Embed Size (px)
Citation preview

Eocene-Oligocene Transition Deep Sea Temperature
and Saturation State Changes from Benthic
Foraminiferal Trace Metal Analysis
John S. Crowe
Master of Earth Science
Marine Geoscience (International)
April 2015
School of Earth and Ocean Sciences, Cardiff University, Main Building, Park
Place, Cardiff, UK, CF10 3AT

We live on a planet that has a more or less infinite capacity to surprise. What
reasoning person could possibly want it any other way?
-Bill Bryson

3
DECLARATION
STATEMENT
This work has not previously been accepted in substance for any degree and is not being
concurrently submitted in candidature for any degree.
Signed: ________________________________ (candidate)
Date: __________________________________
STATEMENT
This dissertation is being submitted in partial fulfilment of the requirements for the degree of
Master of Earth Sciences.
Signed: ________________________________ (candidate)
Date: __________________________________
STATEMENT
This dissertation is the result of my own independent work, except where otherwise stated.
Signed: ________________________________ (candidate)
Date: __________________________________

4
Abstract
The climate transition that occurs at the Eocene-Oligocene boundary (~33.7 Ma) is marked by the first
significant Antarctic glaciation of the Cenozoic. Across the Eocene-Oligocene transition, two upward
shifts in deep sea benthic foraminiferal isotope (δ18O) values occur, Step 1 and Step 2. These shifts in
δ18O reflect a combination of bottom water temperature and δ18O of seawater. Published
paleotemperature records, calculated from foraminiferal Mg/Ca, do not show a cooling across the
Eocene-Oligocene transition, potentially indicating a saturation state effect on benthic foraminiferal
Mg/Ca during this period.
This thesis attempts to ascertain the saturation state history of Ocean Drilling Program Site 757
through the use of benthic foraminiferal trace metal climate proxies. Inductively coupled plasma mass
spectrometry was used to analyse the trace metal geochemistry of two benthic foraminifera species,
Bulimina jarvisi (n=56) and Cibicidoides havanensis (n=62). An infaunal and epifaunal benthic
foraminifera species were chosen, enabling an assessment to be made of how saturation state change
effects the different microhabitats inhabited by foraminifera. Quantification of saturation state change
enabled benthic foraminiferal Mg/Ca values to be corrected for any saturation state effect, allowing
accurate paleotemperature change to be determined. By calculating accurate changes in past bottom
water temperature, it was possible to quantify change in δ18O of seawater and thus ice growth and sea
level change.
Here we present data that shows a clear rise in infaunal (0.12 mmol/mol) and epifaunal (0.06
mmol/mol) benthic foraminiferal Mg/Ca across the Eocene-Oligocene transition. Further, a clear
increase in saturation state (18.17 µmol kg-1) was shown for the same period. By correcting Mg/Ca
paleotemperatures for saturation state change, it was found that bottom water temperature cooled
across the Eocene-Oligocene transition (~1.5 ⁰C). The timings of the cooling and warming events
across the Eocene-Oligocene transition, with regard to the two δ18O isotope shifts, do not easily
correlate with published records. To some degree, both δ18O shifts appear to be associated with
warming and ice growth. Across the Eocene-Oligocene transition, sea level has been shown to fall
(~50m), however sea level appears to rise as well as fall during this period.
This study concluded that non saturation state corrected temperatures calculated from infaunal
foraminifera are not accurate or reliable indicators of changes in bottom water temperature. Despite
accounting for saturation state change, warming periods across the Eocene-Oligocene climate
transition were discovered here. These warming periods are associated with the two shifts in δ18O
values, Step 1 and Step 2. Significant periods of ice growth were associated also with the shifts.

5
Table of Contents
1. INTRODUCTION ................................................................................................................ 8
1.1. Foraminiferal Stable Isotope Analysis ......................................................................... 9
1.1.1. Oxygen ................................................................................................................. 9
1.1.2. Carbon ................................................................................................................ 10
1.2. Carbonate System ...................................................................................................... 11
1.3. Foraminiferal Test Mass as a Proxy for Carbonate Saturation State ......................... 13
1.4. Benthic Foraminiferal Abundance as a Proxy for Productivity ................................. 14
1.5. Trace Metal/Calcium Proxies Using Benthic Foraminifera....................................... 15
1.5.1. Mg/Ca ................................................................................................................. 15
1.5.2. Li/Ca ................................................................................................................... 19
1.5.3. B/Ca .................................................................................................................... 21
1.5.4. Sr/Ca ................................................................................................................... 23
1.5.5. U/Ca .................................................................................................................... 24
1.6. Eocene-Oligocene Transition .................................................................................... 27
1.6.1. Oxygen Stable Isotope Records .......................................................................... 27
1.6.2. Trace Metal Records ........................................................................................... 28
1.6.3. Proposed Mechanisms for EOT Initiation .......................................................... 29
1.7. Regional and Geological Setting ............................................................................... 31
1.8. Benthic Foraminiferal Biostratigraphy of ODP Hole 757B ...................................... 32
1.9. Pilot Ostracod Study .................................................................................................. 33
1.10. Motivation .............................................................................................................. 37
1.10.1. Hypothesis ...................................................................................................... 37
2. MATERIALS AND METHODS ....................................................................................... 39
2.1. Sampling .................................................................................................................... 40

6
2.1.1. Species Selection ................................................................................................ 40
2.2. Microscopy ................................................................................................................ 40
2.3. Weighing .................................................................................................................... 41
2.4. Sample Preparation and Chemical Cleaning.............................................................. 41
2.4.1. Crushing and Pre-Cleaning ................................................................................. 41
2.4.2. Cleaning Procedure............................................................................................. 41
2.5. Sample Dissolution and Calcium Concentration Analysis ........................................ 44
2.6. Trace Metal Analysis ................................................................................................. 44
2.7. Age Model ................................................................................................................. 46
2.8. Calculation of Temperature, Saturation State and Ice Volume ................................. 47
3. RESULTS ........................................................................................................................... 50
3.1. Microscopy ................................................................................................................ 51
3.2. Foraminiferal Average Test Masses .......................................................................... 52
3.3. Species Abundances .................................................................................................. 54
3.4. Foraminiferal Mg/Ca ................................................................................................. 56
3.5. Foraminiferal Li/Ca ................................................................................................... 58
3.6. Foraminiferal B/Ca .................................................................................................... 60
3.7. Foraminiferal Sr/Ca ................................................................................................... 62
3.8. Foraminiferal U/Ca .................................................................................................... 64
4. DISCUSSION ..................................................................................................................... 66
4.1. Bottom Water Temperature and Saturation State History at ODP Site 757 .............. 67
4.1.1. Bottom Water Temperature Calculated Using Mg/Ca of B. jarvisi ................... 67
4.1.2. Saturation State Change Calculated Using B/Ca of C. havanensis .................... 70
4.1.3. Support for Saturation State Change from Foraminiferal Test Mass ................. 72
4.1.4. Inter-Site Comparisons of Saturation State Change ........................................... 72
4.1.5. Corrected Bottom Water Temperature Change Using C. Havanensis ............... 74

7
4.1.6. Inter-Site Comparisons of Bottom Water Temperature Change ........................ 76
4.1.7. Changes in Ice Volume across the EOT ............................................................. 78
4.1.8. Inter-Site Comparisons of δ18Osw ....................................................................... 82
4.2. Indications from the Foraminiferal Li/Ca, Sr/Ca and U/Ca Records......................... 83
4.2.1. Li/Ca ................................................................................................................... 83
4.2.2. Sr/Ca ................................................................................................................... 85
4.2.3. U/Ca .................................................................................................................... 86
4.3. Surface Productivity Changes across the EOT .......................................................... 88
5. SUMMARY ....................................................................................................................... 90
5.1. Conclusions ................................................................................................................ 91
5.2. Further Work and Future Direction ........................................................................... 93
6. REFERENCES ................................................................................................................... 94
7. ACKNOWLEDGMENTS ................................................................................................ 102
8. APENDICES .................................................................................................................... 103
8.1. Appendix 1 ............................................................................................................... 103
8.2. Appendix 2 ............................................................................................................... 105
8.3. Appendix 3 ............................................................................................................... 114

1. INTRODUCTION
8
1. INTRODUCTION
Foraminiferal stable isotope analysis
Carbonate system
Foraminiferal test mass
Foraminiferal abundance
Trace metal/calcium proxies using benthic foraminifera
Eocene-Oligocene transition
Regional and geological setting
Biostratigraphy
Pilot ostracod study
Motivation

1. INTRODUCTION
9
The creation of an accurate record of past climate is essential to modern day climate science.
Records of past climate are used to test models which predict future change. Direct records of
past climate run from the mid-15th century to present. No indication of climate change outside
the last millennium is available from direct records. The climatic conditions which prevailed
during the Eocene-Oligocene transition (EOT) are studied in this work. The Eocene-
Oligocene transition coincided with the first glaciation of Antarctica during the Cenozoic. The
climatic conditions are ascertained by the use of foraminiferal geochemical proxies. This
introduction provides information on the proxies used, the regional setting of the sample sites
and the aims of the project.
1.1. Foraminiferal Stable Isotope Analysis
1.1.1. Oxygen
Oxygen stable isotope analysis is the most established geochemical proxy for recording past
climate conditions from foraminifera. Foraminiferal oxygen isotopes (δ18O) record both
temperature and sea water isotopic composition (δ18OSW). Ice growth increases δ18OSW
because 16O is concentrated in ice sheets by the hydrological cycle (Pearson, 2012). Melting
ice returns O16 dominated meltwater to the oceans, changing the ratio of the 16O to 18O. A
change in δ18O values can thus be temperature or ice growth related, providing an excellent
method for calculating past temperatures and ice volumes. A caveat when using stable oxygen
isotope ratios is that, in order to calculate either temperature or ice volume, the other variable
must be known. This variable can be calculated using other geochemical proxies, however
these are less accurate than using stable isotopes alone. During periods where no ice volume
change occurred, the 18O of a foraminifera may be used independently of seawater 18O to
quantify temperature. The 18O of a sample is calculated by comparing the ratio of both
oxygen isotopes to a standard of known isotopic composition (Equation 1.1).
EQN. (1.1)
18O sample (‰) = [(
𝑂18
𝑂16) 𝑠𝑎𝑚𝑝𝑙𝑒 − (𝑂18
𝑂16) 𝑠𝑡𝑎𝑛𝑑𝑎𝑟𝑑
(𝑂18
𝑂16) 𝑠𝑡𝑎𝑛𝑑𝑎𝑟𝑑]
The relationship between foraminiferal 18O, temperature and the 18O of seawater was
shown by Epstein et al. (1953) to follow equation 1.2:

1. INTRODUCTION
10
EQN. (1.2)
𝑇(0𝐶) = 𝐴 + 𝐵(δ18𝑂𝑠𝑎𝑚𝑝𝑙𝑒 − δ18𝑂𝑆𝑒𝑎𝑤𝑎𝑡𝑒𝑟) + 𝐶(δ18𝑂𝑠𝑎𝑚𝑝𝑙𝑒 − δ18𝑂𝑆𝑒𝑎𝑤𝑎𝑡𝑒𝑟)2
Where A, B and C represent experimentally derived coefficients.
In practice these relationships equate to ~0.21-0.23 ‰ decrease in the δ18O of calcite for
every 1 ⁰C increase in temperature.
1.1.2. Carbon
Two commonly used carbon isotopes (C12 and C13) are helpful in unravelling past climatic
trends. Equation 1.3 shows how 13C of a foraminifera is calculated from the isotopic ratio of
C13 to C12 and a standard of known isotopic composition (Rohling and Cooke, 2003). This
proxy monitors past changes in carbon reservoirs. Three major signals are recorded by 13C.
Firstly, isotopically light carbon is added to bottom waters when organic matter is oxidised.
Secondly, younger waters are isotopically heavier than older waters. If differently aged waters
are mixed a new 13C value is created. Finally, high productivity concentrates C12 within
organic matter. This process in turn concentrates C13 within the total dissolved carbonate that
reaches the bottom waters. Therefore, at any one time, benthic foraminiferal 13C may record
the 13C of the total dissolved carbonate in the a water mass, ocean circulation patterns and
productivity change (Ravelo and Hillaire-Marcel, 2007).
EQN. (1.3)
13C sample (‰) = [(
𝐶13
𝐶12) 𝑠𝑎𝑚𝑝𝑙𝑒 − (𝐶13
𝐶12) 𝑠𝑡𝑎𝑛𝑑𝑎𝑟𝑑
(𝐶13
𝐶12) 𝑠𝑡𝑎𝑛𝑑𝑎𝑟𝑑]

1. INTRODUCTION
11
1.2. Carbonate System
An understanding of the carbonate system is required in order to decipher the mechanisms
controlling the Eocene-Oligocene Transition climate. The carbonate system plays an
important role in calcareous microfossil shell chemistry, a key tool used to identify past
climatic events.
Calcium carbonate (CaCO3) solubility increases in waters with low pH or low temperatures or
increased pressure. The point at which no more CaCO3 can be dissolved into solution is
termed the calcium carbonate saturation point. A water mass containing a higher
concentration of solute than the saturation point is oversaturated while a water mass with a
lower concentration is termed undersaturated. Therefore, for dissolution to occur, a water
mass must be undersaturated with respect to calcium carbonate and environmental solubility
conditions must be favourable.
The saturation state of seawater (Ω) was summarized by (Barker et al., 2003a) as:
EQN. (1.4)
Ω = [CO3
2−][C𝑎2+]
𝐾𝑠𝑝
where a Ω value of 1 represents the saturation point, Ω > 1 representing oversaturation and Ω
< 1 undersaturation, while Ksp represents the stoichiometric solubility product for CaCO3 (a
function of temperature and pressure). The surface oceans are supersaturated with respect to
both [Ca2+] and [CO32-]. Supersaturation means that the surface oceans contain more of these
ions than could be dissolved by the ocean under normal atmospheric conditions. As [Ca2+] is
broadly conservative in the Earth’s oceans it is the non-conservative [CO32-] that controls the
saturation state of calcium carbonate (Barker et al., 2003a).

1. INTRODUCTION
12
Changing partial pressures of atmospheric CO2 (PCO2atm) affect ocean carbonate chemistry
through the following reactions:
EQN. (1.5)
𝐶𝑂2 𝑎𝑡𝑚 + 𝐻2𝑂 ↔ 𝐻2𝐶𝑂3 ↔ 𝐻+ + 𝐻𝐶𝑂3
− ↔ 2𝐻+ + 𝐶𝑂3 2−
When CO2 dissolves in the oceans, carbonic acid (𝐻2𝐶𝑂3) is formed. Subsequently, the
carbonic acid disassociates into bicarbonate (𝐻𝐶𝑂3 −) and hydrogen (H+) ions. Le Chatelier's
principle states, any change to one side of a reaction will promote an opposing reaction, which
will shift equilibrium thus minimizing the effect of the change. With this in mind, an increase
in PCO2atm will result in the reactions presented in equation 1.5 shifting toward the right. The
[𝐶𝑂3 2−] will be lowered which will lower saturation state, promoting dissolution of CaCO3.
The carbonate compensation depth (CCD) is the depth of the water column at which rate of
dissolution of CaCO3 is equal to the rate of accumulation. Below the CCD, there is no
preservation of calcareous planktonic or benthic skeletons. Another feature of the water
column is the lysocline. At the lysocline, the rate of dissolution increases dramatically. Above
the lysocline, calcareous microfossils remain practically unaltered. The CCD and lysocline are
key indicators of changes in the atmosphere due to the explicit relationship between PCO2atm
and saturation state change (Δ[CO32−]).

1. INTRODUCTION
13
1.3. Foraminiferal Test Mass as a Proxy for Carbonate Saturation State
Planktonic foraminiferal test mass has been correlated with [𝐶𝑂3 2−]. The study presented in
Barker and Elderfield (2002) investigated foraminiferal mass across Termination 1 (the last
glacial-interglacial change). It was suggested that foraminiferal growth rate is a function of
[𝐶𝑂3 2−]. A high [𝐶𝑂3
2−] infers a low rate of dissolution, which means there are a greater
number of carbonate ions that foraminifera may utilise to build tests. Therefore, organisms,
which utilize calcium carbonate, are found to be heavier when the saturation state is high. The
strong correlation between planktonic foraminiferal test mass and [𝐶𝑂3 2−] shown by Barker
and Elderfield (2002) was attributed to this growth rate mechanism (Fig. 1.1).
Figure 1.1 Size normalised weights of the planktonic foraminifera Globorotalia bulloides
against [𝐶𝑂3 2−] from (Barker and Elderfield, 2002).
The relationship between planktonic foraminiferal test mass and [𝐶𝑂3 2−] was observed also in
benthic test mass (Williams, 2008). Using the species Melonis barleeanum, it was found that
test mass decreased with decreasing saturation state. This trend is the same as the one
presented for planktonic foraminifera (Barker and Elderfield, 2002). Williams (2008)
investigated the 1 km deepening of the CCD during the EOT, discovered by Van Andel
(1975). This deepening of the CCD is known to have increased saturation state at depth.
Increased benthic foraminifera test mass indicates higher calcification rate at Ocean Drilling
Program (ODP) Site 1218 for the EOT (Williams, 2008). The site 1218 data supports the use
of benthic foraminiferal test mass as an indicator of carbonate ion change.

1. INTRODUCTION
14
1.4. Benthic Foraminiferal Abundance as a Proxy for Productivity
Organic carbon export flux is vital to sustain most benthic life. The majority of any organic
export flux reaching the seabed is used by the benthic ecosystem upon deposition. Labile
organic matter is the most quickly and easily consumed part of the particulate organic matter
(POM) deposited on the deep-sea floor. POM is a major source of non-living food for
foraminifera (Loubere and Fariduddin, 1999). Therefore, a correlation exists between organic
export flux and benthic biomass (Van Der Zwaan et al., 1999). As an estimate, deposition of 1
mg of organic carbon correlates to deposition of one benthic foraminiferal shell >150 µm
(Herguera and Berger, 1991). This relationship allows benthic foraminiferal abundance to be
used to indicate organic carbon flux to the sediment. Benthic foraminiferal records can thus be
used as a proxy for changes in ocean productivity as deposition of organic carbon is a
consequence of productivity in the surface oceans (Herguera, 1992). However, because other
environmental variables (e.g. temperature, salinity) affect export flux, the use of benthic
foraminiferal abundance as a proxy is limited (Murray, 2001). Indications of past surface
productivity are restricted to waters below 1000 m where environmental factors show little
variance (Altenbach et al., 1999).
Regions of high productivity and sustained export flux to the sediment are synonymous with
certain genera of foraminifera (Thomas et al., 1995). One such genus is Bulimina. Further
investigation led Fariduddin and Loubere (1997) to suggest that high productivity surface
waters are characterized by high abundances of all infaunal foraminifera. Infaunal
foraminifera are those that live in the substrate while epifaunal species inhabit the substrate
surface.

1. INTRODUCTION
15
1.5. Trace Metal/Calcium Proxies Using Benthic Foraminifera
Since the turn of the 20th century, there has been sustained research into trace metal chemistry
in biogenic carbonate. As early as 1917, it was proposed that temperature may play a role in
the concentration of trace metals found within biogenic calcite (Clarke and Wheeler, 1922).
As a tool for paleooceanographic and climate science, this knowledge had a restricted use for
many years because the quantity of material needed for analysis on early mass spectrometers
far exceeded that available at any sites of interest. Trace metal chemistry became a useful tool
for the paleooceanographic and climate fields by the 1970s. Both methods and equipment,
used for trace metal analysis, had advanced enough for trace metal chemistry to become a
useful tool. Early investigations focused on analysing planktonic foraminifera (Savin and
Douglas, 1973, Bender et al., 1975). It was not until 1988 that a use for the trace metal
composition of benthic foraminifera was proposed. Izuka (1988) suggested temperature as the
primary control on magnesium within the benthic foraminifera Cassidulina spp. Since then
trace metal proxies involving elements including lithium, boron, strontium and uranium have
all been developed. While all of these proxies have significant caveats to their use, trace metal
geochemistry remains a powerful instrument in deciphering ocean and climate history. The
following sub-sections aim to outline the uses, advantages and possible pitfalls of the major
benthic foraminiferal trace metal proxies.
1.5.1. Mg/Ca
Incorporation of Mg in Benthic Foraminifera
The most commonly used benthic foraminifera trace metal proxy involves the ratio of
magnesium to calcium contained within a foraminifera test. A positive relationship between
Mg/Ca and temperature exists in planktonic foraminifera (Nürnberg et al., 1996). This
relationship was reproduced in benthic foraminifera, collected from the Little Bahamas Bank,
by Rosenthal et al. (1997) (Fig. 1.2). The relationship has allowed foraminiferal Mg/Ca to be
used as a paleothermometer. A number of other variables have been shown to affect
incorporation of magnesium into foraminifera tests. The inconsistency found between Mg/Ca
concentrations of inorganic and biogenic calcite formed under different temperatures infers
non temperature controls on magnesium incorporation. Vital effects (Rosenthal et al., 1997),

1. INTRODUCTION
16
Δ[CO32−] (Martin et al., 2002) and salinity (Ferguson et al., 2008) have all been proposed as
significant variables affecting the ratio of magnesium to calcium in both planktonic and
benthic foraminifera. However, the effect of salinity on benthic foraminifera is yet to be
quantified.
Figure 1.2 Mg/Ca incorporation into the benthic foraminiferal species Cibicidoides
pachyderma (floridanus) as a function of water temperature. The solid line represents an
exponential fit while the dashed lines are a forecast interval at the 96% level from (Rosenthal
et al., 1997).
Limitations
The use of Mg/Ca as a paleothermometer is complicated by a number of factors. Vital effects
specific to individual species of foraminifera have been shown to affect incorporation of
magnesium into the test calcite (Lear et al., 2002). Genetic variability exists between different
foraminifera species. This may account for the differences in magnesium incorporation
observed. However, genetic variability affecting Mg/Ca ratios is yet to be proven. Interspecies
differences in Mg/Ca are corrected for using species specific calibrations.

1. INTRODUCTION
17
Diagenetic processes, the physical and chemical changes that occur after burial, may also
contribute to the concentration of Mg/Ca within each shell. Of note is dissolution, which may
lead to lower mg/ca ratios. Brown and Elderfield (1996) showed that calcite containing more
trace metals dissolved preferentially over pure calcite. The joint processes of neomorphism
and cementation may also alter Mg/Ca ratios. These processes raise Mg/Ca by replacing
biogenic calcite with the inorganic form of the crystal. Inorganic calcite has a higher ratio of
Mg/Ca due to a larger participation coefficient for the magnesium ion (Katz, 1973).
Investigation of deeply buried foraminifera showed only a small increase in Mg/Ca between
poorly and well preserved foraminifera. Therefore, it is likely diagenetic effects can be
disregarded due to their minimal impact.
A salinity related impact on Mg/Ca has been discovered in planktonic foraminifera inhabiting
high salinity areas (Rosenthal et al., 2000). This impact was supported by Ferguson et al.
(2008) who suggested salinity may have significant implications for the interpretation of
downcore records. The effect of salinity on benthic foraminifera is unknown. However, there
is unlikely to be any salinity based effect in low salinity areas.
Changes in the proportion of Mg2+ to Ca2+ in seawater can affect the incorporation magnesium
into a test. There are no direct records of seawater Mg/Ca. This makes indirect evidence vital
to the Mg/Ca paleothermometer. Attempts have been made to ascertain past seawater Mg/Ca
using evaporite fluid inclusions (Lowenstein et al., 2001), Mg/Ca from biogenic carbonates
formed during the greenhouse climates of the early Cenozoic (Lear et al., 2002), models of
processes affecting Mg/Ca in the oceans (Demicco et al., 2005) and calcium carbonate veins
(Coggon et al., 2010). The application of these methods has proved difficult and a large range
of values representing past seawater Mg/Ca have been produced. The relatively long
residence times of Mg2+ and Ca2+ in the ocean (~1 and ~10 Ma respectively) allows
temperature calculations to be relatively accurate despite questions remaining as to the
concentrations in past seawater (Broecker et al., 1982). By adding a seawater concentration
component to Mg/Ca calibration equations the effect of changes is minimised.
The final and largest limitation of using test Mg/Ca as a paleothermometer is saturation state
change. Saturation state change has been offered as a mechanism for Mg/Ca records where
Mg/Ca rises when a fall is expected due to negative temperature change (Coxall et al., 2005).
The saturation state change hypothesis was proposed by Elderfield et al. (2006), who showed

1. INTRODUCTION
18
that the foraminifera Cibicidoides wuellerstorfi has a Mg/Ca sensitivity to saturation state
change of 0.0086 ± 0.0006 mmol/mol/μmol/kg. A higher saturation state increases the ratio of
Mg/Ca within benthic foraminifera. Accurate Mg/Ca paleothermometry requires
quantification of saturation state change in order to be effective. It has been proposed that
where saturation state change cannot be quantified, infaunal species of benthic foraminifera
can be used because they are relatively buffered against saturation state change (Elderfield et
al., 2010).
Calibration
A number of species specific calibrations exist in order to calculate temperature from benthic
foraminiferal Mg/Ca. Using table 1.1 and equation 1.6, bottom water temperature can be
quantified for individual foraminifera species. Almost all benthic Mg/Ca calibrations are
given an exponential fit and therefore use the general equation:
EQN. (1.6)
𝑀𝑔/𝐶𝑎𝐹𝑂𝑅𝐴𝑀 = 𝐵 exp (𝐴 × 𝑇)
where T is temperature. A and B can be found in table 1.1. Lear (2007) proposed the addition
of a past seawater Mg/Ca component to this general equation (Equation 1.7) so as to account
for changes in ocean chemistry.
EQN. (1.7)
𝑀𝑔/𝐶𝑎𝐹𝑂𝑅𝐴𝑀 = 𝑀𝑔/𝐶𝑎𝑠𝑤−𝑇
𝑀𝑔/𝐶𝑎𝑠𝑤−0 × 𝐵 exp (𝐴 × 𝑇)
where sw-0 is modern Mg/Ca in seawater and sw-T is estimated Mg/Ca in seawater for
sample age. When investigating short term shifts in Mg/Ca, past seawater concentrations of
magnesium and calcium are not required due to their reasonably long residence times.
Evidence suggest that early Cenozoic seawater Mg/Ca was not less than two thirds of today’s
value of approximately 5.2 mol/mol (Lear et al., 2002).
Care should be taken because the Mg/Ca of some species has been shown to agree more
readily with a linear fit than an exponential fit (Marchitto et al., 2007). A linear fit produces
unrealistic bottom water temperatures for other species (Lear et al., 2008).

