Embed Size (px)
Citation preview

Eab
JEa
b
c
a
ARRAA
KEOAOT
1
skrieltaiep1
M1
0h
Immunology Letters 157 (2014) 51– 59
Contents lists available at ScienceDirect
Immunology Letters
jo u r n al homep age: www.elsev ier .com/ locate / immlet
pigallocatechin-3-gallate ameliorates both obesity andutoinflammatory arthritis aggravated by obesity by altering thealance among CD4+ T-cell subsets
ae-Kyeong Byuna,1, Bo-Young Yoonc,1, Joo-Yeon Jhuna, Hye-Joa Oha,un-kyoung Kima, Jun-Ki Mina, Mi-La Choa,b,∗
The Rheumatism Research Center, Catholic Research Institute of Medical Science, The Catholic University of Korea Seoul, South KoreaConversant Research Consortium in Immunologic Disease, College of Medicine, The Catholic University of Korea Seoul, South KoreaDepartment of Internal Medicine, Inje University Ilsan Paik Hospital, Goyang 411-706, Republic of Korea
r t i c l e i n f o
rticle history:eceived 3 August 2013eceived in revised form 1 November 2013ccepted 4 November 2013vailable online 12 November 2013
eywords:GCG
a b s t r a c t
Epigallocatechin-3-gallate (EGCG) is the most biologically active catechin in green tea. EGCG has beenshown to have therapeutic effects in autoinflammatory diseases and obesity. Obesity is currentlyregarded—partly—as an inflammatory condition because of the inflammatory cytokines and higher Th1cell differentiation detected in obese animal models and human cohort studies. In this work, the effectsof EGCG on diet-induced obesity (DIO) mice and obese collagen-induced arthritis (CIA) mice were inves-tigated. EGCG reduced the body weight and fat infiltration in liver tissue while improving serum lipidprofiles in DIO mice. EGCG also induced a higher Treg/Th17 cell ratio in CD4+ T-cell differentiation by
beseutoinflammatorybese CIAh17/Treg balance
decreasing the ratio of STAT3/STAT5 expression in DIO mice. EGCG was also effective in obese CIA mice.Reducing Th17 cells and increasing regulatory T (Treg) cells by affecting the STAT protein ratio wereimportant effects of EGCG that might result in improved arthritic scores and levels of several inflamma-tory indicators. Thus, EGCG has an anti-inflammatory effect by suppressing STAT3 proteins and Th17-celldifferentiation. EGCG thus shows promise for treating autoimmune conditions related to STAT3 or Th17cells, such as metabolic syndrome, inflammatory arthritis, and some neoplastic diseases.
. Introduction
Green tea (Camellia sinensis) is one of the most commonly con-umed beverages globally and is a rich source of polyphenols,nown as catechins, which are thought to be the major componentsesponsible for the biological effects of tea [1]. These catechinsnclude epicatechin, epigallocatechin, epicatechin-3-gallate, andpigallocatechin-3-gallate (EGCG), of which EGCG is the most bio-ogically active and most abundant (accounting for 50–80% of totalea catechins) [2]. EGCG has been shown to improve symptomsnd reduce pathological conditions associated with autoimmunenflammatory disease [3–6]. In an experimental autoimmune
ncephalomyelitis (EAE) model, EGCG reduced the production ofro-inflammatory cytokines, such as interferon-�, IL-17, IL-6, IL-�, and tumor necrosis factor (TNF)-�, as well as altering the∗ Corresponding author at: Rheumatism Research Center, Catholic Institutes ofedical Science, The Catholic University of Korea, 505 Banpo-dong, Seocho-gu, Seoul
37-040, South Korea. Tel.: +82 2 2258 7467; fax: +82 2 599 4287.E-mail address: [email protected] (M.-L. Cho).
1 These authors contributed equally to this work.
165-2478/$ – see front matter © 2013 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.imlet.2013.11.006
© 2013 Elsevier B.V. All rights reserved.
balance among CD4+ T-cell subsets [7]. Additionally, EGCG has beenshown to affect angiogenesis [8] and apoptosis [9], and to act as anantioxidant [10]. Some investigators have also reported that EGCGis effective against metabolic syndrome and obesity [11–13].
Obesity is known to be a low-grade inflammatory conditionthat can aggravate or induce further inflammation. Epidemiologicalstudies have reported relationships between the levels of pro-inflammatory biomarkers, such as C-reactive protein, IL-6, andTNF-�, and abnormal glucose homeostasis, obesity, and atheroscle-rosis [14]. Adipocytokines, including adiponectin, leptin, resistin,and visfatin, are linking mediators between adipose tissue andinflammation [15–17]. Leptin, an important adipocytokine, hasbeen shown to induce inflammation by increasing levels of TNF, IL-6, IL-12 [18], and various reactive oxygen species (ROS) [19]. Studieshave also reported an effect of leptin on Th1 differentiation of CD4+
T cells [20]. These results demonstrate that other factors play rolesin obesity and in insulin resistance, dyslipidemia, and atherosclero-sis. Thus, we hypothesized that materials with anti-inflammatory
properties could attenuate obesity or treat metabolic issuesassociated with obesity. That is, anti-inflammatory substances thatinfluence the immune system could ameliorate the inflammatorystate in obesity.
5 ology
atmtTemnli
gtoevta(o(GstToalmaEt
2
2
ocWaaUt
2
ta(4r(Cmig
2
s
2 J.-K. Byun et al. / Immun
One study showed a relationship between metabolic disordersnd inflammatory diseases, where obesity aggravated inflamma-ion in joint inflammation, using a collagen-induced arthritis (CIA)
odel [21]. This showed an additive effect of obesity on inflamma-ion in a CIA model through bovine type II collagen (CII)-specific
cell (Th17 cells) differentiation. Using this obese CIA model, theffects of anti-inflammatory materials on obesity and the inflam-atory arthritis aggravated by obesity can be evaluated. Using
on-pharmaceutical methodologies, such as specific foods, the bio-ogical mechanisms in obese and obese CIA models are focusing onnflammatory cytokines and T cell differentiation.
In recent decades, there has been a dramatic increase androwing interest in the use of alternative treatments and herbalherapies in arthritis patients [22,23]. Trustworthy documentationf traditional medical uses, together with extensive modern sci-ntific/pharmacological experimentation, however, is necessary toalidate or refute the supposed medicinal value of many traditionalherapies [24]. The effects and mechanisms of action of severalnti-inflammatory materials have been evaluated in both obesityDIO) mice models and obese CIA mice models. First, our previ-us study demonstrated that grape seed proanthocyanidin extractGSPE) was effective in DIO and obese CIA mouse models (in press).SPE improved obesity in DIO mice and the arthritis score andeveral inflammatory indicators in obese CIA mice by reducinghe numbers of Th17 cells and increasing numbers of regulatory
(Treg) cells via regulating STAT protein. Following this, EGCG wasf interest due to its history of beneficial effects in both obesity andutoimmune inflammatory arthritis. We hypothesized that EGCG,ike GSPE, could have a therapeutic effect on obesity and obese CIA
ice through immunological mechanisms. To assess this, DIO micend our new murine model, obese CIA mice, were treated withGCG and T cell differentiation, levels of signaling molecules, andherapeutic effects were evaluated.
