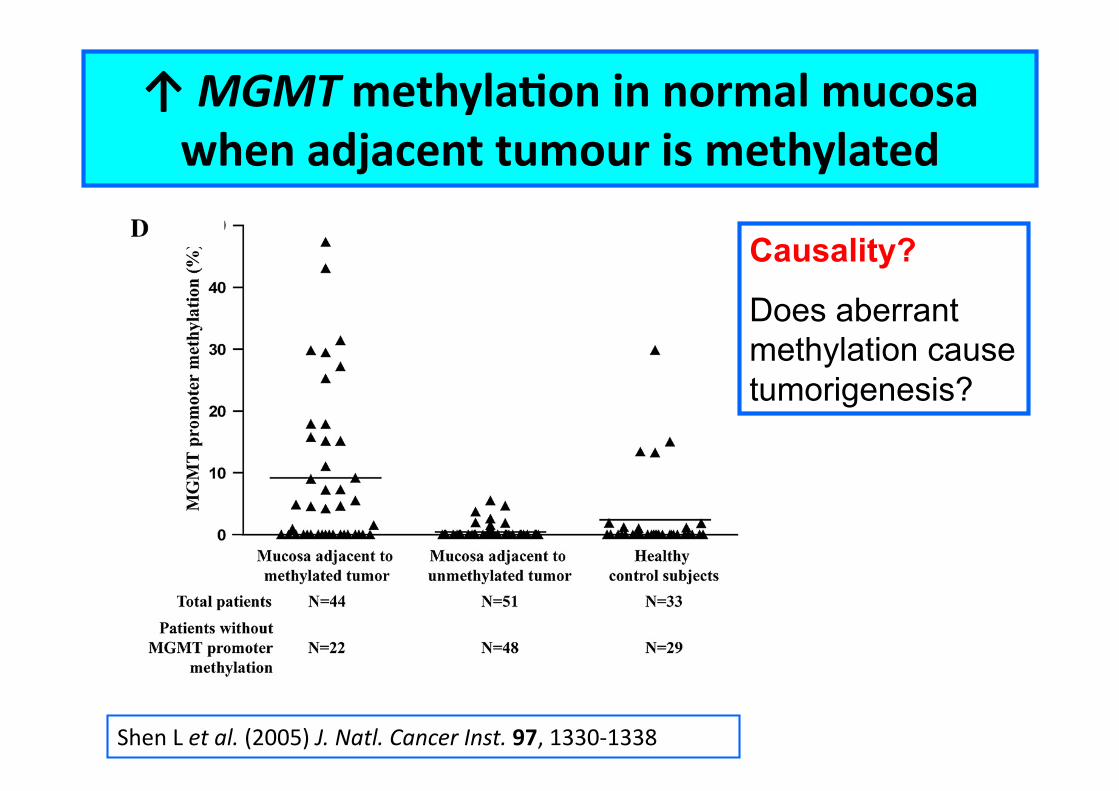
Shen L et al. (2005) J. Natl. Cancer Inst. 97, 1330-‐1338
↑ MGMT methylaCon in normal mucosa when adjacent tumour is methylated
Causality?
Does aberrant methylation cause tumorigenesis?
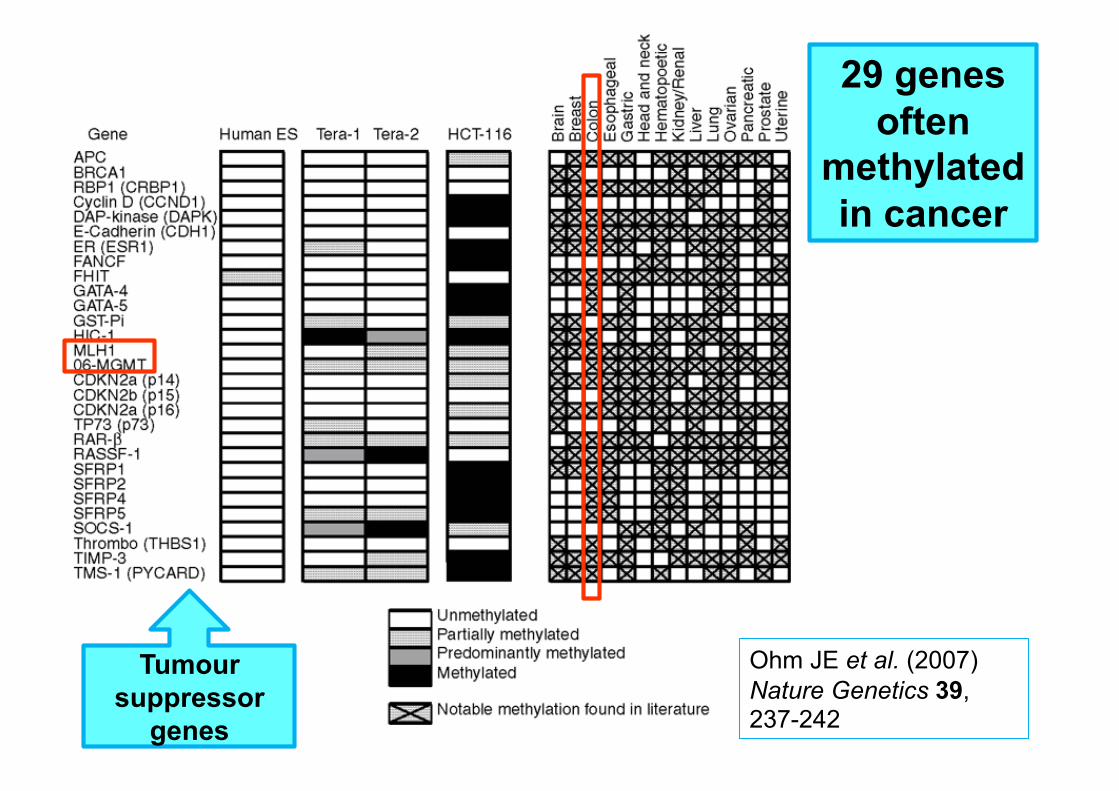
29 genes often
methylated in cancer
Ohm JE et al. (2007) Nature Genetics 39, 237-242
Tumour suppressor
genes
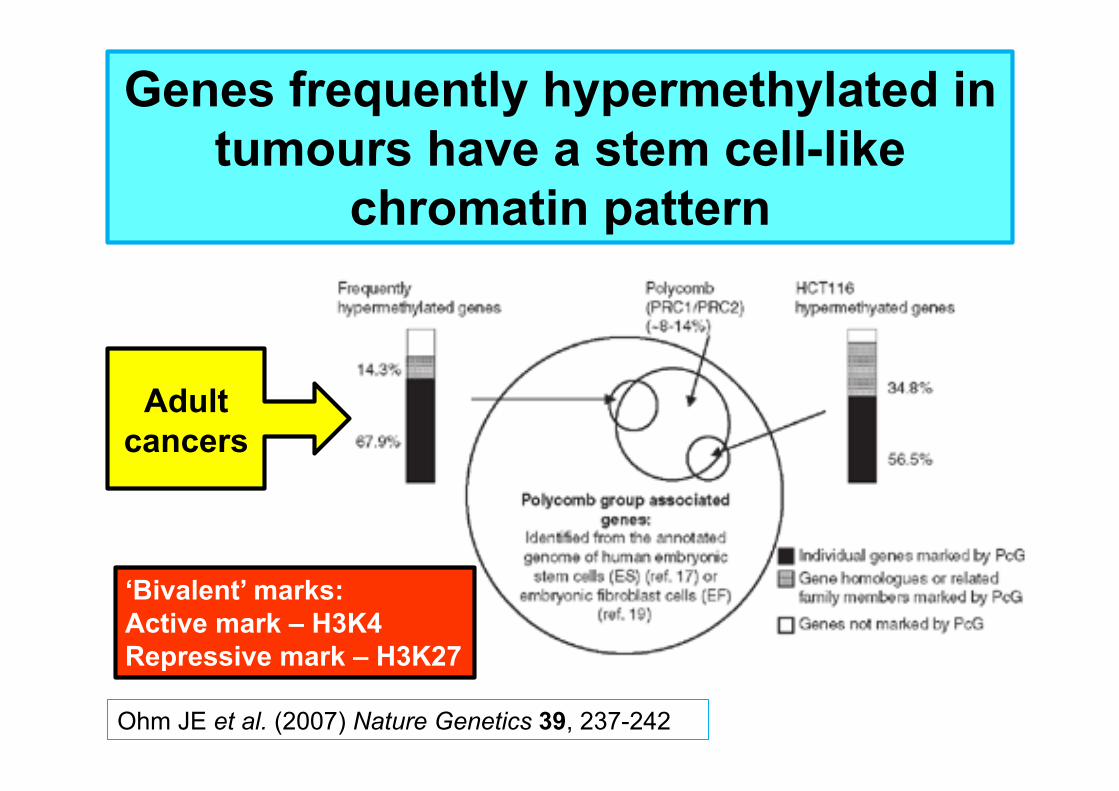
Ohm JE et al. (2007) Nature Genetics 39, 237-242