1. INTRODUCTION
19
The accuracy of the early calibrations of Rosenthal et al. (1997), using data collected from the
Little Bahamas Bank, has been called into question due to inorganic calcite contamination
(Lear et al., 2002). More calibration studies are required to create equations for new species as
well as to confirm the accuracy of previous studies.
Table 1.1 Correlation coefficients used for calibration of temperature from foraminiferal calcite Mg/Ca
Species A B Source
Exponential
(Mg/Ca = B x
expAT)
C. pachyderma 0.1 1.36 (Rosenthal et al., 1997)
C. pachyderma, C. wuellerstorfi 0.11 0.85 (Martin et al., 2002)
C. pachyderma, C. wuellerstorfi, C.
compressus
0.11 0.87 (Lear et al., 2002)
Planulina Spp. 0.12 0.79 (Lear et al., 2002)
Oridorsalis umbonatus 0.114 1.01 (Lear et al., 2002)
Oridorsalis umbonatus 0.09 1.53 (Rathmann et al., 2004)
Melonis spp. 0.101 0.98 (Lear et al., 2002)
Planulina ariminensis 0.062 0.91 (Lear et al., 2002)
Uvigerina Spp. 0.061 0.92 (Lear et al., 2002)
Linear
(Mg/Ca = AT
+B)
C.pachyderma 0.14 1.2 (Marchitto et al., 2007)
Planoglabratella opercularis 2.9 89.7 (Toyofuku et al., 2000)
Quinqueloculina yabei 1.6 66 (Toyofuku et al., 2000)
Planoglabratella opercularis 0.0034 81.5 (Toyofuku and Kitazato,
2005)
1.5.2. Li/Ca
Incorporation of Li in Benthic Foraminifera
Li/Ca ratios in benthic foraminifera can be used as a proxy for Δ[CO32−]. Downcore Li/Ca
foraminiferal records from planktonic and benthic species showed a decrease of 13-14%
across the Pleistocene deglaciation (Hall and Chan, 2004). Systematic changes in Li/Ca with

1. INTRODUCTION
20
δ18O were too large to be temperature derived alone, another variable must be affecting Li/Ca
(Hall and Chan, 2004). As an alternative, carbonate saturation state has been proposed as the
driver for variability seen in Li/Ca records. A link between benthic foraminiferal Li/Ca and
carbonate saturation state was presented by Lear and Rosenthal (2006) using core top data
from a water depth transect in the Norwegian Sea. This core top data suggests a linear
relationship between Li/Ca and carbonate saturation state (Fig. 1.3). Comparisons between
downcore records and the Van Andel (1975) historic CCD data led to the suggestions that
Li/Ca in benthic foraminifera is dependent on Δ[CO32−] (Lear and Rosenthal, 2006).
Figure 1.3 Record of Li/Ca in the benthic foraminifera Oridorsalis umbonatus, temperature
(thick line) and Δ[CO32−] (thin line) plotted against water depth (boxes) adapted from (Lear
and Rosenthal, 2006). Scatter points represent changing benthic foraminiferal Li/Ca with
water depth.
Limitations
The effectiveness of this proxy is limited by a number of factors. Firstly, the concentration of
Li/Ca in benthic foraminifera has a temperature component. Li/Ca decreases with increasing
temperature (Marriott et al., 2004a, Marriott et al., 2004b). Therefore, any quantification of
changes in Δ[CO32−] requires the temperature component to be calculated using a more

1. INTRODUCTION
21
constrained proxy, such as Mg/Ca. An initial attempt to decipher EOT temperature and
Δ[CO32−] from deep sea benthic foraminiferal Mg/Ca and Li/Ca was presented in Lear et al.
(2010). The attempt was not entirely successful in splitting apart the temperature and
Δ[CO32−] signals, succeeding at some sites but not at others. Secondary effects and calibration
thresholds were proposed as the cause of any failures. Secondly, when applied over periods
longer than 1.5 Ma (the approximate residence time of lithium in the oceans (Huh et al.,
1998)), changes in seawater concentration of lithium/calcium must be considered. A seawater
lithium record covering the past 68 Ma showed an increase of 9 ppm (Misra and Froelich,
2012). This record may be of value with respect to the paired Li/Ca Mg/Ca proxy. Thirdly,
species specific vital effects are known to affect trace metal uptake in benthic foraminifera.
This means species specific calibrations are needed to quantify Δ[CO32−].
Calibration
To date, the only calibration using benthic foraminiferal Li/Ca and Mg/Ca to unravel
Δ[CO32−] uses Oridorsalis umbonatus (Lear et al., 2010). The bottom water Δ[CO3
2−] is
calculated from O. umbonatus Li/Ca and Mg/Ca using the following equation:
EQN. (1.8)
Δ[𝐶𝑂3 2−] =
ΔMg + 0.162ΔLi
0.0162
Using this approach the bottom water Δ[CO32−] can be calculated if the change in magnesium
and lithium are known.
1.5.3. B/Ca
Incorporation of B in Benthic Foraminifera
As with Li/Ca, boron/calcium can be used as proxy for Δ[CO32−]. Yu and Elderfield (2007)
showed that benthic foraminiferal B/Ca is linearly correlated with Δ[CO32−] as described in
equation 1.9.
EQN. (1.9)
𝐵: 𝐶𝑎 = 𝐴 × Δ[C𝑂3 2−] + 𝐵

1. INTRODUCTION
22
while values for A and B can be found in table 1.2. The response of epifaunal benthic
foraminiferal B/Ca to Δ[CO32−] is well documented (Yu and Elderfield, 2007, Yu et al., 2010,
Brown et al., 2011). The pore waters, which infaunal species inhabit, are often saturated with
respect to the carbonate ion (Martin and Sayles, 1996). As with infaunal Mg/Ca (Elderfield et
al., 2010), infaunal boron appears to be relatively buffered against Δ[CO32−].
Limitations
As with other foraminiferal trace metal proxies, the usefulness of B/Ca is limited by a number
of factors. Firstly, infaunal B/Ca is expected to be low concentration (Uvigerina spp. B/Ca
∼20 µmol/mol) (Yu and Elderfield, 2007). Therefore, the concentrations recovered may be
under the detection limits of traditional foraminiferal trace metal methodologies. A method
for collecting more precise data has been proposed (Misra et al., 2014). Without such a
method, infaunal B/Ca records are not suitable for analysis. Secondly, concentrations of B/Ca
in the oceans will affect the B/Ca record in foraminiferal calcite. Boron has a long residence
time in the oceans. Lemarchand et al. (2000) calculated a boron residence time of ~14 Ma.
The model presented in Lemarchand et al. (2002) predicts a relatively stable concentration of
oceanic boron across the Cenozoic (~10% variation). Thirdly, species specific vital effects are
known to affect trace metal uptake in benthic foraminifera where there is a large variance in
B/Ca between species. Cases of different B/Ca have been reported between morphotypes of
the same species adding further difficulty when trying to empirically ascertain Δ[CO32−] (Rae
et al., 2011). This means species specific calibrations are needed to quantify Δ[CO32−].
Calibration
Using equation 1.9 and table 1.2, the bottom water Δ[CO32−] can be quantified using
foraminiferal B/Ca. The large range of the coefficients shown in table 1.2 highlights the need
for species specific calibrations. The infaunal species O. umbonatus and Uvigerina spp. have
markedly lower calibration coefficients than the epifaunal species. The B/Ca responses to
Δ[CO32−] for Nuttallides umbonifera and Cibicidoides wuellerstorfi are comparable while the
remaining epifaunal species, Cibicidoides mundulus, has significantly lower coefficients.

1. INTRODUCTION
23
Table 1.2 Correlation coefficients used for calibration of Δ[CO32−] from foraminiferal calcite B/Ca
Species A B Source
Nuttallides umbonifera 1.23 ± 0.15 134 ± 2.7 (Brown et al., 2011)
Cibicidoides wuellerstorfi 1.14 ± 0.048 177.1 ± 1.41 (Yu and Elderfield, 2007)
Cibicidoides mundulus 0.69 ± 0.072 119.1 ± 2.62 (Yu and Elderfield, 2007)
Oridorsalis umbonatus 0.29 ± 0.2 56.4 ± 5.5 (Brown et al., 2011)
Uvigerina spp. 0.27 ± 0.076 19.4 ± 2.99 (Yu and Elderfield, 2007)
1.5.4. Sr/Ca
Incorporation of Sr in Benthic Foraminifera
Another possible proxy for Δ[CO32−] is the Sr/Ca ratio found in foraminifera shells. Core top
studies of benthic foraminiferal Sr/Ca have shown a negative correlation between shell Sr/Ca
and water depth (Rosenthal et al., 1997, Lear et al., 2003, Yu et al., 2014). Unlike B/Ca and
Li/Ca, there no evidence for a temperature component controlling Sr/Ca concentrations in
calcitic benthic foraminifera. The lack of temperature dependency suggests pressure or
Δ[CO32−] is the dominant control on Sr/Ca concentrations in calcitic foraminifera. Strontium
provides a unique record of Δ[CO32−], albeit a limited one.
Limitations
The partitioning of strontium in calcitic foraminifera is poorly understood. It is highly likely
that species specific vital effects play a role in strontium incorporation as with other trace
metal proxies. To ascertain the Δ[CO32−] reliably from foraminiferal Sr/Ca, the past
concentration of strontium in the ocean must be calculated. Using the Sr/Ca record from a
gastropod, Sosdian et al. (2012) calculated past seawater strontium concentration and
suggested that there were minimal variations from the Eocene to the Pliocene.
Calibration
Over periods where seawater strontium concentration remains constant, the work of Yu et al.
(2014) advised that, C. wuellerstorfi and C. mundulus Sr/Ca is controlled mainly by Δ[CO32−]

1. INTRODUCTION
24
(Fig. 1.4). Therefore, Sr/Ca from benthic foraminifera may be used as an auxiliary proxy,
despite no calibration existing to quantify Δ[CO32−].
Figure 1.4 The response of foraminiferal Sr/Ca to deep water Δ[CO32−].
(A) C. wuellerstorfi Sr/Ca and (B) C. mundulus Sr/Ca. Error bars represent ±1σ of
replicates. DSr = (Sr/Ca) shell / (Sr/Ca) seawater. Solid lines in A and B represent the best linear
fits for each data set from (Yu et al., 2014).
1.5.5. U/Ca
Incorporation of U in Benthic Foraminifera
Initial studies of Uranium within benthic foraminifera shells, by Russell et al. (1994),
indicated that U/Ca is controlled by seawater uranium concentration only. Later investigations
showed a marked increase in U/Ca (~25%) from the last glacial period to the Holocene
(Russell et al., 1996). The authors found a correlation between downcore records of U/Ca and
Mg/Ca, this strongly points toward changes in metal incorporation or dissolution as opposed
1. INTRODUCTION
25
to changes in seawater uranium concentration. Anoxic exchange with pore water uranium was
ruled out due to the oxic depositional environment of the studied cores. Dissolution effects are
discounted because well preserved cores with opposite depositional histories show the same
trend. The assumption of temperature as the dominant control on U/Ca in foraminifera is
proposed (Yu et al., 2008). This proposal is supported by the finding that planktonic
foraminiferal U/Ca is inversely proportional to temperature (Yu et al., 2008). Further work by
Russell et al. (2004), with planktonic foraminifera, showed a strong negative correlation
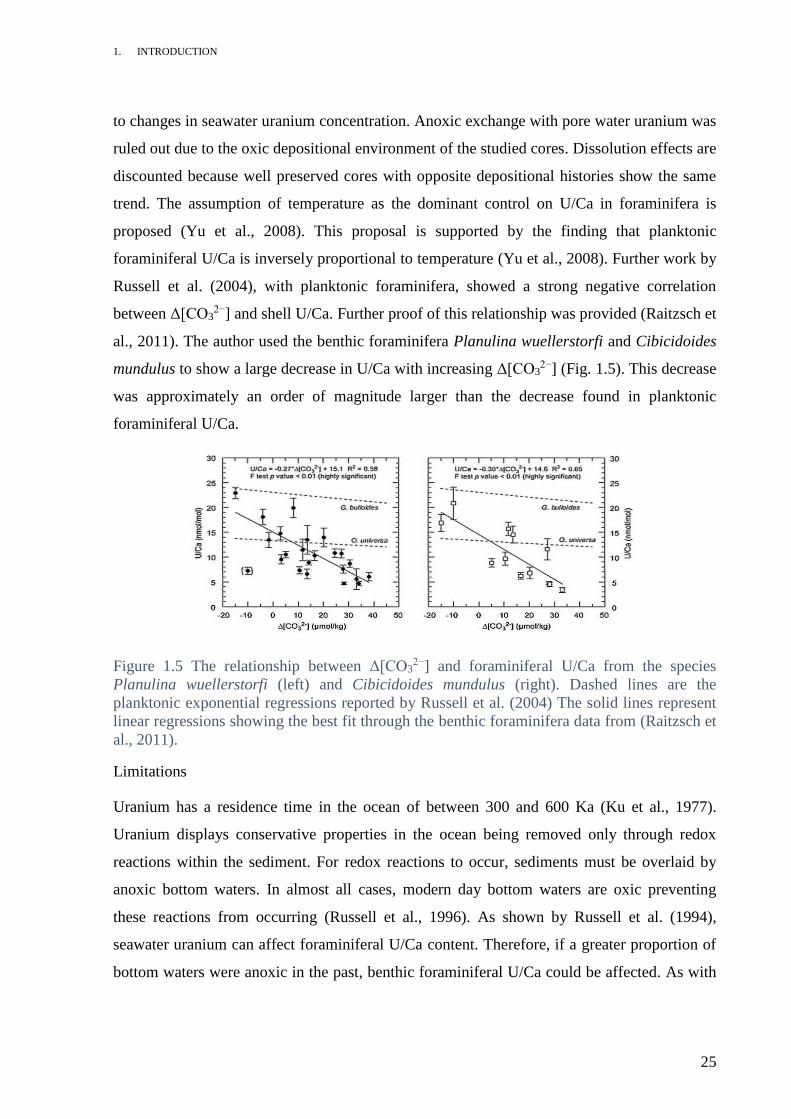
between Δ[CO32−] and shell U/Ca. Further proof of this relationship was provided (Raitzsch et
al., 2011). The author used the benthic foraminifera Planulina wuellerstorfi and Cibicidoides
mundulus to show a large decrease in U/Ca with increasing Δ[CO32−] (Fig. 1.5). This decrease
was approximately an order of magnitude larger than the decrease found in planktonic
foraminiferal U/Ca.
Figure 1.5 The relationship between Δ[CO32−] and foraminiferal U/Ca from the species
Planulina wuellerstorfi (left) and Cibicidoides mundulus (right). Dashed lines are the
planktonic exponential regressions reported by Russell et al. (2004) The solid lines represent
linear regressions showing the best fit through the benthic foraminifera data from (Raitzsch et
al., 2011).
Limitations
Uranium has a residence time in the ocean of between 300 and 600 Ka (Ku et al., 1977).
Uranium displays conservative properties in the ocean being removed only through redox
reactions within the sediment. For redox reactions to occur, sediments must be overlaid by
anoxic bottom waters. In almost all cases, modern day bottom waters are oxic preventing
these reactions from occurring (Russell et al., 1996). As shown by Russell et al. (1994),
seawater uranium can affect foraminiferal U/Ca content. Therefore, if a greater proportion of
bottom waters were anoxic in the past, benthic foraminiferal U/Ca could be affected. As with

1. INTRODUCTION
26
all of the discussed trace metal proxies, there appears to be significant inter species vital
effects controlling incorporation of uranium into foraminifera. This can be resolved by using
species specific calibrations. However few of these currently exist.
Calibration
P. wuellerstorfi and C. mundulus U/Ca is mainly controlled by Δ[CO32−] (Raitzsch et al.,
2011). The authors believe that any correlation with temperature, as shown empirically by Yu
et al. (2008), is due to the relationship between temperature and Δ[CO32−], suggesting that
temperature is not a direct control on U/Ca in benthic foraminifera. Therefore, Raitzsch et al.
(2011) proposed the following linear equation as a suitable calibration of Δ[CO32−] from
U/Ca:
EQN. (1.10)
𝑈: 𝐶𝑎 = 𝐴 × Δ[C𝑂3 2−] + 𝐵
Table 1.3 contains the species specific correlation coefficients for use with equation 1.10
when calculating Δ[CO32−] from benthic foraminiferal U/Ca.
Table 1.3 Correlation coefficients used for calibration of Δ[CO32−] from foraminiferal calcite
U/Ca
Species A B Source
Planulina wuellerstorfi -0.27 ± 0.04 15.1 ± 1.4 (Raitzsch et al., 2011)
Cibicidoides mundulus -0.30 ± 0.06 14.6 ± 1.8 (Raitzsch et al., 2011)

1. INTRODUCTION
27
1.6. Eocene-Oligocene Transition
The Eocene-Oligocene (E-O) boundary (~34 Ma) is known as Earth’s greenhouse-icehouse
transition (Francis et al., 2008). During the Eocene-Oligocene transition (EOT), the first
Cenozoic ice sheets appeared on the Antarctic continent (Kennett and Shackleton, 1976). The
Eastern Antarctic continent was entirely buried under ice by the onset of the Oligocene
(Ehrmann and Mackensen, 1992). After decades of study, terminology associated with the
EOT has become confused. The terminology used here follows that presented in Lear et al.
(2008). The E-O boundary is formally identified by the extinction event associated with the
planktonic foraminiferal family Hankeninidae (Coccioni et al., 1988).
1.6.1. Oxygen Stable Isotope Records
A geochemical signal exists recording the EOT. Two shifts exist in deep water foraminiferal
oxygen isotope values (δ18O), Oligocene isotope shift 1 (Oi-1) (termed here as Step 2),
represents the shift to maximum values of δ18O at the Early Oligocene Glacial Maximum
(EOGM) while a precursor shift , termed here as Step 1, is also observed (Fig. 1.6a) (Coxall et
al., 2005). Coxall et al. (2005) noted that the oxygen isotope shifts Step 1 and Step 2
coincided with a ~1 km deepening of the carbonate compensation depth (CCD) discovered by
Van Andel (1975) (Fig. 1.6b).
Figure 1.6(a) high-resolution data from ODP site 1218 showing a stepwise δ18O increase in
benthic foraminiferal calcite and (b) published CCD data from Deep Sea Drilling Program
(DSDP) sites across the last 50 Ma (Van Andel, 1975). The figure shows a ~1 km deepening
of the CCD at the E-O boundary (timing and duration of the shift is poorly constrained). The
shaded bar represents an uncertainty of ~3 Ma adapted from (Coxall et al., 2005).
Step 2 Step 1
(a) (b)
δ18
O (‰ VPDB)

1. INTRODUCTION
28
The ~1 km deepening of the CCD is associated with the drawdown of atmospheric carbon
dioxide (CO2atm). The magnitude of this drawdown (~25 µatm) is unlikely large enough to
initiate large scale glaciation of Antarctica (Sigman and Boyle, 2000). It appears more likely
that the increase in CCD is glaciation related. One theory is that the availability of continental
shelf (where neritic benthic calcifiers are found) decreased upon glaciation of Antarctica,
which shifted CaCO3 deposition into the pelagic deep sea realm from the benthic neritic realm
(James, 1978, Merico et al., 2008).
1.6.2. Trace Metal Records
The size of shift in the δ18O record is not accounted for by Antarctic glaciation. Either
significant Northern Hemisphere ice accumulation was occurring concurrently with Antarctic
glaciation or there was a temperature component to the δ18O shift. Climate modelling does not
support Northern Hemisphere ice growth across the EOT (Deconto et al., 2008), although
some minor glaciation has been discovered on Greenland during the period (Eldrett et al.,
2007). For bipolar glaciation to occur, the models of Deconto et al. (2008) predict a much
lower PCO2atm than occurred at the EOT. Therefore, it follows that a significant cooling event
impacted upon records of δ18O during the EOT.
Concentration of magnesium with respect to calcium is known to be temperature dependant,
with increased magnesium calcium representing increasing temperature (Burton and Walter,
1991). Foraminiferal Mg/Ca can therefore be used as a paleothermometer. This enables the
temperature and δ18OSW components of δ18O to be separated. Using foraminiferal Mg/Ca
ratios from very well preserved Tanzanian foraminifera deposited well above the CCD, it has
been indicated that Step 1 was predominantly associated with cooling global temperatures
(~2.5⁰C) (Lear et al., 2008), while Step 2 was linked to rapid ice growth and sea level decline
(Coxall et al., 2005). Microfacies, sedimentological and biotic analysis also confirm the
existence of a coupled cooling and ice growth stimulus for initial changes in δ18O with no
sustained cooling associated with Step 2 (Houben et al., 2012). The cooling trend at the first
isotope shift is supported by planktonic foraminiferal Mg/Ca proxy information from ODP
sites 738, 744, and 748 (the Kerguelen Plateau) (Bohaty et al., 2012). However the cooling
event is not observed in benthic foraminiferal Mg/Ca ratios at the same sites.

1. INTRODUCTION
29
Early attempts to apply Mg/Ca paleothermometry to the EOT also supplied no evidence of a
cooling trend from the Eocene to the Oligocene (Lear et al., 2004, Lear et al., 2000, Billups
and Schrag, 2003), while the record at ODP Site 1218 showed a slight increase in temperature
(Lear et al., 2004). One explanation for this relates to the concurrent deepening of the CCD. A
Δ[CO32−] effect is able to affect Mg/Ca records (Elderfield et al., 2006). By quantifying
Δ[CO32−], Lear et al. (2010) corrected the temperature records presented for DSDP Site 522
and ODP Site 1218 (Lear et al., 2004, Lear et al., 2000). After Δ[CO32−] correction, cooling
was shown across the EOT at these sites. Three methods have been proposed to correct for
Δ[CO32−] effect (1) the use of ostracod Mg/Ca because these appear unaffected by Δ[CO3
2−]
(Dwyer et al., 2002), (2) correction of benthic foraminiferal Mg/Ca records using B/Ca or
Li/Ca proxies (used to calculate Δ[CO32−]) (Yu and Elderfield, 2007, Lear and Rosenthal,
2006, Lear et al., 2010) and (3) use of infaunal benthic foraminiferal Mg/Ca ratios as these are
potentially buffered against any change in Δ[CO32−] (Elderfield et al., 2010). Poor proxy
resolution or complications regarding Mg/Ca paleothermometry at low temperatures may also
explain these results.
1.6.3. Proposed Mechanisms for EOT Initiation
The cause of the EOT is unknown. Theories include: declining PCO2atm
levels (Deconto and
Pollard, 2003), the opening of gateways around Antarctica enabling the formation of the
Antarctic Circumpolar Current (ACC), which subsequently thermally isolated the continent
(Toggweiler and Bjornsson, 2000) and increased upwelling of warm water, which promoted
the transport of moisture across the already cold continent, initiating ice expansion (Prentice
and Matthews, 1991). The establishment of the ACC at this time has also been linked to
changing nutrient concentrations in the ocean (Egan et al., 2013, Scher and Martin, 2006). If
the altered nutrient system increased carbon export to the sediments then this may explain the
decreasing PCO2atm proposed by Deconto and Pollard (2003). It has, however, been
postulated that the opening of gateways had a less significant impact on water mass
circulation change than has been reported previously. It was shown that ice sheet growth and
not the establishment of the ACC caused increased transport northward of Antarctic
intermediate water, stimulating the formation of Antarctic bottom water (Goldner et al.,
2014). This evidence supports declining PCO2 across the EOT as the driver for ice sheet
growth rather than the proposed circulation change created by the establishment of the ACC.

