Embed Size (px)
Citation preview

133
Biochimica et Biophysica Acta, 497 (1977) 133--143 © Elsevier/North-Holland Biomedical Press
BBA 28181
ENZYME ACTIVITIES IN THE SHEEP PLACENTA DURING THE LAST THREE MONTHS OF PREGNANCY
ELIZABETH M. EDWARDS, J.M. RATTENBURY~ G. CAROLE E. VARNAM, USHA K. DHAND, MARJORIE K. JEACOCK and D.A.L. SHEPHERD
Department of Physiology and Biochemistry, University of Reading, Whiteknights, Reading, RG6 2AJ (U.K.)
Summary
In order to assess the extent to which metabolism within the sheep placenta may influence the transfer of metabolites between mother and foetus at differ- ent stages of gestation the activities of enzymes concerned with some aspects of carbohydrate, amino acid and ketone body metabolism were determined in placental cotyledons resected from ewes during the last three months of preg- nancy.
The activities of pyruvate kinase (EC 2.7.1.40), lactate dehydrogenase (EC 1.1.1.27), malate dehydrogenase (EC 1.1.1.37), ATP citrate (pro-3S)- lyase (EC 4.1.3.8), citrate (si)-synthase (EC 4.1.3.7), acetyl-CoA synthetase (EC 6.2.1.1), acetyl-CoA acetyltransferase (EC 2.3.1.9) and 3-keto acid CoA-transferase (EC 2.8.3.5) per gram wet weight cotyledon do not change during the period stud- ied. The activities of alanine aminotransferase (EC 2.6.1.2), aspartate amino- transferase (EC 2.6.1.1), isocitrate dehydrogenase (NADP ÷) (EC 1.1.1.42), ornithine-oxoacid aminotransferase (EC 2.6.1.13) and 3-hydroxybutyrate dehydrogenase (EC 1.1.1.30) show an increase in activity between the third and fourth months of pregnancy whilst the activities of arginase (EC 3.5.3.1) and possibly pyruvate carboxylase (EC 6.4.1.1) show an increase in activity between the fourth and final months of pregnancy. Ornithine decarboxylase (EC 4.1.1.17) activity declines to one tenth of its activity during this later period. The absence of detectable activities of phosphoenolpyruvate carboxy- kinase (EC 4.1.1.32) and ornithine carbamoyltransferase (EC 2.1.3.3) indicate that gluconeogenesis and urea synthesis from ammonia do no occur in the sheep placenta.
It appears that the ability of the placenta to metabolise several substrates is achieved by the time the placenta reaches its maximum size at approximately 90 days.

134
Introduction
Three stages are clearly discernable in the development of the sheep placen- ta, namely implantation, growth and maturation [1]. The implantation stage terminates at approximately the 50th day of pregnancy when the placental cotyledons have reached a fixed number. The second stage is characterised by rapid placental growth and terminates at approximately 90 days of gestation when the placental weight has reached its maximum [2,3]. The last stage which extends from 90 days to term may be called the maturation stage because although the placenta reaches its maximum size at three months, the foetus continues to grow throughout gestation and hence the funct ion of the placenta is greatly increased relative to its weight during the last two months of preg- nancy.
The placenta does not only function as a simple organ of exchange between the ewe and her foetus. Metabolism within the placenta may result in the release into the foetal circulation of substrates derived from maternal blood which have been chemically modified by the placenta. An established example of this is the production of fructose from maternal glucose by the ovine placen- ta [4] bu t other important metaboli te interconversions may exist. In addition the utilization by the placenta of substrates present in maternal blood may also prevent or limit their transfer to the foetus.
Any changes in the activities of placental enzymes during pregnancy may well indicate whether particular metabolic pathways are of greater or lesser sig- nificance as pregnancy proceeds. It is generally accepted that the units of a par- ticular enzyme activity measured in vitro reflect the amount of that particular enzymically active protein in the tissue. Changes in the amounts of certain enzyme proteins in tissues are a means whereby the rate of flux of metabolites through a metabolic pathway can be regulated. Such changes occur over rela- tively long periods of time and this means of metabolic regulation provides 'coarse' control of metabolism. In addition to the amount of enzyme protein present the rate of a reaction may also be regulated by the concentrat ion of reactants and in some cases by the concentrat ion of effectors. 'Fine' control of metabolic flux is believed to be brought about in this way. Even though measureable enzyme activity is bu t one factor indicating the rate of utilization of a particular substrate it has been argued that knowledge of enzyme activities may reveal clues about the metabolic potential of an organ during development [5]. In order to provide some indication of the importance of the metabolism of pyruvate, citrate, acetate, ketone bodies and certain amino acids in the placenta at different stages of pregnancy we have studied the activities of 18 enzymes concerned with their metabolism during the growth and maturat ion stages.
Material and Methods
Chemicals Tris, sodium pyruvate, oxaloacetic acid, monosodium phosphoenolypyruvate,
acetyl-CoA, ATP, ADP, NAD ÷, NADH, GSH, CoA, acetyl phosphate (dilithium salt}, carbamyl phosphate (dilithium salt) and all enzymes were obtained from