Genes frequently hypermethylated in tumours have a stem cell-like
chromatin pattern
Adult cancers
‘Bivalent’ marks: Active mark – H3K4 Repressive mark – H3K27
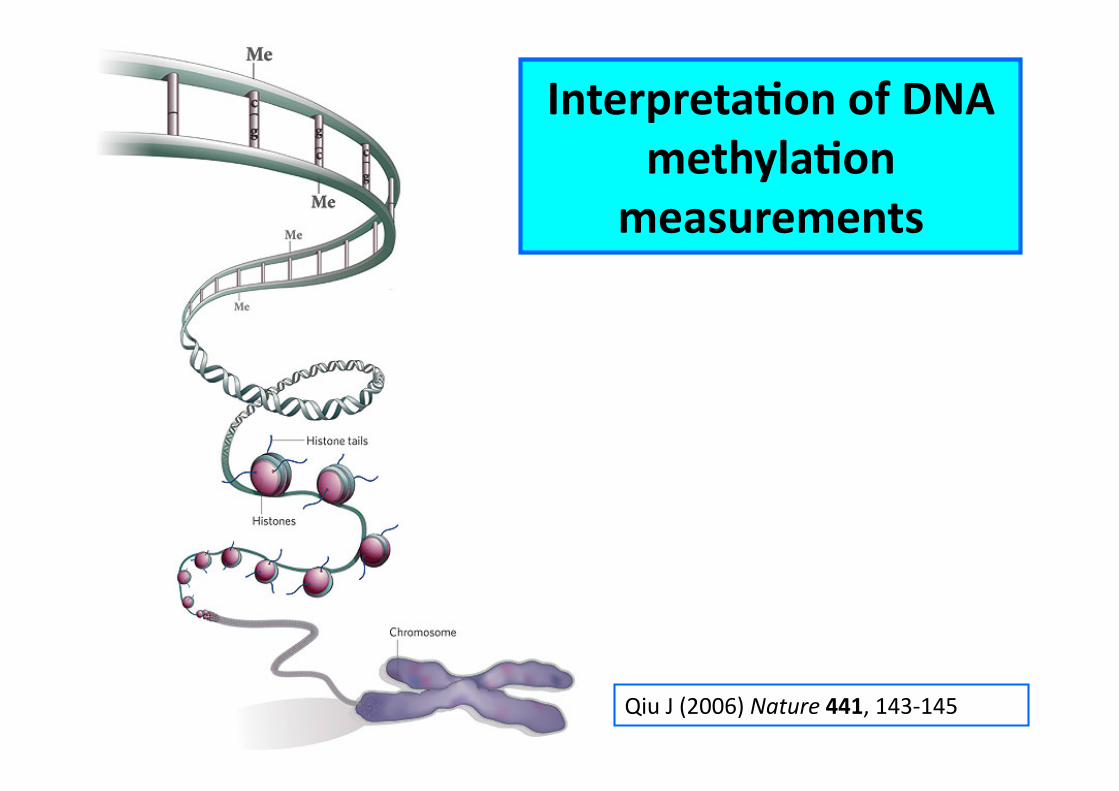
Qiu J (2006) Nature 441, 143-‐145
InterpretaCon of DNA methylaCon
measurements
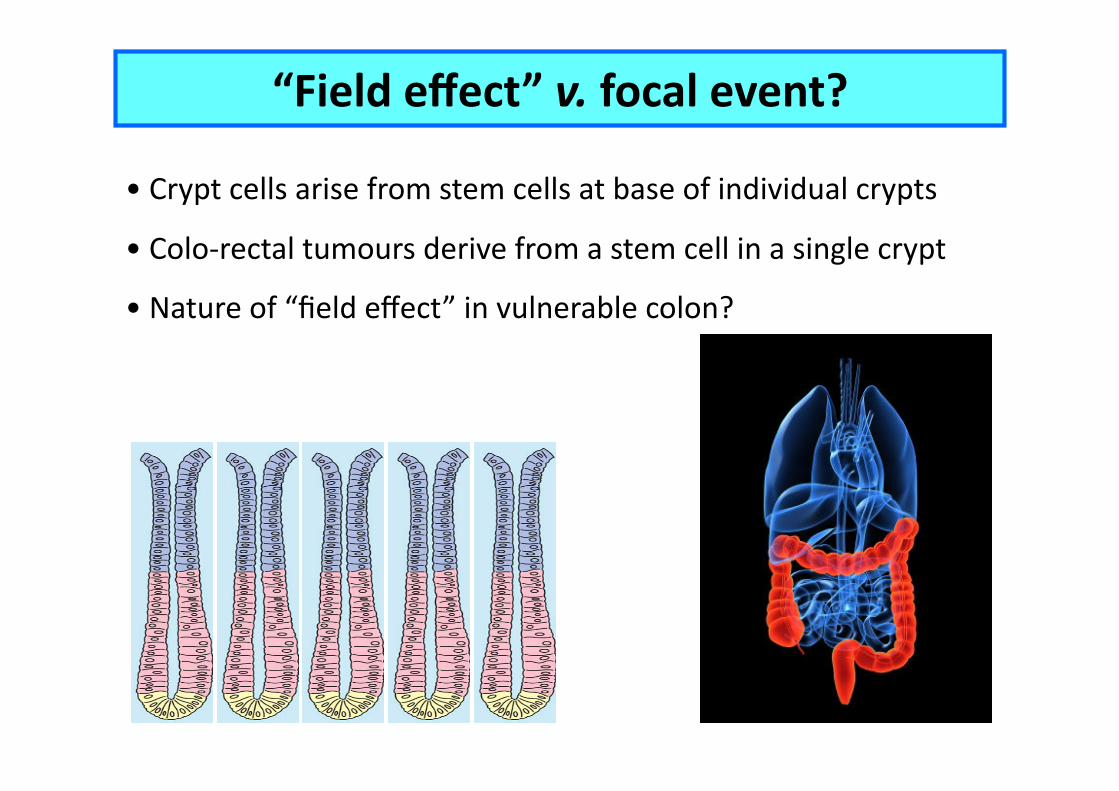
“Field effect” v. focal event?
• Crypt cells arise from stem cells at base of individual crypts
• Colo-‐rectal tumours derive from a stem cell in a single crypt
• Nature of “field effect” in vulnerable colon?