1. INTRODUCTION
30
Geochemical proxy data, from very well preserved foraminifera collected by the Tanzanian
Drilling Project, also supports the Deconto and Pollard (2003) model’s assumption that
declining PCO2atm
is central to the expansion of the Antarctic ice sheet (Pearson et al., 2009).
The reality is that initiation of EOT ice sheet expansion is poorly constrained and glaciation of
Antarctica could be a response to any of the proposed mechanisms.

1. INTRODUCTION
31
1.7. Regional and Geological Setting
The microfossil material analysed in this study was recovered from ODP Hole 757B, which
currently lies upon the Ninety East Ridge, in a water depth of 1644 m (17°01.458'S,
88°10.899'E) (Fig. 1.7a) (Peirce et al., 1989). Analysed core sections ranged between 13H-1
(5-7 cm) and 14H-cc (6-8 cm). During the EOT, the site was ~13°S of its current position
(Fig. 1.7b). Preservation of foraminifera was reported as being good to moderate by the
shipboard scientific party (Peirce et al., 1989). The shipboard party also reported a paleo
water depth of ~1500 m, for the samples under investigation in this study. This paleo water
depth is well above the Eocene and Oligocene paleo CCDs of 3400 m and 4400 m (Van
Andel, 1975).
Figure 1.7 (a) the current positions of ODP Holes 757B and 763A within the Indian Ocean
and (b) a paleo reconstruction of the same sites at 33.9 million years before present plotted
using the ODSN advanced plate reconstruction tool (Hay, 2000).
(a)
(b)

1. INTRODUCTION
32
1.8. Benthic Foraminiferal Biostratigraphy of ODP Hole 757B
From the Lower Eocene until the end of the Pleistocene, sediments comprise calcareous and
nannofossil oozes (Fig. 1.8a) (Peirce et al., 1989). The chronostratigraphy presented in figure
1.4 uses the timescale presented in (Berggren et al., 1985). Long ranging foraminiferal species
such as Gyroidinoides soldanii and Oridasalis umbonatus are found throughout the Eocene
and Oligiocene (Fig.1.8b). Foraminiferal assemblages within the Eocene are dominated by
Cibicidoides subspiratus, C. truncanus and Nuttallides truempyi; Oligocene assemblages are
dominated by Cibicidoides havanensis and Cibicidoides praemundulus. A multivariate
analysis, conducted by Nomura (1995), revealed a dominant foraminiferal assemblage across
the EOT (Fig. 1.8c). The assemblage consists of Cibicidoides havanensis, Cibicidoides
praemundulus, Oridasalis umbonatus and Bulimina jarvisi. The maximum abundance of the
Cibicidoides species, discovered within the assemblage, was 21% of total foraminifera and
was found in core sections 13H-1 (70-75 cm) (111.2 mbsf, 34.2 Ma) and 14H-1 (70-75 cm)
(120.8 mbsf, 35.5 Ma) (Nomura, 1995).
Figure 1.8 (a) lithology of ODP Hole 757B (b) frequency distribution of the significant
foraminiferal species across the EOT at ODP Site 757 and (c) the stratigraphic distribution of
the C. havanensis - C. praemundulus assemblage, indicating the developed interval of the
assemblage (dotted section)(percentage indicates the variance while the horizontal scale is
represented by factor loading). Adapted from (Nomura, 1995).
(a)

1. INTRODUCTION
33
1.9. Pilot Ostracod Study
A Cardiff Undergraduate Research Opportunities Programme (CUROP) funded project was
completed by the author in 2014. It investigated the Mg/Ca ratio in the deep-sea ostracod
genus Krithe at ODP Site 757. The aim of this project was to create an EOT Mg/Ca record
unaffected by Δ[CO32−]. Ostracods are a type of bi-valved crustacean predominantly formed
from the calcium carbonate (CaCO3) polymorph calcite (Fig. 1.9). They contain co-
precipitated magnesium similar to foraminifera. Due to ostracod’s infaunal habitat, it has been
proposed that they are relatively buffered against Δ[CO32−] (Dwyer et al., 2002).
Figure 1.9 Light micrographs of Krithe spp. recovered from ODP Hole 757B Core 14 Section
1 44-46 cm.
A Δ[CO32−] effect on calcitic Mg/Ca is expected across the EOT, at Site 757, because of the
predicted ~1km deepening of the CCD. Concurrent with the CCD deepening, foraminiferal
δ18O shifts at ODP site 757 (Holmström, 2014). The two isotope shifts observed at ODP Site
757 can also be seen in the isotope record of ODP Site 1218 (Fig. 1.10) (Coxall et al., 2005).
For the shifts shown at the two sites, discrepancies between the ages are due to differing age
model methodologies.
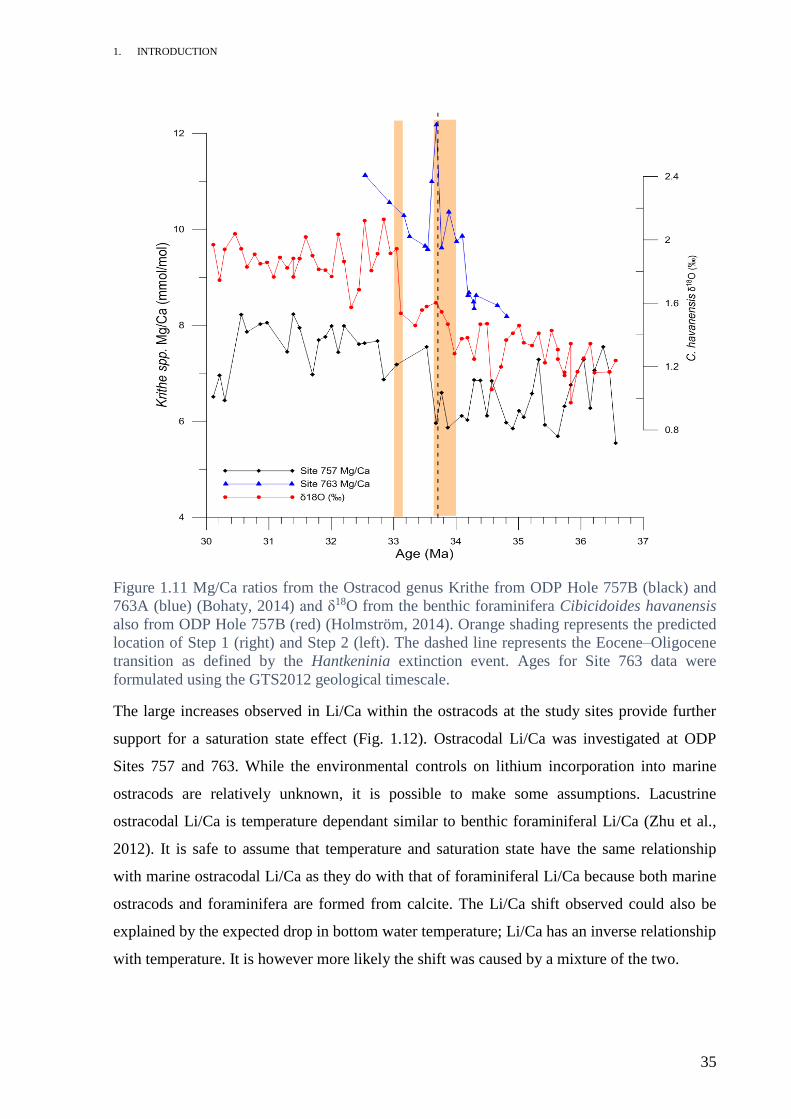
An increase in Krithe Mg/Ca was observed across the Eocene/Oligocene boundary at ODP
site 757 (Fig. 1.11). This increase was also observed in Krithe collected from ODP Site 763
(S. Bohaty, pers. comm., 2014). The offset in Mg/Ca between the two sites may be due to

1. INTRODUCTION
34
vital effects affecting different Krithe species or differing depths of the sites. The increase in
Mg/Ca may have been caused by: temperature increase across the EOT, a seawater Mg/Ca
change or a Δ[CO32−] effect on ostracods. A saturation state effect is contrary to the findings
of Dwyer et al. (2002).
It has been demonstrated that the assumption of no carbonate ion effect on ostracods is
incorrect at low temperatures (Elmore et al., 2012). The data from ODP Sites 757 and 763
supports the conclusions presented by Elmore et al. (2012).
Figure 1.10 the oxygen isotope records for ODP Sites 757 (red) and 1218 (blue). Site 757 data
was produced by Holmström (2014) and plotted against the ages calculated by Schedwin
(2014), which used the Geomagnetic Polarity Timescale presented in Wade et al. (2011). The
record from ODP Site 1218 was published by Coxall et al. (2005) and used an astronomical
timescale. The dashed line represents the EO boundary (33.7 Ma) as defined in Wade et al.
(2011). Two isotopic shifts are observed in both records, Step 1 and Step 2.
Step 2
Step 1
1. INTRODUCTION
35
Figure 1.11 Mg/Ca ratios from the Ostracod genus Krithe from ODP Hole 757B (black) and
763A (blue) (Bohaty, 2014) and δ18O from the benthic foraminifera Cibicidoides havanensis
also from ODP Hole 757B (red) (Holmström, 2014). Orange shading represents the predicted
location of Step 1 (right) and Step 2 (left). The dashed line represents the Eocene–Oligocene
transition as defined by the Hantkeninia extinction event. Ages for Site 763 data were
formulated using the GTS2012 geological timescale.
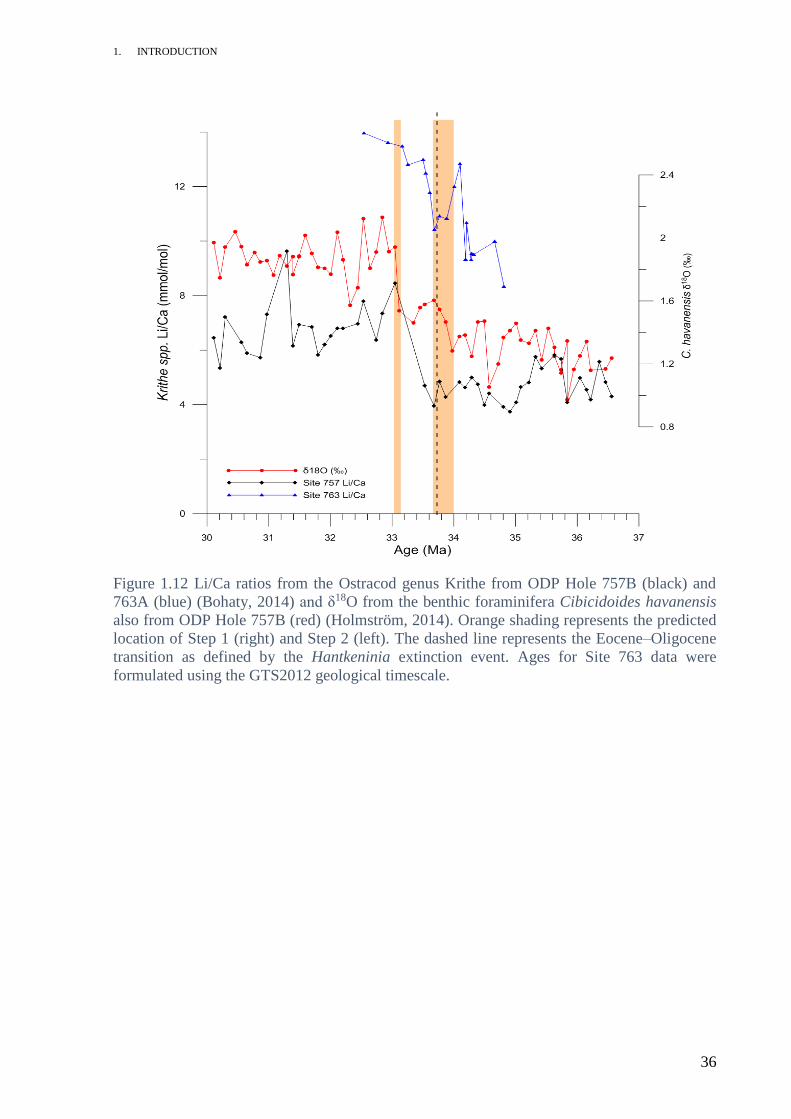
The large increases observed in Li/Ca within the ostracods at the study sites provide further
support for a saturation state effect (Fig. 1.12). Ostracodal Li/Ca was investigated at ODP
Sites 757 and 763. While the environmental controls on lithium incorporation into marine
ostracods are relatively unknown, it is possible to make some assumptions. Lacustrine
ostracodal Li/Ca is temperature dependant similar to benthic foraminiferal Li/Ca (Zhu et al.,
2012). It is safe to assume that temperature and saturation state have the same relationship
with marine ostracodal Li/Ca as they do with that of foraminiferal Li/Ca because both marine
ostracods and foraminifera are formed from calcite. The Li/Ca shift observed could also be
explained by the expected drop in bottom water temperature; Li/Ca has an inverse relationship
with temperature. It is however more likely the shift was caused by a mixture of the two.
1. INTRODUCTION
36
Figure 1.12 Li/Ca ratios from the Ostracod genus Krithe from ODP Hole 757B (black) and
763A (blue) (Bohaty, 2014) and δ18O from the benthic foraminifera Cibicidoides havanensis
also from ODP Hole 757B (red) (Holmström, 2014). Orange shading represents the predicted
location of Step 1 (right) and Step 2 (left). The dashed line represents the Eocene–Oligocene
transition as defined by the Hantkeninia extinction event. Ages for Site 763 data were
formulated using the GTS2012 geological timescale.

1. INTRODUCTION
37
1.10. Motivation
By unravelling the saturation state effect, it is hoped, the climate conditions that occurred
during the EOT can be determined. As discussed, ODP site 757 contains both infaunal and
epifaunal benthic foraminifera. The opportunity to analyse both at the same site is not
common. By comparing both infaunal and epifaunal foraminiferal chemical composition,
changes in water column chemistry can be quantified. The difference in Mg/Ca between
infaunal and epifaunal species may record the effect of saturation state change on the
foraminiferal Mg/Ca because infaunal species are relatively buffered against bottom water
Δ[CO32−] change (Elderfield et al., 2010). Hence, the difference in Mg/Ca can be used to
correct Mg/Ca for Δ[CO32−] change. If infaunal and epifaunal foraminifera records are the
same then Δ[CO32−] change is not responsible for heightened Mg/Ca records in the ostracod
records at ODP Sites 757 and 763. The Li/Ca and B/Ca ratios, from foraminifera at ODP site
757, can be used also to calculate saturation state change.
1.10.1. Hypothesis
A cooling effect is expected as Antarctica becomes glaciated. Consistently, benthic
foraminiferal records show an increase in Mg/Ca during this event and it has been proposed
that a carbonate saturation state effect may play a role (Lear et al., 2004, Elderfield et al.,
2006). It is hypothesised here that saturation state change is responsible for heightened
ostracodal and foraminiferal Mg/Ca ratios found across the Eocene/Oligocene boundary and
not increasing temperature or changing sea water Mg/Ca. A cooling effect associated with the
EOT glaciation of Antarctica rules out temperature increase, while the long residence times of
Mg2+ (~1 Ma) and Ca2+ (~10 Ma) in the ocean (Broecker et al., 1982) make changing seawater
Mg/Ca a less likely driver of rising foraminiferal Mg/Ca than saturation state change. The
consequence of proving this hypothesis is the ability to correct Mg/Ca records for Δ[CO32−]
effect and thus calculate accurate paleo-temperatures and ice volumes for the EOT.
It is predicted that the infaunal benthic Mg/Ca record collected by this study will be buffered
against saturation state change and therefore will not display an increase in Mg/Ca. It is
expected also that the epifaunal benthic B/Ca will show a marked increase across the EOT
while Li/Ca may also increase. Epifaunal benthic foraminiferal U/Ca is predicted to decrease.
Benthic foraminiferal test mass is anticipated to increase due to higher calcification rates

1. INTRODUCTION
38
associated with the deepening CCD. Infaunal test mass is predicted to increase less than
epifaunal mass across the EOT due to the buffering associated with the infaunal micro habitat.

2. MATERIALS AND METHODS
39
2. MATERIALS AND METHODS
Sampling
Microscopy
Weighing
Sample preparation and chemical cleaning
Trace metal analysis
Age model
Calculation of temperature, saturation state and ice volume

2. MATERIALS AND METHODS
40
2.1. Sampling
Cores were recovered in 1989 by the JOIDES Resolution deep sea drilling ship (operated by
the ODP) (Fig 2.1). The coarse fraction of the core collected from ODP Hole 757B was made
available from 110.55 to 129.54 metres below sea floor (mbsf). The coarse fraction contained
all material >63 µm diameter between these depths. The course fraction recovered from 63
intervals, each 2 cm of depth, was provided for this study. The intervals were spread
intermittently across the depth range. The coarse fraction was obtained by washing and
sieving the core in 15.0 MΩ deionized (DI) water (Fig. 2.1). Each sample was viewed under
two times magnification using a Nikon SM7645 light-microscope. All benthic foraminifera
tests were removed from the 250-500 µm size fraction (obtained through sieving the course
fraction) using a fine paintbrush and 15.0 MΩ DI water (Fig 2.1). Next, tests of dominant
benthic foraminifera species (Bulimina jarvisi, Cibicidoides havanensis, Gyroidinoides
soldani and Oridorsalis umbonatus) were removed. Tests with significant dissolution or
discolouring were disregarded. Picked tests with irregular morphological features (such as
size, shape or chamber composition) were disregarded also. Where this was not possible, a
record of any test abnormalities was made.
2.1.1. Species Selection
B. jarvisi and C. havanensis were chosen for trace metal analysis because of their dominance
within the core’s foraminiferal assemblage, as well as the need for an infaunal and epifaunal
species. B. jarvisi is also suitable as it is a relatively deep dwelling infaunal species and may
be more buffered against Δ[CO32−] than a shallower dwelling infaunal species. B. jarvisi tests
were picked from the 250-355 µm size fraction while C. havanensis tests were picked from
the 250-500 µm size fraction.
2.2. Microscopy
Light micrographs of the two species used for geochemical analysis, B. jarvisi and C.
havanensis, were taken. Species were cleaned of any obvious debris before reflected light
microscopy. Relatively well preserved specimens were chosen for this purpose. Images were
taking using a multifocus (montage) method on a Leica MZ16 stereo microscope using
Cardiff University Earth Microscope Computer 1.

2. MATERIALS AND METHODS
41
2.3. Weighing
The coarse fraction of each sample was transferred from a sample bottle to a metal tray. The
fraction was then weighed, in grams to two decimal places, using an A&D EK-400H balance.
Samples were then returned to sample bottles using a brush and glassine paper.
Picked B. jarvisi and C. havanensis tests from each sample were transferred from a
microscope slide to an aluminium foil container using a fine paintbrush. Samples were then
weighed, in micrograms, using a Mettler Toledo XP6/Z micro-balance. Samples were restored
to microscope slides using a brush and glassine paper. The foil container was weighed after
the transfer of tests to ensure all the foraminifera tests were recovered.
2.4. Sample Preparation and Chemical Cleaning
The chemical cleaning procedures used here borrow heavily from the methods presented by
Boyle (1981) and Boyle and Keigwin (1985). These methods were summarized in detail by
(Barker et al., 2003b). Samples must be prepared in batches of no more than ~30.
2.4.1. Crushing and Pre-Cleaning
The aim of crushing was to allow any foraminifera chamber fill to be released during the
cleaning process. Foraminiferal tests were placed in a single layer upon a moist, clean, glass
plate. A second clean glass plate was then used to open the foraminiferal chambers by gently
bringing the plates together (Fig 2.1). Large silicates and clays were removed from the
foraminifera fragments using a fine paintbrush and 18.2 MΩ DI water, viewed under a Nikon
SM7645 light-microscope. Subsequently, the test fragments were transferred to acid cleaned
500 µl micro-centrifuge tubes. The micro-centrifuge tube acid cleaning procedure is detailed
in Appendix 1.
2.4.2. Cleaning Procedure
Test fragments were cleaned chemically to remove any contaminants that could distort the
trace metal record. The cleaning procedure included: (1) removal of fine clays by
ultrasonication, both in 18.2 MΩ DI water and methanol, (2) reductive elimination of metal
oxides using a solution of hydrous hydrazine and citric acid in ammonia, (3) oxidation of
organic matter using a solution of hydrogen peroxide in sodium hydroxide and (4) an acid
leach step using Optima pure grade nitric acid. The full cleaning procedure is available in

2. MATERIALS AND METHODS
42
Appendix 2. Additionally, after the clay removal step, any obviously non-carbonate particles
were removed from samples viewed under a Nikon SM7645 light-microscope, 18.2 MΩ DI
water and a fine paintbrush. Reagents used were trace metal grade, unless otherwise stated.

2. MATERIALS AND METHODS
43
Figure 2.1 Sample recovery and analysis. Clockwise from top left: Drill Ship JOIDES
Resolution used to recover cores on ODP Leg 121 from (Crawford, 2013), sample washing
using a 63 µm sieve, glass slides and microscope used during the crushing procedure, Thermo
Finnegan XR high resolution-inductively coupled plasma-mass spectrometer used for trace
metal analysis, chemical cleaning and benthic foraminifera picking using a fine brush and
light microscope.

2. MATERIALS AND METHODS
44
2.5. Sample Dissolution and Calcium Concentration Analysis
One day in advance of trace metal analysis, each sample was dissolved in Optima grade nitric
acid. The full dissolution procedure is available in Appendix 3. Two aliquots, one 10 µl, the
other 100 µl, were removed from each solution and placed in new, acid cleaned tubes. The
smaller of the two aliquots was used to ascertain the calcium concentration ([Ca]) within the
samples while the larger was used to determine trace metal ratios.
Each sample was diluted with Optima grade nitric acid. The [Ca] of each sample was
analysed using a Thermo Element XR high resolution inductively coupled plasma mass
spectrometer (HR-ICP-MS). The [Ca] of each sample was quantified by comparing drift and
blank corrected intensity data (in counts per second of Ca43) to that of a standard containing
80 ppm calcium. Samples were loaded in blocks of five, separated by a blank and a standard
(24 mmol mixed calibration standard, MCS) (Fig. 2.2). A detailed methodology for calcium
concentration analysis is available in Appendix 3.
2.6. Trace Metal Analysis
On the day of trace metal analysis by the HR-ICP-MS, each sample was diluted using Optima
grade nitric acid. Samples were analysed using a Thermo Element XR HR-ICP-MS.
Independent consistency standards (CS1 and CS2) were analysed alongside the MCS at the
start and end of each run. This allowed the long term accuracy and precision of the run to be
quantified. Optima grade nitric acid blanks were analysed after every 2 samples. Matrix
matched standards (MMS) were created using Optima grade nitric acid diluted MCS. The
MMS provided a standard with similar [Ca] for each individual sample. Each MMS was run
directly after its corresponding sample and was used as a barometer for the count accuracy of
the mass spectrometer during the run (Fig. 2.2). This method accounts for the normal
variation in matrix effects observed better than a single matrix effect correction (Lear et al.,
2002). Full elemental analysis included measuring intensities of 6Li, 7Li, 11B, 24Mg, 25Mg,
27Al, 43Ca, 46Ca, 48Ca, 47Ti, 55Mn, 87Sr, 88Sr, 111Cd, 138Ba, 146Nd and 238U. Each
isotope count was blank corrected using the previous blank in the run sequence. A blank
corrected intensity ratio was calculated using the Ca43 intensity and the blank corrected count
for each isotope. An elemental ratio was subsequently formulated using the MMS and the
blank corrected intensity ratios.

2. MATERIALS AND METHODS
45
Figure 2.2 A representative schematic of a normal analysis sequence used to determine: [Ca]
in dissolved foraminifera (left) and trace metal/Ca ratios in dissolved foraminifera (right).
MCS is the Cardiff University mixed calibration standard, CS1 and CS2 are consistency
standards used to quantify long term precision of the mass spectrometry equipment while
MMS represents the matrix matched standards used for calculating trace metal concentration
ratios.

