Embed Size (px)
Citation preview

Plant Physiol. (1975) 56, 579-583
Enzymes of Carbohydrate Metabolism in the DevelopingRice Grain
Received for publication March 24, 1975 and in revised form June 9, 1975
CONSUELO M. PEREZ, ALICIA A. PERDON, ADORACION P. RESURRECCION, RuTH M. VILLAREAL,AND BIENVENIDO 0. JULIANODepartment of Chemistry, International Rice Research Institute, Los Baiios, Laguna, Philippines
ABSTRACT
The levels of reducing and nonreducing sugars, starch,soluble protein, and selected enzymes involved in the me-tabolism of sucrose, glucose-l-P, and glucose nucleotideswere assayed in dehulled developing rice grains (Oryzasativa L. line IR1541-76-3) during the first 3 weeks afterflowering. The level of reducing sugars in the grain washighest 5 to 6 days after flowering. The level of nonreducingsugars and the rate of starch accumulation were maximum11 to 12 days after flowering, when the level of soluble pro-tein was also the highest. The activities of bound and freeinvertase, sucrose-UDP and sucrose-ADP glucosyltrans-ferases, hexokinase, phosphoglucomutase, nucleoside di-phosphokinase, and UDP-glucose and ADP-glucose pyro-phosphorylases were high throughout starch deposition,and were maximum, except for nucleoside diphosphokinasewhich did not increase in activity, between 8 and 18 daysafter flowering. Soluble primed phosphorylase and ADPglucose-a-glucosyltransferase (starch synthetase) wereboth present during starch accumulation. Phosphorylaseactivity was at least 2-fold that of soluble starch synthetasebut the synthetase followed more closely the rate of starchaccumulation in the grain. The activity of starch synthetasebound to the starch granule also increased progressivelywith increased starch content of the grain.
Changes in the activities of enzymes involved in the conversionof sucrose into starch have been analyzed during grain develop-ment in corn (26, 35) and barley (5-7), but not in rice. Akazawa(1) suggested that during starch synthesis in rice, sucrose is uti-lized by way of sucrose-UDP (ADP) glucosyltransferase. In corn(35), invertase has been proposed as the enzyme that metabolizessucrose during the early stage of starch synthesis, while in barley(6) the enzyme proposed is sucrose-UDP glucosyltransferase.The presence of various intermediates in the transformation offructose to glucose-l-P have been verified in the developing ricegrain by Murata et al. (24). Although Murata et al. (24) couldnot detect a transglucosylation reaction which converts UDPglucose to ADP glucose in rice, Baxter and Duffus (6) suggestedthat, in developing barley, the UDP glucose formed from sucroseis first converted to glucose-l-P by UDP glucose pyrophosphoryl-ase and then the glucose-l-P is converted to ADP glucose byADP glucose pyrophosphorylase (Fig. 1). Nucleoside diphospho-kinase has been detected in developing barley grain (6).ADP glucose-a-glucan-4-a-glucosyltransferase (starch syn-
thetase) is now generally considered to be the major pathway ofstarch synthesis instead of phosphorylase (30), although some invivo studies favor phosphorylase (4, 10). Recently, isozymes ofstarch synthetase (9, 30) and phosphorylase (7-10, 35) which donot require a primer have been identified in developing starchgranules and have been implicated with the initial synthesis ofstarch directly from ADP glucose or glucose-i-P. Although onlyUDP glucose starch synthetase and phosphorylase are reportedto be present during early starch synthesis in the developing grainof barley (6) and corn (35), Ozbun et al. (26) found adequatelevels ofADP glucose starch synthetase in the same corn samples.The changes in reducing and nonreducing sugars, starch, solu-
ble protein, and enzymes involved in the conversion of sucroseto starch in the developing rice grain (line IR1541-76-3) duringthe period of starch accumulation were investigated in this study.It complements a previous study on developing IR8 rice grain(3), in which only enzymes of starch metabolism were assayed.
MATERIALS AND METHODS
Developing rice grains (Oryza sativa L. line IR1541-76-3[IR24/TKM6]) were harvested from the experimental field ofthe Institute during the first 3 weeks after flowering in the 1974dry season (mean temperature, 28 C). The grains were threshedand classified on the basis of the size of the caryopsis into 1 to 3days, 5 to 6 days (early milky), 8 to 9 days (midmilky), 11 to 12days (late milky), 17 to 18 days (dough), and 21 days (yellow)after flowering. They were then frozen in Dry Ice and storedat -20 C until used. Prior to analysis, the grains were classifiedand dehulled manually at 0 C. Duplicate batches of grains wereweighed and analyzed at each developmental stage. Data weresubjected to analysis of variance and LSD (5%) values werecalculated.