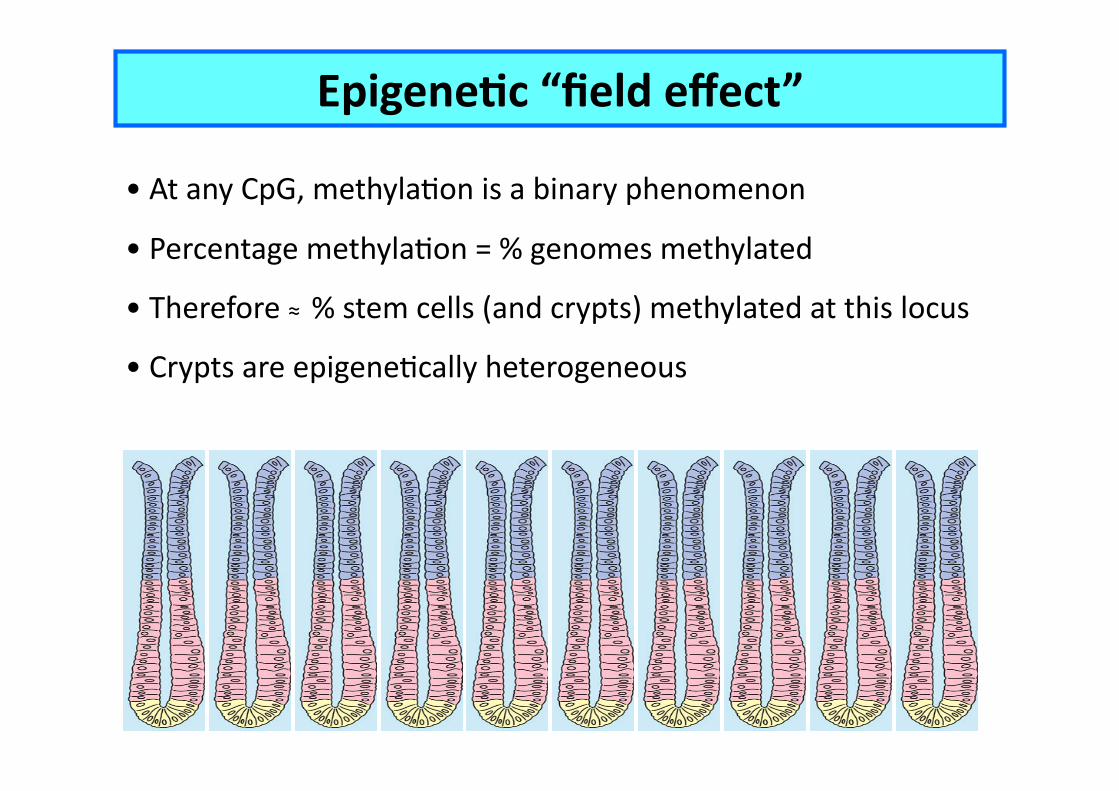
EpigeneCc “field effect”
• At any CpG, methylaEon is a binary phenomenon
• Percentage methylaEon = % genomes methylated
• Therefore ≈ % stem cells (and crypts) methylated at this locus
• Crypts are epigeneEcally heterogeneous
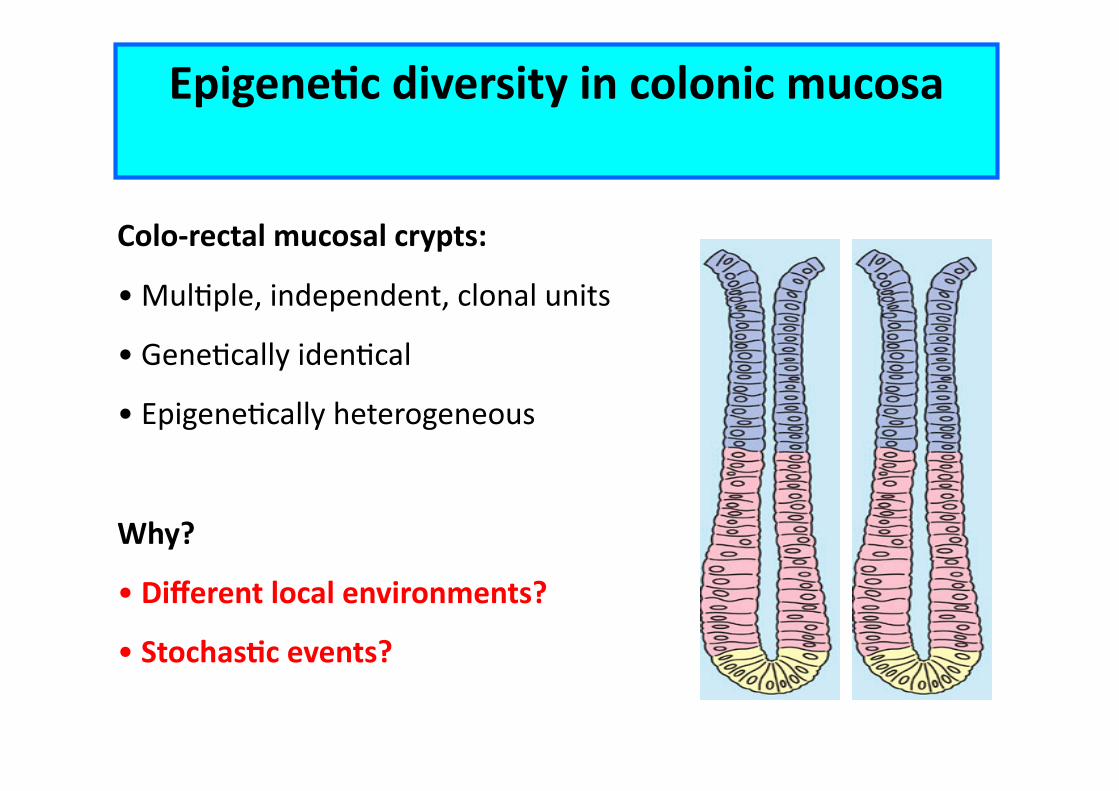
EpigeneCc diversity in colonic mucosa
Colo-‐rectal mucosal crypts:
• MulEple, independent, clonal units
• GeneEcally idenEcal • EpigeneEcally heterogeneous
Why?
• Different local environments?
• StochasCc events?
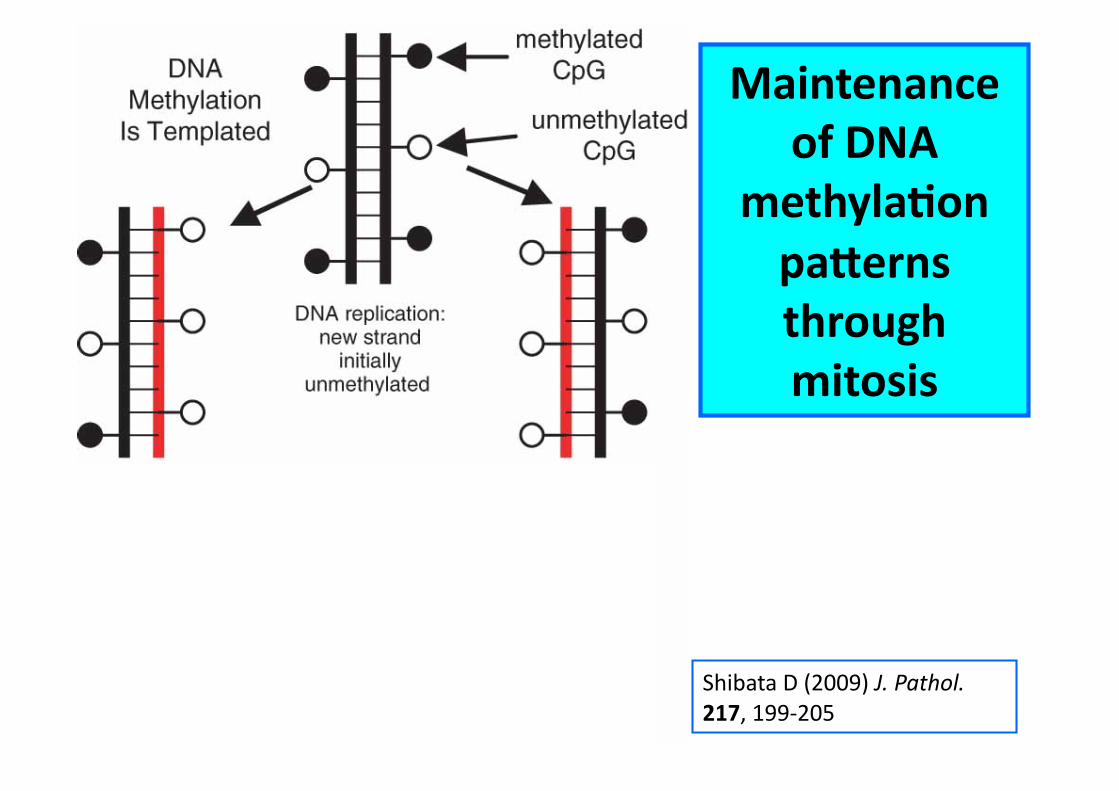
Shibata D (2009) J. Pathol. 217, 199-‐205
Maintenance of DNA
methylaCon pa^erns through mitosis
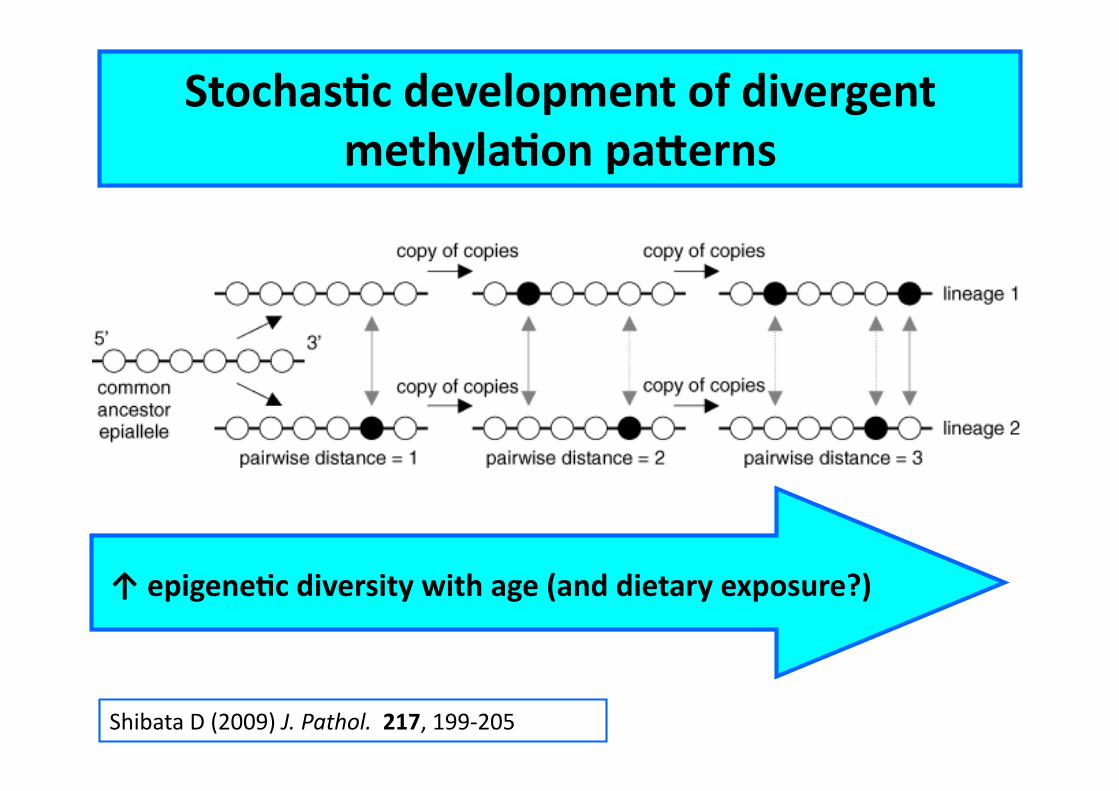
Shibata D (2009) J. Pathol. 217, 199-‐205
StochasCc development of divergent methylaCon pa^erns