. Materials and methods
.1. Animals
C57BL/6 mice (males, n = 6; SLC, Inc., Shozuoka, Japan), 4 weeksld, were housed in polycarbonate cages and fed standard mousehow (Ralston Purina, St. Louis, MO, USA) and water ad libitum.
e also induced obesity with a 60 Kcal high-fat diet with foodnd water available ad libitum. All experimental procedures werepproved by the Animal Research Ethics Committee of the Catholicniversity of Korea (permit number: CUMC:20), and conformed to
he US National Institutes of Health guidelines.
.2. Induction of obese CIA
Chicken type II collagen (CII) immunization was performed forhe first time when the mouse weighed 15 g. CIA was induceds described previously [25]. Briefly, complete Freund’s adjuvantCFA; Chondrex, Redmond, WA, USA) was prepared by grinding
mg of heat-killed Mycobacterium tuberculosis (H37Ra; Difco Labo-atories, Detroit, MI, USA) in 2 mL of incomplete Freund’s adjuvantIFA; Chondrex). An emulsion was formed by dissolving 2 mg/mLII (Chondrex) overnight at 4 ◦C in 0.5 M acetic acid, followed byixing this with an equal volume of CFA. The mice were injected
ntradermally in the tail and SC. Then, 2 weeks later, the mice wereiven a booster injection of 100 �g of CII emulsified in IFA (1:1, v/v).
.3. EGCG treatment
EGCG was obtained from Sigma Chemical Co. EGCG, dissolved inaline, was given intraperitoneally at 20 mg/kg after the secondary
Letters 157 (2014) 51– 59
immunization. The mice were injected with 20 mg/kg EGCG threetimes per week.
2.4. Arthritis histopathology
Mouse joint tissues were fixed using 10% formalin, decalci-fied in EDTA, and embedded in paraffin wax. The sections weredewaxed using xylene and rehydrated through a graded series ofalcohol solutions. The sections were then stained with hematoxylinand eosin (H&E), safranin O, and toluidine blue to detect proteo-glycans. H&E-stained sections were scored for inflammation andbone erosion. Inflammation was scored according to the follow-ing scale: 0 = no inflammation, 1 = slight thickening of the lininglayer or some infiltrating cells in the underlying layer, 2 = slightthickening of the lining layer plus some infiltrating cells in theunderlying layer, 3 = thickening of the lining layer, an influx ofcells in the underlying layer and the presence of cells in the syno-vial space, and 4 = synovium infiltrated with many inflammatorycells. Cartilage damage was determined using safranin O stainingand toluidine blue and the extent of cartilage damage was scoredaccording to the following scale: 0 = no destruction, 1 = minimalerosion limited to single spots, 2 = slight-to-moderate erosion in alimited area, 3 = more extensive erosion, and 4 = general destruction[26]. Immunohistochemistry was performed using the VectastainABC kit (Vector Laboratories, Burlingame, CA, USA). Tissues wereincubated with anti-IL-17, anti-IL-6, anti-IL-1b, anti-TNF-�, andanti-nitrotyrosine antibodies (Santa Cruz Biotechnology Inc., SantaCruz, CA, USA) and then with a biotinylated secondary linking Ab,and streptavidin-peroxidase complex. The final color product wasdeveloped using the DAB chromogen (DAKO, Carpinteria, CA, USA).Images were captured using a DP7 1 digital camera (Olympus, Cen-ter Valley, PA, USA) attached to an Olympus BX4 1 microscope.
2.5. Biochemical analyses
Blood samples were collected 11 weeks after immunizationfrom both treated and control mice and stored at −70 ◦C until use.Serum total cholesterol levels were measured using commercialkits (Wako Co., Osaka, Japan), and LDL-cholesterol and triglyce-rides were measured using kits from Asan Pharmaceutical Co.(Hwangseong-si Gyeonggi-do, Korea).
2.6. T-cell proliferation
Mouse spleens were collected for cell preparation and washedwith PBS. The spleens were minced and red blood cells were lysedwith 0.83% ammonium chloride. The cells were filtered through acell strainer and then centrifuged (1300 rpm, 5 min, 4 ◦C). The cellpellets were resuspended in RPMI 1640 medium. Splenocytes ofobese and EGCG-treated mice were cultured at 2 × 105 cells/wellin 96-well plates at 37 ◦C for 72 h in the absence or presence ofplate-bound CD3 0.5 �g/mL. Cells were cultured for 72 h, and 18 hbefore the end of the culture, 1 �Ci [3H]-thymidine (AmershamPharmacia Biotech, Little Chalfont, UK) was added to each well. Theincorporation of [3H]-thymidine was determined using a Betaplatescintillation counter (Perkin–Elmer, Wellesley, MA, USA).
2.7. Flow cytometry
Cells were stained with PerCP-conjugated anti-CD4 Ab (BDPharmingen), then stained with APC-conjugated anti-CD25 and PE-
conjugated anti-Foxp3 (all from eBiosciences, San Diego, CA, USA),followed by fixation and permeabilization using the mouse Foxp3Buffer Set (BD Biosciences) according to the manufacturer’s instruc-tions. All samples were run on a FACSCalibur (BD Pharmingen), and
ology L
dO
2
sP(Afm×
2
oTptfffug
2
tuIkpoasT4
2
tuJLdCACaCRGTrcA(TE
J.-K. Byun et al. / Immun
ata were analyzed using the FlowJo software (Tree Star, Ashland,R, USA).
.8. Confocal microscopy
For confocal staining, 7-�m tissue sections of spleens weretained using FITC-conjugated anti-CD4, PE-conjugated anti-IL-17,E-conjugated anti-p-STAT3 (Ser727), PE-conjugated anti-p-STAT3Tyr705), PE-conjugated anti-p-STAT5, PE conjugated anti-CD4,PC-conjugated anti-CD25, and FITC-conjugated anti-Foxp3 (all
rom eBiosciences). Stained sections were analyzed using a Zeissicroscope (LSM 510 Meta; Carl Zeiss, Oberkochen, Germany) at400 magnification.
.9. Clinical assessment of arthritis
The severity of arthritis was assessed by two independentbservers. The development of CIA was assessed twice per week.he severity of arthritis was assessed on a scale of 0–4, as describedreviously [27]: 0 = normal (no edema), 1 = slight edema and ery-hema limited to the foot or ankle, 2 = slight edema and erythemarom the ankle to the tarsal bone, 3 = moderate edema and erythemarom the ankle to the tarsal bone, and 4 = edema and erythemarom the ankle to the entire leg. The mean arthritis index wassed to compare the data among the control and experimentalroups.