Chemical Analysis. Total soluble sugars were extracted byhomogenizing the dehulled grains with hot 80%r (v/v) ethanol,and the starch was extracted from the residue with 52%o HC104.The carbohydrate concentration was determined by the anthronemethod (21), and the reducing sugar was assayed on the ethanolextract by Nelson's arsenomolybdate method (13). Soluble pro-tein was determined colorimetrically by the method of Lowry etal. (20). Rate of starch accumulation per grain in vivo was calcu-lated for each sampling interval from the starch content data.The mean rate of starch accumulation per sampling time repre-sented the mean of the rates of starch accumulation before andafter the sampling period.Enzyme Preparation. A known number of dehulled grains was
homogenized at 0 C in a mortar and pestle with 10 mm tris-maleate buffer, pH 7, containing 1 mM dithiothreitol. The ho-mogenate was centrifuged at 30,000g for 30 min at 0 C and thesupernatant liquid was decanted. The residue was washed threetimes with the extracting buffer and centrifuged as before. The
579
www.plantphysiol.orgon June 11, 2018 - Published by Downloaded from Copyright © 1975 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon June 11, 2018 - Published by Downloaded from Copyright © 1975 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon June 11, 2018 - Published by Downloaded from Copyright © 1975 American Society of Plant Biologists. All rights reserved.

50PEFALPlant Physiol. Vol. 56, 1975
enzymes were assayed in the combined extract plus washings, ex-cept for soluble invertase and soluble sucrose-UDP (ADP) glu-cosyltransferase which were assayed in the dialyzed extracts. Forthe soluble starch synthetase, extraction and washing periodswere extended to 30 mmn each, since preliminary trials showedthat 50% more activity based on the extract may be recoveredfrom the washings (3). Bound enzymes were assayed in thewashed residue. Starch synthetase bound to starch granules wasprepared from the washed residue according to Leloir et at. (18).
Phosphorylase was extracted from freeze-dried dehulled grains,since the activity of the enzyme decreases progressively withstorage of the frozen rice grain at -20 C (3).Enzyme Assays. Preliminary assays were done on all enzymes
to determine conditions where linear rates with respect to timeand substrate concentrations are obtained. All enzyme activitieswere expressed in nanomoles per minute per rice grain. Boundand soluble invertase and sucrose-ADP (UDP) glucosyltrans-ferase were determined by the method of Shannon and Dougherty(33). For invertase, a mixture of 200,l of 0.2M sodium acetatebuffer, pH 4.8, 100 Al of 60mm sucrose, and 100,l of enzymepreparation was incubated for 10 min at 35 C. For transferase,a mixture of 200Ml of 0.3M MES buffer, pH 6.5, 100Ml of5mM UDP or ADP, 100,ul of 0.2M sucrose, and 100,ul of enzymewas incubated for 15 min at 35 C. The assays were terminated byadding Nelson's copper reagent (13) and the concentration ofreducing sugars was determined.
Hexokinase was assayed by coupling the reaction with glucose-6-P dehydrogenase and following the change in absorbance ofNADP at 340 nm for 3 min at 25 C (35). The reaction mixturecontained 120 Mmoles HEPES buffer, pH 7.5, 10 Mmoles ofMgCl92, 10, moles of ATP, 3 Mmoles of glucose, 0.5 Amole ofNADP, 3 units of glucose-6-P dehydrogenase (Sigma ChemicalCo.), and 0.2 ml of enzyme in a total volume of 2 ml. Slightlylower hexokinase activity was obtained with fructose as substratewithout added glucose-P-isomerase. Assays with mixtures ofglucose and fructose showed no apparent competitive inhibitionbetween the two sugars.
Glucose-P isomerase activity was assayed by incubating thedialyzed enzyme preparation with glucose-6-P and measuring thefructose-6-P formation (12).