↑ epigeneCc diversity with age (and dietary exposure?)
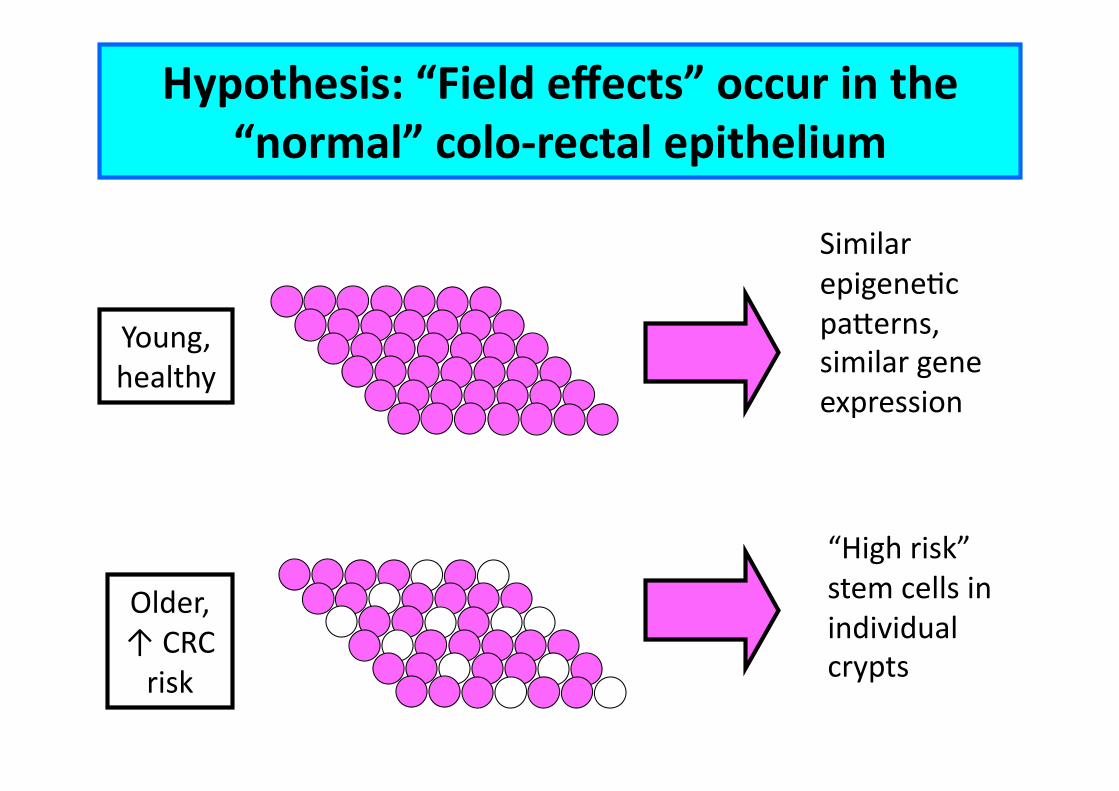
Hypothesis: “Field effects” occur in the “normal” colo-‐rectal epithelium
Young, healthy
Older, ↑ CRC risk
Similar epigeneEc pa"erns, similar gene expression
“High risk” stem cells in individual crypts
Stool as a surrogate “Cssue” for DNA methylaCon measurements
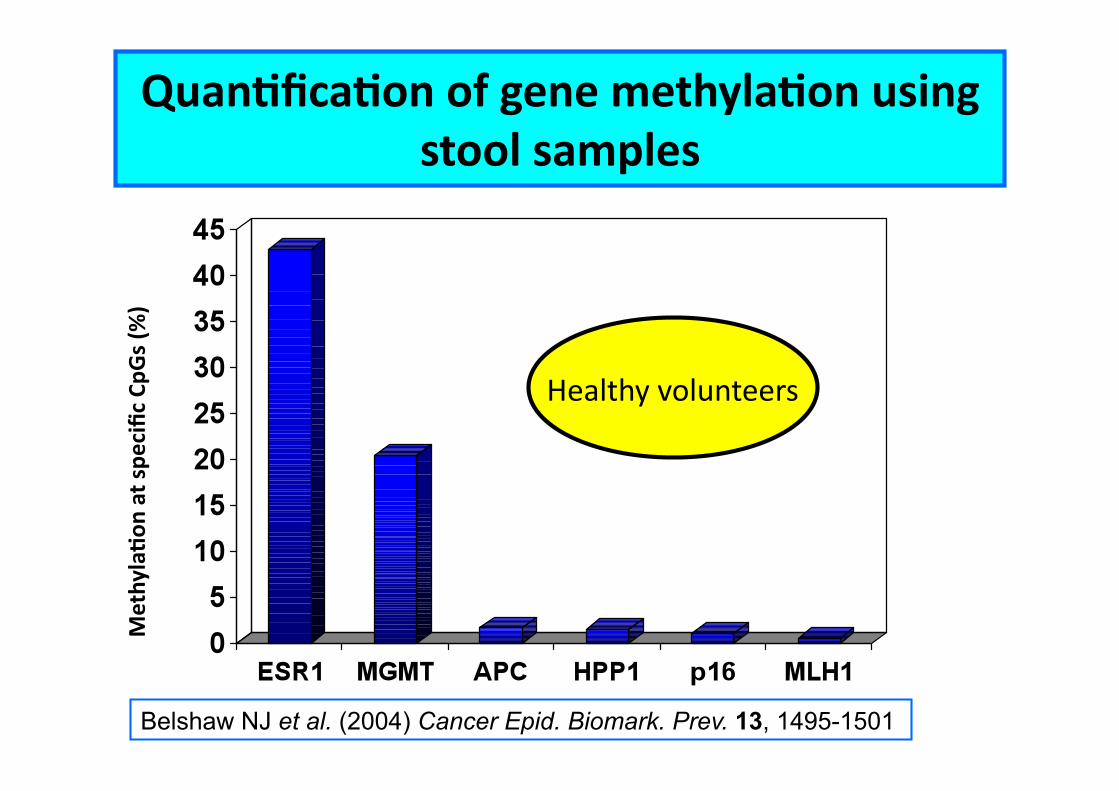
MethylaCon
at spe
cific CpG
s (%
)
QuanCficaCon of gene methylaCon using stool samples
Healthy volunteers
Belshaw NJ et al. (2004) Cancer Epid. Biomark. Prev. 13, 1495-1501
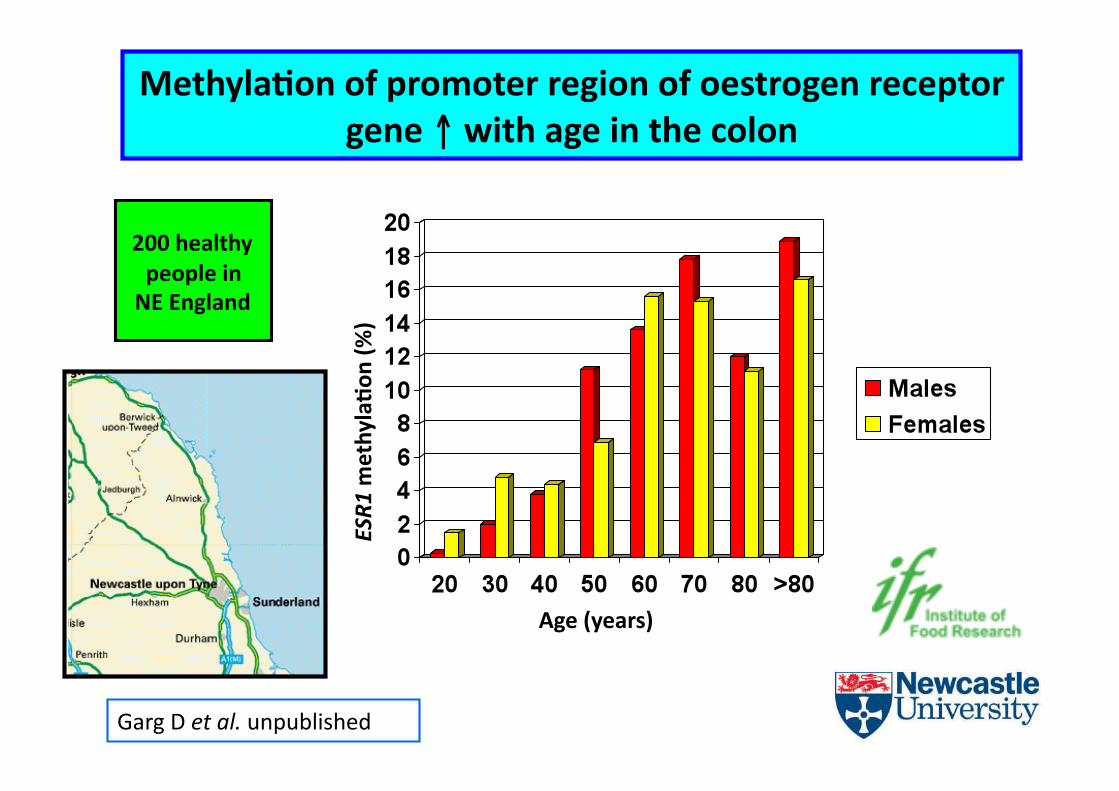
MethylaCon of promoter region of oestrogen receptor gene ↑ with age in the colon
200 healthy people in NE England
Age (years)
ESR1
methylaCon
(%)