.10. Anti-Cll antibody measurement
Blood samples were taken from the orbital sinuses of EGCG-reated and obese CIA mice and the sera were stored at −20 ◦Cntil use. Anti-CII IgG2a antibodies were measured using the mouse
gG2a enzyme-linked immunosorbent assay (ELISA) quantitationit (Bethyl Lab. Co., Montgomery, TX, USA) [27]. Briefly, microtiterlates were coated with type II collagen (2 �g/mL in PBS) at 4 ◦Cvernight, followed by a blocking step for 30 min at room temper-ture. Serum samples were then diluted 1:100,000 in Tris-bufferedaline (pH 8.0) containing 1% bovine serum albumin and 0.5%ween-20. The optical density (OD) of each well was measured at50 nm.
.11. Analysis of gene expression by real-time quantitative PCR
Total RNA was extracted using TRIzol (Molecular Research Cen-er, Cincinnati, OH, USA). Total RNA (2 �g) was reverse transcribedsing the Superscript Reverse Transcription system (Takara, Shiga,
apan). Quantitative real-time PCR (qRT-PCR) was performed withightCycler FastStart DNAmaster SYBR green I (Takara) fluorescentye using an ABI PCR machine. Primers for IL-17 (forward: 5′-CCTAA AGC TCA GCG TGT CC-3′, reverse: 5′-GAG CTC ACT TTT GCG CCAG-3′), forkhead box P3 (Foxp3, forward: 5′-GGC CCT TCT CCA GGAAG A-3′, reverse: 5′-GCT GAT CAT GGC TGG GTT GT-3′), retinoiccid-related orphan receptor (ROR�t), forward: 5′-TGT CCT GGGTA CCC TAC TG-3′, reverse: 5′-GTG CAG GAG TAG GCC ACA TT-3′,unt-related transcription factor 1 (RUNX1, forward: 5′-TAC CTGGA TCC ATC ACC TC-3′, reverse: 5′-GAC GGC AGA GTA GGG AACG-3′), IL-21 (forward: 5′-AAG ATT CCT GAG GAT CCG AGA AG-3′,everse: 5′-GCA TTC GTG AGC GTC TAT AGT GTC-3′), suppressor ofytokine signaling-3 (SOCS3, forward: 5′-CTC AAG ACC TTC AGC TCC
A-3′, reverse: 5′-TTC TCA TAG GAG TCC AGG TA-3′) and �-actinforward: 5′-GAA ATC GTG CGT GAC ATC AAA G-3′, reverse: 5′-GT AGT TTC ATG GAT GCC ACA G-3′) were designed using Primerxpress (Applied Biosystems, Foster City, CA, USA).
etters 157 (2014) 51– 59 53
2.12. Statistical analysis
Results are expressed as means ± SD (or means ± SEM). Stu-dent’s t-tests were performed to assess the significance ofdifferences between the means of the various variables, and pvalues less than 0.05 (two-tailed) were considered to indicatestatistical significance. Groups of data were compared using theMann–Whitney U-test. Differences were considered statisticallysignificant at p < 0.05.
3. Results
3.1. Effect of EGCG in obese mice
C57BL/6 (n = 6) mice were fed a high-fat diet from 4 weeks ofage. EGCG treatment was first performed when the mouse weighed15 g. Mice were administered EGCG at 20 mg/kg during the exper-imental period. The EGCG-treated mice group showed significantweight reduction (Fig. 1A). Adipose infiltration in liver tissue byoil red O staining was investigated in obese mice with or with-out EGCG treatment. EGCG-treated mice showed fewer oil redO-positive cells than obese mice (Fig. 1B). We also performedimmunohistochemical staining for nitrotyrosine, a marker of oxi-dant stress. Nitrotyrosine staining in the liver and spleen wasreduced in the EGCG-treated group (Fig. 1B). Plasma lipid levelsin mice fed the high-fat diet with and without EGCG treatmentwere measured. Total cholesterol, LDL-cholesterol, and triglyceridelevels were decreased significantly by EGCG treatment (Fig. 1C).
3.2. EGCG suppressed the proliferative response
Splenocytes were cultured with anti-CD3 in the presence orabsence of EGCG. The cells obtained from the spleens of obesemice treated with EGCG showed decreased T-cell proliferation inresponse to anti-CD3 stimulation compared with cells obtainedfrom obese mice without EGCG treatment (Fig. 1D).
3.3. EGCG increased the frequency of Foxp3+ Treg cells anddecreased the frequency of Th17 cells in obese mice
We then performed intracellular Foxp3 staining on splenocytesfrom obese and EGCG-treated mice. Mean fluorescence intensities(MFI) were measured using flow cytometry. This indicated thatEGCG increased the frequency of Foxp3+ Treg cells (Fig. 2A). Popula-tions of Foxp3+ Treg cells and Th17 cells were observed in the spleentissue of the EGCG-treated obese mice. An increased population ofTreg cells was detected in the spleens of EGCG-treated mice. Thegraphs present the numbers of positive cells (Fig. 2B). A decrease inthe population of Th17-related transcription factor phospho-STAT3Y705 and S727 cells was detected in the spleens of EGCG-treatedmice. In contrast, the population of Treg-related transcription fac-tor phospho-STAT5+ cells was increased in EGCG-treated mice. Thegraphs present the numbers of positive cells (Fig. 2C).
