Embed Size (px)
Citation preview

Plant Physiol. (1982) 70, 388-3920032-0889/82/70/0388/05/$00.50/0
Enzymes for Acetaldehyde and Ethanol Formation in LegumeNodules
Received for publication December 21, 1981 and in revised form March 8, 1982
SHIGEYUKI TAJIMA' AND THOMAS A. LARUEBoyce Thompson Institutefor Plant Research, Ithaca, New York 14853
ABSTRACT
Soybean (Glycine max L. var. Wilkin) nodules contain acetaldehyde andethanol. The cytosol of soybean and other legume nodules contains pyruvicdecarboxylase (EC 4.1.1.1) and alcohol dehydrogenase (EC 1.1.1.1). Someof the properties of these enzymes from soybean nodules are described.Their presence indicates that in the microaerobic nodule cytosol somecarbohydrate is metabolized by fermentative pathways like those in theroots of flood-tolerant plants.
Many cells, ranging from bacteria to some in higher plants andanimals, can metabolize carbon substrates by aerobic or anaerobicpathways. First described in yeast by Pasteur, the phenomenon isdue to the integration of two multienzyme systems, with a regu-latory control determining the contribution of the two pathways.In many organisms, the 'branch point' for the two pathways ispyruvic acid. In aerobic metabolism, it is oxidatively decarboxyl-ated to acetyl-CoA which is then oxidized in the citric acid cycle.Under anaerobic conditions, fermentation pathways may yieldthree carbon products (alanine or lactic acid), two carbon products(acetaldehyde, ethanol, or acetate), or four carbon products (malicacid) (Fig. 1). These compounds are reported products of anaer-obic metabolism in roots (13).
Because nitrogenase is denatured by 02, nitrogen fixing orga-nisms require a mechanism for maintaining a low PO2. The legumenodule apparently achieves this by using leghemoglobin, whichbinds strongly to 02 (1). The leghemoglobin aids the transfer of02 to the bacteroid, but keeps the free 02 concentration in thecytosol at approximately 10 nm (1).
DeVries et al. (4) suggested that because of its essentiallyanaerobic nature, the nodule cytosol might have an anaerobicmetabolism similar to that in roots of "flood-tolerant" plants.They demonstrated in pea nodule cytosol the presence of malicacid and high levels of PEP2 carboxylase (EC 4.1.1.31) and malatedehydrogenase (EC 1.1.1.37). The Rhizobiumjaponicum bacteroidsisolated from soybean nodules metabolize ethanol and acetalde-hyde (15). In this paper, we present evidence that the soybeannodule contains ethanol and acetaldehyde and that the plantcytosol contains the enzymes necessary for the production of thesecompounds.
' Permanent address: Faculty of Agriculture, Kagawa University, Miki-cho, Kita-Gun, Kagawa, Japan.
2 Abbreviation: pCMB, p-chloromercuribenzoate; PDC, pyruvate de-carboxylase; PEP, phosphoenol pyruvate; TPP, thiamine pyrophosphate;ADH, alcohol dehydrogenase.
MATERIALS AND METHODS
Chemicals. Acetaldehyde was obtained from Eastman KodakCo., Rochester, NY, and ethanol was obtained from U.S. Indus-tries Chemical Co., Louisville, KY. Both were distilled for theenzyme assay. Mops, Mes, Hepes, lactic dehydrogenase, alcoholdehydrogenase, aldehyde dehydrogenase, NAD, NADH, NADP,mercaptoethanol, DTT, Polyclar AT, pCMB, and phenylmethyl-sulfonylfluoride were purchased from Sigma. Calcium phosphategel was prepared by the method of Siegelman et al. (18). DEAE-Sephacel, Sephadex G-25, G-150 were products of Pharmacia(Piscataway, NJ). Chromosorb 104 and Porapak N were obtainedfrom Applied Science Laboratories Inc., State College, PA. Arand N2 gas (prepurified grade) were purchased from Union Car-bide Corporation, Linde Division, New York, NY. All otherchemicals were analytical grade. 1-14C sodium pyruvate and 3-14Csodium pyruvate were obtained from New England Nuclear.