Garg D et al. unpublished
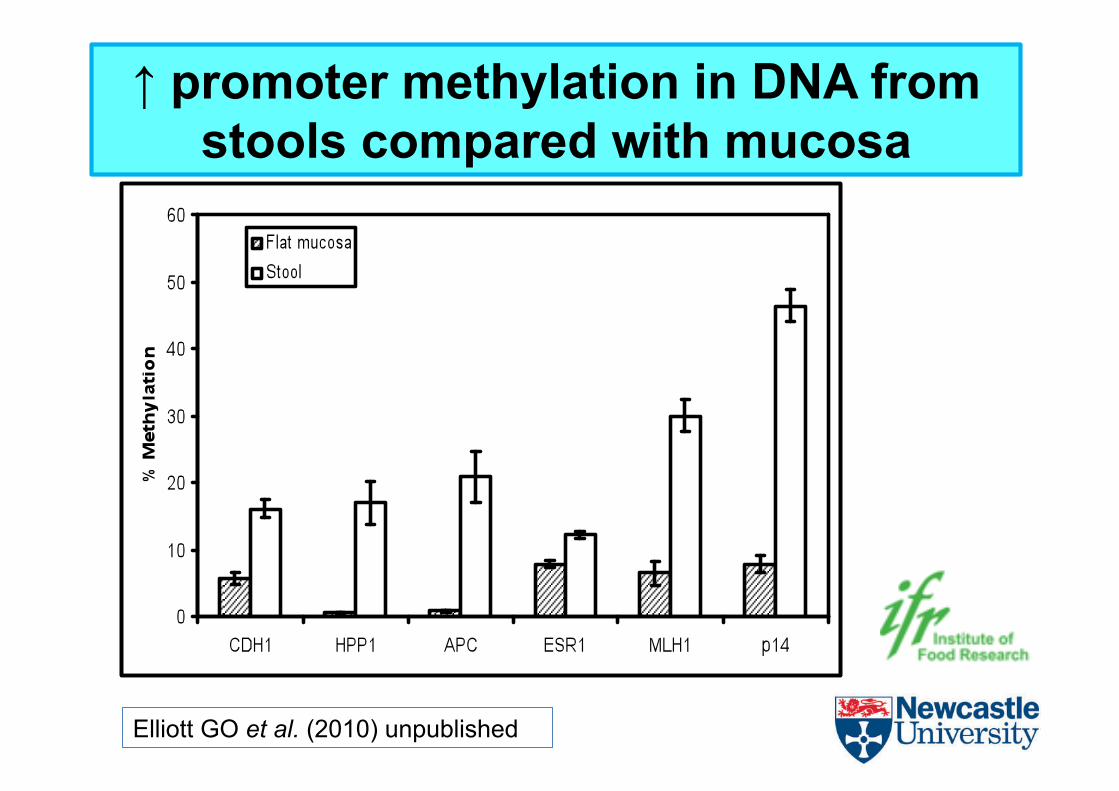
Elliott GO et al. (2010) unpublished
↑ promoter methylation in DNA from stools compared with mucosa
0
10
20
30
40
APC CDH1 HPP1 ESR1 MLH1 p14
% M
eth
0
10
20
30
40
50
60
70
80
APC CDH1 HPP1 ESR1 MLH1 p14
% M
eth
50
60
70
80
% M
eth
**
*** *** ***
***
***
***
*
C
B
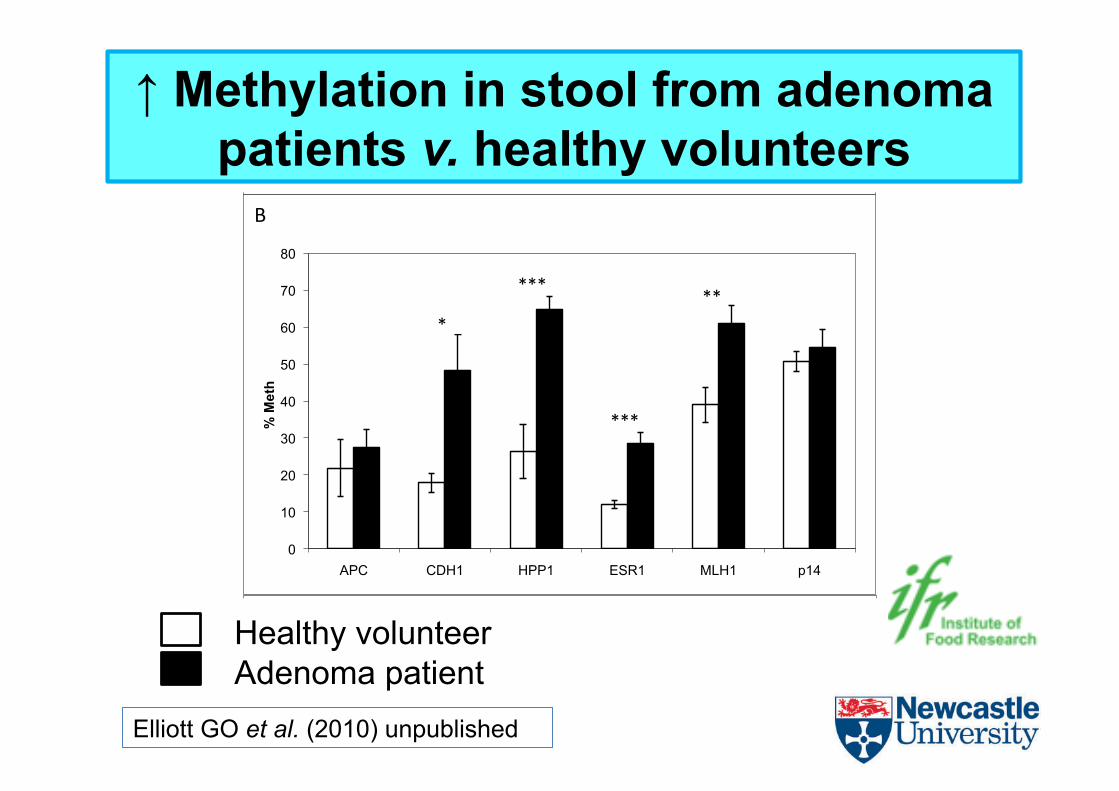
Elliott GO et al. (2010) unpublished
↑ Methylation in stool from adenoma patients v. healthy volunteers
Healthy volunteer Adenoma patient

Stool-‐based DNA methylaCon measurements
• Human DNA can be harvested from stool
• Quality of DNA is adequate for quanEficaEon of promoter methylaEon
• Some results as anEcipated e.g. age-‐dependent ↑ in ESR1 methylaEon