Phosphoglucomutase was assayed by the method of Tsai et al.(35). The reaction mixture (1 ml), containing 60,umoles ofHEPES buffer, pH 7.4, 5,umoles of MgCl2, 5,umoles of cysteine,4,umoles of glucose-1-P, and 0.1 ml of enzyme, was incubatedfor 5 min at 30 C and 1 ml of5 N H2SO4 was added to terminatethe reaction. Then 3 ml of water were added, and the sample washydrolyzed in a boiling water bath for 10 min. The Pi released
Table I. Changes in Soluble Sugars, Soluble Protein, and Starchin Developing Rice Grains, LineIR1541-76-3
Property per Grain
Days afterFlowering
Sugars StarchSoluble Starch accumu-proteinlain
reducing mean rate
dmolesnrnolesA,ntoles glucose
1-35-68-911-1217-1821
LSD (5%)
0.290.560.611.020.720.580.09
1.231.631.280.740.740.560.12
lg
3211619225420415919
glucose
4.216.629.548.575.379.44.4
gluicoselinin1.962.723.692.760.960.40
after acid hydrolysis was assayed by the method of Galanos andKapoulas (11). Phosphoglucomutase activity was expressed asnanomoles Pi incorporated into glucose-6-P.ADP (UDP) glucose pyrophosphorylase was measured by
the rate at which nucleotide diphospho-'4C-glucose was formedfrom 14C-glucose-1-P and ATP (UTP) according to Murata et at.(23). The reaction mixture (50 Ml) contained 7 Mmoles of glycyl-glycine buffer, pH 7.8, 0.5 Mmole of 14C-glucose-1-P, 1 Mmole ofATP (UTP), 2 Mmoles of MgCl2, 0.2,u mole of EDTA, 4, molesof NaF, and enzyme. After being incubated for 10min at 37 C,the mixture was immersed in a boiling water bath for 45 sec tostop the reaction. After centrifuging atlOOog for 10min, a 20-
Mul portion was developed by descending paper chromatographyon acid-washed Whatman no. 1 paper for 18 hr using a 95%ethanol-IM ammonium acetate mixture (15:6,v/v) as the de-veloper (27). The spots corresponding to ADP (UDP) glucosewere located with a Packard Model 460 chromatogram scanner,cut, eluted with 50% ethanol, and counted in a PackardTri-Carb liquid scintillation spectrometer.
Nucleoside diphosphokinase was assayed by coupling the re-action with the formation of 6-phosphogluconic acid from glucoseby the combined action of hexokinase and glucose-6-P dehydro-genase (22). ATP liberated from the nucleoside diphosphokinaseaction produces one NADPH in the glucose-6-P dehydrogenasereaction. The reaction mixture (3.8 ml) contained 2.4umoles oftris buffer, pH 7.8, 15,molesof MgCl2, 16.6,umoles of glucose,0.2 unit of hexokinase, 0.1 unit of glucose-6-P dehydrogenase, 3
,Mmoles of UTP, 0.15,u mole of ADP, 0.5,u mole of NADP, and1.1 ml of enzyme. After the mixture was incubated for 10 min at25 C, the absorbance was read at 340 nm. Controls were runwherein ADP was omitted from the incubation mixture. Thenucleoside diphosphokinase activity was expressed as nanomoles(NADPH) produced.Phosphorylase assay was essentially that of Baxter and Duffus
(6, 7). The incubation mixture consisted of 0.4 ml of 0.1M tris-maleate buffer, pH 6.2, with1 mm NaF, 0.3 ml of 2.5%- amylo-pectin, 0.1 ml of 0.05 N glucose-1-P, and 0.2 ml of enzyme. Aftershaking at 30 C for1 hr, the reaction was stopped by adding 0.5ml 5%7, trichloroacetic acid. Pi was determined by the method ofLowry and Lopez (19). Amylopectin was omitted from the assayof unprimed phosphorylase.
Starch synthetases were assayed according to the method ofBaun et al. (3). Unprimed starch synthetase was assayed byomitting the oyster glycogen primer. Synthetase activity was ex-pressed in nanomoles of glucose added to glycogen.
RESULTSChemical Changes. The highest level of nonreducing sugars in
the developing rice grain occurred 11 to 12 days after flowering,whereas the level of reducing sugars was highest5 to 6 days afterflowering. Baun et al. (3) did not observe a distinct peak forsoluble sugars in developing grains ofIR8 rice, whereas Akazawaet al. (2) found peak levels of both reducing and nonreducingsugars 9 days after flowering of Peta rice, one of the parents ofIR8. LaBerge et al. (17) also found the peak level of sucrose laterthan that of glucose and fructose in developing barley grains.The peak level of soluble protein occurred concomitantly with
that of the reducing sugars, 11 to 12 days after flowering (TableI). In developing IR8 rice grains, the peak of the soluble proteinlevel was found to occur about 10 days after flowering (3).
Starch synthesis in the rice grain was most rapid during thefirst 2 weeks after flowering, particularly at the midmilky stage(Table I). Based on starch content, the calculated weighted meanrate of starch accumulation was highest 8 to 9 days after flower-ing, but the biggest increase was between 8 to 9 days and 11 to12 days after flowering.
580 PEREZ ET AL.
www.plantphysiol.orgon June 11, 2018 - Published by Downloaded from Copyright © 1975 American Society of Plant Biologists. All rights reserved.