Biological Materials. Seeds were obtained commercially ofGlycine max var. Wilkin; Viciafaba var. Diana; Phaseolus limensis,Pisum sativum var. Alaska; Lupinus albus, Trifolium repens, andVigna unguiculata var. Chinese Red. Plants were grown in green-houses in Turface (IMC Corp., Havelock, Ontario) with CaCO3added to keep the pH near neutral. They were watered two tothree times daily and supplied with nutrient solution three timesweekly. The nutrient was that described by Evans et al. (6), exceptthat iron sequestrine was increased 10-fold. Seeds were inoculatedwith appropriate commercial inocula (Nitragin Co., Milwaukee,WI). Nodules were collected from 5- to 6-week-old soybean plantswhich were flowering or in early stage of pod fill. Nodules werecollected from other legumes at flowering.To estimate nitrogenase, detached soybean nodules were
weighed, sealed in a 100-ml serum bottle and 10 ml of gas phasereplaced by 10 ml of C2H2. After 10 min, l-ml gas samples weretaken for C2H4 measurement by flame ionization detector aftergas CG on Porapak N. The nodules were frozen in liquid N2 forsubsequent analysis of pyruvic decarboxylase and alcohol dehy-drogenase.
For preparation of nodule cytosol, bacteroid extract and rootextract, all buffers were thoroughly sparged with N2, and alltreatments were performed under a continuous stream of N2. Forcentrifugation, sealed N2-flushed tubes were used.Nodule cytosol and bacteroids were prepared from freshly
harvested nodules. Two grams of nodules were ground in a chilledmortar with a pestle with 10-ml grinding medium (200 mm K-phosphate [pH 7.5], containing 0.3 M sucrose, 5 mM DTT, I mMMgCl2, and 0.4 mm EDTA) and 0.6 g of acid-washed insolublePVP (Polyclar AT). The homogenate was squeezed through fourlayers of cheesecloth. The nodule homogenate was centrifuged for30 min at 16,000g and the supernatant used as nodule cytosol.
Bacteroids were prepared from the homogenate according tothe method reported by Peterson and LaRue (15), except thebuffer system was the grinding medium described above. The finalbacteroid fraction was sonicated 10 times for 10 s each time. The
388 www.plantphysiol.orgon December 28, 2019 - Published by Downloaded from Copyright © 1982 American Society of Plant Biologists. All rights reserved.

ALCOHOL AND ACETALDEHYDE IN NODULES
Sucrose
ADP ATPi
PEP
Lactate
NAD
-NADH
Co2
Ethanol
NAD
1 NADH
J....-. AcetaldehydeKu
NAD
< NADH
INADHAcetate
NAD
Malate
FIG. 1. Some possible pathways of carbon metabolism under aerobicor anaerobic conditions. I, Pyruvate decarboxylase; 2, alcohol dehydrog-enase; 3, lactate dehydrogenase; 4, PEP carboxylase; 5, malate dehydrog-enase; 6, NADP-specific malic enzyme; 7, aldehyde dehydrogenase; 8,pyruvate kinase.
supematant after centrifugation (30 min at 16,000g) was used as
the bacteroid extract.To obtain root extract, 2 g of non-nodulated roots were homog-
enized by the Polytron with 10 ml of the grinding medium lackingsucrose. The supernatant after centrifugation (30 min at 16,000g)was used for assays.The extracts were passed through a Sephadex G-25 column (1.2
x 30 cm) equilibrated with the grinding medium, immediatelybefore enzyme assays.
Partial Purification of Pyruvate Decarboxylase. Soybean nod-ules (100 g) frozen in liquid N2 and stored at -80°C were
homogenized by the Polytron with 500 ml ofthe grinding medium,and the plant cytosol fraction obtained. All buffers in the purifi-cation steps described below contained 1 mm MgCl2, 10 ,UM TPP,1 mm DTT, 0.1 mm phenylmethylsulfonylfluoride, and 5% glyc-erol. All steps were performed at 4°C, and the manipulations insteps 1 to 3 (see Table IV) were performed anaerobically under astream of N2 (17).