• Levels of methylaEon are consistently higher than those seen in corresponding mucosal biopsies
• ? DifferenEal survival of methylated sequences?
• PotenEal use in developing biomarkers of CRC risk

“All seeing, all controlling”
Claudia Bentley (2006)

MicroRNA (miRNA)
Large family of small (≈ 22 nucleotides long) non-coding RNAs;
At least 721 miRNA in human genome;
Regulate transcription of ≈30% of all protein-encoding genes through sequence-specific binding to RNA;
Inhibit translation and/or signal degradation of target mRNA;
Regulate almost all cellular processes investigated.
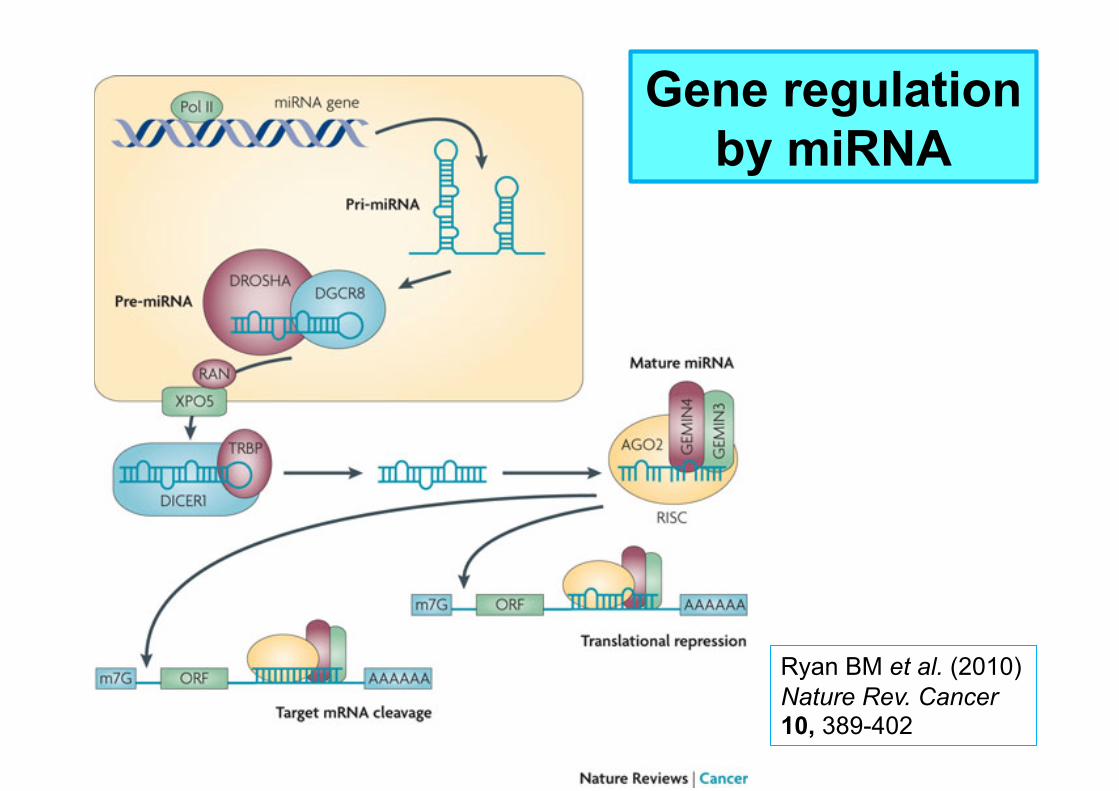
Gene regulation by miRNA
Ryan BM et al. (2010) Nature Rev. Cancer 10, 389-402

miRNA and cancer Some miRNA which are normally “silent” in adult tissues become re-expressed