2. MATERIALS AND METHODS
46
2.7. Age Model
The age model (Fig. 2.3) which was formulated by Schedwin (2014) using the planktonic
foraminiferal and nannofossil biostratigraphy recorded by the shipboard scientific party of
ODP Leg 121 was applied to the samples analysed in this study (Peirce et al., 1989). The
model utilises the revised geomagnetic polarity time scale (GPTS) presented in Wade et al.
(2011). By measuring the midpoint depth (mbsf) of the samples that correlated with the
nannofossil and planktonic foraminiferal datums, a calibration equation was created allowing
for interpolation within the spread of datum events. A sedimentation rate of 2.9 m/Ma was
predicted for site 757B using the model (Schedwin, 2014).
Figure 2.3 The age model constructed, for ODP Site 757, using nannofossil and planktonic
foraminifera datum events adapted from (Schedwin, 2014). The abbreviation LO represents
the lowest occurrence found of the specific foraminifera. A linear fit was applied to all the
datums. The success of this regression was measured with the multivariate R2, which was
0.98.
y = 0.3452x - 8.157
R² = 0.9873
27
29
31
33
35
37
39
105 110 115 120 125 130 135
Age
(Ma)
Depth (mbsf)
LO Hantkeninia spp.
LO Globerigeinatheka index
Nannofossil datums
Linear (all datums)

2. MATERIALS AND METHODS
47
2.8. Calculation of Temperature, Saturation State and Ice Volume
Changes in temperature, saturation state and ice volume can be calculated from raw trace
metal data using a number of calibrations. Listed here are those calibrations that were applied
to the data collected throughout this study. Each calibration was chosen to best match the
species and data under investigation.
Bottom water temperature (T) was calculated from the Mg/Ca of B. jarvisi through the
following equation:
EQN. (2.1)
𝑀𝑔/𝐶𝑎𝐵. 𝑗𝑎𝑟𝑣𝑖𝑠𝑖 = 1.008 exp (0.114 × 𝑇)
(Lear et al., 2002)
The calibration presented in equation 2.1 was derived for the infaunal foraminifera species O.
umbonatus. O. umbonatus have a higher concentration of Mg/Ca than Cibicidoides spp.
Uvigerina spp. is a second infaunal genera for which a temperature calibration exists.
Uvigerina have lower concentrations of Mg/Ca than Cibicidoides (Lear et al., 2002).
Cibicidoides havanensis was shown, by this study, to have a significantly lower Mg/Ca
concentration than B. jarvisi. Therefore, a calibration using the infaunal species O. umbonatus
was chosen because the calibration better fits the data collected.
A first order approximation of early Cenozoic (~49 Ma) Mg/Ca in seawater is 3.5 mol mol-1
while modern concentrations of Mg/Ca in the oceans was measured at approximately 5.2 mol
mol-1. This was combined into equation 2.1 so as to account for changes in oceanic Mg/Ca
from the Eocene to present (Equation 2.2).
EQN. (2.2)
𝑀𝑔/𝐶𝑎𝐵. 𝑗𝑎𝑟𝑣𝑖𝑠𝑖 =3.5
5.2 × 1.008 exp (0.114 × 𝑇)
(Lear et al., 2002)
Saturation state was calculated from the B/Ca of C. havanensis through the following
equation:

2. MATERIALS AND METHODS
48
EQN. (2.3)
𝐵/𝐶𝑎𝐶. ℎ𝑎𝑣𝑎𝑛𝑒𝑛𝑠𝑖𝑠 = 0.69 × ∆[𝐶𝑂3 2−] + 119.1
(Yu and Elderfield, 2007)
The calibration presented in equation 2.3 was derived for the epifaunal foraminifera species
C. mundulus. C. mundulus are genetically the closest relative to C. havanensis for which a
B/Ca saturation state calibration exists (Holbourn et al., 2013). For this reason, the C.
mundulus saturation state calibration was chosen. The range of values used to formulate the
C. mundulus saturation state calibration were higher than those recorded for C. havanensis
(Yu and Elderfield, 2007). Therefore, it is likely that a true calculation of saturation state
change would require lower sensitivities than those used in equation 2.3.
Bottom water temperature (T) was calculated using the following equation:
EQN. (2.4)
𝑀𝑔/𝐶𝑎𝐶. ℎ𝑎𝑣𝑎𝑛𝑒𝑛𝑠𝑖𝑠 = 0.82 + 0.056𝑇 + 0.087∆[𝐶𝑂3 2−]
(Elderfield et al., 2006)
The calibration presented in equation 2.4 was derived for the epifaunal foraminifera species
C. wuellerstorfi. Due to a lack of more suitable species specific calibrations, it was necessary
to use C. wuellerstorfi. It is highly likely that, due to vital effects, sensitivities to saturation
state change and temperature differ from C. havanensis.
Due to the lack of species specific calibrations for B. jarvisi and C. havanensis, it must be
noted that the above calibrations are best employed to monitor relative changes in bottom
water temperature and saturation state. By setting the value of the first sample to zero, relative
changes can be quantified. Due to its use in calculating relative change, the paired Mg/Ca and
saturation state equation applied to the data collected from C. havanensis omitted a past ocean
Mg/Ca component. It should be noted that if this were included, as with equation 2.2, absolute
temperatures would be notably higher.

2. MATERIALS AND METHODS
49
The output from equation 2.4 was used to calculate the δ18O of seawater using the benthic
equation of Shackleton (1974):
EQN (2.5)
𝑇 = 16.9 − 4 (δ18𝑂𝐹𝑂𝑅𝐴𝑀 − δ18𝑂𝑆𝐸𝐴𝑊𝐴𝑇𝐸𝑅)
to which a +0.5‰ equilibrium offset for Cibicidoides was applied.

3. RESULTS
50
3. RESULTS
Microscopy
Foraminiferal average test masses
Species abundances
Foraminiferal Mg/Ca
Foraminiferal Li/Ca
Foraminiferal B/Ca
Foraminiferal Sr/Ca
Foraminiferal U/Ca

3. RESULTS
51
3.1. Microscopy
C. havanensis test form was found to be trochospiral. Specimens had a distinctive convex
spiral side and a less convex umbilical side. The majority of specimens had a thick plug of
secondary calcite in the centre of the umbilical side. Chambers, which increase gradually in
size, were separated by flush sutures on the umbilical side and deeper but thinner sutures on
the spiral side. Specimens were generally well perforated, especially on the spiral suture. (Fig.
3.1).
B. jarvisi test form was found to be a slender, triserial series. The initial portion of the test
was acute, while the apertural end was sub rounded in the majority of specimens. Chambers
increased in size towards the aperture and were separated by depressed sutures. Chamber
walls showed course perforations. An adapical spine is associated with this species and was
observed in most specimens. Each specimen had approximately 6 whorls. (Fig. 3.1).
Figure 3.1 Reflected light micrograph images of the benthic foraminifera species Cibicidoides
havanensis (top) and Bulimina jarvisi (bottom). Images on top row are a single species from
ODP Site 757 Hole B Section 13H Core 4 135-137 cm. (left hand top image: spiral view,
right hand top image: umbilical view). Bottom row images are two specimens from ODP Site
757 Hole B Section 13H Core 2 103-105 cm

3. RESULTS
52
3.2. Foraminiferal Average Test Masses
The epifaunal foraminiferal test mass, recovered from C. havanensis, showed variation
between 30 and 37 million years before present. The dataset included 62 samples and returned
a mean mass of 84.85 µg (Fig. 3.2a). Mass values ranged from 49.83 to 144.83 µg. The mean
mass of epifaunal tests before the initiation of the EOT was 63.49 µg, while during the EOT
the epifaunal test mass rose to 88.64 µg. After the EOT, the mean epifaunal test mass
increased further to 102.23 µg. Post EOT, epifaunal test mass was significantly higher
compared to pre EOT concentrations. The relative standard deviation (RSD) before the EOT
was 16% (n=25) while after the EOT RSD fell to 15% (n=29). During the EOT the RSD of
the data was significantly higher at 27% (n=8).
The infaunal foraminiferal test mass, recovered from B. jarvisi, showed some variation
between 30 and 37 million years before present. The dataset included 59 samples and returned
a mean mass of 34.54 µg (Fig. 3.2b). Values ranged from 26 to 41.57 µg. The mean mass of
infaunal tests before the initiation of the EOT was 32.57 µg, while during the EOT the mean
infaunal test mass dropped to 31.65 µg. After the EOT, the mean infaunal test mass increased
to 36.95 µg. Post EOT, mean infaunal test mass was higher compared to pre EOT
concentrations. The relative standard deviation (RSD) before the EOT was 10% (n=25) while
after the EOT RSD had fallen to 6% (n=27). During the EOT the RSD was 8% (n=7).

3. RESULTS
53
Figure 3.2 Average mass of the benthic foraminifera species (a) Cibicidoides havanensis and
(b) B. jarvisi at ODP Site 757 Hole B. Dashed line represents the EO boundary as defined by
Wade et al. (2011). Ages use the Wade et al. (2011) timescale. Solid black lines and grey
boxes represent the arithmetic mean and relative standard deviation respectively. The data set
is split into three before (blue), during (red) and after (black) the Eocene-Oligocene transition.
Vertical orange bars represent Steps 1 and 2 of the EOT, as shown by the δ18O record of
Holmström (2014).
(a)
(b)

3. RESULTS
54
3.3. Species Abundances
The abundance of C. havanensis showed variation between 30 and 37 million years before
present. The dataset included 63 samples and returned a mean abundance of 12.73 specimens
per gram of coarse fraction (Fig. 3.3). Abundance values ranged from 1.02 to 53.85 specimens
per gram of coarse fraction. The mean abundance of C. havanensis before the initiation of the
EOT was 16.28 specimens per gram, while during the EOT the mean abundance of C.
havanensis fell to 14.64. After the EOT, the mean abundance of C. havanensis decreased
further to 9.25 specimens per gram. Post EOT, abundance of C. havanensis was significantly
lower compared to pre EOT concentrations. During Step 1, abundance of C. havanensis
dropped by 13.63 specimens per gram, while during Step 2 C. havanensis abundance
increased by 5.42 specimens per gram.
The abundance of B. jarvisi showed variation between 30 and 37 million years before present.
The dataset included 59 samples and returned a mean abundance of 18.55 specimens per gram
of coarse fraction (Fig. 3.3). Abundance values ranged from 4.08 to 53.85 specimens per gram
coarse fraction. The mean abundance of B. jarvisi before the initiation of the EOT was 22.64
specimens per gram, while during the EOT the mean abundance of B. jarvisi rose to 22.81.
After the EOT, the mean abundance of B. jarvisi decreased to 14.29 specimens per gram. Post
EOT, abundance of B. jarvisi was significantly lower compared to pre EOT concentrations.
During Step 1 abundance of B. jarvisi drops by 34 specimens per gram, while during Step 2
B. jarvisi abundance increases by 5.17 specimens per gram.

3. RESULTS
55
Figure 3.3 Number of the benthic foraminifera species Cibicidoides havanensis (purple) and B. jarvisi (red) per gram of course fraction at ODP Site
757 Hole B. Vertical yellow bars represent Steps 1 and 2 of the EOT, as shown by the δ18O record of Holmström (2014). Ages use the Wade et al.
(2011) timescale.
0
10
20
30
40
50
60
29 30 31 32 33 34 35 36 37
Fora
min
ife
ra p
er
Gra
m o
f C
oar
se F
ract
ion
Age (Ma)
B. jarvisi C. havanensis

3. RESULTS
56
3.4. Foraminiferal Mg/Ca
The epifaunal magnesium calcium record, recovered from C. havanensis, showed variation
between 30 and 37 million years before present. The dataset included 62 samples and returned
a mean Mg/Ca ratio of 1.1 mmol/mol (Fig. 3.4). Mg/Ca values ranged from 0.92 to 1.29
mmol/mol. The mean value of epifaunal foraminiferal Mg/Ca was 1.07 mmol/mol before the
initiation of the EOT, while during the EOT the mean Mg/Ca concentration rose to 1.1
mmol/mol. After the EOT the mean Mg/Ca remained relatively elevated at 1.13 mmol/mol.
Post EOT Mg/Ca concentrations were slightly raised compared with pre EOT concentrations.
During Step 1 of the EOT epifaunal Mg/Ca increased by 0.14 mmol/mol from 1.14
mmol/mol. While the trend across Step 1 was positive, the data peaked and troughed during
this period. Increases during Step 1 were generally two times the magnitude of decreases.
Across Step 2, an increase of 0.1 mmol/mol Mg/Ca was observed, rising to 1.18 mmol/mol.
In between the two steps, a marked decrease in Mg/Ca occurred falling by 0.08 mmol/mol to
1.08 mmol/mol before rising to 1.17 mmol/mol. This decrease took place directly after 33.7
Ma, the EO boundary. After the EOT, Mg/Ca continued to increase, peaking at 1.29
mmol/mol (32.3 Ma), before decreasing to 0.99 mmol/mol between 32.3 and 31.5 Ma.
The infaunal Mg/Ca record, recovered from B. jarvisi, also showed significant variation
between 30 and 37 million years before present. The dataset included 56 samples and returned
a mean Mg/Ca ratio of 1.78 mmol/mol (Fig. 3.4). Mg/Ca values ranged from 1.5 to 2.13
mmol/mol. The mean value of infaunal foraminiferal Mg/Ca was 1.73 mmol/mol before the
initiation of the EOT, while during the EOT the mean Mg/Ca concentration rose to 1.8
mmol/mol. After the EOT the mean Mg/Ca increased further to 1.85 mmol/mol. Post EOT
Mg/Ca levels were raised compared with pre EOT levels. During Step 1 of the EOT, infaunal
Mg/Ca increased by 0.07 mmol/mol to 1.77 mmol/mol. While the trend across Step 1 was
positive, the data peaked and troughed during this period. Between Steps 1 and 2 Bulimina
jarvisi disappeared from the fossil record and therefore Mg/Ca concentrations were not
recorded. This disappearance takes place directly after 33.7 Ma, the EO boundary. The
species does not reappear until the commencement of Step 2. An increase of 0.12 mmol/mol
occurred between Steps 1 and 2. Across Step 2, infaunal Mg/Ca increased to 2.13 mmol/mol
from 1.89 mmol/mol. After the EOT (from 33 Ma to 30Ma) Mg/Ca decreased steadily
(Range: 2.13 mmol/mol to 1.5 mmol/mol).

3. RESULTS
57
Figure 3.4 Record of benthic foraminiferal Mg/Ca collected from the species Cibicidoides havanensis (black) and B. jarvisi (red) at ODP Site 757 Hole
B. Vertical orange bars represent Steps 1 and 2 of the EOT, as shown by the δ18O record of Holmström (2014). Dashed line represents the EO
boundary as defined by Wade et al. (2011). Ages use the Wade et al. (2011) timescale.

3. RESULTS
58
3.5. Foraminiferal Li/Ca
The epifaunal lithium calcium record, recovered from C. havanensis, showed variation
between 30 and 37 million years before present. The dataset included 62 samples and returned
a mean Li/Ca ratio of 8.29 mmol/mol (Fig. 3.5). Li/Ca values ranged from 7.4 to 10.1
mmol/mol. The mean value of epifaunal foraminiferal Li/Ca was 8 mmol/mol before the
initiation of the EOT, while during the EOT the mean Li/Ca concentration rose to 8.32
mmol/mol. After the EOT, the mean Li/Ca remained relatively elevated at 8.56 mmol/mol.
Post EOT Li/Ca levels were raised compared with pre EOT levels. During Step 1 of the
Eocene Oligocene transition, epifaunal Li/Ca decreased by 0.12 mmol/mol to 7.6 mmol/mol.
In between the two steps, a marked increase in Li/Ca occurred, rising 1.57 by mmol/mol to
9.16 mmol/mol. This increase took place directly after 33.7 Ma, the EO boundary. Across
Step 2, Li/Ca decreased by 0.24 mmol/mol, however this was followed immediately by an
increase of 0.24 mmol/mol. After the EOT, Li/Ca decreased to 7.98 mmol/mol, a drop of 1.18
mmol/mol (32.3 Ma), before levelling off between 32.3 and 31.07 Ma. A strongly positive
trend in Li/Ca values was observed from 31 Ma onward. Outliers, well outside the general
trend of the data, were observed at 30.1, 30.46 and 36.05 Ma.
The infaunal lithium calcium record, recovered from B. jarvisi, also showed significant
variation between 30 and 37 million years before present. The dataset includes 53 samples
and returned a mean Li/Ca ratio of 7.4 mmol/mol (Fig. 3.5). Li /Ca values ranged from 6.22 to
9.04 mmol/mol. The mean value of infaunal foraminiferal Li/Ca was 6.84 mmol/mol before
the initiation of the EOT, while during the EOT mean Li/Ca concentration rose to 7.36
mmol/mol. After the EOT, the mean Li/Ca increased further to 7.94 mmol/mol. Post EOT,
Li/Ca levels were notably raised compared with pre EOT levels. During Step 1 of the Eocene
Oligocene transition, infaunal Li/Ca increased from 6.75mmol/mol by 0.34 mmol/mol.
Between Steps 1 and 2, Bulimina jarvisi disappeared from the fossil record and therefore
Li/Ca concentration was not recorded. This disappearance took place directly after 33.7 Ma,
the EO boundary. The species does not reappear until the commencement of Step 2. Upon the
re-emergence of the species, Li/Ca concentration was significantly raised by 0.77 mmol/mol.
After the EOT, Li/Ca remains relatively constant with the exception of two peaks at 32 Ma
and 31.7 Ma.

3. RESULTS
59
Figure 3.5 Record of benthic foraminiferal Li/Ca collected from the species Cibicidoides havanensis (black) and B. jarvisi (red) at ODP Site 757 Hole
B. Vertical orange bars represent Steps 1 and 2 of the EOT, as shown by the δ18O record of Holmström (2014). Dashed line represents the EO
boundary as defined by Wade et al. (2011). Ages use the Wade et al. (2011) timescale.

3. RESULTS
60
3.6. Foraminiferal B/Ca
The epifaunal boron calcium record, recovered from C. havanensis, showed variation between
30 and 37 million years before present. The dataset included 62 samples and returned a mean
B/Ca ratio of 103.75 mmol/mol (Fig. 3.6). B/Ca values ranged from 86.94 to 129.57
mmol/mol. The mean value of epifaunal foraminiferal B/Ca was 97.59 mmol/mol before the
initiation of the EOT, while during the EOT the mean B/Ca concentration rose to 105.73
mmol/mol. After the EOT, the mean B/Ca remained elevated at 108.73 mmol/mol. Post EOT
B/Ca levels were raised compared with pre EOT levels. During Step 1 of the Eocene
Oligocene transition, epifaunal B/Ca decreased by 0.77 mmol/mol to 97.83 mmol/mol. In
between the two steps, a marked increase in B/Ca occurred rising to 116.41 mmol/mol before
falling by 6.79 mmol/mol to 109.62 mmol/mol. By the beginning of Step 2, epifaunal B/Ca
had increased by 8.83 mmol/mol. Across Step 2, B/Ca decreases by 7.31 mmol/mol followed
immediately by an increase of 4.32 mmol/mol. After the EOT, B/Ca generally decreased,
before increasing again after 31 Ma. A strongly positive trend in B/Ca values is observed
from 31 Ma onward. Two outliers were recorded after 30.65 Ma that were significantly lower
than the general trend of the data, while one outlier before the EOT (35.06 Ma) was
significantly higher than the surrounding concentrations.
The infaunal boron calcium record, recovered from B. jarvisi, also showed significant
variation between 30 and 37 million years before present. The dataset included 50 samples
and returned a mean B/Ca ratio of 8.01 mmol/mol (Fig. 3.6). B/Ca values ranged from 6.28 to
10.84 mmol/mol. The mean value of infaunal foraminiferal B/Ca was 7.36 mmol/mol before
the initiation of the EOT, while during the EOT mean B/Ca concentration rose to 7.39
mmol/mol. After the EOT, the mean B/Ca increased further to 8.79 mmol/mol. Post EOT
B/Ca levels were notably raised compared with pre EOT levels. During Step 1 of the Eocene
Oligocene transition, infaunal B/Ca increased from 7.05 mmol/mol by 0.13 mmol/mol.
Between Steps 1 and 2, Bulimina jarvisi disappeared from the fossil record and therefore
B/Ca concentration was not recorded. This disappearance took place directly after 33.7 Ma,
the EO boundary. The species does not reappear until Step 2. Upon the re-emergence of the
species, B/Ca concentration was significantly raised by 0.48 mmol/mol. After the EOT, B/Ca
continued to increase steadily to a high of 10.84 mmol/mol (30.29 Ma).

3. RESULTS
61
Figure 3.6 Record of benthic foraminiferal B/Ca collected from the species Cibicidoides havanensis (black) and B. jarvisi (red) at ODP Site 757 Hole
B. Vertical orange bars represent Steps 1 and 2 of the EOT, as shown by the δ18O record of Holmström (2014). Dashed line represents the EO
boundary as defined by Wade et al. (2011). Ages use the Wade et al. (2011) timescale.

3. RESULTS
62
3.7. Foraminiferal Sr/Ca
The epifaunal strontium calcium record, recovered from C. havanensis, showed some
variation between 30 and 37 million years before present. The dataset included 62 samples
and returned a mean Sr/Ca ratio of 0.85 mmol/mol (Fig.3.7). Sr/Ca values ranged from 0.76 to
0.94 mmol/mol. The mean value of epifaunal foraminiferal Sr/Ca was 0.85 mmol/mol before
the initiation of the EOT, while during the EOT the mean Sr/Ca concentration rose to 0.87
mmol/mol. After the EOT, the mean Sr/Ca decreased to 0.86 mmol/mol. Post EOT, Sr/Ca
concentrations were similar to pre EOT concentrations. During Step 1 of the Eocene
Oligocene transition, epifaunal Sr/Ca decreased from 0.87 mmol/mol by 0.08 mmol/mol. In
between the two steps, a marked increase in Sr/Ca occurred between 33.7 and 33.5 Ma rising
by 0.13 mmol/mol to 0.92 mmol/mol before falling by 0.02 mmol/mol. By the beginning of
Step 2, epifaunal Sr/Ca had increased to 0.94 mmol/mol. Across Step 2, Sr/Ca decreases by
0.06 mmol/mol. After the EOT, Sr/Ca remained generally stable at pre EOT concentrations.
An outlier was recorded at 30.06 Ma that was significantly lower than the general trend of the
data.
The infaunal strontium calcium record, recovered from B. jarvisi, also showed some variation
between 30 and 37 million years before present. The dataset included 56 samples and returned
a mean Sr/Ca ratio of 0.52 mmol/mol (Fig. 3.7). Sr/Ca values ranged from 0.45 to 0.62
mmol/mol. The mean value of infaunal foraminiferal Sr/Ca was 0.5 mmol/mol before the
initiation of the EOT, while during the EOT mean Sr/Ca concentration rose to 0.51
mmol/mol. After the EOT, the mean Sr/Ca increased further to 0.54 mmol/mol. Post EOT
Sr/Ca levels were slightly raised compared with pre EOT levels. During Step 1 of the Eocene
Oligocene transition, infaunal Sr/Ca decreased by 0.07 mmol/mol to 0.46 mmol/mol. The
negative trend began at 33.8 Ma, slightly after the commencement of Step 1. In between
Steps 1 and 2, Bulimina jarvisi disappeared from the fossil record and therefore Sr/Ca
concentration was not recorded. This disappearance took place directly after 33.7 Ma, the EO
boundary. The species does not reappear until the commencement of Step 2. Upon the re-
emergence of the species, Sr/Ca concentration was raised by 0.06 mmol/mol. After the EOT,
Sr/Ca concentration remained relatively constant, with a large peak between 30 and 31 Ma.

3. RESULTS
63
Figure 3.7 Record of benthic foraminiferal Sr/Ca collected from the species Cibicidoides havanensis (black) and B. jarvisi (red) at ODP Site 757 Hole
B. Vertical orange bars represent Steps 1 and 2 of the EOT, as shown by the δ18O record of Holmström (2014). Dashed line represents the EO
boundary as defined by Wade et al. (2011). Ages use the Wade et al. (2011) timescale.