3.4. Effect of EGCG in obese CIA mice
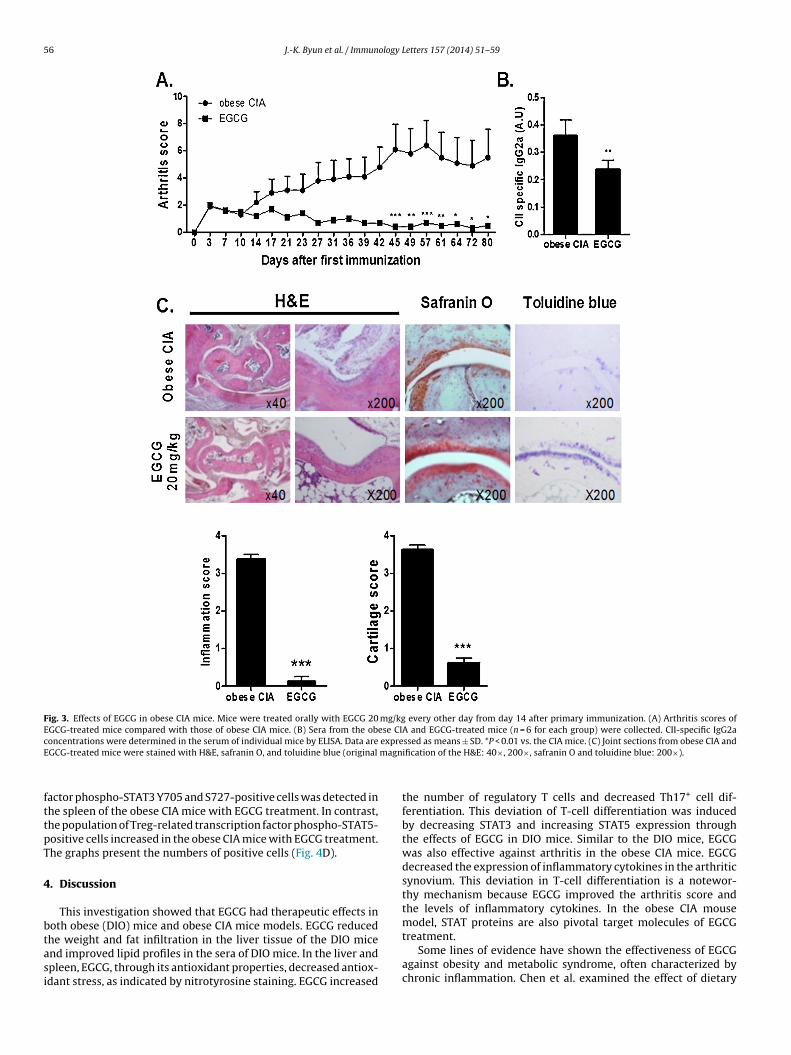
An obese inflammatory arthritis model was created accordingto a previous report [21]. C57BL/6 (wild-type, n = 6) mice fed a high-fat diet (60 kcal) were immunized with bovine type II collagen (CII),and arthritis developed. There were significant reductions in themean arthritis scores in the EGCG-treated mice compared with theobese CIA mice (Fig. 3A). The level of CII-specific IgG2a in the serum,
reflecting the Th1 response, decreased significantly in the obeseCIA mice treated with EGCG versus untreated obese CIA mice. Thisindicates the effects of EGCG treatment on serum levels of type IIcollagen-specific IgG subtypes (Fig. 3B).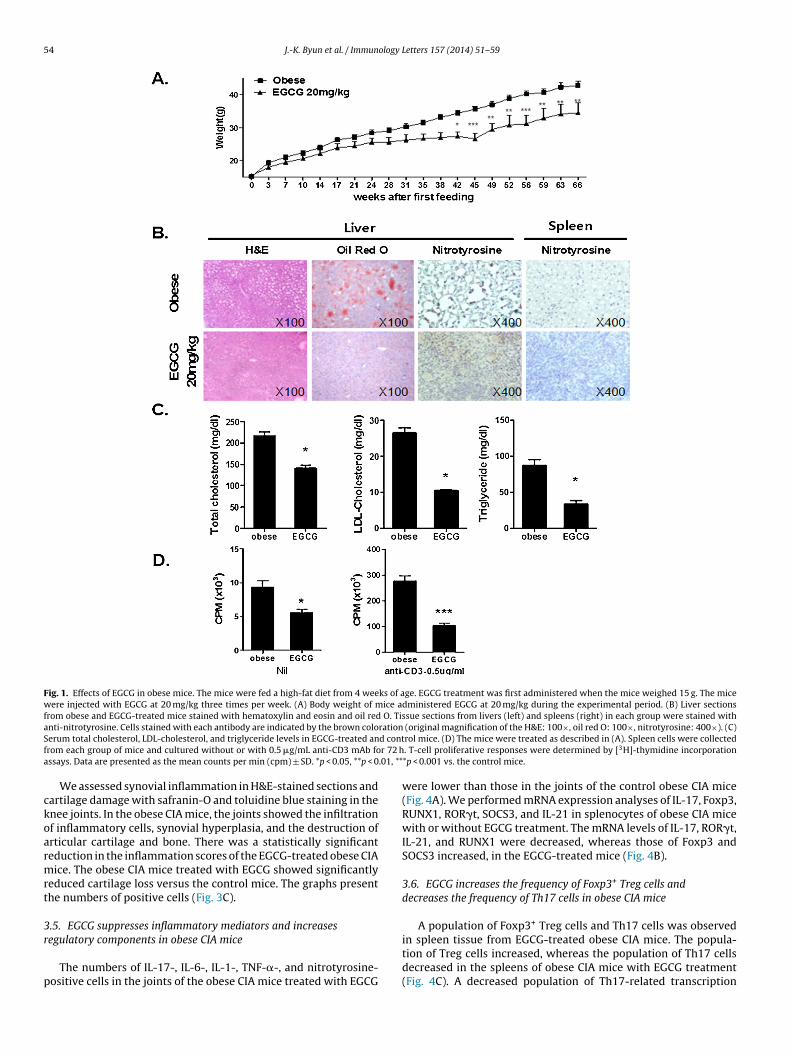
54 J.-K. Byun et al. / Immunology Letters 157 (2014) 51– 59
Fig. 1. Effects of EGCG in obese mice. The mice were fed a high-fat diet from 4 weeks of age. EGCG treatment was first administered when the mice weighed 15 g. The micewere injected with EGCG at 20 mg/kg three times per week. (A) Body weight of mice administered EGCG at 20 mg/kg during the experimental period. (B) Liver sectionsfrom obese and EGCG-treated mice stained with hematoxylin and eosin and oil red O. Tissue sections from livers (left) and spleens (right) in each group were stained withanti-nitrotyrosine. Cells stained with each antibody are indicated by the brown coloration (original magnification of the H&E: 100×, oil red O: 100×, nitrotyrosine: 400×). (C)S d contf r 72 ha .01, **
ckoarmrt
3r
p
erum total cholesterol, LDL-cholesterol, and triglyceride levels in EGCG-treated anrom each group of mice and cultured without or with 0.5 �g/mL anti-CD3 mAb fossays. Data are presented as the mean counts per min (cpm) ± SD. *p < 0.05, **p < 0
We assessed synovial inflammation in H&E-stained sections andartilage damage with safranin-O and toluidine blue staining in thenee joints. In the obese CIA mice, the joints showed the infiltrationf inflammatory cells, synovial hyperplasia, and the destruction ofrticular cartilage and bone. There was a statistically significanteduction in the inflammation scores of the EGCG-treated obese CIAice. The obese CIA mice treated with EGCG showed significantly
educed cartilage loss versus the control mice. The graphs presenthe numbers of positive cells (Fig. 3C).