CARBOHYDRATE METABOLISM IN RICE GRAIN
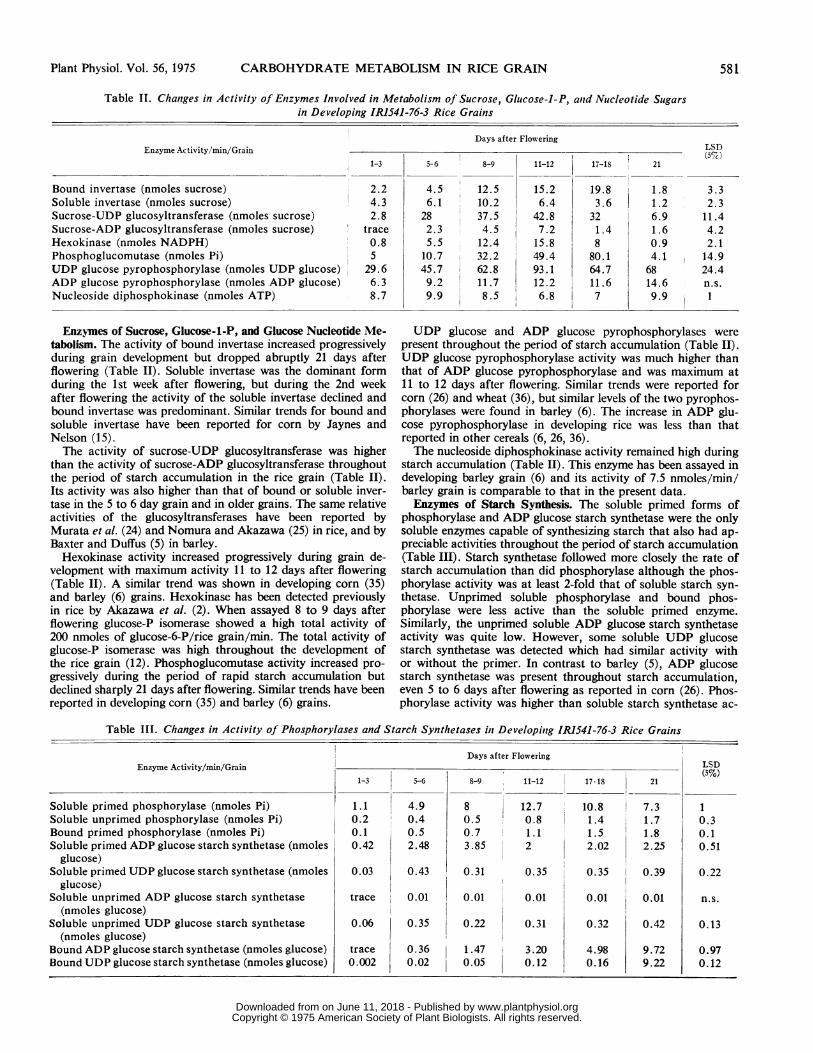
Table II. Changes in Activity of Enzymes Involved in Metabolism of Sucrose, Glucose-1-P, and Nucleotide Sugarsin Developing IRJ541-76-3 Rice Grains
Days after FloweringEnzyme Activity/min/Grain LSD
1-3 5-6 8-9 11-12 17-18 21
Bound invertase (nmoles sucrose) 2.2 4.5 12.5 15.2 19.8 1.8 3.3Soluble invertase (nmoles sucrose) 4.3 6.1 10.2 6.4 3.6 1.2 2.3Sucrose-UDP glucosyltransferase (nmoles sucrose) 2.8 28 37.5 42.8 32 6.9 11.4Sucrose-ADP glucosyltransferase (nmoles sucrose) trace 2.3 4.5 7.2 1.4 1.6 4.2Hexokinase (nmoles NADPH) 0.8 5.5 12.4 15.8 8 0.9 2.1Phosphoglucomutase (nmoles Pi) 5 10.7 32.2 49.4 80.1 4.1 14.9UDP glucose pyrophosphorylase (nmoles UDP glucose) 29.6 45.7 62.8 93.1 64.7 68 24.4ADP glucose pyrophosphorylase (nmoles ADP glucose) 6.3 9.2 11.7 12.2 11.6 14.6 n.s.Nucleoside diphosphokinase (nmoles ATP) 8.7 9.9 8.5 6.8 7 9.9 1
Enzymes of Sucrose, Glucose-1-P, and Glucose Nucleotide Me- UDP glucose and ADP glucose pyrophosphorylases weretabolism. The activity of bound invertase increased progressively present throughout the period of starch accumulation (Table II).during grain development but dropped abruptly 21 days after UDP glucose pyrophosphorylase activity was much higher thanflowering (Table II). Soluble invertase was the dominant form that of ADP glucose pyrophosphorylase and was maximum atduring the 1st week after flowering, but during the 2nd week 11 to 12 days after flowering. Similar trends were reported forafter flowering the activity of the soluble invertase declined and corn (26) and wheat (36), but similar levels of the two pyrophos-bound invertase was predominant. Similar trends for bound and phorylases were found in barley (6). The increase in ADP glu-soluble invertase have been reported for corn by Jaynes and cose pyrophosphorylase in developing rice was less than thatNelson (15). reported in other cereals (6, 26, 36).The activity of sucrose-UDP glucosyltransferase was higher The nucleoside diphosphokinase activity remained high during
than the activity of sucrose-ADP glucosyltransferase throughout starch accumulation (Table II). This enzyme has been assayed inthe period of starch accumulation in the rice grain (Table II). developing barley grain (6) and its activity of 7.5 nmoles/min/Its activity was also higher than that of bound or soluble inver- barley grain is comparable to that in the present data.tase in the 5 to 6 day grain and in older grains. The same relative Enzymes of Starch Synthesis. The soluble primed forms ofactivities of the glucosyltransferases have been reported by phosphorylase and ADP glucose starch synthetase were the onlyMurata et al. (24) and Nomura and Akazawa (25) in rice, and by soluble enzymes capable of synthesizing starch that also had ap-Baxter and Duffus (5) in barley. preciable activities throughout the period of starch accumulationHexokinase activity increased progressively during grain de- (Table III). Starch synthetase followed more closely the rate of