3. RESULTS
64
3.8. Foraminiferal U/Ca
The epifaunal uranium calcium record, recovered from C. havanensis, showed variation
between 30 and 37 million years before present. The dataset included 62 samples and returned
a mean U/Ca ratio of 4.3 mmol/mol (Fig. 3.8). U/Ca values ranged from 2.63 to 7.73
mmol/mol. The mean value of epifaunal foraminiferal U/Ca was 3.4 mmol/mol before the
initiation of the EOT, while during the EOT the mean U/Ca concentration rose to 3.77
mmol/mol. After the EOT, the mean U/Ca increased to 5.24 mmol/mol. Post EOT U/Ca
concentrations were higher compared to pre EOT concentrations. During Step 1 of the Eocene
Oligocene transition, epifaunal U/Ca increased from 2.95 mmol/mol to 3.52 mmol/mol, a rise
of 0.57 mmol/mol. In between the two steps, an increase in U/Ca concentration, from 3.52
mmol/mol to 4.43 mmol/mol, occurred. Across Step 2, U/Ca increased by 0.66 mmol/mol
rising to 4.69 mmol/mol. Pre EOT concentrations of U/Ca were stable. After the EOT, U/Ca
continued to increase with a positive trend. By 30.21 Ma, epifaunal U/Ca concentration had
risen as high as 7.73 mmol/mol. An outlier was recorded at 30.06 Ma that was significantly
lower than the general trend of the data.
The infaunal uranium calcium record, recovered from B. jarvisi, also showed variation
between 30 and 37 million years before present. The dataset included 56 samples and returned
a mean U/Ca ratio of 4.54 mmol/mol (Fig 3.8). U/Ca values ranged from 3.17 to 6.15
mmol/mol. The mean value of infaunal foraminiferal U/Ca was 4.06 mmol/mol before the
initiation of the EOT, while during the EOT mean U/Ca concentration rose to 4.45 mmol/mol.
After the EOT, the mean U/Ca increased further to 5.01 mmol/mol. Post EOT, U/Ca levels
were raised compared with pre EOT levels. Upon commencement of Step 1 of the Eocene
Oligocene transition, infaunal U/Ca decreased by 1.11 mmol/mol to 3.47 mmol/mol before
rising by 1.06 mmol/mol. In between Steps 1 and 2, Bulimina jarvisi disappeared from the
fossil record and therefore U/Ca concentration was not recorded. This disappearance took
place directly after 33.7 Ma, the EO boundary. The species does not reappear until the
initiation of Step 2. Upon the re-emergence of the species, U/Ca concentration was raised by
0.63 mmol/mol. After the EOT, U/Ca concentrations were stable until 31.7 Ma. At this point,
U/Ca shifted upward by 1.65 mmol/mol before decreasing slightly to 5.42 mmol/mol. The
infaunal U/Ca signal remains stable from this point Ma forward.

3. RESULTS
65
Figure 3.8 Record of benthic foraminiferal U/Ca collected from the species Cibicidoides havanensis (black) and B. jarvisi (red) at ODP Site 757 Hole
B. Vertical orange bars represent Steps 1 and 2 of the EOT, as shown by the δ18O record of Holmström (2014). Dashed line represents the EO
boundary as defined by Wade et al. (2011). Ages use on the Wade et al. (2011) timescale.

4. DISCUSSION
66
4. DISCUSSION
Across the EOT at ODP site 757:
Bottom water temperature change
Saturation state change
Ice volume and sea level changes
Unravelling the foraminiferal Li/Ca, Sr/Ca and U/Ca records
Productivity changes

4. DISCUSSION
*Bottom water temperatures were calculated from Krithe Mg/Ca using the calibration of
Dwyer et al. (2002).
67
4.1. Bottom Water Temperature and Saturation State History at ODP Site 757
4.1.1. Bottom Water Temperature Calculated Using Mg/Ca of B. jarvisi
Bottom water temperature was calculated from B. jarvisi Mg/Ca without correction for
∆[CO32-] because it was assumed that B. jarvisi Mg/Ca was buffered against ∆[CO3
2-]
(Elderfield et al., 2010). This produced a temperature change of +0.87 ⁰C between the
beginning and the end of the EOT (Fig. 4.1). At Tanzanian Drilling Project (TDP) Sites 12
and 17, a -1.2 ⁰C change was calibrated for the same period (Fig. 4.1) (Lear et al., 2008).
Across Step 1, which has been strongly linked with a cooling event, the B. jarvisi bottom
water temperatures marginally increased by 0.16 ⁰C. This is in stark contrast with the -2.8 ⁰C
formulated for Step 1 at TDP Sites 12 and 17 (Lear et al., 2008). Dissolution has been shown
to preferentially remove magnesium rich calcite (Brown and Elderfield, 1996). Therefore, it is
possible that some of apparent warming across Step 1 could be attributed to reduced
dissolution rather than temperature change. Step 2 bottom water temperatures from ODP Site
757 are somewhat easier to correlate with the TDP sites than those for Step 1. From the
beginning to the end of Step 2, a +0.45 ⁰C was formulated for ODP Site 757, a change
comparable to the ~0.5 ⁰C calculated by Lear et al. (2008) for TDP Sites 12 and 17.
The pilot investigation conducted by the author for this study, found that Mg/Ca bottom water
temperatures, formulated using the ostracod genus Krithe*, changed by -0.13 ⁰C at Step 1 and
+0.68 ⁰C at Step 2 (Fig. 4.1). Again the temperature change at Step 2 appears to be
comparable with published records, however the decrease at Step 1 is approximately three
times smaller than the one presented in Lear et al. (2008). As with infaunal foraminifera, such
as B. jarvisi, ostracods have been predicted to be relatively buffered against saturation state
change (Dwyer et al., 2002).
If these species’ Mg/Ca concentrations were buffered against saturation state change, the
temperatures produced would correlate with the TDP sites, which were very well preserved
and located well above the CCD. Across Step 1, a significant cooling is expected, due to the
magnitude of the upward shift in foraminiferal δ18O and the lack of ice growth stimulus (Lear
et al., 2008). Some cooling is required to explain the size of the shift in foraminiferal δ18O
across Step 2. The size and/or lack of cooling predicted by these Mg/Ca paleotemperatures
across Steps 1 and 2 is incompatible with other geochemical proxy information published for

4. DISCUSSION
68
the EOT. It is suggested here that, contrary to Elderfield et al. (2010) and Dwyer et al. (1995)
respectively, both B. jarvisi and Krithe spp. Mg/Ca are affected by changes in saturation state.
A reliable way of quantifying changes in saturation state has not been published for either of
these species. Therefore, their use as indicators of past climate is restricted to sites well above
the calcite compensation depth or periods for which carbonate chemistry in the ocean
remained the same.

4. DISCUSSION
69
Figure 4.1 Record of change in bottom water temperature at ODP Site 757B relative to the
oldest sample, for the benthic foraminifera B. jarvisi (red) and the ostracod species Krithe spp.
(blue). Ages use on the Wade et al. (2011) timescale. Vertical orange bars represent Steps 1
and 2 of the EOT, as shown by the δ18O record of Holmström (2014). Dashed line represents
the EO boundary as defined by Wade et al. (2011). Black line shows the absolute bottom
water temperatures calculated for TDP Sites 12 and 17 from Cibicidoides spp. (Lear et al.,
2008) plotted against the timescale of Cande and Kent (1995). Vertical green bars represent
Step 1 and 2 of the EOT, as shown by the δ18O record of Coxall et al. (2005).
Age (Ma), Cande and Kent (1995) time scale
C
ha
nge
in
Bo
tto
m W
ate
r T
em
pe
ratu
re (
⁰C)

4. DISCUSSION
70
4.1.2. Saturation State Change Calculated Using B/Ca of C. havanensis
As the use of B. jarvisi Mg/Ca to calculate paleotemperatures was unsuccessful, it became the
necessary to calculate saturation state change in order to separate the temperature signal from
the signal caused by saturation state change. Through the use of C. havanensis B/Ca,
saturation state change was quantified. Across the EOT, a clear increase in saturation state
occurred, rising by 18.17 µmol kg-1 (Fig.4.2). This is to be expected when considering the ~1
km deepening of the CCD across the same period. The decrease of 23.88 µmol kg-1 that starts
at 34.18 Ma, marginally before the initiation of Step 1, and lasts 400 Ka is less expected. A
dissolution hypothesis explaining why temperatures appear to warm across Step 1 can be
discounted because saturation state decreased. This would have coincided with increased
dissolution across Step 1. The large increase of 29.88 µmol kg-1 between Step 1 and 2
suggests a significant change in carbonate chemistry within the ocean. It seems plausible that
this increase was caused by the predicted deepening of the CCD. Across Step 2, a small
decrease of 4.33 µmol kg-1 was calculated.
The effect of saturation state change on foraminiferal Mg/Ca was quantified using the B/Ca
derived Δ[CO32−]. The relationship of 0.0086 mmol mol-1 Mg/Ca per µmol kg-1 change in
Δ[CO32−] was used (Elderfield et al., 2006). A rise of 0.16 mmol mol-1 Mg/Ca across the EOT
was found, while the ~29 µmol kg-1 increase in saturation state that occurred just before Step
2 caused foraminiferal Mg/Ca to rise by 0.25 mmol mol-1. The decrease ending midway
through Step 1 shifted Mg/Ca concentrations down by 0.21 mmol mol-1.
The implications to Mg/Ca paleotemperatures of these findings are varied. Temperature
change calculated for Step 1 is likely to be higher than the true change, while the same is true
for Step 2. Any temperature change calculated using Mg/Ca records for between Step 1 and 2
will be notably lower than the true change in temperature.

4. DISCUSSION
71
Figure 4.2 Record of saturation state change relative to the oldest sample at ODP Site 757
Hole B, as calculated from the B/Ca of Cibicidoides havanensis (black: red triangles). Change
in foraminiferal δ18O across the Eocene-Oligocene transition at ODP Site 757B (black: black
diamonds) (Holmström, 2014). Vertical orange bars represent Steps 1 and 2 of the EOT, as
shown by the δ18O record of Holmström (2014). Ages use the Wade et al. (2011) timescale.

4. DISCUSSION
72
4.1.3. Support for Saturation State Change from Foraminiferal Test Mass
The saturation state record produced for ODP Site 757 showed a single large increase
between Steps 1 and 2. As stated above it can assumed that this is linked to the deepening of
the CCD during the EOT (Van Andel, 1975). Both B. jarvisi and C. havanensis appear to
have become larger across the EOT. There was a significant increase in the average mass of
each test as sample age decreased (Fig. 3.2). Possibly this was due to the reduced stress
associated with an increased carbonate saturation state. However, it seems strange that mass
appeared to increase linearly across the entire data set rather than in one distinct shift. This
suggests that the environmental conditions for calcite formation were improving throughout
the late Eocene and well into the early Oligocene. Saturation state did increase linearly across
the entire data set (P > 0.01), which supports the assumption of ever improving conditions for
benthic foraminifera during this period.
4.1.4. Inter-Site Comparisons of Saturation State Change
ODP Site 1263 is located on the Walvis Ridge in the South Eastern Atlantic Ocean at a water
depth of 2700 m. Site 1263 had a paleo water depth of ~1400 m during the Paleogene; this
depth was well above the lysocline (~2 km) (Zachos et al., 2004). A record was published
estimating an ~29 µmol kg-1 change in saturation state across the Eocene-Oligocene transition
at ODP Site 1263 (Peck et al., 2010). This record utilized a Li/Ca proxy to calculate saturation
state change.
The similarities between ODP Sites 757 and 1263 are striking. Both sites were located at
approximately the same paleo water depth during the Paleogene and lay well above the
carbonate dissolution horizons of the CCD and lysocline. It appears highly likely that the
increase of ~29 µmol kg-1 estimated by Peck et al. (2010) is associated with the same change
in carbonate chemistry as the 29.88 µmol kg-1 change presented here. An increase in bottom
water saturation state was also found in the equatorial deep waters of the pacific (~37 µmol
kg-1) (Lear and Rosenthal, 2006, Lear et al., 2010). Several sites, 744: Indian Ocean (Salamy
and Zachos, 1999) ; 1218: Equatorial Pacific (Coxall et al., 2005); 1263: Southern Atlantic
(Peck et al., 2010), show a transient decrease in %CaCO3 at (or marginally prior to) Step 1
(Fig. 4.3). Such a decrease is highly indicative of a dissolution effect. Peck et al. (2010)
attributed this decrease to an abrupt decrease in carbonate ion saturation state coincident with

4. DISCUSSION
73
the increases in foraminiferal δ18O. This observation ties closely with the abrupt decrease in
saturation state discovered, just before Step 1, in the samples from Site 757.
Figure 4.3 Evidence for dissolution at the onset of the Eocene-Oligocene transition in
different ocean basins. These include the Indian Ocean (Site 744) (Salamy and Zachos, 1999),
Pacific Ocean (Site 1218) (Coxall et al., 2005) and the Atlantic Ocean(Site 1263) (Riesselman
et al., 2007, Peck et al., 2010). Data shows (a) %CaCO3 and (b) benthic δ18O from Site 744;
(c) %CaCO3 and (d) benthic δ18O from Site 1218; and (e) %CaCO3 and (f) benthic δ18O from
Site 1263. Dashed grey line indicates coring disturbance. The vertical blue bar represents Oi-
1, termed in this paper as Step 2. Vertical yellow bar represents the dissolution event
proposed by Peck et al. (2010). ODP Site 1263 data is plotted against depth as opposed to
core age. Figure adapted from (Peck et al., 2010).
δ18O foram (‰)

4. DISCUSSION
74
4.1.5. Corrected Bottom Water Temperature Change Using C. Havanensis
Using the B/Ca proxy, it became possible to correct the Mg/Ca record recovered to account
for changes in saturation state (Fig. 4.4). The epifaunal record was chosen for Mg/Ca
paleothermometry because the effect of temperature and saturation state are better understood
for Cibicidoides than for Bulimina. The average temperature before the EOT was calculated
to be 1.5 ⁰C higher than the average temperature after the EOT. Beginning ~400 Ka before
Step 1, temperature dropped significantly by 6 ⁰C. This temperature decrease preceded the
start of decreasing saturation state by ~100 Ka. A further decrease of 3.5 ⁰C occurred before
temperatures rose by 3.1 ⁰C at the start of Step 1. Temperature continued to rise (+3.1 ⁰C)
during Step 1 before remaining constant for the last ~300 Ka of the step. Between the two
steps temperature drops dramatically by 6.2 ⁰C. This drop occurred at the same point as the
distinct 29 µmol kg-1 increase in saturation state. Step 2 saw a 3.9 ⁰C increase in temperature
after which temperature remains relatively stable. The temperature increase at Step 2 is
correlated with only a minor fall in saturation state. It is suggested that across Step 2
saturation state did not have a major effect on the Mg/Ca record. While a saturation state
effect appears to have affected clearly Mg/Ca across the transition, it occurred mainly during
Step 1 and the period directly following it.
Previous studies have indicated that the major climatic changes across the EOT were mainly
associated with the oxygen isotope shifts at Step 1 and Step 2. The proxy information
collected for ODP Site 757 appears to suggest differently. The largest changes in temperature
and saturation state occur between the two steps. Another strong indication that the variations
between the two steps represented major changes in the environmental conditions inhabited
by benthic foraminifera is the lack of Bulimina jarvisi. B. jarvisi are completely absent from
the record at ODP Site 757 from the end of Step 1, where temperature and saturation state
change significantly. It can be inferred that its disappearance was due to an inability to
survive in the post change conditions. It seems unlikely that increased saturation state, which
theoretically should be favourable to calcitic organisms, was responsible for the
disappearance. This allows the conclusion to be drawn that cooling bottom water temperature
was responsible for the loss of B. jarvisi from the fossil record. It should be noted that the
infaunal ostracod genera Krithe was missing also during this period.

4. DISCUSSION
75
Figure 4.4 Record of bottom water temperature change (green: circles) and saturation state change relative to the oldest sample at ODP Site
757 Hole B, as calculated from the Mg/Ca and B/Ca of Cibicidoides havanensis (black: red triangles). Change in foraminiferal δ18O across
the Eocene Oligocene transition at ODP Site 757B (black: black diamonds) (Holmström, 2014). Vertical orange bars represent Steps 1 and
2 of the EOT, as shown by the δ18O record of Holmström (2014). Ages use the Wade et al. (2011) timescale.

4. DISCUSSION
76
4.1.6. Inter-Site Comparisons of Bottom Water Temperature Change
The benthic foraminiferal Mg/Ca record recovered from ODP Site 1263 had a significant
decrease in Mg/Ca across Step 1 (Peck et al., 2010). The authors attributed this decrease to a
lowered saturation state rather than a cooling of bottom waters. This conclusion is supported
by the data from Site 757, which showed a saturation state decrease and a warming across the
same period. From the end of Step 1 to the end of Step 2, Mg/Ca at Site 1263 increased. Data
from Site 757 supports Mg/Ca increase at Site 1263. Both a significant increase in saturation
state at depth and the slight warming observed during Step 2, at Site 757, would have
magnified the increase in Mg/Ca discovered at Site 1263.
Some support for the Site 757 overall cooling of 1.5 ⁰C across the EOT exists from saturation
state corrected temperatures from other sites. A ~1.5 ⁰C decrease in temperature was found to
be associated with Step 2 at ODP Sites 1090 and 1265 (South Atlantic) (Pusz et al., 2011).
The record from Sites 1090 and 1265 did not indicate major temperature change between the
beginning of Step 1 and the beginning of Step 2 (Fig. 4.5). Although the temperature
increases associated with Steps 1 and 2 found in this study were not found at Sites 1090 and
1265, the trend across the entire EOT is broadly similar.
The TDP records published by Lear et al. (2008) strongly indicated a cooling stimulus for the
oxygen isotope shift at Step 1. TDP sites are not directly comparable with ODP sites due to
different depositional histories. However, it seems likely that if global cooling of bottom
waters existed at Step 1, it would be present in calibrated temperatures from both sets of sites.
As this is not the case with samples from ODP Site 757, other explanations must be
considered. It was shown that foraminiferal Mg/Ca from Site 1263, located well above the
lysocline, was affected by changes in ocean carbonate chemistry despite lying above the
dissolution horizon. If the Mg/Ca collected from the TDP sites was affected in a similar way
to ODP Site 1263 then paleotemperatures calculated without carbonate saturation state effect
correction will be invalid.

4. DISCUSSION
77
Figure 4.5 The records of benthic foraminiferal δ18O, saturation state corrected Mg/Ca and
saturation state corrected bottom water temperatures from ODP Sites 1090 (blue) and 1265
(red). Isotope values are reported relative to the Vienna PeeDee belemnite (VPDB) standard.
A three point running average is presented using a black line. Grey horizontal bars are
indicative of EOT 1, 2 and Oi-1. Oi-1 is termed here as Step 2. For the purposes of this thesis
EOT- 1 and 2 may be ignored. Figure adapted from (Pusz et al., 2011).

4. DISCUSSION
78
4.1.7. Changes in Ice Volume across the EOT
By applying the Mg/Ca bottom water temperatures, corrected for ∆[CO32−], to the
paleotemperature equation of Shackleton (1974), it was possible to partition the foraminiferal
δ18O into the temperature and δ18Osw components. An approximation of δ18Osw at ODP Site
757 allowed ice volume and sea level change to be calculated (Fig. 4.6).
The average value of δ18Osw before the EOT was found to be 0.2 ‰ lower than after the
transition. Directly before the EOT it was found that δ18Osw decreased markedly. This was
followed by an increase in δ18Osw (0.9 ‰) during the first half of Step 1. At the end of Step 1
δ18Osw drops dramatically by 1.8 ‰. Across Step 2, δ18Osw increased by 1.39 ‰. The oxygen
isotopic ratio within seawater is generally considered to reflect ice volume change. Increases
in global ice volume lower sea level.
For the Pleistocene it was found that a 0.1 ‰ increase in δ18Osw was approximately equal to a
10 m decrease in sea level (Shackleton and Opdyke, 1973). This relationship can be used to
estimate sea level change for other periods where δ18Osw is known. Using the average value of
δ18Osw before and after the EOT, the decrease in sea level across the Eocene-Oligocene
transition would appear to be ~20 m. The increase in δ18Osw during Step 1 represents a 90 m
fall in sea level, while the increase during Step 2 indicates a fall of ~140 m. Between the two
steps, a sea level increase appears to be indicated by the record, rising approximately 180 m.
This large increase corresponds with dramatic falls in temperatures and rises in saturation
state. Cumulatively the changes above indicate a sea level fall of ~50 m during the EOT.
A number of difficulties exist in interpreting these sea level changes. Firstly, concentrating
on the 140 m decrease in sea level at Step 2, it should be noted that a decrease of this size is
not possible when considering Antarctic ice growth alone. More ice would be required than
can be accommodated onto modern day Antarctica. Others have suggested that simultaneous
bipolar glaciation during the early Oligocene would provide enough ice growth to account for
the changes observed here (Coxall et al., 2005, Tripati et al., 2005). However, Edgar et al.
(2007) stated that no +evidence for extreme bipolar glaciation existed for the main Eocene
calcite compensation shift. Two plausible explanations for the ice growth signal observed at
ODP Site 757 remain. Firstly, it is possible that small northern hemisphere valley glaciers and
ephemeral ice caps existed. Some evidence for valley glaciers such as these has been
presented for Greenland during the Eocene-Oligocene transition (Eldrett et al., 2007). The

4. DISCUSSION
79
other explanation centres on the suggestion that the west Antarctic continent was up to 20%
larger in area during the late Eocene than it is in the modern day (Wilson and Luyendyk,
2009), which would allow for significantly more ice growth. A combination of these two may
be a better reflection of the true nature of EOT ice growth. It is safe to say that while a
significant period of Antarctic ice growth and sea level fall did occur during Step 2, it may not
have been of the magnitude shown here. Another factor other than Antarctic ice growth must
have played a role in moderating seawater oxygen isotopes or sea level.
Much the same can be said for the proposed sea level fall during Step 1 as for Step 2.
Previously published records have hypothesised a cooling driven change in foraminiferal
oxygen isotopes during Step 1 (Lear et al., 2008). However, little evidence for this is shown in
the record from ODP Site 757. Changes at Site 757 appear to be driven by ice growth similar
to like Step 2. The magnitude of the sea level fall, predicted by the Site 757 record for Step 1,
is incompatible with current predictions for the level of ice growth across the step. Therefore,
other controls on δ18Osw must be considered. Within the deep ocean δ18Osw is broadly
conservative, changing only due to changes in temperature and mixing (Ravelo and Hillaire-
Marcel, 2007). It is possible that the opening of oceanic gateways and the establishment of the
Antarctic Circumpolar Current modified deep water circulation during the Eocene-Oligocene
transition. Changes in δ18Osw broadly mirror the trends seen in the foraminiferal carbon
isotope record. Since carbon isotopic ratios are affected by mixing water masses, these
changes may provide an indication of mixing processes.
The δ13C ratios for ODP Site 757 (Fig. 4.6) are broadly similar to those for Sites 1265 and
522, while δ13C is approximately 1 ‰ lower at Sites 1090 and 1218. Pusz et al. (2011)
attributed the higher values of δ13C at Sites 1265 and 522 to differing source regions from
those of Sites 1090 and 1218. Therefore, it can be asserted that Site 757 has the same source
region as Sites 1265 and 522. High δ13C ratios are indicative of nutrient depleted waters,
which are likely to have a northerly origin. Unlike Sites 1265 and 522, foraminiferal δ13C is
raised after the EOT. This suggests a more northerly nutrient depleted source region after the
EOT than before it.
Changes in the δ18Osw record may reflect the regional salinity of Site 757 during the EOT.
Both ice growth and precipitation change salinity. Any water mass with a modified salinity
will change δ18Osw if it is mixed with the original bottom waters. Considering this, the

4. DISCUSSION
80
increase in δ18Osw shown at the beginning of Step 1 may indicate a higher regional salinity,
while any decrease in oxygen isotope values could indicate a lower regional salinity. The
increase in δ18Osw during Step 1 could result from the mixing of high salinity waters with
those originally at Site 757. High salinity waters would either indicate ice growth at the
source region or a decrease in precipitation. If the source region for Site 757 bottom had
changed, as could be the case if circulation changed upon the opening of oceanic gateways,
this could result also in modified salinity.

4. DISCUSSION
81
Figure 4.6 Record of bottom water temperature change relative to the oldest sample at ODP
Site 757 Hole B, as calculated from the Mg/Ca of Cibicidoides havanensis (black: diamonds).
Change in foraminiferal δ18O (red) and δ13C (green: triangles) across the Eocene Oligocene
transition at ODP Site 757B (Holmström, 2014). The change in the δ18O of seawater across
the EOT (blue), calculated using the equation of Shackleton (1974). Horizontal blue bars
represent Steps 1 and 2 of the EOT, as shown by the δ18O record of Holmström (2014). Ages
use the Wade et al. (2011) timescale.