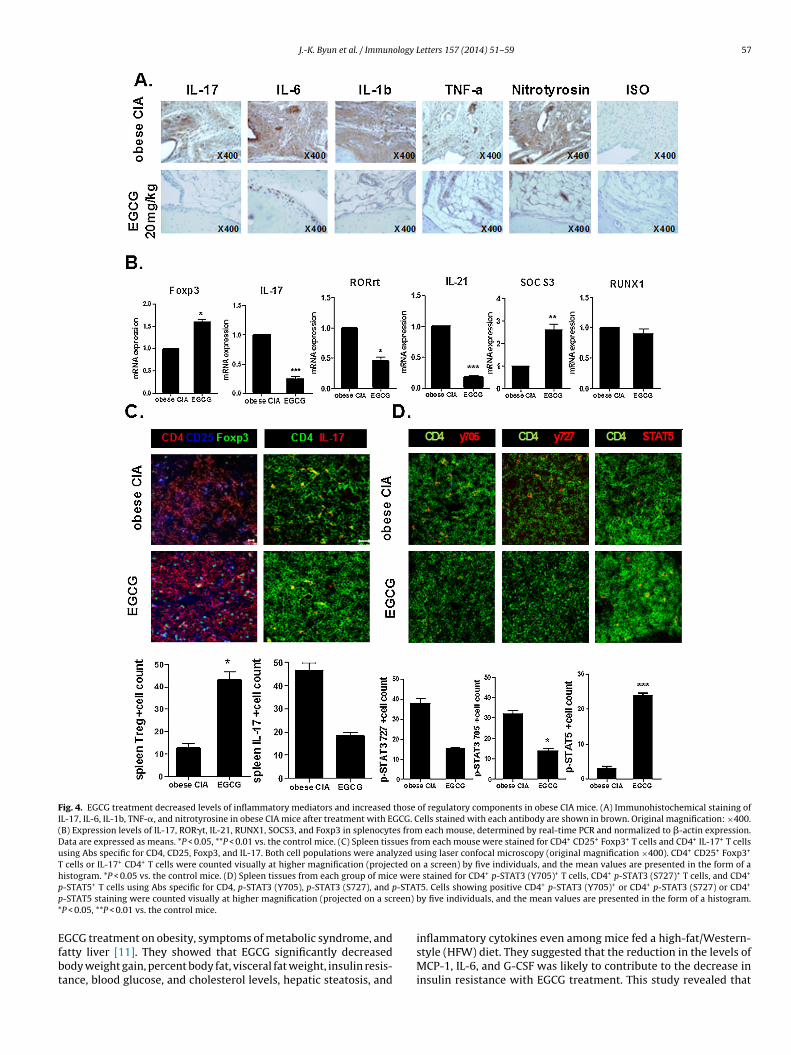
.5. EGCG suppresses inflammatory mediators and increases
egulatory components in obese CIA miceThe numbers of IL-17-, IL-6-, IL-1-, TNF-�-, and nitrotyrosine-ositive cells in the joints of the obese CIA mice treated with EGCG
rol mice. (D) The mice were treated as described in (A). Spleen cells were collected. T-cell proliferative responses were determined by [3H]-thymidine incorporation*p < 0.001 vs. the control mice.
were lower than those in the joints of the control obese CIA mice(Fig. 4A). We performed mRNA expression analyses of IL-17, Foxp3,RUNX1, ROR�t, SOCS3, and IL-21 in splenocytes of obese CIA micewith or without EGCG treatment. The mRNA levels of IL-17, ROR�t,IL-21, and RUNX1 were decreased, whereas those of Foxp3 andSOCS3 increased, in the EGCG-treated mice (Fig. 4B).
3.6. EGCG increases the frequency of Foxp3+ Treg cells anddecreases the frequency of Th17 cells in obese CIA mice
A population of Foxp3+ Treg cells and Th17 cells was observed
in spleen tissue from EGCG-treated obese CIA mice. The popula-tion of Treg cells increased, whereas the population of Th17 cellsdecreased in the spleens of obese CIA mice with EGCG treatment(Fig. 4C). A decreased population of Th17-related transcription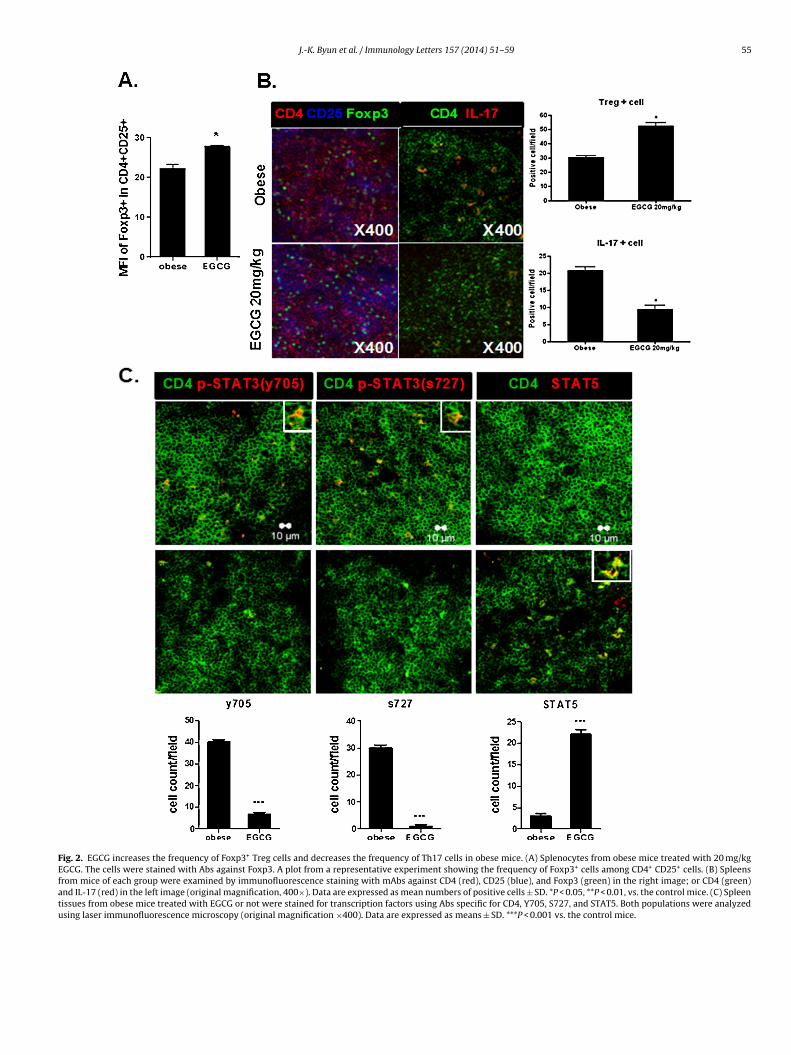
J.-K. Byun et al. / Immunology Letters 157 (2014) 51– 59 55
Fig. 2. EGCG increases the frequency of Foxp3+ Treg cells and decreases the frequency of Th17 cells in obese mice. (A) Splenocytes from obese mice treated with 20 mg/kgEGCG. The cells were stained with Abs against Foxp3. A plot from a representative experiment showing the frequency of Foxp3+ cells among CD4+ CD25+ cells. (B) Spleensfrom mice of each group were examined by immunofluorescence staining with mAbs against CD4 (red), CD25 (blue), and Foxp3 (green) in the right image; or CD4 (green)and IL-17 (red) in the left image (original magnification, 400×). Data are expressed as mean numbers of positive cells ± SD. *P < 0.05, **P < 0.01, vs. the control mice. (C) Spleentissues from obese mice treated with EGCG or not were stained for transcription factors using Abs specific for CD4, Y705, S727, and STAT5. Both populations were analyzedusing laser immunofluorescence microscopy (original magnification ×400). Data are expressed as means ± SD. ***P < 0.001 vs. the control mice.