velopment with maximum activity 11 to 12 days after flowering starch accumulation than did phosphorylase although the phos-(Table II). A similar trend was shown in developing corn (35) phorylase activity was at least 2-fold that of soluble starch syn-and barley (6) grains. Hexokinase has been detected previously thetase. Unprimed soluble phosphorylase and bound phos-in rice by Akazawa et al. (2). When assayed 8 to 9 days after phorylase were less active than the soluble primed enzyme.flowering glucose-P isomerase showed a high total activity of Similarly, the unprimed soluble ADP glucose starch synthetase200 nmoles of glucose-6-P/rice grain/min. The total activity of activity was quite low. However, some soluble UDP glucoseglucose-P isomerase was high throughout the development of starch synthetase was detected which had similar activity withthe rice grain (12). Phosphoglucomutase activity increased pro- or without the primer. In contrast to barley (5), ADP glucosegressively during the period of rapid starch accumulation but starch synthetase was present throughout starch accumulation,declined sharply 21 days after flowering. Similar trends have been even 5 to 6 days after flowering as reported in corn (26). Phos-reported in developing corn (35) and barley (6) grains. phorylase activity was higher than soluble starch synthetase ac-
Table III. Changes in Activity of Phlosphorylases and Starch Synthetases in Developing IR1541-76-3 Rice Grains
Days after FloweringEnzyme Activity/min/Grain (5--)
1-3 5-6 8-9 11-12 17-18 21
Soluble primed phosphorylase (nmoles Pi) 1.1 4.9 8 12.7 10.8 7.3 1Soluble unprimed phosphorylase (nmoles Pi) 0.2 0.4 0.5 0.8 1.4 1.7 0.3Bound primed phosphorylase (nmoles Pi) 0.1 0.5 0.7 1.1 1.5 1.8 0.1Soluble primed ADP glucose starch synthetase (nmoles 0.42 2.48 3.85 2 2.02 2.25 0.51
glucose)Soluble primed UDP glucose starch synthetase (nmoles 0.03 0.43 0.31 0.35 0.35 0.39 0.22
glucose)Soluble unprimed ADP glucose starch synthetase trace 0.01 0.01 0.01 0.01 0.01 n.s.
(nmoles glucose)Soluble unprimed UDP glucose starch synthetase 0.06 0.35 0.22 0.31 0.32 0.42 0.13
(nmoles glucose)Bound ADP glucose starch synthetase (nmoles glucose) trace 0.36 1.47 3.20 4.98 9.72 0.97Bound UDP glucose starch synthetase (nmoles glucose) 0.002 0.02 0.05 0.12 0.16 9.22 0.12
Plant Physiol. Vol. 56, 1975 581
www.plantphysiol.orgon June 11, 2018 - Published by Downloaded from Copyright © 1975 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 56, 1975
tivity, as was observed in corn (26), and in contrast to the resultsof Baun etal. (3) withIR8 rice. Varietal differences may be in-volved since Pisigan (29) did not find unprimed soluble starchsynthetase in a crude extract from developingIR8 rice grains.The incorporation of glucose into glycogen from UDP glucose
by soluble starch synthetase was interesting since soluble starchsynthetase has been known to be very specific for ADP glucose.However, Tsai (34) also reported some activity of soluble starchsynthetase from corn towards UDP glucose.ADP glucose starch synthetase bound to the starch granule in-
creased progressively during grain development (TableIII). Itsspecificity to ADP glucose was higher than toward UDP glucoseexcept between1 and 3 days after flowering. The total starchsynthetase (soluble and bound) activity was lower than that ofphosphorylase except in the 21-day rice grain. The specificity forADP glucose of bound synthetase from theIR1541-76-3 ricegrain was greater than that fromIR8 rice (ADP glucose/UDPglucose of 2:1 (3) and 1:1 (28)), and developing barley grains(5). In developingIR8 rice grains, soluble starch synthetase wasonly a fraction of the bound synthetase even in the 7-day grain(3), whereas soluble synthetase was higher in IR1541-76-3 ricethan bound synthetase up to 8 to 9 days after flowering.