Solid ammonium sulfate was added to the cytosol fraction to201% saturation. After centrifugation (10 min, 16,000g) the super-natant was brought to 40% saturation. The resulting precipitatewas collected, dissolved in buffer, and passed through a SephadexG-25 column (5.0 x 50 cm) equilibrated with 30 mm K-phosphate(pH 7.5). The protein fraction was applied to a DEAE-Sephacelcolumn (2.6 x 30 cm) equilibrated with 20 mm K-phosphate (pH7.5), and eluted with a linear gradient of 250 ml 20 mm K-phosphate (pH 7.5) and 200 mm K-phosphate (pH 7.5). Theenzymically active fractions, contained within a single peak, were
pooled and brought to 50%o saturation by adding solid ammoniumsulfate. The precipitate was dissolved in buffer and passed througha Sephadex G-25 column (2.6 x 30 cm) equilibrated with 20 mmK-phosphate. The protein fraction was loaded on a calciumphosphate gel column (2.6 x 40 cm) equilibrated with 20 mm K-phosphate, and eluted with a linear gradient of 250 ml, 20 mm K-phosphate (pH 7.5) and 250 ml 400 mm K-phosphate (pH 7.5).The active fractions were pooled and concentrated by ultrafiltra-tion with an Amicon B 15 concentrator (Amicon Corporation,Lexington, MA). The concentrated solution was applied to a
Sephadex G- 150 column (2.6 x 40 cm) equilibrated with the 20mm K-phosphate (pH 7.5), and eluted with the same buffer. Theenzyme fractions were pooled and concentrated with the AmiconB 15 concentrator.The partly purified enzyme was stored in the form of a precip-
itate in 60% ammonium sulfate solution at -20°C. In this condi-tion the enzyme kept 60% of its activity after 1 month.
Biochemical Enzyme Assays. All enzyme assays were performedat room temperature (22°C). A Gilford 250 spectrophotometerwas used to determine oxidation or reduction of pyridine nucleo-tide at 340 nm. For determination of NADH degradation due toenzyme, an anaerobic cuvette was used to eliminate the effect ofNADH oxidase activity. NADH solutions were prepared imme-diately before use and all buffers for anaerobic enzyme assayswere sparged with Ar. Protein was determined according to Lowryet al. (11) with BSA as standard.
Alcohol dehydrogenase (EC 1.1.1.1) was measured with theassay system of Cossins et al. (2): 100 mm ethanol, 1.26 mM NAD,50 mM Hepes (pH 8.7); or 20 mm acetaldehyde, 0.12 mM NADH,50 mM Mes (pH 6.5).PDC was assayed by the method of John and Greenway (8). In
this assay, added yeast alcohol dehydrogenase catalyzes the re-duction of acetaldehyde, and oxidation of NADH is measured at340 nm. The manometric assay of PDC (9) was performed byusing a Gilson Differential Respirometer to measure CO2.
NADP-specific malic enzyme (EC 1.1.1.40) was determined bythe method of Johnson et al. (9). Malate dehydrogenase wasdetermined with oxalacetic acid as substrate (16). Lactic dehy-drogenase (EC 1.1.1.27) was assayed by the method of Gutmannand Wablefeld (7).
Determination of Ethanol and Lactate Formation in NoduleExtracts. Frozen nodules were ground with cold extraction buffercontaining sucrose, centrifuged, and the supernatant passedthrough Sephadex G-25. The nodule cytosol was incubated in atotal volume of 2 ml with 2 ,umol NADH and the reaction startedby adding 200 ,umol pyruvate. The oxidation of NADH wasmeasured by the change in A at 340 nm. The reaction was stoppedby adding 0.5 ml 1 M HC104. Ethanol was determined by alcoholdehydrogenase (24) and lactic acid by lactic dehydrogenase (7).Gas Chromatographic Identification of Acetaldehyde and
Ethanol in Soybean Nodules. Freshly harvested nodules (2-3 g)were frozen in liquid N2 and ground in a mortar. A test tube (2.6x 10 cm) containing the fme powder was connected through avacuum line to a cold finger. Under a vacuum (50 mm Hg)volatiles from the fine powder were trapped by the cold fingerdipped in liquid N2. After keeping the cold finger for 5 min at-20°C, 1 ml of distilled H20 was injected into the cold fingerfollowed by shaking at 0°C. The water was transferred to a 20-mlchilled serum bottle and sealed with a Teflon-coated septum.(Weaton Scientific, Millville, NJ). After 10-min incubation at60°C, 0.5 ml of head-space sample was injected to a Varian 1400gas chromatograph equipped with a flame ionization detector. A3.0 x 3000 mm stainless steel column packed with Chromosorb104 or 3 x 1500 mm stainless steel column packed with PorapackN was used to separate acetaldehyde and ethanol. The columntemperatures were 150 and 140°C, respectively. For injection ofthe sample the syringes were heated to 1 10°C (5).Enzymic Determination of Acetaldehyde and Ethanol Content
in Nodules. Freshly harvested nodules were frozen in liquid N2,and ground in a mortar. Four to 5 grams of the fine powder wereshaken with 20 ml ice-chilled 0.6 M HC104, squeezed through twolayers of Miracloth (Chicopee Mills, Inc., New York, NY), cen-trifuged for 10 min at lO,OOOg in a capped tube. The supernatantwas neutralized to pH 6.0 to 7.0 with 4 M KOH solution, andcentrifuged again. The supernatant (15 ml) was distilled to yielda total of 6.0 ml in the collecting cylinder that contained 1.0 ml ofa buffer solution (21). For the acetaldehyde assay, the tip of thedistillation apparatus was immersed into a buffer solution thatcontained 0.1% sodium bisulfite, 1.25 mm NAD, 0.1 M KCI, and20 mmEDTA. Determination of acetaldehyde was performed byusing aldehyde dehydrogenase (12), and the fluorescence ofNADH formed was measured by using a Perkin-Elmer MPF 44B
389
www.plantphysiol.orgon December 28, 2019 - Published by Downloaded from Copyright © 1982 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 70, 1982
spectrophotometer (22). Excitation and emission wavelengths were350 and 460 nm, respectively.For ethanol assay, the buffer solution was 200 mm PPi (pH 9.0).