Persistent stem cell-like de-differentiated state
miRNA over-expressed in tumours may act like oncogenes
↑ proliferation, ↓ apoptosis
miRNA with tumour suppressor (TS) regulatory activity may beome down regulated
loss of TS activity
Jeffrey SS (2008) Nature Biotech. 26, 400-401
Oncomirs = miRNA with a role in cancer
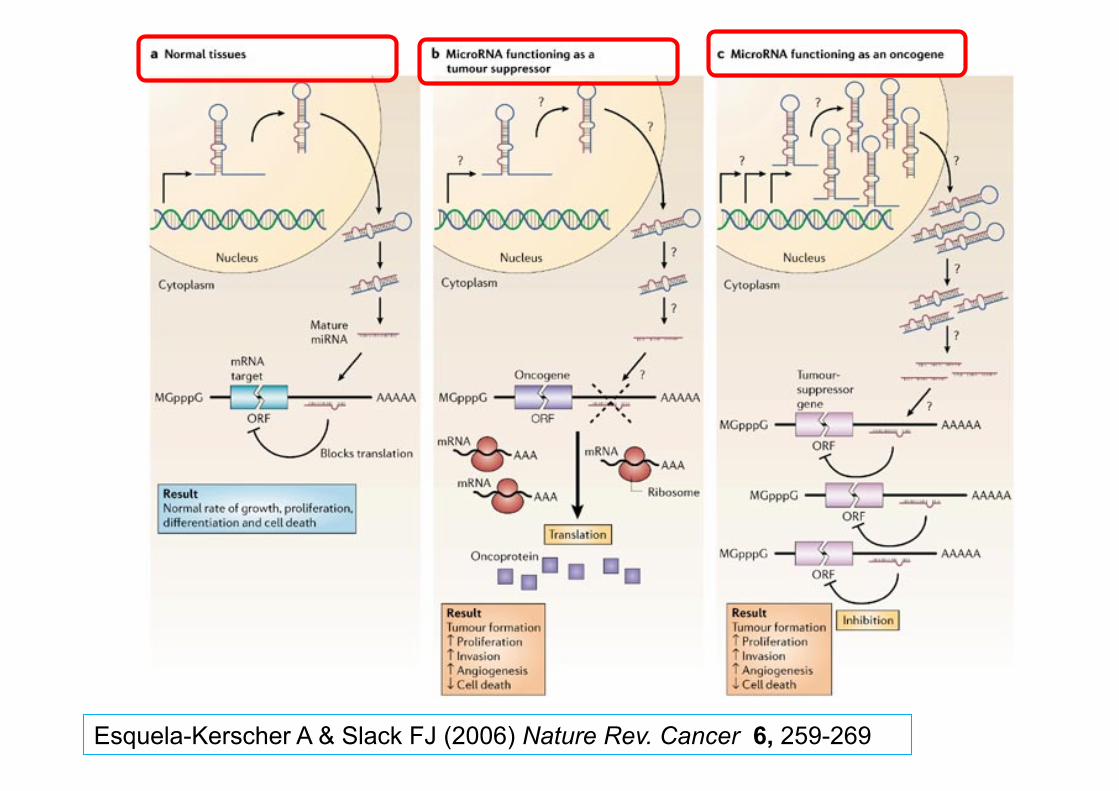
Esquela-Kerscher A & Slack FJ (2006) Nature Rev. Cancer 6, 259-269
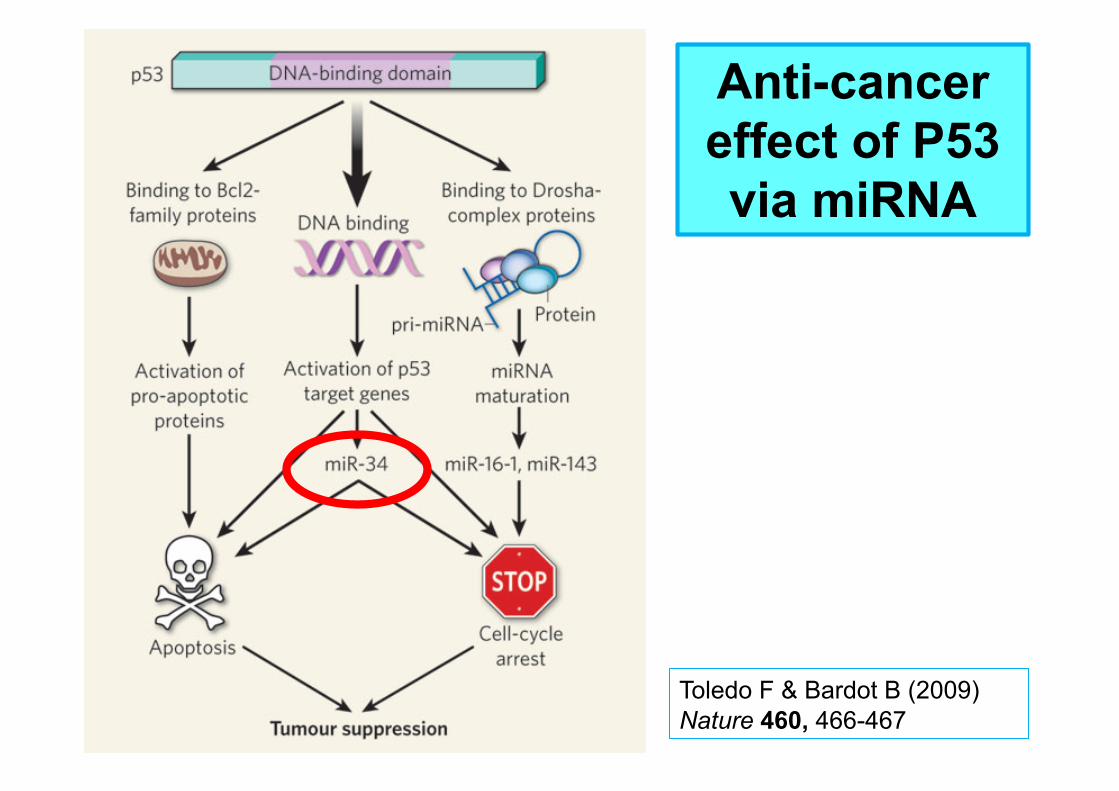
Toledo F & Bardot B (2009) Nature 460, 466-467
Anti-cancer effect of P53 via miRNA
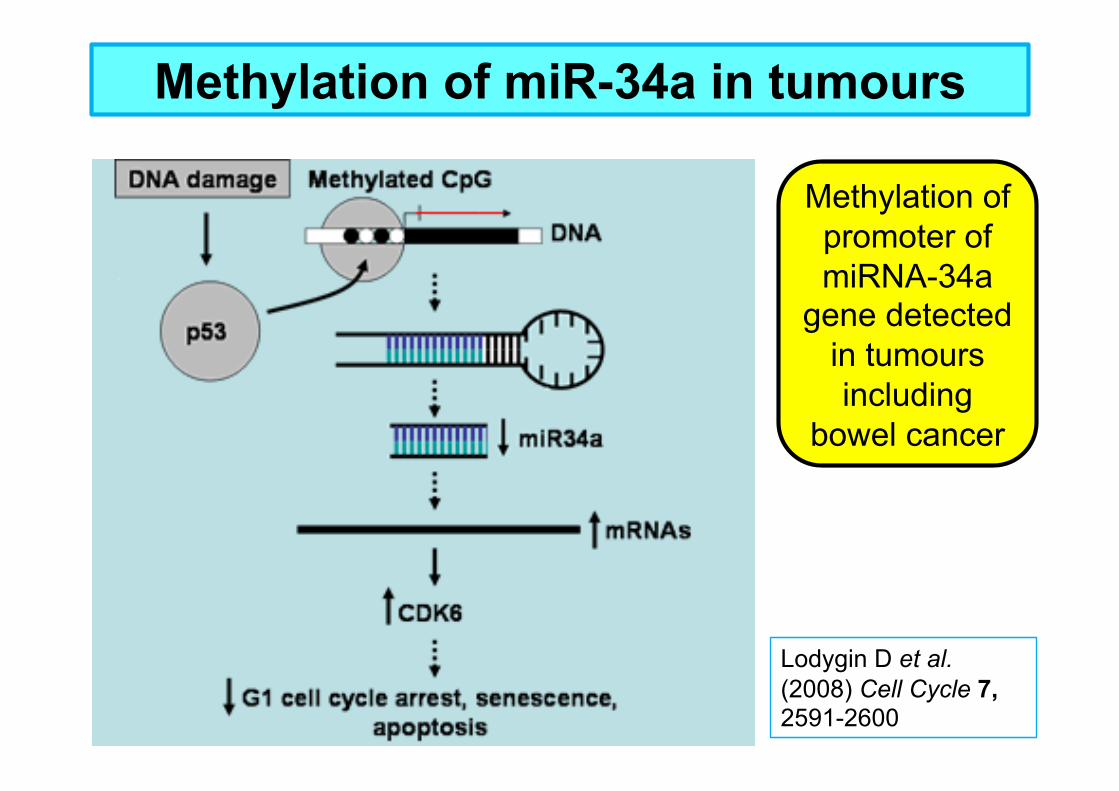
Methylation of miR-34a in tumours
Lodygin D et al. (2008) Cell Cycle 7, 2591-2600
Methylation of promoter of miRNA-34a
gene detected in tumours including
bowel cancer

Diet, miRNA and bowel cancer risk
?