DISCUSSION
Sucrose-UDP glucosyltransferase probably plays a major rolein sucrose metabolism in the developing rice grain (Table II).Murata etal. (23) reported that the content ofUDP is consistentlylower than that of ADP in the ripening rice grain while the UDPglucose content is about three to five times higher than the con-tent of ADP glucose. In addition, UDP specifically inhibitssucrose-ADP glucose glucosyltransferase (24). Probably, bothsucrose-UDP and sucrose-ADP glucosyltransferases are involvedin the conversion of sucrose into nucleotide sugars, except at 1to 3 days after flowering when sucrose-ADP glucosyltransferaseactivity was low. Part of the UDP glucose is probably utilizedfor wall glucan formation during the period of cell enlargement.However, this period, which spans from 5 to 13 days afterflowering (14), does not exactly coincide with the activity curveof sucrose-UDP glucosyltransferase (TableII).The contribution of invertase in the initial step of sucrose me-
tabolism in the developing rice grain cannot be disregarded. Inaddition, the presence of bound invertase also suggests the pos-
UDP
UDP ?
ADP
Sucroe
I t _Fructose
\ _4~~~3ilucose CT' 2r-ructose-6-P
<ADP 14Glucose -5 -F *5 G\\coCO
FIG. 1. Sucrose starch conversion in the developing rice grain. 1:Invertase; 2: sucrose-UDP (ADP) glucosyltransferase (sucrose synthe-tase); 3: hexokinase; 4: glucose-P isomerase; 5: phosphoglucomutase;6: UDP (ADP) glucose pyrophosphorylase (synthetase); 7: nucleosidediphosphokinase; 8: phosphorylase; 9: ADP (UDP) glucose starchsynthetases.
sibility of sucrose hydrolysis during its movement into the endo-sperm as was reported in corn (33). Murata et al. (24) foundradioactive glucose and fructose after supplying radioactivesucrose to rice panicles, indicating that some hydrolysis of sucroseoccurred in the developing grain. However, Jenner (16) showedthat inversion was not necessary for the translocation of sucrosein developing wheat endosperm.The present study provides evidence that the levels of UDP
glucose pyrophosphorylase and ADP glucose pyrophosphorylasewere adequate to account for the conversion of UDP glucose toADP glucose, with glucose-i-P as intermediate (TableIt, Fig. 1)as was observed in barley. The amount of ADP glucose pyrophos-phorylase was adequate to account for all the starch producedfrom both the glucose and the fructose moieties of sucrose. Inaddition, the activity of nucleoside diphosphokinase was quitehigh throughout starch deposition.ADP glucose starch synthetase activity was adequate to ac-
count for starch accumulation in the developing rice grain, ex-cept in the1 to 3 day sample, where the activity of this enzymeis less than the rate of starch accumulation. Since starch granulesstart appearing in the rice endosperm only by the 4th day afterflowering (31, 32), most of this starch analyzed in the 1 to 3-daysample may be from the pericarp and the nucellus and outer-integument cells (32). During this time soluble phosphorylaseand UDP glucose starch synthetase were quite active. AlthoughADP glucose starch synthetase is probably the major enzyme ofnucleotide sugar starch synthetase in IR1541-76-3 rice, because ofthe poor specificity to UDP glucose, the UDP glucose systemprobably contributes substantially to starch synthesis, particu-larly if UDP glucose-ADP glucose conversion occurs in the de-veloping rice grain (Fig. 1). Direct comparison of in vitro enzymeactivities and in vivo rate of starch accumulation may not neces-sarily reflect the in vivo situation because of the limitations ofintracellular concentration of substrate, metal ions, and allostericeffectors.
LITERATURE CITED
1. AKAZAWA, T. 1972. Enzymes of rice. In: D. F. Houston, ed., Rice Chemistry andTechnology. American Association of Cereal Chemists, Inc., St. Paul, Minn.pp. 75-101.
2. AKAZAWA, T., T. MINAMIKAWA, AND T. MURATA. 1964. Enzymic mechanism ofstarch synthesis in ripening rice grains. Plant Physiol. 39: 371-378.