Ethanol content was determined by using alcohol dehydrogenase(24). Recoveries of acetaldehyde and ethanol after distillationwere 91% ± 3% and 80%o ± 10%, respectively. A blank was
obtained by using 4 ml of H20 instead of the nodule powder.Standard curves were prepared for each assay using aqueous
solutions of acetaldehyde and ethanol.That ethanol arises from pyruvic acid was confirmed using "C-
labeled Na pyruvate. A serum bottled contained I ,umol NADH,23 ,umol pyruvate containing 500,000 ± 2,500 cpm [1-14C] or 3-14C] sodium pyruvate, 5 nmol TPP in 3 ml of 0.1 M Tes, pH 6.0.The bottle contained a small tube containing KOH solution to
absorb CO2. The serum bottle was sparged 5 min with N2. Thenthe reaction was started by injecting nodule cytosol (0.8 mg
protein) immediately after its passage through Shephadex G25.After 15 min incubation, the reaction was terminated by inject-
ing 1 ml I M HC104 and incubated 15 min to trap CO2 in thealkali.The mixture was treated with Dowex- 1 x 8 to remove unreacted
pyruvate, neutralized with 4 M KOH, and the ethanol separatedby distillation. The radioactivity of the distillate and the trappedCO2 was as determined.
RESULTS
Organic volatiles from soybean nodules were analyzed by GC.Each of the two columns separated two peaks. The retention timesof these peaks were identical to those of acetaldehyde and ethanol.
In three separate batches of nodules, the amounts of acetalde-hyde determined enzymically were 53.4 nmol, 31.5, and 39.8nmol/g fresh weight nodules. The ethanol detected was 18.7, 25.1,and 36.2 nmol/g fresh weight nodules. Assuming that the noduleis 90% H20, the concentrations of the compounds approximate 36to 60 /.M and 20 to 40 ttM.
The enzymes for ethanol formation were mainly localized inthe nodule cytosol (Table I). Pyruvic decarboxylase was found innodule cytosol, but not in the bacteroid or root tissue. Alcoholdehydrogenase and malate dehydrogenase were found in bacte-roids, cytosol, and root extract, whereas lactic dehydrogenase was
absent. Weak activity of NADP-specific malic enzyme was de-tected in the nodule cytosol.
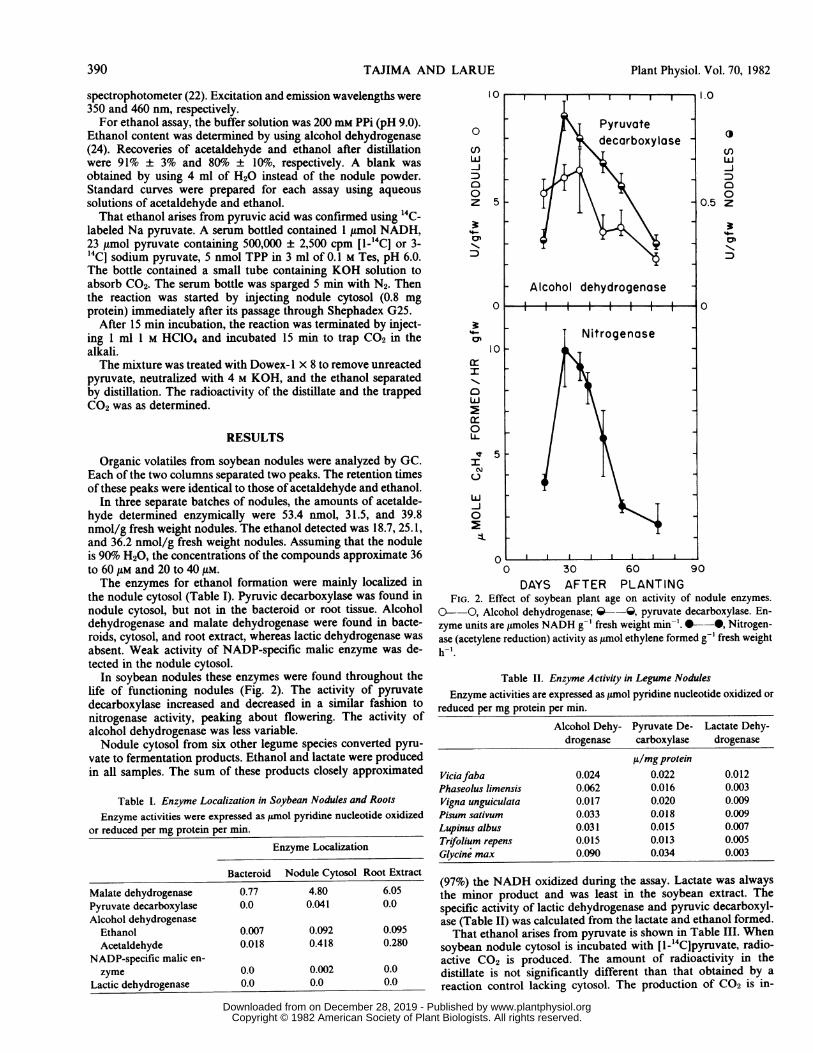