56 J.-K. Byun et al. / Immunology Letters 157 (2014) 51– 59
Fig. 3. Effects of EGCG in obese CIA mice. Mice were treated orally with EGCG 20 mg/kg every other day from day 14 after primary immunization. (A) Arthritis scores ofEGCG-treated mice compared with those of obese CIA mice. (B) Sera from the obese CIA and EGCG-treated mice (n = 6 for each group) were collected. CII-specific IgG2ac expreE magn
fttpT
4
btasi
oncentrations were determined in the serum of individual mice by ELISA. Data areGCG-treated mice were stained with H&E, safranin O, and toluidine blue (original
actor phospho-STAT3 Y705 and S727-positive cells was detected inhe spleen of the obese CIA mice with EGCG treatment. In contrast,he population of Treg-related transcription factor phospho-STAT5-ositive cells increased in the obese CIA mice with EGCG treatment.he graphs present the numbers of positive cells (Fig. 4D).
. Discussion
This investigation showed that EGCG had therapeutic effects inoth obese (DIO) mice and obese CIA mice models. EGCG reduced
he weight and fat infiltration in the liver tissue of the DIO micend improved lipid profiles in the sera of DIO mice. In the liver andpleen, EGCG, through its antioxidant properties, decreased antiox-dant stress, as indicated by nitrotyrosine staining. EGCG increasedssed as means ± SD. *P < 0.01 vs. the CIA mice. (C) Joint sections from obese CIA andification of the H&E: 40×, 200×, safranin O and toluidine blue: 200×).
the number of regulatory T cells and decreased Th17+ cell dif-ferentiation. This deviation of T-cell differentiation was inducedby decreasing STAT3 and increasing STAT5 expression throughthe effects of EGCG in DIO mice. Similar to the DIO mice, EGCGwas also effective against arthritis in the obese CIA mice. EGCGdecreased the expression of inflammatory cytokines in the arthriticsynovium. This deviation in T-cell differentiation is a notewor-thy mechanism because EGCG improved the arthritis score andthe levels of inflammatory cytokines. In the obese CIA mousemodel, STAT proteins are also pivotal target molecules of EGCG
treatment.Some lines of evidence have shown the effectiveness of EGCGagainst obesity and metabolic syndrome, often characterized bychronic inflammation. Chen et al. examined the effect of dietary
J.-K. Byun et al. / Immunology Letters 157 (2014) 51– 59 57
Fig. 4. EGCG treatment decreased levels of inflammatory mediators and increased those of regulatory components in obese CIA mice. (A) Immunohistochemical staining ofIL-17, IL-6, IL-1b, TNF-�, and nitrotyrosine in obese CIA mice after treatment with EGCG. Cells stained with each antibody are shown in brown. Original magnification: ×400.(B) Expression levels of IL-17, ROR�t, IL-21, RUNX1, SOCS3, and Foxp3 in splenocytes from each mouse, determined by real-time PCR and normalized to �-actin expression.Data are expressed as means. *P < 0.05, **P < 0.01 vs. the control mice. (C) Spleen tissues from each mouse were stained for CD4+ CD25+ Foxp3+ T cells and CD4+ IL-17+ T cellsusing Abs specific for CD4, CD25, Foxp3, and IL-17. Both cell populations were analyzed using laser confocal microscopy (original magnification ×400). CD4+ CD25+ Foxp3+
T cells or IL-17+ CD4+ T cells were counted visually at higher magnification (projected on a screen) by five individuals, and the mean values are presented in the form of ahistogram. *P < 0.05 vs. the control mice. (D) Spleen tissues from each group of mice were stained for CD4+ p-STAT3 (Y705)+ T cells, CD4+ p-STAT3 (S727)+ T cells, and CD4+
p -STATp reen)
*
Efbt
-STAT5+ T cells using Abs specific for CD4, p-STAT3 (Y705), p-STAT3 (S727), and p-STAT5 staining were counted visually at higher magnification (projected on a scP < 0.05, **P < 0.01 vs. the control mice.
GCG treatment on obesity, symptoms of metabolic syndrome, andatty liver [11]. They showed that EGCG significantly decreasedody weight gain, percent body fat, visceral fat weight, insulin resis-ance, blood glucose, and cholesterol levels, hepatic steatosis, and
5. Cells showing positive CD4+ p-STAT3 (Y705)+ or CD4+ p-STAT3 (S727) or CD4+
by five individuals, and the mean values are presented in the form of a histogram.
inflammatory cytokines even among mice fed a high-fat/Western-style (HFW) diet. They suggested that the reduction in the levels ofMCP-1, IL-6, and G-CSF was likely to contribute to the decrease ininsulin resistance with EGCG treatment. This study revealed that

5 ology
ot
daetpwEr(rtimatsisGittg
enetctpctTawTcresow
EmEi
amISCptEBd
grt
8 J.-K. Byun et al. / Immun
besity involves inflammation and can be treated with EGCG, dueo its anti-inflammatory properties.
Some clinical trials have reported that EGCG significantlyecreased weight and body mass index (BMI) [12,28,29]. Recently,
randomized controlled cross-over trial with EGCG examined theffects of EGCG in overweight and obese males [13]. Althoughhe investigators suspected a positive effect on obesity and bloodressure, the results showed that EGCG induced only a bodyeight change. Several anti-obesity mechanisms influenced by
GCG have been proposed, such as inhibition of extracellular signal-elated kinases (ERKs), activating AMP-activated protein kinaseAMPK), modulation of adipocyte marker proteins, and down-egulation of lipogenic enzymes [30]. Here, EGCG exhibited aherapeutic effect against obesity by reducing body weight andmproving hepatic steatosis and serum cholesterol in the DIO
ouse model. However, we were interested in anti-inflammatorynd immune regulatory effects of EGCG in obesity because ofhe link between inflammation and obesity. Previously, GSPE washown to have anti-inflammatory or immune regulatory effectsn the DIO mouse model (in press). We repeated essentially theame investigation with EGCG in the DIO mouse model. Similar toSPE, EGCG not only reduced the body weight of DIO mice, but also
ncreased Treg differentiation and decreased Th17 cell differentia-ion. EGCG affects the immune state in obesity and can amelioratehe deviation in T cell differentiation and the inflammatory stateenerally.