135
the Boehringer Corp. (London) Ltd. (Lewes, Sussex, U.K.); 2-mercaptoethanol, pyridoxal 5-phosphate, IDP and bovine serum albumin {crystallized and lyophil- ized) were from Sigma (London) Chemical Co. Ltd. (London, SW6, U.K.); o-aminobenzaldehyde was from K and K Laboratories Inc. (Plainview, Holly- wood, California, U.S.A.); 5,5'-dithiobis-(2-nitrobenzoic acid) and diketene were from Ralph N. Emanuel Ltd. (Alperton, Middlesex, U.K.); diacetyl monox- ime, hyamine hydroxide and dioxan-based scintillator were from Koch-Light Laboratories (Colnbrook, U.K.), and I~[1-'4C]ornithine monohydrochlor ide and NaH'4CO3 were from the Radiochemical Centre {Amersham, Bucks, U.K.). All other chemicals were obtained from BDH Chemicals Ltd. (Poole, Dorset, U.K.) and were of Analar grade whenever possible.
Sodium acetoacetate was prepared from methyl acetoacetate by the method of Hall [6] and was found to be at least 95% pure. Acetoacetyl-CoA was syn- thesised from CoA and diketene by the method of Simon and Shemin [7] and the concentrat ion of acetoacetyl-CoA was determined spectrophotometrical ly [81.
Animals and collection o f tissue Dorset Horn ewes tupped by Suffolk rams on known dates were purchased
from Reading University farms. The stage of gestation was confirmed by radio- logical examination at the Grassland Research Institute, Hurley. The ewes were housed and fed as described previously [9]. Immediately after foetuses had been delivered by Caesarian section [9] cotyledons were resected and placed in ice-cold 0.15 M KC1. Wedge shaped samples for the preparation of tissue slices and homogenates were cut through the cotyledons after they had been tr immed free of the cotyledonary branches of the umbilical vessels. No at tempt was made to separate maternal f rom foetal cotyledonary tissue since it was con- sidered that the maternal and foetal tissue together form the functional unit.
Measurement of urea production Slices of placental cotyledons (0.25 mm thick, 50--150 mg wet weight), cut
with a Stadie-Riggs microtome, were incubated at 37°C in 5.5 ml bicarbonate buffer [10] containing 18.2 mM ammonium chloride and 1.82 mM L-ornithine hydrochloride with 95% 02/5% CO2 as the gas phase. After 1 h, 0.5 ml 9 M perchloric acid was added to each of the incubation flasks and urea was deter- mined in the protein-free supernatants by condensation with diacetyl monox- ime [11]. Four replicate incubations were performed for each cotyledon, and blank incubations were carried out wi thout addition of substrate.
Measurement o f acetoacetate production Samples of tissue were homogenised in a high speed mixed emulsifier (Silver-
son Machines Ltd., London, S.E.1.) in 20 volumes of a medium consisting of 0.25 M sucrose, 1 mM 2-mercaptoethanol and 10 mM Tris adjusted to pH 7.4 with HC1. The homogenate was then sonicated with intermittent bursts using a 9 mm probe at 6--7 kcycles, 8--9 p amplitude for a total t ime of 45 s (100 W ultrasonic disintegrator, Measuring and Scientific Equipment Ltd. London, S.W.1.) and centrifuged at 38 000 × g for 30 min. These procedures were car- ried out at 0--4 ° C. Aliquots of supernatant were incubated with an acetyl-CoA