In soybean nodules these enzymes were found throughout thelife of functioning nodules (Fig. 2). The activity of pyruvatedecarboxylase increased and decreased in a similar fashion tonitrogenase activity, peaking about flowering. The activity ofalcohol dehydrogenase was less variable.Nodule cytosol from six other legume species converted pyru-
vate to fermentation products. Ethanol and lactate were producedin all samples. The sum of these products closely approximated
Table I. Enzyme Localization in Soybean Nodules and Roots
Enzyme activities were expressed as pmol pyridine nucleotide oxidizedor reduced per mg protein per min.
Enzyme Localization
Bacteroid Nodule Cytosol Root Extract
Malate dehydrogenase 0.77 4.80 6.05
Pyruvate decarboxylase 0.0 0.041 0.0
Alcohol dehydrogenaseEthanol 0.007 0.092 0.095
Acetaldehyde 0.018 0.418 0.280
NADP-specific malic en-
zyme 0.0 0.002 0.0
Lactic dehydrogenase 0.0 0.0 0.0
0
(I)
0
0
z
0'N-
'4-
I
CPN
0
0
IT
Lii
-J
0
Ut)w
-J
0
z
'4-
0 30 60 90
DAYS AFTER PLANTINGFIG. 2. Effect of soybean plant age on activity of nodule enzymes.
0- O, Alcohol dehydrogenase; Q--Q, pyruvate decarboxylase. En-zyme units are ytmoles NADH g-' fresh weight min-'.*--, Nitrogen-ase (acetylene reduction) activity as tmol ethylene formed g-' fresh weighth-'.
Table II. Enzyme Activity in Legume NodulesEnzyme activities are expressed as,umol pyridine nucleotide oxidized or
reduced per mg protein per min.
Alcohol Dehy- Pyruvate De- Lactate Dehy-drogenase carboxylase drogenase
,u/mgproteinViciafaba 0.024 0.022 0.012Phaseolus limensis 0.062 0.016 0.003Vigna unguiculata 0.017 0.020 0.009Pisum sativum 0.033 0.018 0.009Lupinus albus 0.031 0.015 0.007Tr!folium repens 0.015 0.013 0.005Glycine max 0.090 0.034 0.003
(97%) the NADH oxidized during the assay. Lactate was alwaysthe minor product and was least in the soybean extract. Thespecific activity of lactic dehydrogenase and pyruvic decarboxyl-ase (Table II) was calculated from the lactate and ethanol formed.That ethanol arises from pyruvate is shown in Table III. When
soybean nodule cytosol is incubated with [1_14C]pyruvate, radio-active CO2 is produced. The amount of radioactivity in thedistillate is not significantly different than that obtained by a
reaction control lacking cytosol. The production of CO2 is in-
390 TAJIMA AND LARUE
www.plantphysiol.orgon December 28, 2019 - Published by Downloaded from Copyright © 1982 American Society of Plant Biologists. All rights reserved.