Winer et al. demonstrated that obesity promoted selectivexpansion of the Th17 T-cell sublineage, a subset with promi-ent pro-inflammatory roles [31]. T cells from DIO mice showedxpanded Th17-cell pools and produced progressively more IL-17han lean littermates, in an IL-6-dependent process. Dendritic cellsould be regulators of adipose tissue inflammation by regulatinghe switch toward Th17-cell responses [32]. Because Th17 cellslay an important role in obesity, decreased numbers of Th17 cellsould be an effective target for treating obesity. Moreover, inves-igators using an EAE model showed that EGCG inhibited Th1 andh17 differentiation by down-regulating STAT1 and T-bet for Th1,nd STAT3 and ROR�t for Th17 [33,34]. The expression of STAT3as decreased, and that of STAT5 increased, by GSPE (in press).
hus, use of therapeutic materials, such as EGCG and GSPE, couldhange the Th17/Treg balance because of their ability to down-egulate the expression of STAT3. In this study, EGCG decreased thexpression of STAT3 and increased the expression of STAT5 in theplenocytes of the DIO mice. Thus, a possible mechanism of actionf EGCG in treating obesity through regulating signaling moleculesas demonstrated.
The observed effectiveness of EGCG in DIO mice suggested thatGCG could be effective in treating obese CIA mice, via a similarechanism to the immunological correction after treatment with
GCG in obese mice; i.e., decreased numbers of Th17 cells andncreased numbers of regulatory T cells.
This study also showed using immunohistochemistry that EGCGmeliorated arthritis scores and joint inflammation. After treat-ent with EGCG, there was a decrease in the expression of ROR�t,
L-17, and IL-21, and an increase in the expression of Foxp3 andOCS3 in the synovium of obese CIA mice versus control obeseIA mice (not EGCG-treated). Because EGCG has anti-inflammatoryroperties, EGCG may improve obesity and inhibit the aggrava-ion of arthritis induced by obesity in obese CIA mice. Furthermore,GCG may directly improve joint inflammation in obese CIA mice.ased on this study, EGCG may act both indirectly in obesity andirectly in arthritis.
This study suggests that obesity shares a common signaling andene transcription pathway with inflammation. STAT3 signaling isequired for leptin regulation of energy [35]. Selective inhibitors ofhe JAK2-STAT3 signaling pathway and STAT3 activation, as well as
Letters 157 (2014) 51– 59
STAT3 siRNA and a dominant-negative STAT3, all suppressed adipo-genesis in vitro [35]. These studies indicated that STAT3 activationinduced adipogenesis. It appears that STAT5 activation converselyis involved in inhibiting adipogenesis. Studies in the liver indicatedthat JAK2 and STAT5 proteins act to prevent hepatic lipid accumu-lation in mice [36]. Moreover, the activation of STAT3 induced thesecretion of pro-inflammatory cytokines, such as TNF, IL-6, and IL-12. STAT3 signaling is a pivotal transcription process in Th17-celldifferentiation; thus, depletion of STAT3 can prevent Th17 cell dif-ferentiation. Some investigators have reported that the modulationof STAT3 in CD4+ T cells affected the differentiation of Th17 and Tregcells in patients with rheumatoid arthritis (RA) and STAT3 may bea new therapeutic target for the management of RA [37]. This workdemonstrated that EGCG reduced STAT3 expression in the spleensof obese CIA mice. Thus, EGCG can directly affect adipogenesis andregulate T cell differentiation by controlling the STAT protein ratio:lower STAT3 and higher STAT5.
In conclusion, EGCG from green tea is effective against bothobesity (DIO mice) and inflammatory arthritis aggravated by obe-sity (obese CIA mice). The mechanism of the therapeutic efficacyinvolves the amelioration of the deviated T-cell differentiation,which T-cell differentiation may be related to a STAT proteinimbalance. So, EGCG results in decreased STAT3 and increasedSTAT5 proteins and ameliorates the deviated T-cell differentiation.Moreover, EGCG may have value in treating immunological condi-tions associated with STAT protein imbalances, including severalmetabolic, inflammatory, and even neoplastic diseases.
Conflicts of interest statement
The authors have no conflict to disclose.
Acknowledgments
This research was supported by grants from the Ministry ofHealth, Welfare, and Family Affairs (Korea Healthcare TechnologyR&D project grants A092258) and Sanofi-Aventis PharmaceuticalsCorporation.
References
[1] Manning J, Roberts JC. Analysis of catechin content of commercial green teaproducts. Journal of Herbal Pharmacotherapy 2003;3:19–32.
[2] Yang CS, Maliakal P, Meng X. Inhibition of carcinogenesis by tea. Annual Reviewof Pharmacology and Toxicology 2002;42:25–54.
[3] Ikeda M, Egawa T, Yamauchi A, Sohda Y, Koga A, Tominaga K, et al. Selec-tive serotonin reuptake inhibitors fluvoxamine and paroxetine restore forcedexercise-induced temperature dysregulation in ovariectomized mice. Euro-pean Journal of Pharmacology 2008;579:439–44.
[4] Aktas O, Prozorovski T, Smorodchenko A, Savaskan NE, Lauster R, Kloetzel PM,et al. Green tea epigallocatechin-3-gallate mediates T cellular NF-kappa B inhi-bition and exerts neuroprotection in autoimmune encephalomyelitis. Journalof Immunology 2004;173:5794–800.
[5] Haqqi TM, Anthony DD, Gupta S, Ahmad N, Lee MS, Kumar GK, et al. Preventionof collagen-induced arthritis in mice by a polyphenolic fraction from greentea. Proceedings of the National Academy of Sciences of the United States ofAmerica 1999;96:4524–9.
[6] Morinobu A, Biao W, Tanaka S, Horiuchi M, Jun L, Tsuji G, et al. (−)-Epigallocatechin-3-gallate suppresses osteoclast differentiation andameliorates experimental arthritis in mice. Arthritis and Rheumatism2008;58:2012–8.
[7] Wang J, Ren Z, Xu Y, Xiao S, Meydani SN, Wu D. Epigallocatechin-3-gallateameliorates experimental autoimmune encephalomyelitis by altering balanceamong CD4+ T-cell subsets. American Journal of Pathology 2012;180:221–34.
[8] Shankar S, Chen Q, Srivastava RK. Inhibition of PI3K/AKT and MEK/ERKpathways act synergistically to enhance antiangiogenic effects of EGCGthrough activation of FOXO transcription factor. Journal of Molecular Signaling
2008;3:7.[9] Ahmed S, Silverman MD, Marotte H, Kwan K, Matuszczak N, Koch AE. Down-regulation of myeloid cell leukemia 1 by epigallocatechin-3-gallate sensitizesrheumatoid arthritis synovial fibroblasts to tumor necrosis factor alpha-induced apoptosis. Arthritis and Rheumatism 2009;60:1282–93.