4. DISCUSSION
82
4.1.8. Inter-Site Comparisons of δ18Osw
A record of δ18Osw was published for ODP Sites 1090 and 1295 across the EOT (Pusz et al.,
2011). At these sites eustatic sea level fall was shown to be approximately 45-90 m across
Step 2 while no significant ice growth was shown during any other part of the EOT. This ~70
m fall coincident with Step 2 is comparable with the 55-70 m decrease in sea level estimated
for other sites (Pekar et al., 2002, Miller et al., 2008, Katz et al., 2008). It is plausible that the
majority of the Step 2 shift in foraminiferal δ18O is accounted for by the ice volume change
observed at other sites. The studies mentioned above support the assumption of lack of ice
growth during Step 1 (Lear et al., 2008). The records from the above studies differ
significantly from the record recovered from ODP Site 757. Without the cooling associated
with Step 1, it becomes difficult to explain the shift in δ18Osw at Site 757 without large scale
ice growth. The saturation state corrected bottom water temperatures clearly show a
significant warming Over Step 1, in direct conflict with published records. Another problem
is presented by the cooling occurring between the two steps. It seems highly unlikely that ice
volume decreased globally with no indication from any other records. One explanation could
be that the initial prediction of the duration of Step 1 was shorter than the true length. Were
this to be the case, a cooling trend would have occurred across Step 1 going someway to
corroborating evidence from Site 757 with records from other studies. This argument,
however, lacks evidence due to the lack of a rise in foraminiferal δ18O after 33.7 Ma. A higher
resolution isotope record may shed light on this issue.

4. DISCUSSION
83
4.2. Indications from the Foraminiferal Li/Ca, Sr/Ca and U/Ca Records
4.2.1. Li/Ca
The temperature and saturation state changes, formulated using Mg/Ca and B/Ca, are
supported by the lithium calcium record of C. havanensis (Fig. 4.7a). As stated in section
4.1.6., before Step 1, temperature increased and saturation state fell. Over the same period the
Li/Ca signal decreased. Falling foraminiferal Li/Ca ratios are associated with decreasing
carbonate saturation state and increasing temperature. Therefore, the decrease in the Li/Ca
record presented here for Step 1 at Site 757 indicates strongly that the rising temperatures and
falling carbonate saturation state changes calculated from Mg/Ca are valid. The same can be
applied to the changes that occur between the two steps and for Step 2. The large cooling that
took place just after the EO boundary is coupled with a significant increase in saturation state.
As expected Li/Ca concentrations also increased. Changes in foraminiferal Li/Ca across Step
2 were in line with changes to bottom water temperature and saturation state. However, the
magnitude of these changes was negligible with regard to the resolution of the record. The
Li/Ca record consistently supports the calculated bottom water temperature and carbonate
saturation state records. Away from the EOT, the carbonate saturation state increases
significantly around 30 Ma. This is supported by increases in the foraminiferal Li/Ca record.
Therefore, it can be assumed that a significant saturation state increase did occur well after the
EOT, towards the middle Oligocene.
Comparing the foraminiferal Li/Ca collected from Site 757 samples with those collected from
Site 1263 reveals similarities in the trends observed at Step 1 (Fig. 4.7b). Li/Ca from Site
1263 increases by 2 mmol/mol prior to Step 1, before decreasing by 1 mmol/mol during Step
1. The initial rise in Li/Ca coincides with a rise in %CaCO3. Decreases in Li/Ca coincide with
decreasing %CaCO3 during Step 1. The Site 757 Li/Ca values mirror this trend, however
increases and decreases are about half the magnitude of those at Site 1263. This may be
explained by the use of different species or the differing site location. Interestingly, the
decreasing Li/Ca during Step 1 at Site 757 ties closely with the decreasing %CaCO3 shown to
exist at many other sites (Fig. 4.3). This further supports the theory that before the EOT a
large scale dissolution event occurred.

4. DISCUSSION
84
Figure 4.7 (a) Record of bottom water temperature (blue) and saturation state change (black)
relative to the oldest sample at ODP Site 757 Hole B, as calculated from the Mg/Ca and B/Ca
of Cibicidoides havanensis. Change in foraminiferal Li/Ca (red) across the Eocene Oligocene
transition at ODP Site 757B. Horizontal blue bars represent Steps 1 and 2 of the EOT, as
shown by the δ18O record of Holmström (2014). Ages use the Wade et al. (2011) timescale.
(b) a record of %CaCO3 (Riesselman et al., 2007) and benthic foraminiferal Li/Ca (Peck et al.,
2010) from ODP Site 1263. Figure 4.7b adapted from (Peck et al., 2010).
(b)
(a)

4. DISCUSSION
85
4.2.2. Sr/Ca
The foraminiferal Sr/Ca ratios from ODP Site 757 were rising until the beginning of Step 1 of
the EOT before dropping dramatically during Step 1. The concentrations of foraminiferal
Sr/Ca then increase between Steps 1 and 2 by around the same magnitude as the preceding
decrease. These trends are observed in both infaunal and epifaunal records. Compared with
these changes, foraminiferal Sr/Ca after the EOT is relatively stable. This indicates that a
saturation state decrease occurred during Step 1, while an increase in saturation state occurred
between the two steps. The assumption that the increases and decreases observed in
foraminiferal Sr/Ca ratios across the EOT are caused by a saturation state effect are supported
by the saturation state changes calculated from the B/Ca record, as well as the trends observed
within benthic foraminiferal Li/Ca. Foraminiferal Sr/Ca ratios are affected also by changing
water depth (of the order of -0.1 mmol/mol per km depth increase (Lear et al., 2003)).
Therefore, if paleowater depth changed across the EOT at Site 757, it may account for some
of the change observed in foraminiferal Sr/Ca ratios, which previously has been attributed to
saturation state change.
The rapid decreases in foraminiferal Sr/Ca discovered at Sites 1219 and 1220 after the
initiation of Step 2 are not seen in the record from Site 757 (Mawbey, 2012). The Site 757
record more closely resembles the trends observed at Site 1218, however these are not shown
clearly. The foraminiferal Sr/Ca concentrations at Site 1218 are relatively constant before and
after the transition, while between Steps 1 and 2 Sr/Ca decreases and then increases (Mawbey,
2012). These changes are of a similar magnitude. The Site 1218 decrease-increase trend is
remarkably similar to the one presented for ODP Site 757. If not for the significantly different
time span of the increase-decrease and that the two sites Sr/Ca match different parts of the
foraminiferal 18O record, it would be easy to see a clear correlation between the
foraminiferal Sr/Ca of the two sites. Higher resolution oxygen isotope analysis and plotting
both records on the same age model may yet show that these events occurred at the same
time. The radically differing trends shown at different sites of Leg 199 (Sites 1215-1222)
allowed Mawbey (2012) to conclude that changing seawater Sr/Ca was not the causal
mechanism of the decreases. The lack of the rapid decrease in foraminiferal Sr/Ca in the Site
757 record supports this conclusion.

4. DISCUSSION
86
4.2.3. U/Ca
The uranium calcium concentration within various Cibicidoides species (mundulus and
wuellerstorfi) has been shown to increase with decreasing carbonate saturation state (Fig 1.5)
(Raitzsch et al., 2011). With this in mind, both the infaunal record recovered from B. jarvisi
and the epifaunal record recovered from C. havanensis indicate a fall in carbonate saturation
state across the EOT at Site 757. The epifaunal record is relatively stable until the EO
boundary, at 33.7 Ma, at which point it rises steadily until 30 Ma. Due to the disappearance of
B. jarvisi from the fossil record between the two steps of the EOT, the infaunal U/Ca appears
to shift upward dramatically. This shift may be gentler, however this is impossible to prove
due to the lack of data. Whether gentle or steep, the increase in the infaunal U/Ca record
supports the assumption that saturation state decrease occurred across the EOT. The upward
trend in U/Ca concentrations was reproduced at sites on ODP Leg 199 (Mawbey, 2012).
Increasing mass accumulation rate of carbonate, CCD deepening, % calcium carbonate and
better shell preservation were all listed as reasons why increasing U/Ca could not indicate a
saturation state decrease across the EOT (Mawbey, 2012). The data collected from
foraminifera at Site 757 strongly supports the lack of a saturation state decrease across the
EOT. Calculated changes in saturation state show a clear increase between Step 1 and 2 of the
EOT. The increase is supported by the Li/Ca and Sr/Ca ratios. As sites in both the equatorial
Pacific Ocean and the Indian Ocean both show the increasing trend in U/Ca and saturation
state increase, it can be concluded that another mechanism is controlling U/Ca during this
period.
The relationship the U/Ca is negatively correlated with saturation state may not be true for the
species investigated in this study. However, due to the close genetic relationship of C.
havanensis and C. mundulus this can be ruled out. While it is expected that only redox
chemistry of the seafloor and saturation state have an effect on benthic foraminifera, it is
possible that bottom water temperature plays a role. The U/Ca of some planktonic
foraminifera has a temperature component, which results in a positive relationship between
temperature and U/Ca (Yu et al., 2008). Were this the case with the benthic foraminifera
studied at Site 757 and Leg 199, then a warming is indicated by the data. As a warming is not
shown by the Site 757 saturation state corrected bottom waters temperatures, this seems
unlikely. Records of bottom water temperatures from sites unaffected by saturation state

4. DISCUSSION
87
change indicate a cooling across the EOT, further ruling out warming as the causal
mechanism for increasing U/Ca (Lear et al., 2008).
Changes in the oxygen concentrations of pore waters can increase or decrease the levels of
authigenic, uranium bearing material being precipitated. However, no evidence exists for
changing redox conditions at the seafloor during the EOT. Further, the fact that the trend in
U/Ca occurs in two ocean basins seems to suggest that this change in pore water oxygen
concentration would have to be a global event, a highly unlikely scenario. Mawbey (2012)
attributed the increases in U/Ca concentration at leg 199 to a decreasing dissolution effect
across the EOT. It was noted that dissolution preferentially removes high uranium carbonate
from foraminifera (Russell et al., 2004, Yu et al., 2008). Were this the case, as saturation state
increases dissolution would fall in turn increasing U/Ca. This seems a likely explanation for
the trends in foraminiferal U/Ca observed at Site 757. It is suggested here that use of U/Ca as
an indicator of saturation state change is highly limited by the conflicting signal from
dissolution. While work is required to quantify the effect of dissolution on foraminiferal
U/Ca, it may be the case that U/Ca is better suited as a proxy for dissolution than it is for
saturation state.
A dissolution effect, as proposed above, has serious implications on the bottom water
temperatures calculated using Mg/Ca paleothermometry. If Mg/Ca is affected by dissolution
in a similar way to U/Ca, which has previously been proposed by Brown and Elderfield
(1996), some of the increase in foraminiferal Mg/Ca discovered at Site 757 may be due to a
dissolution affect. Mawbey (2012) attributed ~0.4 mmol/mol of the increase in foraminiferal
Mg/Ca at ODP Site 1218 to a dissolution affect such as this. Bar the U/Ca evidence, proof for
a dissolution effect is somewhat lacking. Therefore, more study is required before a definitive
conclusion can be drawn as to whether dissolution affected trace metal ratios in benthic
foraminifera across the EOT.

4. DISCUSSION
88
4.3. Surface Productivity Changes across the EOT
Abundance of benthic foraminifera provides a first order indication of surface productivity.
Here B. jarvisi abundance is discussed because it has been proposed as an excellent indicator
of surface productivity (Thomas et al., 1995). Abundances were clearly variable before the
EOT. These short term fluctuations occur at approximately 200 – 400 Ka cycles, suggesting
they are caused by orbitally driven climatic variations. A similar record of Late Eocene cyclic
variations in surface productivity was produced for ODP Sites 689 and 744 using numerous
methods (opal analysis, benthic foraminiferal accumulation rate and accumulation of other
calcitic organisms) (Diester-Haass, 1996, Diester-Haass and Zahn, 1996). Diester-Haass
(1996) published a record showing a notable upward spike in abundance at ~36 Ma. This
spike appears clearly on the record from Site 757. As Site 757 and 744 are both located in the
southern Indian Ocean in seems likely that the signal from the two sites is linked.
Stratification and increased vertical mixing during the late Eocene were proposed as a
mechanism for this spike (Diester-Haass, 1996).
During the EOT, B. jarvisi abundance shifted upward significantly. This shift occurred at the
same point as the saturation state increase shown by the record calculated using foraminiferal
B/Ca (Fig. 3.6). Increasing bottom water saturation state may have increased the abundance
signal of benthic foraminifera because of increased preservation of shells or more favourable
conditions for calcite formation. As both of these hypotheses centre around an effect on
benthic foraminifera only and not an increase in carbon flux to the seafloor, any increase in
abundance that occurs coincident with increasing saturation state can be ruled out as an
indicator of surface productivity increase.
From the end of the EOT, excluding two peaks at ~32.7 and ~32 Ma, abundance of benthic
foraminifera generally decreases. The abundance signal can be considered stable at low levels
from 32 Ma onward. Site 744 benthic foraminifera accumulation rates also showed the strong
peak at ~32 Ma (Diester-Haass, 1996). While the cause of the isolated peak is hard to qualify,
the occurrence at two Indian Ocean sites suggests strongly that an abrupt and short term
change in surface productivity occurred at this point in time. The decreasing abundances at
Site 757 do not correlate with the Site 744 accumulation rates, constraining the cause of the
trend is thus difficult. The surface productivity records from Sites 763 and 592 showed clear
increases from Eocene to Oligocene shedding further doubt on the assumptions that can be

4. DISCUSSION
89
drawn from the data produced here (Diester-Haass and Zahn, 2001). It is possible that
increased carbon flux, due to increased surface productivity, raised acidity locally at Site 757.
Carbonate dissolution can be increased when CO2 is produced by bacterial decay of organic
matter. If organic matter is increased, hypothetically dissolution could increase thus reducing
the preservation of benthic foraminifera (Diester‐Haass, 1992). More work is required to
ascertain the paleoproductivity of Site 757 during the EOT because the methodology used to
calculate abundances here is far from fool proof. A method involving the measurement of
course fraction directly after initial sieving and the use of all benthic foraminifera to calculate
a marine benthic accumulation rate would have been a better indicator of surface productivity.
It is hard to reconcile the records of δ13C with the abundance data collected from B. jarvisi
and C. havanensis. As stated above, abundance decreased indicating that surface productivity
decreased. The benthic foraminiferal δ13C increased markedly across the EOT before
decreasing to pre EOT levels from 32.5 Ma onward. In normal circumstances, an increase in
carbon isotope ratio would indicate a surface productivity increase because high surface
productivity concentrates 13C in the ocean. As abundance data suggests surface productivity
decreased, another mechanism for increasing δ13C must be sought. Ice growth occurred during
the EOT resulting in a decrease in sea level. This decrease exposed fresh limestone on the
continental shelf. Erosion of this limestone could increase global river inputs of dissolved
carbonate, which in turn would have increased seawater δ13C (Merico et al., 2008). This
seems a likely explanation when considering the ice growth calculated by this study at this
site.

5. SUMMARY
90
5. SUMMARY
Conclusions
Further work and future directions

5. SUMMARY
91
5.1. Conclusions
1. The alternative hypothesis presented in Section 1.10.1 stated that, saturation state
change was responsible for heightened ostracodal and foraminiferal Mg/Ca ratios
found across the Eocene/Oligocene boundary and not increasing temperature or
changing sea water Mg/Ca. Using the benthic foraminiferal B/Ca, a large increase in
deep water saturation state was discovered. Therefore, the alternative hypothesis
presented in this work can be accepted.
2. The hypotheses presented by Elderfield et al. (2010) and Dwyer et al. (1995), which
suggested that the environment inhabited by infaunal benthic foraminifera and
ostracods are relatively buffered against changes in saturation state and thus may be
used for Mg/Ca paleothermometry without saturation state correction, have been
disproven. The Mg/Ca ratios of infaunal benthic foraminifera and ostracods are clearly
affected by changing saturation state at ODP Site 757.
3. As predicted in Section 1.10.1, B/Ca and Li/Ca ratios in the benthic foraminifera C.
havanensis increased across the EOT. Foraminiferal U/Ca did not decrease, as
expected, but rose consistently throughout the EOT and beyond. The rise in
Foraminiferal U/Ca has been attributed to a dissolution affect across the EOT. It is
strongly suggested here that use of U/Ca as an indicator of saturation state change is
highly limited by the conflicting signal from dissolution. While work is required to
quantify the effect of dissolution on foraminiferal U/Ca, it may be the case that U/Ca
is better suited as a proxy for dissolution than it is for saturation state. The potential
discovery of a dissolution affect at both ODP Sites 757 and 1218 (Mawbey, 2012)
casts a doubt over the accuracy and magnitude of bottom water temperature changes
calculated from Mg/Ca ratios from benthic foraminifera. This is because Mg/Ca may
be affected by dissolution in a similar way to benthic foraminiferal U/Ca.
4. The trends in benthic foraminiferal test mass agreed with that predicted in section
1.10.1. Benthic foraminiferal test mass increased across the EOT. This increase in test
mass has been ascribed to increased calcification rates, a more readily available source
of carbonate ions and a less aggressive dissolution environment. The evidence

5. SUMMARY
92
gathered from test masses supports the conclusion that a significant saturation state
increase occurred across the EOT, coincident with deepening of the CCD. Infaunal
test mass did increase to a lesser degree than epifaunal test mass. Therefore, it can be
concluded that while not significant enough to reduce the saturation state effect on
Mg/Ca ratios some pore water buffering does exist in the infaunal micro habitat.
5. The benthic foraminiferal Mg/Ca record recovered from ODP Site 1263 had a
significant decrease in Mg/Ca across Step 1 (Peck et al., 2010). The authors attributed
this decrease to a lowered saturation state rather than a cooling of bottom waters. This
conclusion is supported by the data from Site 757, which showed a saturation state
decrease and a warming across the same period.
6. The sea level and ice growth picture at Site 757 is somewhat convoluted. There
appears to be significant ice growth across both Step 1 and Step 2 of the EOT. This is
in direct conflict with the majority of recently published studies, which find that Step
1 is associated with cooling and Step 2 ice growth. A rise in sea level between the two
steps is also not shown in previously published records. Furthermore, the magnitude of
sea level rises and falls calculated from the seawater oxygen isotope ratios are
unrealistic when considering Antarctic glaciation only. It is concluded here that,
another mechanism must have been affecting δ18Osw during the EOT, however more
evidence is required before a plausible mechanism can be reliably proposed.
7. Using a basic foraminiferal abundance approximation, it can be concluded that the
post EOT surface oceans were less productive than those before Antarctic glaciation.
Because abundance data suggests surface productivity decreased, another mechanism
for increasing δ13C was sought. Ice growth occurred during the EOT resulting in a
decrease in sea level. It can be concluded that, this decrease exposed fresh limestone
on the continental shelf. Erosion of this limestone could increase global river inputs of
dissolved carbonate, which in turn would have increased seawater δ13C. This seems a
plausible mechanism by which carbon isotopic ratios could rise without productivity
increases.

5. SUMMARY
93
5.2. Further Work and Future Direction
It is now clear that, almost all benthic foraminiferal Mg/Ca records must be corrected for
changes in dissolution and saturation state before they can be used for pelothermometry. The
use of infaunal species for Mg/Ca paleothermometry under the assumption of no saturation
state effect is wrong. It seems likely that any record of bottom water temperature or ice
volume published reliant on non-saturation state corrected Mg/Ca temperatures is likely to be
unreliable and of little use. With this in mind, it is suggested that future work concentrates on
making the paired Li/Ca Mg/Ca proxy for saturation state corrected temperatures more
accurate. The same can be said for the saturation state change calibrations that utilize B/Ca.
More species specific calibrations for all of the trace metal proxies commonly used to
reconstruct past climate are a must as this really holds back our ability to quantify the real
values for climatic variables in the past. Some time should be given to developing U/Ca ratios
in benthic foraminifera as a potential proxy for dissolution. The end goal of such a
development would be to correct Mg/Ca paleotemperatures so that dissolution affects no
longer cloud our interpretation of climate records.
With regard to the data presented here for ODP Site 757, it is hard to reconcile the changes in
bottom water temperature and ice volume with those of previous records. The production of
more records combined with an increased understanding of trace metal chemistry in
foraminifera will only make interpreting Site 757 easier. It is hoped with time and further
study, the exact mechanism which caused the Eocene-Oligocene transition can be qualified,
along with the exact changes in climate that occurred.

6. REFERENCES
94
6. REFERENCES
Altenbach, A. V., Pflaumann, U., Schiebel, R., Thies, A., Timm, S. and Trauth, M. 1999.
Scaling percentages and distributional patterns of benthic foraminifera with flux rates
of organic carbon. The Journal of Foraminiferal Research, 29(3), pp. 173-185.
Barker, S. and Elderfield, H. 2002. Foraminiferal calcification response to glacial-interglacial
changes in atmospheric CO2. Science, 297(5582), pp. 833-836.
Barker, S., Elderfield, H. and Higgins, J. 2003a. The future of the carbon cycle: review,
calcification response, ballast and feedback on atmospheric CO2. Philosophical
Transactions of the Royal Society of London. Series A: Mathematical, Physical and
Engineering Sciences, 361(1810), pp. 1977-1999.
Barker, S., Greaves, M. and Elderfield, H. 2003b. A study of cleaning procedures used for
foraminiferal Mg/Ca paleothermometry. Geochemistry, Geophysics, Geosystems, 4(9).
Bender, M. L., Lorens, R. B. and Williams, D. F. 1975. Sodium, magnesium and strontium in
the tests of planktonic foraminifera. Micropaleontology, pp. 448-459.
Berggren, W. A., Kent, D. V., Flynn, J. J. and Van Couvering, J. A. 1985. Cenozoic
geochronology. Geological Society of America Bulletin, 96, pp. 1407-1418.
Billups, K. and Schrag, D. 2003. Application of benthic foraminiferal Mg/Ca ratios to
questions of Cenozoic climate change. Earth and Planetary Science Letters, 209(1),
pp. 181-195.
Bohaty, S. 2014. RE: Site 763 ostracods. Type to CROWE, J.
Bohaty, S. M., Zachos, J. C. and Delaney, M. L. 2012. Foraminiferal Mg/Ca evidence for
Southern Ocean cooling across the Eocene–Oligocene transition. Earth and Planetary
Science Letters, 317, pp. 251-261.
Boyle, E. and Keigwin, L. 1985. Comparison of Atlantic and Pacific paleochemical records
for the last 215,000 years: Changes in deep ocean circulation and chemical
inventories. Earth and Planetary Science Letters, 76(1), pp. 135-150.
Boyle, E. A. 1981. Cadmium, zinc, copper, and barium in foraminifera tests. Earth and
Planetary Science Letters, 53(1), pp. 11-35.
Broecker, W. S., Peng, T.-H. and Beng, Z. 1982. Tracers in the Sea, Lamont-Doherty
Geological Observatory, Columbia University.
Brown, R. E., Anderson, L. D., Thomas, E. and Zachos, J. C. 2011. A core-top calibration of
B/Ca in the benthic foraminifers Nuttallides umbonifera and Oridorsalis umbonatus: A
proxy for Cenozoic bottom water carbonate saturation. Earth and Planetary Science
Letters, 310(3), pp. 360-368.
Brown, S. J. and Elderfield, H. 1996. Variations in Mg/Ca and Sr/Ca ratios of planktonic
foraminifera caused by postdepositional dissolution: Evidence of shallow
Mg‐dependent dissolution. Paleoceanography, 11(5), pp. 543-551.
Burton, E. A. and Walter, L. M. 1991. The effects of PCO2 and temperature on magnesium
incorporation in calcite in seawater and MgCl2-CaCl2 solutions. Geochimica et
Cosmochimica Acta, 55(3), pp. 777-785.
Clarke, F. W. and Wheeler, W. C. 1922. The inorganic constituents of marine invertebrates,
U. S Gov't. Print. Off.
Coccioni, R., Monaco, P., Monechi, S., Nocchi, M. and Parisi, G. 1988. Biostratigraphy of the
Eocene-Oligocene boundary at Massignano (Ancona, Italy). The Eocene-Oligocene
boundary in the Umbria-Marche Basin (Italy): Ancona, International Union of
Geological Sciences Special Publication: Ancona, Italy, Fratelli Annibelli Publishers,
pp. 59-74.