ALCOHOL AND ACETALDEHYDE IN NODULES
Table III. Radioactivity of Productsfrom "C-Labeled Pyruvic Acid afterIncubation with Cytosolfrom Soybean Nodules
Substrate
Reaction Mixture [l- "C]Pyruvate [3-'4C]Pyruvate
CO2 Distillate CO2 Distillate
cpmComplete 12,570 654 1,017 17,855-NADH 10,390 <100 321 1,138+Pyrazole, 50 mm 11,440 <100 167 3,655-Extract 756 < 100 145 355+pCMB, 50 Mm 2,355 <100 215 1,615
hibited by 50 Mm pCMB, is not dependent on the presence ofNADH, and is not inhibited by pyrazole, an inhibitor of alcoholdehydrogenase.When [3-1'Clpyruvate is the substrate, radioactive ethanol is
produced, but relatively little radioactive CO2 (Table III). Theproduction of ethanol is dependent on the presence of NADH,and is reduced by inhibitors of pyruvate decarboxylase (pCMB)and of alcohol dehydrogenase (pyrazole).
Partial Purification of PDC. PDC was purified 136 fold (TableIV). The crude nodule extract lost 80%o of the enzyme activity in24 h if DTT and 0.3 M sucrose were not added. Even in thepresence of these compounds, 20%o of the activity was lost underair, and 80% under pure O2. Therefore, all the steps were per-formed in 3 d, and steps I to 3 were under a stream ofN2 gas. Thepresence of TPP was necessary to maintain the enzyme activity.More than 95% of the activity was precipitated between 20%o and40%o saturation of ammonium sulfate, and enzymically activefractions were obtained in a single peak in DEAE-Sephacel andcalcium phosphate column chromatography. The enzyme fractionwas eluted just after the fraction of void volume in SephadexG 150 column chromatography, suggesting the molecular weightof this enzyme is approximately 200,000 to 300,000.The fractions containing PDC in steps I to 4 also contained
alcohol dehydrogenase activity. The final preparation was freefrom that enzyme, whereas malate dehydrogenase activity was
detected in the preparation. NADP-specific malic enzyme andlactic dehydrogenase (pyruvate as substrate) were not detected inthe final preparation.A manometric assay of CO2 evolved by the preparation showed
92% of the activity that was assayed with alcohol dehydrogenase.The crude extracts could not be analyzed for PDC by a mano-
metric technique, nor by the evolution of 14CO2 from [1-14C]-pyruvate because of the presence of carboxylases.
Addition of 5% mannitol, glycerol, or sorbitol to the enzyme
preparation showed a stabilizing effect similar to that of sucrose.
Characteristics of PDC. All assays, unless otherwise indicated,
were performed by using alcohol dehydrogenase (8). Optimumrates were obtained only if the preparation was preincubated withNADH, and the reaction started by addition ofpyruvate. Initiationof the enzyme reaction by adding NADH as the final componentshowed only 20 to 30o of the maximum activity in the first 1 min.The manometric assay also showed a 40 to 50%o decrease of theenzyme activity when NADH was absent in the assay system.The Km value for pyruvate was 1.18 mm. The optimum pH was
at 6.0. The enzyme solution after calcium phosphate columnchromatography (2.10 ,umol NADH formed/mg protein * min)was precipitated by adding solid ammonium sulfate. The PDCprotein can be disassociated from TPP by mild alkalinity (19).After centrifugation, the pellet was dissolved in 20 mM Hepesbuffer (pH 8.2), containing 1 mm DTT and dialyzed by passingthrough Sephadex G-25 column (1.2 x 20 cm) equilibrated withthe same buffer. The enzyme lost 29% of its activity by alkalitreatment and only 7.6% of the initial activity remained afterpassage through the Sephadex.pCMB at 10 uM inhibited 82% of the enzyme activity (Table V),
and oxamic acid at 10 mm decreased the activity 75%. Acetalde-hyde, the product of the enzyme reaction, inhibited 88% of theactivity at 1 mM.
Characteristics of Alcohol Dehydrogenase from Cytosol. Whenethanol was used as substrate for the enzyme assay, ADH showedan optimum pH at 9.4, and the Km was 3.7 mm. Pyrazole at 0.05mm inhibited the enzyme activity 900o. In contrast, when acetal-dehyde was used as substrate for the enzyme assay, ADH showeda pH optimum at 7.0, and the Km value was calculated as 0.33mM. Pyrazole at 0.05 mm inhibited 82% of the enzyme activity.The characteristics are similar to those reported for alcohol de-hydrogenase in soybean seedlings (10).