3. BAUN, L. C., E. P. PALMIANO, C. M. PEREZ, AND B. 0. JULIANO. 1970. Enzymes
of starch metabolism in the developing rice grain. Plant Physiol. 46: 429-434.4. BADENHUIZEN, N. P. 1973. Fundamental problems in the biosynthesis of starch
granules. Ann. N. Y. Acad. Sci. 210:11-16.5. BAXTER, E. D. AND C. M. DuFFUS. 1971. Starch synthetase in developing barley
amyloplast. Phytochemistry 10: 2641-2644.6. BAXTER, E. D. AND C. M. DUFFUS. 1973. Enzymes of carbohydrate metabolism
in developing Hordeum distichum grain. Phytochemistry 12: 1923-1928.7. BAXTER, E. D. AND C. M. DuFFus. 1973. Phosphorylase activity in relation to
starch synthesis in developing Hordeum distichum grain. Phytochemistry 12:2321-2330.
8. BURR, B. AND 0. E. NELSON. 1973. The phosphorylases of developing maize seeds.Ann. N. Y. Acad. Sci. 210:129-138.
9. Fox,J., L. D. KENNEDY,J. S. HAWKER,J. L. OZBUN, E. GREENBERG, C. LAM-
MEL, AND J. PREISS. 1973. De noro synthesis of bacterial glycogen and plant
starch by ADPG:a-glucan 4-glucosyl transferase. Ann. N. Y. Acad. Sci. 210:90-103.
10. FRYDMAN, R. B. AND E. SLABNIK. 1973. The role of phosphorylase in starch bio-synthesis. Ann. N. Y. Acad. Sci. 210:153-169.
11. GALANOS, D. S. AND V. M. KAPOULAS. 1966. Arapid method for the determinationof organic nitrogen and phosphorus based on a single perchloric acid digestion.Anal. Chim. Acta 34: 360-366.
12. GENTINETTA, E. AND F. SALAMINI. 1973. Phosphoglucose isomerase I and II indeveloping and germinating rice endosperm. Riso 22:303-311.
13. HODGE, J. E. AND B. T. HOFREITER. 1962. Determination of reducing sugars andcarbohydrates. In: R. L. Whistler and M. L. Wolfrom, eds., Methods in Carbo-hydrate Chemistry, Vol.1. Academic Press, New York, pp. 380-394.
14. HOSHIKAWA, K. 1967. Studies on the development of endosperm in rice. 2. Proc-ess of endosperm tissue formation with special reference to the enlargement ofcells. Nippon Sakumotsu Gakkai Kiji 36: 203-209.
15. JAYNEs, T. A. AND 0. E. NELSON. 1971. Invertase activity in normal and mutantmaize endosperm during development. Plant Physiol. 47: 623-628.
582 PEREZ ET AL.
A-
www.plantphysiol.orgon June 11, 2018 - Published by Downloaded from Copyright © 1975 American Society of Plant Biologists. All rights reserved.

CARBOHYDRATE METABOLISM IN RICE GRAIN
16. JENNER, C. F. 1974. An investigation of the association between the hydrolysis ofsucrose and its absorption by grains of wheat. Aust. J. Plant Physiol. 1: 319-329.
17. LABERGE, D. E., A. W. MACGREGOR, AND W. 0. S. MEREDITH. 1973. Changesin the free sugar content of barley kernels during maturation. J. Inst. Brew. 79:471-477.
18. LELOIR, L. F., M. A. R. DE FEKETE, AND C. E. CARDINI. 1961. Starch and oligo-saccharide synthesis from uridine diphosphate glucose. J. Biol. Chem. 236:636-641.
19. LOWRY. 0. H. AND J. A. LOPEZ. 1946. The determination of inorganic phosphatein the presence of labile phosphate esters. J. Biol. Chem. 162: 421-428.
20. LowRy, 0. H., N. J. ROsEBROUGH, A. L. FARR, AND R. J. RANDALL. 1951. Pro-tein measurement with the Folin phenol reagent. J. Biol. Chem. 193: 265-290.
21. MCCREADY, R. M.,J. GUGGOLZ, V. SILVEIRA, AND H. S. OwENS. 1950. Determina-tion of starch and amylose in vegetables. Anal. Chem. 22: 1156-1158.
22. MILLs, G. T. AND E. F. B. SmITH. 1965. Uridine diphosphoglucose, uridine di-phosphogalactose, uridine triphosphate and uridine diphosphoglucuronic acid.In: H. U. Bergmeyer, ed., Methods of Enzymatic Analysis. Academic Press,New York, pp. 581-595.
23. NIURATA, T., T. MINAMIKAWA, T. AKAZAWA, AND T. SUGIYAMA. 1964. Isolation ofadenosine diphosphate glucose from ripening rice grains and its enzymic syn-thesis. Arch. Biochem. Biophys. 106: 371-378.
24. MURATA, T., T. SUGIYAMA, T. MINAMIKAWA, AND T. AKAZAWA. 1966. Enzymicmechanism of starch synthesis in ripening rice grains. III. Mechanism of thesucrose-starch conversion. Arch. Biochem. Biophys. 113: 34-44.