6. REFERENCES
95
Coggon, R. M., Teagle, D. A., Smith-Duque, C. E., Alt, J. C. and Cooper, M. J. 2010.
Reconstructing past seawater Mg/Ca and Sr/Ca from mid-ocean ridge flank calcium
carbonate veins. Science, 327(5969), pp. 1114-1117.
Coxall, H. K., Wilson, P. A., Pälike, H., Lear, C. H. and Backman, J. 2005. Rapid stepwise
onset of Antarctic glaciation and deeper calcite compensation in the Pacific Ocean.
Nature, 433(7021), pp. 53-57.
Crawford, W. 2013. National Science Foundation To Continue Operations Of The Drillship
JOIDES Resolution [Online]. Available at: http://oceanleadership.org/national-
science-foundation-continue-operations-drillship-joides-resolution/ [Accessed 05 Feb.
2015].
Deconto, R. M. and Pollard, D. 2003. Rapid Cenozoic glaciation of Antarctica induced by
declining atmospheric CO2. Nature, 421(6920), pp. 245-249.
Deconto, R. M., Pollard, D., Wilson, P. A., Pälike, H., Lear, C. H. and Pagani, M. 2008.
Thresholds for Cenozoic bipolar glaciation. Nature, 455(7213), pp. 652-656.
Demicco, R. V., Lowenstein, T. K., Hardie, L. A. and Spencer, R. J. 2005. Model of seawater
composition for the Phanerozoic. Geology, 33(11), pp. 877-880.
Diester-Haass, L. 1996. Late Eocene-Oligocene paleoceanography in the southern Indian
Ocean (ODP Site 744). Marine geology, 130(1), pp. 99-119.
Diester-Haass, L. and Zahn, R. 1996. Eocene-Oligocene transition in the Southern Ocean:
History of water mass circulation and biological productivity. Geology, 24(2), pp.
163-166.
Diester-Haass, L. and Zahn, R. 2001. Paleoproductivity increase at the Eocene–Oligocene
climatic transition: ODP/DSDP Sites 763 and 592. Palaeogeography,
Palaeoclimatology, Palaeoecology, 172(1), pp. 153-170.
Diester‐Haass, L. 1992. Late Eocene–Oligocene sedimentation in the Antarctic ocean,
Atlantic sector (Maud Rise, ODP Leg 113, Site 689): development of surface and
bottom water circulation. The Antarctic Paleoenvironment: A Perspective on Global
Change: Part One, pp. 185-202.
Dwyer, G. S., Cronin, T. M., Baker, P. A., Raymo, M. E., Buzas, J. S. and Correge, T. 1995.
North Atlantic deepwater temperature change during late Pliocene and late Quaternary
climatic cycles. Science, 268, pp. 1888.
Dwyer, G. S., Cronin, T. M. and Baker, P. A. 2002. Trace elements in marine ostracodes. The
Ostracoda: Applications in Quaternary Research, pp. 205-225.
Edgar, K. M., Wilson, P. A., Sexton, P. F. and Suganuma, Y. 2007. No extreme bipolar
glaciation during the main Eocene calcite compensation shift. Nature, 448(7156), pp.
908-911.
Egan, K. E., Rickaby, R. E., Hendry, K. R. and Halliday, A. N. 2013. Opening the gateways
for diatoms primes Earth for Antarctic glaciation. Earth and Planetary Science
Letters, 375, pp. 34-43.
Ehrmann, W. U. and Mackensen, A. 1992. Sedimentological evidence for the formation of an
East Antarctic ice sheet in Eocene/Oligocene time. Palaeogeography,
Palaeoclimatology, Palaeoecology, 93(1), pp. 85-112.
Elderfield, H., Yu, J., Anand, P., Kiefer, T. and Nyland, B. 2006. Calibrations for benthic
foraminiferal Mg/Ca paleothermometry and the carbonate ion hypothesis. Earth and
Planetary Science Letters, 250(3), pp. 633-649.
Elderfield, H., Greaves, M., Barker, S., Hall, I., Tripati, A., Ferretti, P., Crowhurst, S., Booth,
L. and Daunt, C. 2010. A record of bottom water temperature and seawater δ18O for

6. REFERENCES
96
the Southern Ocean over the past 440kyr based on Mg/Ca of benthic foraminiferal
Uvigerina spp. Quaternary Science Reviews, 29(1), pp. 160-169.
Eldrett, J. S., Harding, I. C., Wilson, P. A., Butler, E. and Roberts, A. P. 2007. Continental ice
in Greenland during the Eocene and Oligocene. Nature, 446(7132), pp. 176-179.
Elmore, A., Sosdian, S., Rosenthal, Y. and Wright, J. 2012. A global evaluation of
temperature and carbonate ion control on Mg/Ca ratios of ostracoda genus Krithe.
Geochemistry, Geophysics, Geosystems, 13(9).
Epstein, S., Buchsbaum, R., Lowenstam, H. A. and Urey, H. C. 1953. Revised carbonate-
water isotopic temperature scale. Geological Society of America Bulletin, 64(11), pp.
1315-1326.
Fariduddin, M. and Loubere, P. 1997. The surface ocean productivity response of deeper
water benthic foraminifera in the Atlantic Ocean. Marine Micropaleontology, 32(3–4),
pp. 289-310.
Ferguson, J., Henderson, G., Kucera, M. and Rickaby, R. 2008. Systematic change of
foraminiferal Mg/Ca ratios across a strong salinity gradient. Earth and Planetary
Science Letters, 265(1), pp. 153-166.
Francis, J., Marenssi, S., Levy, R., Hambrey, M., Thorn, V., Mohr, B., Brinkhuis, H.,
Warnaar, J., Zachos, J. and Bohaty, S. 2008. From greenhouse to icehouse–the
Eocene/Oligocene in Antarctica. Developments in Earth and Environmental Sciences,
8, pp. 309-368.
Goldner, A., Herold, N. and Huber, M. 2014. Antarctic glaciation caused ocean circulation
changes at the Eocene-Oligocene transition. Nature, 511(7511), pp. 574-577.
Hall, J. M. and Chan, L.-H. 2004. Li/Ca in multiple species of benthic and planktonic
foraminifera: Thermocline, latitudinal, and glacial-interglacial variation. Geochimica
et Cosmochimica Acta, 68(3), pp. 529-545.
Hay, W. W., Deconto, R., Wold, C.N., Wilson, K.M., Voigt, S., Schulz, M., Wold-Rossby,
A., Dullo, W.C., Ronov, A.B., Balukhovsky, A.N. , Soeding, E. 2000. ODSN Plate
Tectonic Reconstruction Service [Online]. Available at:
http://www.odsn.de/odsn/services/paleomap/adv_map.html [Accessed 12 Nov. 2014].
Herguera, J. C. and Berger, W. 1991. Paleoproductivity from benthic foraminifera abundance:
Glacial to postglacial change in the west-equatorial Pacific. Geology, 19(12), pp.
1173-1176.
Herguera, J. C. 1992. Deep-sea benthic foraminifera and biogenic opal: Glacial to postglacial
productivity changes in the western equatorial Pacific. Marine Micropaleontology,
19(1–2), pp. 79-98.
Holbourn, A., Henderson, A. S. and Macleod, N. 2013. Atlas of Benthic Foraminifera,
Hoboken, NJ, Wiley.
Holmström, M. 2014. ODP site 757 δ13C and δ18O records. Bsc project, Stockholm
University.
Houben, A. J., Van Mourik, C. A., Montanari, A., Coccioni, R. and Brinkhuis, H. 2012. The
Eocene–Oligocene transition: Changes in sea level, temperature or both?
Palaeogeography, Palaeoclimatology, Palaeoecology, 335, pp. 75-83.
Huh, Y., Chan, L.-H., Zhang, L. and Edmond, J. M. 1998. Lithium and its isotopes in major
world rivers: implications for weathering and the oceanic budget. Geochimica et
Cosmochimica Acta, 62(12), pp. 2039-2051.
Izuka, S. K. 1988. Relationship of magnesium and other minor elements in tests of
Cassidulina subglobosa and C. oriangulata to physical oceanic properties. The Journal
of Foraminiferal Research, 18(2), pp. 151-157.

6. REFERENCES
97
James, N. P. 1978. Neritic carbonate depositional environments. Sedimentology. Berlin:
Springer, pp. 759-775.
Katz, A. 1973. The interaction of magnesium with calcite during crystal growth at 25–90 C
and one atmosphere. Geochimica et Cosmochimica Acta, 37(6), pp. 1563-1586.
Katz, M. E., Miller, K. G., Wright, J. D., Wade, B. S., Browning, J. V., Cramer, B. S. and
Rosenthal, Y. 2008. Stepwise transition from the Eocene greenhouse to the Oligocene
icehouse. Nature Geoscience, 1(5), pp. 329-334.
Kennett, J. and Shackleton, N. 1976. Oxygen isotopic evidence for the development of the
psychrosphere 38 Myr ago. Nature, 260, pp. 513-515.
Ku, T.-L., Knauss, K. G. and Mathieu, G. G. 1977. Uranium in open ocean: concentration and
isotopic composition. Deep Sea Research, 24(11), pp. 1005-1017.
Lear, C. H., Elderfield, H. and Wilson, P. 2000. Cenozoic deep-sea temperatures and global
ice volumes from Mg/Ca in benthic foraminiferal calcite. Science, 287(5451), pp. 269-
272.
Lear, C. H., Rosenthal, Y. and Slowey, N. 2002. Benthic foraminiferal Mg/Ca-
paleothermometry: A revised core-top calibration. Geochimica et Cosmochimica Acta,
66(19), pp. 3375-3387.
Lear, C. H., Elderfield, H. and Wilson, P. 2003. A Cenozoic seawater Sr/Ca record from
benthic foraminiferal calcite and its application in determining global weathering
fluxes. Earth and Planetary Science Letters, 208(1), pp. 69-84.
Lear, C. H., Rosenthal, Y., Coxall, H. K. and Wilson, P. 2004. Late Eocene to early Miocene
ice sheet dynamics and the global carbon cycle. Paleoceanography, 19(4).
Lear, C. H. and Rosenthal, Y. 2006. Benthic foraminiferal Li/Ca: Insights into Cenozoic
seawater carbonate saturation state. Geology, 34(11), pp. 985-988.
Lear, C. H. 2007. Mg/Ca paleothermometry: a new window into Cenozoic climate change. In:
WILLIAMS, M., HAYWARD, A., GREGORY, F. & SCHMIDT, D. (eds.) Deep-time
Perspectives on Climate Change: Marrying the Signal from Computer Models and
Biological Proxies. London: Geological Society, pp.
Lear, C. H., Bailey, T. R., Pearson, P. N., Coxall, H. K. and Rosenthal, Y. 2008. Cooling and
ice growth across the Eocene-Oligocene transition. Geology, 36(3), pp. 251-254.
Lear, C. H., Mawbey, E. M. and Rosenthal, Y. 2010. Cenozoic benthic foraminiferal Mg/Ca
and Li/Ca records: Toward unlocking temperatures and saturation states.
Paleoceanography, 25(4).
Lemarchand, D., Gaillardet, J., Lewin, E. and Allegre, C. 2000. The influence of rivers on
marine boron isotopes and implications for reconstructing past ocean pH. Nature,
408(6815), pp. 951-954.
Lemarchand, D., Gaillardet, J., Lewin, E. and Allegre, C. 2002. Boron isotope systematics in
large rivers: implications for the marine boron budget and paleo-pH reconstruction
over the Cenozoic. Chemical Geology, 190(1), pp. 123-140.
Loubere, P. and Fariduddin, M. 1999. Benthic foraminifera and the flux of organic carbon to
the seabed. In: GUPTA, B. K. S. (ed.) Modern Foraminifera. Dordrecht Kluwer, pp.
181–199.
Lowenstein, T. K., Timofeeff, M. N., Brennan, S. T., Hardie, L. A. and Demicco, R. V. 2001.
Oscillations in Phanerozoic seawater chemistry: evidence from fluid inclusions.
Science, 294(5544), pp. 1086-1088.
Marchitto, T., Bryan, S., Curry, W. and Mccorkle, D. 2007. Mg/Ca temperature calibration
for the benthic foraminifer Cibicidoides pachyderma. Paleoceanography, 22(1).

6. REFERENCES
98
Marriott, C. S., Henderson, G. M., Belshaw, N. S. and Tudhope, A. W. 2004a. Temperature
dependence of δ 7 Li, δ 44 Ca and Li/Ca during growth of calcium carbonate. Earth
and Planetary Science Letters, 222(2), pp. 615-624.
Marriott, C. S., Henderson, G. M., Crompton, R., Staubwasser, M. and Shaw, S. 2004b. Effect
of mineralogy, salinity, and temperature on Li/Ca and Li isotope composition of
calcium carbonate. Chemical Geology, 212(1), pp. 5-15.
Martin, P. A., Lea, D. W., Rosenthal, Y., Shackleton, N. J., Sarnthein, M. and Papenfuss, T.
2002. Quaternary deep sea temperature histories derived from benthic foraminiferal
Mg/Ca. Earth and Planetary Science Letters, 198(1–2), pp. 193-209.
Martin, W. R. and Sayles, F. L. 1996. CaCO3 dissolution in sediments of the Ceara Rise,
western equatorial Atlantic. Geochimica et Cosmochimica Acta, 60(2), pp. 243-263.
Mawbey, E. M. 2012. Cenozoic Records of Seawater Chemistry: Chemical Proxies as
Indicators of Past Climate. PhD., Cardiff University.
Merico, A., Tyrrell, T. and Wilson, P. A. 2008. Eocene/Oligocene ocean de-acidification
linked to Antarctic glaciation by sea-level fall. Nature, 452(7190), pp. 979-982.
Miller, K. G., Browning, J. V., Aubry, M.-P., Wade, B. S., Katz, M. E., Kulpecz, A. A. and
Wright, J. D. 2008. Eocene–Oligocene global climate and sea-level changes: St.
Stephens Quarry, Alabama. Geological Society of America Bulletin, 120(1-2), pp. 34-
53.
Misra, S. and Froelich, P. N. 2012. Lithium isotope history of Cenozoic seawater: changes in
silicate weathering and reverse weathering. Science, 335(6070), pp. 818-823.
Misra, S., Greaves, M., Owen, R., Kerr, J., Elmore, A. C. and Elderfield, H. 2014.
Determination of B/Ca of natural carbonates by HR‐ICP‐MS. Geochemistry,
Geophysics, Geosystems, 15(4), pp. 1617-1628.
Murray, J. W. 2001. The niche of benthic foraminifera, critical thresholds and proxies. Marine
Micropaleontology, 41(1), pp. 1-7.
Nomura, R. 1995. Paleogene to Neogene deep-sea paleoceanography in the eastern Indian
Ocean: benthic foraminifera from ODP Sites 747, 757 and 758. Micropaleontology,
41(3), pp. 251-290.
Nürnberg, D., Bijma, J. and Hemleben, C. 1996. Assessing the reliability of magnesium in
foraminiferal calcite as a proxy for water mass temperatures. Geochimica et
Cosmochimica Acta, 60(5), pp. 803-814.
Pearson, P. N., Foster, G. L. and Wade, B. S. 2009. Atmospheric carbon dioxide through the
Eocene–Oligocene climate transition. Nature, 461(7267), pp. 1110-1113.
Pearson, P. N. 2012. Oxygen isotopes in foraminifera: overview and historical review.
Paleontological Society Papers, 18, pp. 1-38.
Peck, V. L., Yu, J., Kender, S. and Riesselman, C. 2010. Shifting ocean carbonate chemistry
during the Eocene‐Oligocene climate transition: Implications for deep‐ocean Mg/Ca
paleothermometry. Paleoceanography, 25(4).
Peirce, J. W., Weissel, J. K., Taylor, E., Dehn, J., Driscoll, N., Farrell, J., Fourtanier, E., Frey,
F. A., Gamson, P. D., Gee, J. S., Gibson, I. L., Janecek, T. R., Klootwijk, C.,
Lawrence, J. R., Littke, R., Newman, J. S., Nomura, R., Owen, R. M., Pospichal, J. J.,
Rea, D. K., Resiwati, P., Saunders, A. D., Smit, J., Smith, G. M., Tamaki, K., Weis, D.
and Wilkinson, C. 1989. Site 757. Broken Ridge and Ninetyeast Ridge; covering Leg
121 of the cruises of the drilling vessel JOIDES Resolution, Fremantle, Australia, to
Port of Singapore, sites 752-758, 30 April to 28 June 1988. College Station, TX:
Ocean Drilling Program.

6. REFERENCES
99
Pekar, S. F., Christie-Blick, N., Kominz, M. A. and Miller, K. G. 2002. Calibration between
eustatic estimates from backstripping and oxygen isotopic records for the Oligocene.
Geology, 30(10), pp. 903-906.
Prentice, M. and Matthews, R. 1991. Tertiary ice sheet dynamics: The snow gun hypothesis.
Journal of Geophysical Research: Solid Earth (1978–2012), 96(B4), pp. 6811-6827.
Pusz, A. E., Thunell, R. C. and Miller, K. G. 2011. Deep water temperature, carbonate ion,
and ice volume changes across the Eocene-Oligocene climate transition.
Paleoceanography, 26(2), pp. PA2205.
Rae, J. W., Foster, G. L., Schmidt, D. N. and Elliott, T. 2011. Boron isotopes and B/Ca in
benthic foraminifera: proxies for the deep ocean carbonate system. Earth and
Planetary Science Letters, 302(3), pp. 403-413.
Raitzsch, M., Kuhnert, H., Hathorne, E. C., Groeneveld, J. and Bickert, T. 2011. U/Ca in
benthic foraminifers: A proxy for the deep‐sea carbonate saturation. Geochemistry,
Geophysics, Geosystems, 12(6).
Rathmann, S., Hess, S., Kuhnert, H. and Mulitza, S. 2004. Mg/Ca ratios of the benthic
foraminifera Oridorsalis umbonatus obtained by laser ablation from core top
sediments: Relationship to bottom water temperature. Geochemistry, Geophysics,
Geosystems, 5(12).
Ravelo, A. C. and Hillaire-Marcel, C. 2007. The Use of Oxygen and Carbon Isotopes of
Foraminifera in Paleoceanography. In: HILLAIRE-MARCEL, C. & DE VERNAL, A.
(eds.) Developments in Marine Geology. Oxford: Elsevier Science Ltd, pp.
Riesselman, C., Dunbar, R., Mucciarone, D. and Kitasei, S. 2007. High resolution stable
isotope and carbonate variability during the early Oligocene climate transition: Walvis
Ridge (ODP Site 1263). Antarctica: A Keystone in a Changing World—Online
Proceedings of the 10th ISAES, edited by AK Cooper, and CR Raymond et al., US
Geol. Surv. Open File Rep, 1047.
Rohling, E. and Cooke, S. 2003. Stable oxygen and carbon isotopes in foraminiferal carbonate
shells. Modern Foraminifera. Springer Netherlands, pp. 239-258.
Rosenthal, Y., Boyle, E. A. and Slowey, N. 1997. Temperature control on the incorporation of
magnesium, strontium, fluorine, and cadmium into benthic foraminiferal shells from
Little Bahama Bank: Prospects for thermocline paleoceanography. Geochimica et
Cosmochimica Acta, 61(17), pp. 3633-3643.
Rosenthal, Y., Lohmann, G., Lohmann, K. and Sherrell, R. 2000. Incorporation and
preservation of Mg in Globigerinoides sacculifer: Implications for reconstructing the
temperature and 18O/16O of seawater. Paleoceanography, 15(1), pp. 135-145.
Russell, A. D., Emerson, S., Nelson, B. K., Erez, J. and Lea, D. W. 1994. Uranium in
foraminiferal calcite as a recorder of seawater uranium concentrations. Geochimica et
Cosmochimica Acta, 58(2), pp. 671-681.
Russell, A. D., Emerson, S., Mix, A. C. and Peterson, L. C. 1996. The use of foraminiferal
uranium/calcium ratios as an indicator of changes in seawater uranium content.
Paleoceanography, 11(6), pp. 649-663.
Russell, A. D., Hönisch, B., Spero, H. J. and Lea, D. W. 2004. Effects of seawater carbonate
ion concentration and temperature on shell U, Mg, and Sr in cultured planktonic
foraminifera. Geochimica et Cosmochimica Acta, 68(21), pp. 4347-4361.
Salamy, K. A. and Zachos, J. C. 1999. Latest Eocene–Early Oligocene climate change and
Southern Ocean fertility: inferences from sediment accumulation and stable isotope
data. Palaeogeography, Palaeoclimatology, Palaeoecology, 145(1), pp. 61-77.

6. REFERENCES
100
Savin, S. M. and Douglas, R. G. 1973. Stable isotope and magnesium geochemistry of recent
planktonic foraminifera from the South Pacific. Geological Society of America
Bulletin, 84(7), pp. 2327-2342.
Schedwin, T. 2014. ODP Hole 757B Age Model. BSc. Project, Stockholm University.
Scher, H. D. and Martin, E. E. 2006. Timing and climatic consequences of the opening of
Drake Passage. Science, 312(5772), pp. 428-430.
Shackleton, N. 1974. Attainment of isotopic equilibrium between ocean water and the
benthonic foraminifera genus Uvigerina: isotopic changes in the ocean during the last
glacial.
Shackleton, N. J. and Opdyke, N. D. 1973. Oxygen isotope and palaeomagnetic stratigraphy
of equatorial Pacific core V28-238: Oxygen isotope temperatures and ice volumes on
a 10 5 year and 10 6 year scale. Quaternary research, 3(1), pp. 39-55.
Sigman, D. M. and Boyle, E. A. 2000. Glacial/interglacial variations in atmospheric carbon
dioxide. Nature, 407(6806), pp. 859-869.
Sosdian, S. M., Lear, C. H., Tao, K., Grossman, E. L., O'dea, A. and Rosenthal, Y. 2012.
Cenozoic seawater Sr/Ca evolution. Geochemistry, Geophysics, Geosystems, 13(10).
Thomas, E., Booth, L., Maslin, M. and Shackleton, N. J. 1995. Northeastern Atlantic benthic
foraminifera during the last 45,000 years: Changes in productivity seen from the
bottom up. Paleoceanography, 10(3), pp. 545-562.
Toggweiler, J. and Bjornsson, H. 2000. Drake Passage and palaeoclimate. Journal of
Quaternary Science, 15(4), pp. 319-328.
Toyofuku, T., Kitazato, H., Kawahata, H., Tsuchiya, M. and Nohara, M. 2000. Evaluation of
Mg/Ca thermometry in foraminifera: Comparison of experimental results and
measurements in nature. Paleoceanography, 15(4), pp. 456-464.
Toyofuku, T. and Kitazato, H. 2005. Micromapping of Mg/Ca values in cultured specimens of
the high‐magnesium benthic foraminifera. Geochemistry, Geophysics, Geosystems,
6(11).
Tripati, A., Backman, J., Elderfield, H. and Ferretti, P. 2005. Eocene bipolar glaciation
associated with global carbon cycle changes. Nature, 436(7049), pp. 341-346.
Van Andel, T. H. 1975. Mesozoic/Cenozoic calcite compensation depth and the global
distribution of calcareous sediments. Earth and Planetary Science Letters, 26(2), pp.
187-194.
Van Der Zwaan, G. J., Duijnstee, I. a. P., Den Dulk, M., Ernst, S. R., Jannink, N. T. and
Kouwenhoven, T. J. 1999. Benthic foraminifers: proxies or problems?: A review of
paleocological concepts. Earth-Science Reviews, 46(1–4), pp. 213-236.
Wade, B. S., Pearson, P. N., Berggren, W. A. and Pälike, H. 2011. Review and revision of
Cenozoic tropical planktonic foraminiferal biostratigraphy and calibration to the
geomagnetic polarity and astronomical time scale. Earth-Science Reviews, 104(1), pp.
111-142.
Williams, F. 2008. Benthic Foraminiferal Shell Weight: A Potential Proxy for Seawater
Carbonate Saturation State and the Global Carbon Cycle. MESci. Thesis, Cardiff
University.
Wilson, D. S. and Luyendyk, B. P. 2009. West Antarctic paleotopography estimated at the
Eocene‐Oligocene climate transition. Geophysical Research Letters, 36(16).
Yu, J. and Elderfield, H. 2007. Benthic foraminiferal B/Ca ratios reflect deep water carbonate
saturation state. Earth and Planetary Science Letters, 258(1), pp. 73-86.

6. REFERENCES
101
Yu, J., Elderfield, H., Jin, Z. and Booth, L. 2008. A strong temperature effect on U/Ca in
planktonic foraminiferal carbonates. Geochimica et Cosmochimica Acta, 72(20), pp.
4988-5000.
Yu, J., Foster, G. L., Elderfield, H., Broecker, W. S. and Clark, E. 2010. An evaluation of
benthic foraminiferal B/Ca and δ 11 B for deep ocean carbonate ion and pH
reconstructions. Earth and Planetary Science Letters, 293(1), pp. 114-120.
Yu, J., Elderfield, H., Jin, Z., Tomascak, P. and Rohling, E. J. 2014. Controls on Sr/Ca in
benthic foraminifera and implications for seawater Sr/Ca during the late Pleistocene.
Quaternary Science Reviews, 98(0), pp. 1-6.
Zachos, J. C., Kroon, D. and Blum, P. 2004. Proc. ODP, Init. Repts., 208. College Station,
TX: Ocean Drilling Program.
Zhu, Z.-J., Chen, J.-A., Li, D.-H., Ren, S.-C. and Liu, F. 2012. Li/Ca ratios of ostracod shells
at Lake Qinghai, NE Tibetan Plateau, China: a potential temperature indicator.
Environmental Earth Sciences, 67(6), pp. 1735-1742.