DISCUSSION
Soybean nodules contained 31 to 53 nmol acetaldehyde and 19to 36 nmol ethanol per gram fresh weight. Straten and Schmidt(23) found these compounds and acetone in the gas phase whenexcised soybean nodules were waterlogged for 2 to 4 h. Theinhibitory effect of waterlogging on acetylene reduction was at-tributed to the toxic effect of ethanol. Sprent and Gallacher (20)confirmed that waterlogged nodules produced ethanol. They alsoreported ethanol production by severely water-stressed roots (45%loss in fresh weight), and suggested that imposed water stress caninduce anaerobiosis. Our finding of alcohol and acetaldehyde infreshly harvested nodules indicates that they are normal metabol-ities of the nodule, and not formed solely in response to prolongedstress.
Peterson and LaRue (15) found that alcohol and acetaldehydesupported respiration and acetylene reduction by R. japonicumbacteroids isolated from soybean nodules. Probably they are con-
verted to acetate, for bacteroids contain alcohol dehydrogenaseand a soluble aldehyde dehydrogenase (J. B. Peterson and T. A.
Table IV. Pur!fication of Pyruvate Decarboxylasefrom Soybean Nodules
Volume Proteina Total Activity Specific Activity Recovery
ml mg/ml pmol ACH/min lrmol ACH/min *mg %protein
1. Crude supernatant 480 7.50 (3600) 85.3 0.0237 1002. Ammonium sulfate precipi-
tation 110 5.00 (550) 78.7 0.143 92.33. DEAE Sephacel chromatog-
raphy 20.0 3.10 (62.0) 70.1 1.13 82.24. Calcium phosphate gel chro-
matography 10.3 1.88 (19.4) 48.9 2.52 57.35. Sephadex G 150 chromatog- 3.21
raphy 2.8 1.07 (3.0) 9.6 (135-fold) 11a Values in parentheses are total protein in milligrams.
391
www.plantphysiol.orgon December 28, 2019 - Published by Downloaded from Copyright © 1982 American Society of Plant Biologists. All rights reserved.

TAJIMA AND LARUE
Table V. Effect of Various Compounds on Activity on PyruvateDecarboxylasefrom Soybean Nodules
Each value was the mean triplicate assays, and performed with the fimalenzyme preparation after 10 min preincubation.
Compounds Concentration PDC activityAdded
mM %
PCMB 0.005 69EDTA 1.0 100Pyrazolea 1.0 100Oxamic acid 10.0 25Acetaldehydea 1.0 12
a For the assay with pyrazole or acetaldehyde, enzyme activity wasdetermined by the manometric method.
LaRue, unpublished results).Tracer experiments show that ethanol, and presumably acetal-
dehyde, arise from pyruvate. The enzymes for ethanol formationwere localized in the nodule cytosol (Table I), showing thatethanol formation is a function of the macro-symbiont. Pyruvatedecarboxylase and alcohol dehydrogenase are present throughoutthe development of the nodule. Because of the strong 02 bindingcapacity ofleghemoglobin, the nodule cytosol is likely to be micro-aerobic. In roots of many plants, alcohol dehydrogenase andpyruvate decarboxylase are formed when 02 availability decreases(13).
Alcohol dehydrogenase was also found in the nodule cytosol offaba bean, lima bean, cowpea, pea, lupin, and clover. This obser-vation suggests that fermentative pathways in nodules are widelydistributed in legumes. Although lactic dehydrogenase was barelydetectable in soybean nodules, in other legumes (Table II) theformation of lactate might be an alternate anaerobic pathway.
Pyruvate decarboxylase in nodule cytosol was inhibited by 10tLM pCMB and high concentrations of DTT were effective insustaining enzyme activity. These indicate that it is a typical SHenzyme. The optimum pH, Km for pyruvate, dependence on Mg2+and TPP, and inhibition by acetaldehyde are characteristics sim-ilar to data reported for PDC from other plants (3, 13, 14).John and Greenway (8) claimed that the control of alcohol
production in rice roots was at least partly mediated by theactivation ofpyruvate decarboxylase by NADH. Our preparationswere too unstable to test this. Studies on the regulation of theenzyme and its characteristics will require further work on thepreparation of the enzyme. Another problem will be to determinewhether the formation of acetaldehyde and ethanol represents amajor or minor pathway of carbohydrate metabolism in theunstressed nodule.