25. NOMURA, T. AND T. AKAZAWA. 1973. Enzymic mechanism of starch synthesis inripening rice grains. VII. Purification and enzymic properties of sucrose synthe-tase. Arch. Biochem. Biophys. 156: 644-652.
26. OZBUN, J. L., J. S. HAWKER, E. GREENBERG, C. LAMMEL, J. PREss, AND E. Y.LEE. 1973. Starch synthetase, phosphorylase, ADP glucose pyrophosphorylase,
583
and UDP glucose pyrophosphorylase in developing maize kernels. Plant Phys-iol. 51: 1-5.
27. PALADINI, A. C. AND L. F. LELOIR. 1952. Studies on uridine diphosphate glucose.Biochem. J. 51: 426-430.
28. PERDON, A. A., E. J. DEL ROSARIO, AND B. 0. JULiANO. 1975. Solubilization ofstarch synthetase bound to Oryza sativa starch granules. Phytochemistry. Inpress.
29. PIsIGAN, R. A., JR. 1974. Isozymes of soluble starch synthetase from developingrice grains. Master's thesis. University of the Philippines, Los Baiios.
30. PREIsS, J., J. L. OZBuN, J. S. HAWKER, E. GREENBERG, AND C. LAMMEL. 1973.ADPG synthetase and ADPG-a-glucan 4-glucosyl transferase: enzymes in-volved in bacterial glycogen and plant starch synthesis. Ann. N. Y. Acad. Sci.210: 265-278.
31. ROSARIO, A. R. DEL, V. P. BRIONEs, A. J. VIDAL, AND B. 0. JULIANo. 1968. Com-position and endosperm structure of developing and mature rice kernel. CerealChem. 45: 225-335.
32. SATO, K. 1964. Studies on starch contained in the tissues of rice plant. 10. Starchdistribution in the tissues of flower and caryopsis with their development ofgrowth. Nippon Sakumotsu Gakkai Kiji 33: 29-34.
33. SHANNON, J. C. AND C. T. DOUGHERTY. 1972. Movement of 14C-labeled assimilatesinto kernels of Zea moys. I. Invertase activity of the pedicel and placento-chalazal tissues. Plant Physiol. 49: 203-206.
34. TSAI, C. Y. 1974. The function of the waxy locus in starch synthesis in maize endo-sperm. Biochem. Genet. 11: 82-96.
35. TSAI, C. Y., F. SALAMINI, AND 0. E. NELSON. 1970. Enzymes of carbohydratemetabolism in the developing endosperm of maize. Plant Physiol. 46: 299-306.
36. TURNER, J. F. 1969. Starch synthesis and changes in uridine diphosphate glucosepyrophosphorylase and adenosine diphosphate glucose pyrophosphorylase inthe developing wheat grain. Aust. J. Biol. Sci. 22: 1321-1327.
Plant Physiol. Vol. 56, 1975
www.plantphysiol.orgon June 11, 2018 - Published by Downloaded from Copyright © 1975 American Society of Plant Biologists. All rights reserved.

CORRECTIONSVol. 55: 1067-1070. 1975
Nobel, Park S., Lawrence J. Zaragoza, and William K. Smith.Relation between Mesophyll Surface Area, PhotosyntheticRate, and Illumination Level during Development for Leavesof Plectranthus parviflorus Henckel.
Page 1070, column 1, paragraph 1, lines 13 to 16 should becorrected to read: Figure 4 indicates a linear relation betweenAm'S/A and leaf thickness for mature third-node leaves ofPlectranthus parviflorus up to nearly 700 Am thick, viz.,Am's/A = 0.044 x (leaf thickness in Am: 40).
Vol. 56: 579-583. 1975
Perez, Consuelo M., Alicia A. Perdon, Adoracion P. Resurrec-cion, Ruth M. Villareal, and Bienvenido 0. Juliano. Enzymesof Carbohydrate Metabolism in the Developing Rice Grain.
Page 581, Table III, column 7, the activity of bound UDP glu-cose starch synthetase 21 days after flowering should be cor-rected to read: 0.
Vol. 57: 41-46. 1976
Rose, Ray and John Possingham. Chloroplast Growth and Rep-lication in Germinating Spinach Cotyledons following Massive-y-Irradiation of the Seed.
Page 41, line 3 should be corrected to read: Received for publi-cation August 11, 1975 and in revised form September 19,1975.
Vol. 57: 344-346. 1976
Zobel, Richard W., Peter Del Tredici, and John G. Torrey.Method for Growing Plants Aeroponically.
Page 344, column 2, line 5 should be corrected to read: as fol-lows: fractional horsepower motor, type NS1-13, with 1/40hp,.
602





