7. ACKNOWLEDGMENTS
102
7. ACKNOWLEDGMENTS
First, I thank my supervisor Dr Caroline Lear for her support, direction and seemingly endless
knowledge of the topic under investigation.
This research used samples provided by the Ocean Drilling Program (ODP). ODP is
sponsored by the U.S. National Science Foundation (NSF) and participating countries under
management of Joint Oceanographic Institutions (JOI), Inc. Therefore, thanks are due to the
ODP and its partners.
My appreciation also goes to the technical staff in the school of Earth and Ocean Science at
Cardiff University. In particular Dr Elaine Mawbey, for cleaning samples and always making
time to help no matter how busy, and Dr Anabel Morte-Ródenas, for her knowledge of ICP-
MS without which analysis would not have been possible.
A mention must also go to Dr Helen Coxall and Dr Steve Bohaty for providing data and
advice related to this project.
Finally, I acknowledge the Cardiff Undergraduate Research Opportunities Programme for
funding the pilot study from which this investigation was based.

8. APPENDICES
103
8. APENDICES
8.1. Appendix 1
Cardiff University Tube Cleaning Procedure
General: Leave flow benches off overnight (extends life of filters), turn on for ~15min before
starting work in them and wash well with D.I. water. Leave 10% HCl in the flow bench, but
with the lid tightly closed, the nozzle (which falls off easily) wrapped in parafilm and bags
over both the lid and nozzle. Fume cupboard needs turning on at the black switch under the
cover, as well as the switch on the front at the left. Leave the lids to bottles upside down on
the work bench, and avoid touching the rim of bottles when filling.
1. Rinse the large beaker and watch glass well with D.I. water.
2. Fill the beaker (~3/4 full) with 10% HCl (remember to unscrew the top to the acid
container).
3. Clean a disposable pipette with a few rinses of 10% HCl – remember to dilute to waste.
4. Squirt 10% HCl into the tubes, and remove any bubbles (either tap the tubes on the base or
suck out bubbles with the pipette), fill the lids too.
5. Drop (tubes) into the beaker of acid.
6. Once you are done, check the tubes to see if any bubbles have got into the tubes, and
remove as necessary.
7. Place the watch glass on the beaker, and the beaker on the hot plate, Set hot plate to:
1) 100 for >1hr
2) 150 for a further >1hr
3) 200 – bubbles will start to form. Hold at this temperature until the end of the day and then
TURN THE HOTPLATE OFF.
8. Leave the tubes in acid overnight (with the fume cupboard on).
9. Next day; clean flow benches again with D.I. water and leave on for ~15mins before
starting work.

8. APPENDICES
104
10. In the fume cupboard, decant the acid back into its container, taking care not to allow any
tubes to follow the acid in (this is most easily achieved by holding the watch glass on top of
the lid).
11. Fill the beaker with D.I. water and agitate (swirl around/shake gently/poke) the tubes in
the diluting water.
12. Pour the dilute acid to waste
13. Repeat 11-12 times 3
14. Carefully pick up a tube, by the pointy end if possible (this is easiest if the beaker is ~3/4
full of water).
15. “Flick” out the dilute acid in the bottom of the tube, and place between thumb and
forefinger (or between fore and middle fingers).
16. Repeat 14-15 until you have 4-6 in your hand.
17. Using your “D.I. water for tubes” bottle squirt water into the tubes, lids, and rinse the
hinge and around the cap.
18. Pour out the water and using a gangster hand gesture flick out the dregs in the bottom. Do
this in the general vicinity of the sink, but not too close as there is Mg in the sink (tap water).
19. Repeat, 17-18 4-6 times, dependant on your current levels of paranoia
20. Place the now empty tubes on the flow bench, open end facing into the airflow to dry.
21. Repeat steps 14-20 until your beaker is empty.
22. When the tubes are dry (they do not take very long) carefully close the caps, avoiding
touching the rim and inside of the lids.
23. Place them in a fresh zip lock bag, and label with your name, what they are, the date and
the number in the bag.
24. Clean the flow bench, fume cupboard and work surface with D.I., rinse out the beaker and
watch glass, switch off the flow bench, D.I. water and if safe to do so the fume cupboard.
25. Switch off all lights.

8. APPENDICES
105
8.2. Appendix 2
Foraminifera Cleaning Procedure for Room 2.10
J. Riker, September 2008
This description is modified from the “Foraminifera Procedure for Cadmium” recorded by
Paula Rosener (1988), as updated to reverse the redox steps (1994). Our procedure differs in
that the sample transfer is performed between reduction and oxidation. Our procedure is also
expanded to include instructions specific to our lab. It is not necessary to perform Step III in
all cases; please confirm the appropriate cleaning process before starting.
I. Before You Start
Set your tubes of crushed foraminifera in a clean, Perspex rack. Ensure samples are
clearly labelled with permanent ink and record a list or diagram of samples before
starting. Randomize your samples prior to cleaning.
Locate your reagents (remake or refill as necessary). All reagents should be prepared
and stored in new, acid-leached PE bottles. You will need:
10% HCl or HNO3 for rinsing pipette tips (250 mL; top up bottle from labelled jars in
the flow bench each day you clean)
DI H2O for rinsing pipette tips (250 mL; rinse and refill bottle with fresh water each
day you clean)
DI H2O for foraminifera cleaning (500 mL wash bottle; rinse and refill with fresh
water each day you clean)
Trace grade methanol for foraminifera cleaning (store in a 250 mL bottle; pour off a
small amount into a 125 mL spray bottle just before use)
Empty 60 mL bottle for reducing reagent
Empty 60 mL bottle for rinsing reducing reagent
Empty 60 mL bottle for oxidizing reagent
DI H2O for sample transfers (250 mL; rinse and refill bottle with fresh water each day
you clean)

8. APPENDICES
106
0.002 M HNO3 for foraminifera leaching (0.001 M for small samples) (250 mL; make
fresh if old or contaminated)
Turn on flow bench and allow to run for at least 15 minutes before using.
Wipe down all work surfaces with DI water before starting (counter surfaces, interior
of flow benches and fume cupboard, equipment surfaces).
If floor has not been mopped recently, mop floor using DI water from the reservoir in
Room 2.12.
II. Removal of Fine Clays
1. Drain the ultrasonic bath in the flow bench and refill with fresh DI H2O. Fill to the
base of your Perspex rack. Use the prop provided and never fill below the minimum
fill line.
2. Tap your sample rack firmly on the bench to shake foraminifera to the base of tubes.
3. Open tube tops slowly in case foraminifera are stuck to the sides or lids.
4. Using your DI H2O for foraminifera cleaning, gently fill each tube most of the way.
5. If foraminifera are visible in the tube lids, add a small amount of water to the lids as
well. Close tubes.
6. Tap rack firmly on the bench to settle foraminifera and get rid of any air bubbles.
CHL: If foraminifera will not settle or bubbles will not rise, tap the side or corners of
the rack firmly on the edge of the flow bench.
7. Turn on siphon (switch is on the rear of the pump, on the right-hand side). Always
make sure the siphon tip is in the flow bench when the pump is on.
8. If siphon waste beaker is full, empty into the labelled waste container beneath the
water purifier. Check the waste level throughout the cleaning process and do not let it
rise above the “max fill” line.
9. Rinse siphon tip in 10% HCl (3x) and then DI H2O (3x) tip rinses. Do not siphon up
too much tip rinse at once, as this can cause siphon waste to splash and contaminate
the pump tubing.

8. APPENDICES
107
10. Siphon off as much water as possible from the tubes. This works best if you avoid
putting the siphon tip directly in the sample. Instead, rest the tip against the front of
the tube, above water level, and siphon down gradually.
11. Open all tubes and fill ~1/3 full with water (not quite up to the rack base). Close
tubes. CHL: The foraminifera will agitate best in minimal H2O.
12. Tap rack as necessary to remove air bubbles.
13. Ultrasonicate for 1 minute (set the bath to “hold”). Fine clays should now be
dislodged and held in suspension.
14. Turn off bath and remove rack. Open all tubes and vigorously squirt DI H2O for
foraminifera cleaning into each tube so as to agitate the sample and mix clays
throughout. Close tubes.
15. Tap rack firmly on the bench, invert and shake, then wait for foraminifera to settle.
Do not wait too long, or suspended clays will also settle. If necessary, tap the rack
again to encourage foraminifera to settle.
16. Clean siphon tip (3x 10% HCl and 3x DI H2O) while waiting for foraminifera to settle.
17. Siphon off as much water as possible. Do not siphon off your foraminifera.
18. Repeat steps 11-17 a total of 3x with DI H2O. To avoid systematic variations in the
effectiveness of clay removal: Begin siphoning at a different row and (or) side of the
rack during each rinse step. Change the orientation of the rack in the sonic bath during
each sonication.
19. Fill 125 mL spray bottle ~1/5 full with trace grade methanol. Loosen cap of spray
bottle when not in use to keep methanol from dripping from the tip.
20. Repeat steps 11-17 1-2x with trace grade methanol, depending on the degree of clay
contamination in your samples. Special instructions for methanol:
Always wear goggles when working with methanol
Fill tubes to the top of the rack with methanol (rather than just 1/3 full)
Do not add additional methanol after ultrasonicating; simply siphon off
existing methanol

8. APPENDICES
108
Methanol is less viscous than water, so take special care when siphoning; do
not go to quite to the bottom of the tube
When siphoning methanol, it may work better to press siphon tip against the
rear of the tube (rather than the front)
Dispose of any leftover methanol in the labelled waste container
21. Repeat steps 11-17 an additional 2x with DI H2O.
22. Pipette off all remaining water using a clean (3x 10% HCl and 3x DI H2O), 100 μl
(yellow) pipette tip. It is not necessary to rinse the tip between samples.
III. Removal of Metal Oxides (Reducing Step)
1. Turn on power source for hotplate in the fume cupboard and set to 300 °C.
2. Rinse and fill the glass evaporating dish in the fume cupboard with DI H2O from the
ELGA tap. Fill to the base of your Perspex rack. Set on hotplate.
3. Rinse and fill tall form beaker containing thermometer with DI H2O from the ELGA
tap. Set on hot plate. Use this to top up the evaporating dish as water evaporates.
4. Drain the ultrasonic bath in the fume cupboard and refill with fresh DI H2O. Fill to
the base of your Perspex rack. Use the prop provided and never fill below the
minimum fill line.
5. Prepare your reducing reagent in the labelled, empty 60 mL bottle. Please note that
hydrous hydrazine is volatile, carcinogenic, and explosive. Always work in the fume
cupboard and take care to minimize exposure. Dispose of all related waste (pipette
tips, parafilm, and gloves) in a plastic bag and seal bag before removing from fume
cupboard.
Pour 10 ml ammonia solution and 10 ml citric acid/ammonia solution (both
stored in the fridge) into the empty bottle; pour these reagents directly from the
bottles (no pipettes) and take care not to touch the lids of reagent bottles to any
other surfaces.
Prepare a waste bag, a fresh strip of parafilm, and a clean (3x 10% HCl and 3x
DI H2O) 1000 μl (blue) pipette tip

8. APPENDICES
109
Remove hydrous hydrazine from fridge
Pipette 1200 μl hydrous hydrazine into the reducing reagent
Dispose of pipette tip
Cap reducing reagent and invert to mix
Re-parafilm hydrous hydrazine and return to fridge
6. Before proceeding, ensure hot water bath is hot (on verge of boiling, 80-90 °C). This
can take about 30 minutes.
7. Open tubes. Using a clean (3x 10% HCl and 3x DI H2O) 100 μl pipette tip, add 100
μL reducing reagent to each tube. Be aware that the reagent has a low viscosity and
tends to drip. Close tubes firmly.
8. Because ammonia has a high vapour pressure, tube caps will tend to blow open in the
hot water both. To prevent this, clamp tubes shut by screwing a Perspex plate to the
top of your rack. Ensure your tubes are firmly closed and that they are in good contact
with the plate surface.
9. Place racks in the hot water bath for a total of 30 minutes. Calcium carbonate is
slightly soluble in ammonia, so avoid letting your foraminifera fragments sit in the
reducing agent for longer than the necessary 30 minutes. Every 2 minutes:
Remove rack
Tighten screws on Perspex clamp
Invert, shake, and tap rack to settle foraminifera and remove bubbles
Ultrasonicate rack for a few seconds (this will agitate the reagent into all parts
of the sample and discourage dissolved oxides from re-precipitating)
Tap rack firmly and return to hot water bath
Top off the water bath as necessary using hot water from the beaker
10. After 30 minutes, remove rack and clamp and carefully open and close all tubes to
release gas. Keep one finger on the top of the tube and use your thumb to open the
tube in a peeling motion.

8. APPENDICES
110
11. Pipette off as much reducing reagent as possible using a clean (3x 10% HCl and 3x DI
H2O) 100 μL pipette tip. Do not use siphon. Eject waste into the reducing reagent
bottle. Eject tip into waste bag.
12. Fill tube caps and tubes (to top of rack or higher) with DI H2O for foraminifera
cleaning. Close tubes. Tap rack firmly to settle foraminifera.
13. Turn on the siphon in the fume cupboard using the labelled control knob on the left-
hand panel. Rinse the siphon tip (3x 10% HCl and 3x DI H2O). Siphon caps and then
siphon off as much water as possible from tubes.
14. Repeat steps 12 and 13 two more times.
15. Fill tubes half full with DI H2O for foraminifera cleaning, close tubes, then set in the
hot water bath for 5 minutes.
16. In the meantime, prepare a fresh strip of parafilm.
17. Remove hydrazine waste container (brown bottle) from fridge and place in fume
cupboard.
18. Dump leftover reducing reagent into waste container.
19. Fill the empty 60 mL bottle for rinsing reducing reagent with DI H2O from the ELGA
tap.
20. Rinse the reducing reagent bottle 2-3x with DI H2O, dumping rinse water into the
waste container.
21. Re-parafilm waste container and return to fridge.
22. If 5 minutes have passed, remove rack from hot water bath, clean siphon tip (3x 10%
HCl and 3x DI H2O), siphon caps, and then siphon off as much water as possible from
tubes.
23. Repeat steps 12 and 13 two more times.
24. Repeat step 15.
25. Repeat step 22. It is now safe to remove the rack from the fume hood.
26. Turn hotplate off or down as appropriate (you will need it again in Section V). Please
remember to turn off the power source as well.

8. APPENDICES
111
IV. Sample Transfer
1. In the flow bench, label a new set of acid-leached tubes for your samples.
2. Using a disposable scalpel, cut off ~1/4 of a 100 μL pipette tip.
3. Set the pipettor to 70 μL and thoroughly clean the pipette tip (6x 10% HCl and 6x DI
H2O).
4. If you have not already, rinse and refill your DI H2O for sample transfers.
5. Open an old tube. Hold pipette tip directly over foraminifera fragments and pipette
and expel fragments (± H2O) into the new tube of the same sample number.
6. Add a small amount of DI H2O for sample transfers to the old tube. Repeat transfer
until no foraminifera fragments are visible in the old tube and then again once more
(usually 2-3x).
7. Between samples, rinse the pipette tip 2-3x in your DI H2O for sample transfers.
8. Once all samples have been transferred into new tubes, turn on the siphon, clean the
siphon tip (3x 10% HCl and 3x DI H2O), and siphon off as much water as possible.
V. Removal of Organic Matter (Oxidizing Step)
1. In the fume cupboard, ensure hot water bath is hot (on verge of boiling, 80-90 °C) and
filled to the base of your Perspex rack.
2. Prepare your oxidizing reagent in the labelled, empty 60 mL bottle.
Pour 15 mL 0.1 N NaOH (stored in the fridge) into the empty bottle; pour this
reagent directly from the bottle (no pipettes) and take care not to touch the lid
of the reagent bottle to any other surface
Using a clean (3x 10% HCl and 3x DI H2O) 100 μL pipette tip, add 50 μL
H2O2; SB: please pour a small quantity of H2O2 into the H2O2 bottle cap,
pipette from the cap, and dispose of cap contents before re-capping the bottle
Cap reagent bottle and invert to mix

8. APPENDICES
112
3. Open tubes and add 250 μL oxidizing reagent to each sample. Close tubes.
4. Set rack in hot water bath for 5 minutes.
5. Remove rack and invert, shake, and tap the rack to settle foraminifera and remove
bubbles. Ultrasonicate rack for a few seconds, then tap rack firmly and return to hot
water bath.
6. Repeat steps 4 and 5.
7. Open tubes and top them off with DI H2O for foraminifera cleaning.
8. Turn on siphon, clean siphon tip (3x 10% HCl and 3x DI H2O), and siphon off
oxidizing reagent.
9. Repeat steps 7 and 8 two more times.
VI. Dilute Acid Leach
1. In the flow bench, clean a 1000 μL pipette tip (3x 10% HCl and 3x DI H2O).
2. Add 250 μL 0.002 N HNO3 to each tube. Because HNO3 will dissolve carbonate, you
may wish to use 0.001 N HNO3 for small samples. You may also wish to skip
ultrasonication and do fewer (or no) repetitions of the leach.
3. Tap the rack firmly and check for air bubbles. If necessary, tap some more.
4. Ultrasonicate the rack for 30 seconds.
5. Remove rack from bath. Invert, shake, and tap rack firmly to settle foraminifera.
6. Open tubes. While waiting for foraminifera to settle, turn on siphon and clean siphon
tip (3x 10% HCl and 3x DI H2O).
7. Once foraminifera have settled, siphon off as much acid as possible.
8. Repeat steps 2-7 4x as quickly as possible to avoid dissolving your samples. To avoid
systematic variations in the effectiveness of the acid leach: Begin siphoning at a
different row and (or) side of the rack during each rinse step. Change the orientation
of the rack in the sonic bath during each sonication.
9. Fill tubes and caps with DI H2O for foraminifera cleaning. Close tubes.

8. APPENDICES
113
10. Tap rack firmly, check for bubbles, and ultrasonicate for a few seconds.
11. Remove rack from bath. Invert, shake, and tap rack firmly to settle foraminifera.
12. Turn on siphon and clean siphon tip (3x 10% HCl and 3x DI H2O). Once foraminifera
have settled, siphon caps and then siphon off as much water as possible from tubes.
13. Repeat steps 10-13.
14. Pipette off all remaining water using a clean (3x 10% HCl and 3x DI H2O), 100 μL
pipette tip. It is important to remove as much water as possible. Use a new, freshly-
cleaned tip for each sample.
Your samples may be stored indefinitely at this point.

8. APPENDICES
114
8.3. Appendix 3
Dissolving and Diluting Benthic Foraminifera for Trace Metal Analysis
J. Riker, May 2008
The following protocol describes how to dissolve and dilute cleaned benthic foraminifera for
trace metal analysis on the Element (using the FULL_FORAMINIFERA_LMR method). This
protocol assumes you have ~20 small tests per sample. If you have large or bulky samples, or
if you are analysing a different species, please consult with a lab manager to ensure that your
final sample concentrations fall within the working range of the instrument.
Dissolving Cleaned Foraminifera
1. Before starting, check to see that fluid was successfully siphoned from all tubes following
the weak acid leach. Remove any excess fluid with a clean pipette tip (6x HNO3 tip rinse, 6x
H2O tip rinse).
2. Add 120 mL 0.065 M Optima HNO3 (see “reagents,” below) to each tube with a clean
pipette tip (6x HNO3 tip rinse, 6x H2O tip rinse). Be aware that static may draw dry
foraminifera up the sides of the tubes; open tubes gently, one at a time, with the cleaned and
acid-filled pipette tip ready to dispense.
3. After filling, close caps, invert each sample, and mix for a few seconds on the vortex tube
stirrer. Centrifuge each tube for 3 minutes.
4. Leave overnight to dissolve. If samples do not dissolve overnight, try the following (in
order):
a. Release any built up CO2 by gently opening and closing tube (trapped CO2 will buffer the
dissolution reaction).
b. Invert and rotate tube (this should expose more of the foraminifera’s surface area to the
dissolution acid and disturb any buffered environment that may exist at the tube base). Watch
to see if foraminifera dissolve.
c. Ultrasonicate tubes (same effect as above). Check to see if foraminifera have dissolved.
d. Place tubes in the fridge overnight (carbonate is more soluble at low temperature). Check to
see if foraminifera have dissolved.

8. APPENDICES
115
e. As a last resort (if 10-20 foraminifera remain undissolved), add an additional 50-100 mL
0.065 M HNO3 to the tube. Transfer foraminifera plus acid to a new, clean centrifuge tube
before adding more acid. Never fill tubes used during the cleaning process above the 120 mL
mark, as micro fractures often form in these tubes during cleaning.
Splitting Dissolved Samples for Ratio and Ca Concentration Analysis
1. If your samples have been refrigerated, allow them to warm to room temperature before
proceeding.
2. Once you have confirmed that samples are dissolved, label two sets of clean tubes with the
appropriate sample numbers. Label one set of tubes with the letters “CC” (for Ca
concentration analysis) and the other set with “TM” (for trace metal ratio analysis).
3. Invert, vortex, and centrifuge all samples.
4. Set pipettor to 100 mL and clean a pipette tip (6x HNO3 tip rinse, 6x H2O tip rinse). Open
the first trace metal ratio tube you wish to fill.
5. Gently remove the corresponding sample tube from the centrifuge (always double check
the sample number), taking care not to disturb the sediment pack at the tube base.
Keep the tube in the same orientation it was sitting at in the centrifuge and walk slowly to the
flow bench.
6. Gently open the tube and withdraw 100 mL of dissolved sample. Keep the pipette tip as
close to the fluid surface as possible, moving the tip down as you pipette. This will help you
to avoid sucking up solid contaminants at the tube base. Pipette the fluid into the opened
empty tube.
7. Repeat with all tubes for trace metal analysis, using a new, cleaned pipette tip for each
sample.
8. Re-centrifuge remaining dissolved samples.
9. Set pipettor to 10 mL and clean a pipette tip (3x HNO3 tip rinse, 3x H2O tip rinse). Open
the first Ca concentration tube you wish to fill.
10. Using the same technique described above, pipette 10 mL of dissolved sample into each
tube for Ca concentration analysis.

8. APPENDICES
116
11. As a rule, always pipette off 100 mL of each dissolved sample for ratio analyses before
pipetting off 10 mL for Ca analyses. This will help to minimize contamination of your ratio
samples, which require more accurate analysis than your Ca samples, with particles or
leachants concentrated at the tube base.
12. Discard (or label and store, if you prefer) the original dissolved sample tubes.
Diluting Dissolved Samples for Ca Concentration Analyses
1. On the day you plan to run your Ca concentration analyses, add 190 mL 0.5 M Optima
HNO3 (see “ reagents”) to each Ca concentration sample tube using a clean pipette tip (6x
HNO3 tip rinse, 6x H2O tip rinse), for a total sample volume of 200 mL.
2. Invert, vortex, and centrifuge sample tubes immediately prior to analysis.
Diluting Dissolved Samples for Ratio Analysis and Matrix-Matching Standards
1. Run Ca concentration analyses to determine the appropriate dilution of a matrix-matched
standard (a standard of roughly equivalent Ca concentration) for each trace metal ratio
sample. To calculate volumes of acid and standard to pipette, use the template spreadsheets
provided by the lab manager.
2. Label a set of clean centrifuge tubes with the appropriate sample numbers and the letter
“S” for standard. If you are planning to run your blanks in tubes (rather than in a vial), prepare
an additional set of tubes for the blanks.
3. On the day you plan to run your ratio analyses, add 250 mL 0.5M Optima HNO3 to each
trace metal ratio sample tube using a clean pipette tip (6x HNO3 tip rinse, 6x H2O tip rinse),
for a total sample volume of 350 mL.
4. Pipette each matrix-matched standard using your calculated volumes of standard and 0.5 M
Optima HNO3, for a total standard volume of 350 mL (same as your samples).
5. If necessary, pipette tube blanks (350 mL 0.5M Optima HNO3).
6. Invert, vortex, and centrifuge sample tubes immediately prior to analysis. Stir standard
tubes on the vortex, but it is not necessary to centrifuge them.
Reagents

8. APPENDICES
117
Before preparing reagents, have a lab manager pour an aliquot of concentrated Optima HNO3
into an acid-cleaned 60 mL Nalgene bottle. Pipette only out of this small bottle.
Never handle the large Optima HNO3 bottle stored in the fridge unless you have been given
explicit permission. Your aliquot can be stored at room temperature in a plastic bag.
0.065 Molar Optima HNO3: Weigh out 249.35 g DI H2O from the ELGA system in an acid-
cleaned 250 mL Nalgene bottle (don not forget to zero the scale with the empty bottle first).
Using a clean pipette tip, add 1036 mL 16 M Optima HNO3.
0.5 Molar Optima HNO3: Weigh out 242.5 g DI H2O from the ELGA system in an acid
cleaned 250 mL Nalgene bottle. Using a clean pipette tip, add 7750 mL 16 M Optima HNO3.





