Understanding aetiological mechanisms Development of novel diet-responsive biomarkers of bowel cancer risk

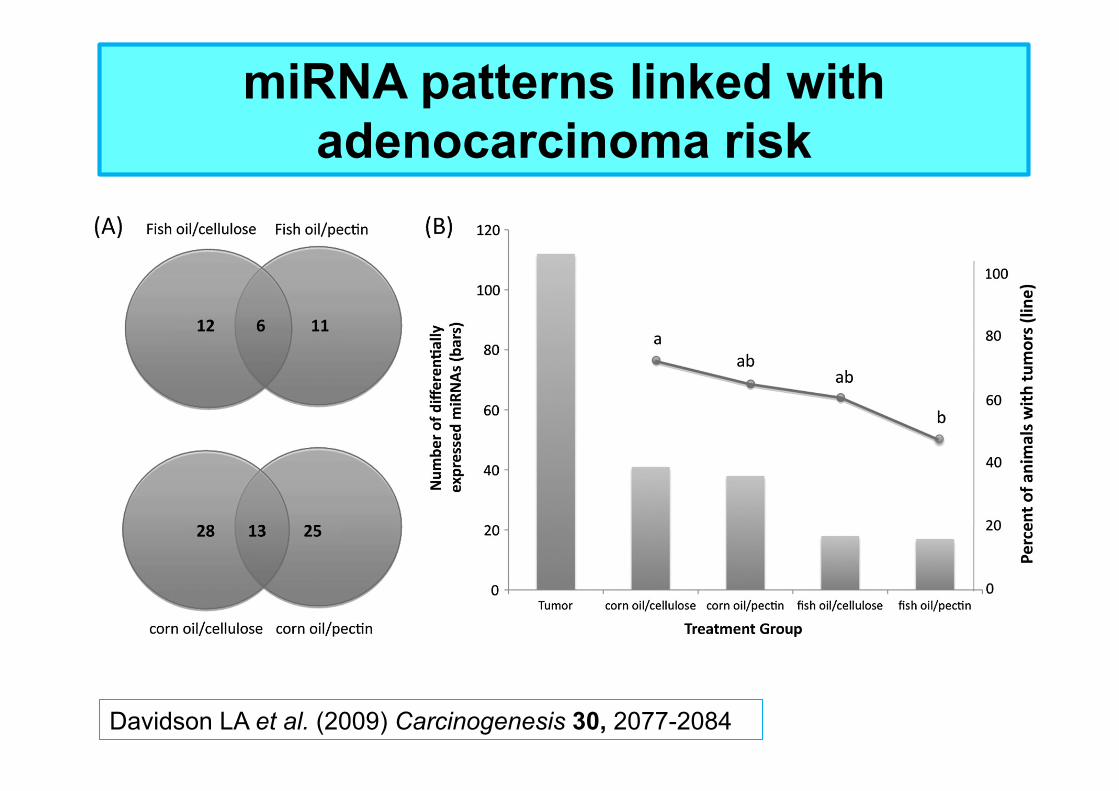
Davidson LA et al. (2009) Carcinogenesis 30, 2077-2084
Effect of dietary factors on miRNA signatures in rat colon
Cellulose Pectin
Corn oil Fish oil
2*2*2 factorial designed study in Sprague-Dawley rats
2 types of dietary fibre
2 types of fat
+ and – AOM treatment
Assayed 368 mature miRNAs in colonic mucosa

Davidson LA et al. (2009) Carcinogenesis 30, 2077-2084
Diet alters miRNA in rat colon
Cellulose
Pectin
Corn oil
Fish oil
Davidson LA et al. (2009) Carcinogenesis 30, 2077-2084
miRNA patterns linked with adenocarcinoma risk
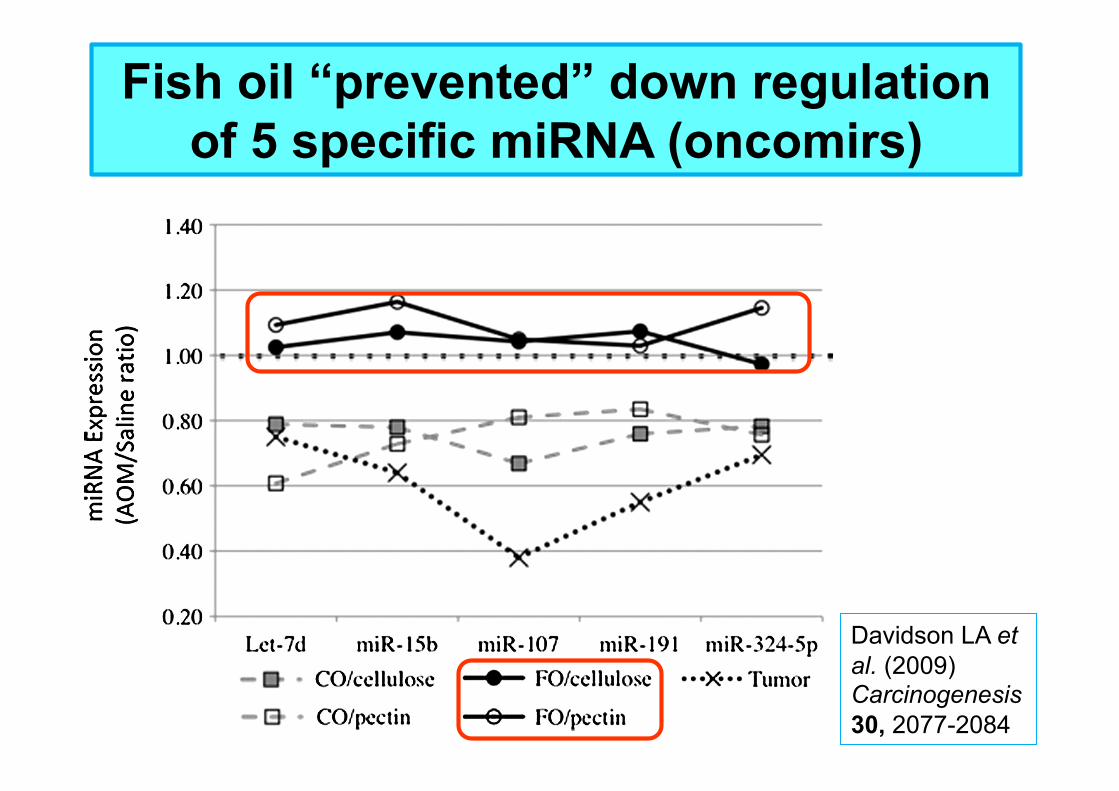
Davidson LA et al. (2009) Carcinogenesis 30, 2077-2084
Fish oil “prevented” down regulation of 5 specific miRNA (oncomirs)

Summary
Diet is a major modulator of bowel cancer risk
Epigenetics mechanisms link dietary exposure with development of bowel cancer
DNA methylation shows promise as route to novel (diet-related) biomarkers of bowel cancer risk
DNA methylation measurements can be made in stool
Altered miRNA patterns occur in cancer and may be diet responsive

Research prioriCes
• Which epigeneEc changes in macroscopically normal mucosa are causal for ↑ bowel cancer risk?
• What are major exposures causing ↑ epigeneEc heterogeneity with age?
• What dietary (and other lifestyle) factors prevent, or reverse, these early epigeneEc changes?
Acknowledgements
Nigel Belshaw
Giles Elliott
Mike Bradburn
Dharmendra Garg
Wendy Bal
Liz Williams
Ian Johnson





