LITERATURE CITED
1. BERGESEN FJ, GL TURNER 1975 Leghaemoglobin and supply of 02 to nitrogenfixing root nodule bacteroids: studies of an experimental system with no gasphase. J Gen Microbiol 89: 31-47
2. COSSINs EA, LC KOPALA, B BLAWACKY, AM SPRONK 1968 Some properties ofa higher plant alcohol dehydrogenase. Phytochemistry 7: 1125-1134
3. DAVIEs DD, RJ CORBETT 1969 Glyoxylate decarboxylase activity in higherplants. Phytochemistry 8: 529-542
4. DEVRIEs GE, P IN 'T VELD, JW KIJNE 1980 Production oforganic acids in Pisumsativum root nodules as a result of oxygen stress. Plant Sci Lett 20: 115-123
5. ERIKSSON CJP, HW SIPPEL, OA FORSANDER 1977 The determination of acetal-dehyde in biological samples by head-space gas chromatography. Anal Bio-chem 80: 116-124
6. EVANS HJ, JB KOCH, R. KLUCAS 1972 Preparation of nitrogenase from nodulesand separation into components. Methods Enzymol 24: 470-476
7. GUTMANN I, AW WABLEFELD 1974 L-(+)-lactate. Determination with lactatedehydrogenase and NAD. In HU Bergmeyer, ed., Methods in EnzymaticAnalysis, 2nd Ed, Vol 3. Weinheim: Verlag Chemie, p. 1464
8. JOHN CD, H GREENWAY 1976 Alcohol fermentation and activity ofsome enzymesin rice roots under anaerobiosis. Aust J Plant Physiol 3: 325-336
9. JOHNSON HS, MD HATCH 1970 Properties and regulation of leaf nicotinamide-adenine dinucleotide phosphate-malate dehydrogenase and "malic" enzyme inplants with the C4-dicarboxylic acid pathway of photosynthesis. Biochem J119: 273-280
10. LEBLOVA S, E PERGLEROVA 1976 Soybean alcohol dehydrogenase. Phytochem-istry 15: 813-815
11. LOWRY OH, NJ ROSEBROUGH, AL FARR, RJ RANDALL 1951 Protein measure-ments with Folin-phenol reagent. J Biol Chem 193: 265-275
12. LUNDQUIST F 1974 Acetaldehyde. Determination with aldehyde dehydrogenase.In HU Bergmeyer, ed, Methods in Enzymatic Analysis, 2nd Ed, Vol 3.Weinheim: Verlag Chemie, pp 1509-1513
13. MCMANMOM M, RMM CRAWFORD 1971 A metabolic theory of flooding toler-ance: the significance of enzyme distribution and behavior. New Phytol 70:299-306
14. OBA K, I URITANI 1975 Purification and characterization of pyruvate decarbox-ylase from sweet potato roots. J Biochem 77: 1205-1213
15. PETERSON JB, TA LARUE 1981 Utilization of alcohols and aldehydes by soybeanbacteroids. Plant Physiol 68: 489-493
16. REEVES HC, R RABIN, WS WEGENER, SJ AJL 1971 Malate dehydrogenase. In JRNorris and DW Ribbons, eds, Methods in Microbiology, Vol. 6A. London,New York, Academic Press, pp 451-452
17. SAKAMI W 1961 Anaerobic gradient elution column chromatography. J BiolChem 236: 50
18. SIGELMAN HW, GA WIECZOREK, BC TURNER 1965 Preparation of calciumphosphate for protein chromatography. Anal Biochem 13: 402-404
19. SINGER TP 1955 a-carboxylase from wheat germ. Methods Enzymol 1: 465-47020. SPRENT JI, A GALLACHER 1975 Anaerobiosis in soybean root nodules under
water stress. Soil Biol Biochem 8: 317-32021. STOTs E 1943 A colorimetric determination of acetaldehyde in blood. J Biol
Chem 148: 585-59122. STOWELL AR, KE CROW, RM GREENWAY, RD BATT 1978 Determination of
acetaldehyde in blood using automated distillation and fluorometry. AnalBiochem 84: 384-392
23. STRATEN JV, EL SCHMIDT 1974 Volatile compounds produced during acetylenereduction by detached soybean nodules. Soil Biol Biochem 6: 347-351
24. WIrr I 1974 Determination of ethanol with a alcohol dehydrogenase and 3-acetyl-pyridine analogue of NADP. In HU Bergmeyer, ed, Methods in Enzy-matic Analysis, 2nd Ed, Vol 3. Weinheim: Verlag Chemie, pp 1502-1505
392 Plant Physiol. Vol. 70, 1982
www.plantphysiol.orgon December 28, 2019 - Published by Downloaded from Copyright © 1982 American Society of Plant Biologists. All rights reserved.





