ology L
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
J.-K. Byun et al. / Immun
10] Singh R, Ahmed S, Islam N, Goldberg VM, Haqqi TM. Epigallocatechin-3-gallateinhibits interleukin-1beta-induced expression of nitric oxide synthase and pro-duction of nitric oxide in human chondrocytes: suppression of nuclear factorkappaB activation by degradation of the inhibitor of nuclear factor kappaB.Arthritis and Rheumatism 2002;46:2079–86.
11] Chen YK, Cheung C, Reuhl KR, Liu AB, Lee MJ, Lu YP, et al. Effects ofgreen tea polyphenol (−)-epigallocatechin-3-gallate on newly developed high-fat/Western-style diet-induced obesity and metabolic syndrome in mice.Journal of Agricultural and Food Chemistry 2011;59:11862–71.
12] Basu A, Sanchez K, Leyva MJ, Wu M, Betts NM, Aston CE, et al. Green teasupplementation affects body weight, lipids, and lipid peroxidation in obesesubjects with metabolic syndrome. Journal of the American College of Nutrition2010;29:31–40.
13] Brown AL, Lane J, Holyoak C, Nicol B, Mayes AE, Dadd T. Health effects of greentea catechins in overweight and obese men: a randomised controlled cross-overtrial. British Journal of Nutrition 2011;106:1880–9.
14] Tilg H, Moschen AR. Adipocytokines: mediators linking adipose tissue, inflam-mation and immunity. Nature Reviews Immunology 2006;6:772–83.
15] La Cava A, Matarese G. The weight of leptin in immunity. Nature ReviewsImmunology 2004;4:371–9.
16] Weisberg SP, McCann D, Desai M, Rosenbaum M, Leibel RL, Ferrante Jr AW.Obesity is associated with macrophage accumulation in adipose tissue. Journalof Clinical Investigation 2003;112:1796–808.
17] Fantuzzi G. Adipose tissue, adipokines, and inflammation. Journal of Allergyand Clinical Immunology 2005;115:911–9, quiz 920.
18] Gainsford T, Willson TA, Metcalf D, Handman E, McFarlane C, Ng A, et al. Leptincan induce proliferation, differentiation, and functional activation of hemopoi-etic cells. Proceedings of the National Academy of Sciences of the United Statesof America 1996;93:14564–8.
19] Matarese G, Moschos S, Mantzoros CS. Leptin in immunology. Journal ofImmunology 2005;174:3137–42.
20] Lord GM, Matarese G, Howard JK, Baker RJ, Bloom SR, Lechler RI. Lep-tin modulates the T-cell immune response and reverses starvation-inducedimmunosuppression. Nature 1998;394:897–901.
21] Jhun JY, Yoon BY, Park MK, Oh HJ, Byun JK, Lee SY, et al. Obesity aggravatesthe joint inflammation in a collagen-induced arthritis model through devia-tion to Th17 differentiation. Experimental and Molecular Medicine 2012;44:424–31.
22] Callahan LF, Wiley-Exley EK, Mielenz TJ, Brady TJ, Xiao C, Currey SS, et al.Use of complementary and alternative medicine among patients with arthritis.Preventing Chronic Disease 2009;6:A44.
23] Kikuchi M, Matsuura K, Matsumoto Y, Inagaki T, Ueda R. Bibliographi-cal investigation of complementary alternative medicines for osteoarthritis
[
etters 157 (2014) 51– 59 59
and rheumatoid arthritis. Geriatrics and Gerontology International 2009;9:29–40.
24] Ahmed S. Green tea polyphenol epigallocatechin 3-gallate in arthritis: progressand promise. Arthritis Research and Therapy 2010;12:208.
25] Inglis JJ, Simelyte E, McCann FE, Criado G, Williams RO. Protocol for the induc-tion of arthritis in C57BL/6 mice. Nature Protocols 2008;3:612–8.
26] Camps M, Ruckle T, Ji H, Ardissone V, Rintelen F, Shaw J, et al. Blockade ofPI3Kgamma suppresses joint inflammation and damage in mouse models ofrheumatoid arthritis. Nature Medicine 2005;11:936–43.
27] Rosloniec EF, Cremer M, Kang A, Myers LK. Collagen-induced arthritis. In: Col-igan JE, et al., editors. Current Protocols in Immunology 2001;Chapter 15(Unit15):15.
28] Kovacs EM, Lejeune MP, Nijs I, Westerterp-Plantenga MS. Effects of green teaon weight maintenance after body-weight loss. British Journal of Nutrition2004;91:431–7.
29] Westerterp-Plantenga MS, Lejeune MP, Kovacs EM. Body weight loss andweight maintenance in relation to habitual caffeine intake and green teasupplementation. Obesity Research 2005;13:1195–204.
30] Moon HS, Lee HG, Choi YJ, Kim TG, Cho CS. Proposed mechanisms of (−)-epigallocatechin-3-gallate for anti-obesity. Chemico-Biological Interactions2007;167:85–98.
31] Winer S, Paltser G, Chan Y, Tsui H, Engleman E, Winer D, et al. Obesity predis-poses to Th17 bias. European Journal of Immunology 2009;39:2629–35.
32] Bertola A, Ciucci T, Rousseau D, Bourlier V, Duffaut C, Bonnafous S, et al. Iden-tification of adipose tissue dendritic cells correlated with obesity-associatedinsulin-resistance and inducing Th17 responses in mice and patients. Diabetes2012;61:2238–47.
33] Wang J, Pae M, Meydani SN, Wu D. Green tea epigallocatechin-3-gallate mod-ulates differentiation of naive CD4(+) T cells into specific lineage effector cells.Journal of Molecular Medicine 2013;91:485–95.
34] Wu D, Wang J, Pae M, Meydani SN. Green tea EGCG, T cells, and T cell-mediatedautoimmune diseases. Molecular Aspects of Medicine 2012;33:107–18.
35] Wang D, Zhou Y, Lei W, Zhang K, Shi J, Hu Y, et al. Signal transducerand activator of transcription 3 (STAT3) regulates adipocyte differentiationvia peroxisome-proliferator-activated receptor gamma (PPARgamma). Biol-ogy of the Cell/Under the Auspices of the European Cell Biology Organization2010;102:1–12.
36] Sos BC, Harris C, Nordstrom SM, Tran JL, Balazs M, Caplazi P, et al. Abrogation of
growth hormone secretion rescues fatty liver in mice with hepatocyte-specificdeletion of JAK2. Journal of Clinical Investigation 2011;121:1412–23.37] Ju JH, Heo YJ, Cho ML, Jhun JY, Park JS, Lee SY, et al. Modulation of STAT-3 inrheumatoid synovial T cells suppresses Th17 differentiation and increases theproportion of Treg cells. Arthritis and Rheumatism 2012;64:3543–52.