136
generating system for 10 min at 37°C in the medium described by Williamson et al. [12] for their assay of hydroxymethylglutaryl-CoA synthase. The aceto- acetate formed was determined enzymicaUy [13].
Measurement of enzyme activities Samples of the cotyledons to be assayed for pyruvate kinase (EC 2.7.1.40),
lactate dehydrogenase (EC 1.1.1.27), malate dehydrogenase (EC 1.1.1.37), aspartate aminotransferase (EC 2.6.1.1), alanine aminotransferase (EC 2.6.1.2), ATP citrate (pro-3S)-lyase (EC 4.1.3.8), citrate (si)-synthase (EC 4.1.3.7), iso- citrate dehydrogenase (NADP +) (EC 1.1.1.42) and acetyl-CoA synthase (EC 6.2.1.1) activities were homogenised in 5--10 volumes of 0.25 M sucrose. Tis- sue to be assayed for 3-hydroxybutyra te dehydrogenase (EC 1.1.1.30) activity was homogenised in 10 volumes of a medium adjusted to pH 7.4 containing 10 mM succinate, 1 mM EDTA, 0.6 mM ATP and 1 mM NAD ÷. The activities of these enzymes were determined in whole homogenates which had been sub- jected to sonication at 0°C except for ATP citrate (pro-3S)-lyase and acetyl- CoA synthetase, activities of which were determined in the supernatant after the homogenates had been centrifuged at 600 X g for 15 min to remove the nuclear fraction. This supernatant was frozen and thawed three times. Tissues to be assayed for pyruvate carboxylase (EC 6.4.1.1) and phosphoenolpyruvate carboxykinase (GTP) (EC 4.1.1.32) activities were homogenised in 5--10 vol- umes of a medium containing 0.2 M sucrose, 20 mM triethanolamine, 1 mM GSH and 1 mM EDTA adjusted to pH 7.5 with HC1. Aliquots of these homog- enates were freeze-dried and resuspended in glass distilled water prior to assay. The preparation of the homogenates was carried out at 0--4°C and the activi- ties of these 12 enzymes were assayed as described previously [9,14]. 3-Keto- acid CoA-transferase (EC 2.8.3.5) and ace ty l -CoA acetyltransferase (EC 2.3.1.9) activities were assayed in tissue supernatants prepared as for the deter- mination of the rate of acetoacetate production. The activity of 3-ketoacid CoA-transferase was determined by measuring the decrease in extinction at 310 nm resulting from the disappearance of the acetoacetyl-CoA-Mg 2÷ complex in the presence of succinate and iodoacetamide [15]. The activity of acetyl-CoA acetyltransferase was determined by measuring the decrease in extinction at 310 nm resulting from the disappearance of the acetoacetyl-CoA-Mg 2÷ complex in the presence of CoA [12]. Tissue to be assayed for arginase (EC 3.5.3.1) activity was homogenised in 20 volumes of a medium containing 0.16 M sodi- um chloride and 36 mM manganous sulphate in a Vortex homogeniser (Mea- suring & Scientific Equipment Ltd. London, S.W.1) at 0--4°C. The tissue homogenate was activated by incubation at 37°C for 30 min. Aliquots were then added to a solution containing 0.2 M L-arginine hydrochloride and 16 mM glycine adjusted to pH 9.45 with NaOH. The reaction was stopped by addi- tion of perchloric acid. Any urea in the supernatant was determined by conden- sation with diacetyl monoxime [11]. Tissue to be assayed for ornithine car- bamoyltransferase (EC 2.1.3.3) activity was homogenised in 10 volumes 3 mM cetyltrimethyl ammonium bromide in a glass-teflon Potter-Elvejhem homog- eniser at 0--4°C. The homogenate was centrifuged at 4000 X g for 15 min at 0--4°C and activity was determined in the supernatant by measuring the rate of production of citrulline in the presence of ornithine and carbamyl phos-

137
phate [16]. Any citrulline formed was determined by the diacetyl monoxime method [11]. Tissue to be assayed for ornithine decarboxylase (EC 4.1.1.17) activity was homogenised at 0--4°C in a Potter-Elvejhem homogeniser in 5 vol- umes of a medium containing 25 mM Tris, 1 mM EDTA and 5 mM dithiothrei- tol adjusted to pH 7.5 with HC1. The homogenate was centrifuged at 20 000 X g for 10 min and the supernatant obtained recentrifuged at 105 000 X g for 60 min at 0--4°C. Activity was determined in the final supernatant fraction by measuring the rate of product ion of radioactive CO2 evolved from L-[1-'4C] - ornithine. 0.5 ml of the supernatant was added to 0.5 ml of a solution contain- ing 100 mM glycylglycine, pH 7.2, 10 mM dithiothreitol, 0.4 mM pyridoxal phosphate and 2 mM L-[1-14C]ornithine (0.2 pCi) in flasks closed with suba- seal stoppers from which were suspended glass vials containing 0.2 ml hyamine hydroxide (1 mol/1 methanol). The flasks were incubated for 60 min at 37°C. The reaction was stopped by addition of 0.5 ml 2.5 M trichloracetic acid and the incubation continued for a further 30 min. The vials containing hyamine hydroxide were then placed in 10 ml dioxan-based liquid scintillator. The radioactivity was counted in a Corumatic 25 liquid scintillation counter (Tra- cerlab, Weybridge, Surrey, U.K.) and corrections for efficiency were made using an external standard. Tissue to be assayed for ornithine aminotransferase (EC 2.6.1.13) activity was homogenised at 0--4°C in a Potter-Elvejhem homog- eniser in a medium containing 0.25 M sucrose, 0.1 M potassium dihydrogen or thophosphate and 10 mM 2-mercaptoethanol adjusted to pH 8.0 with KOH. The activity was determined by measuring the rate of formation of A-pyrroline- 5-carboxylate using o-aminobenzaldehyde [ 17 ].
In all cases the rates of the enzyme catalysed reactions were constant over the incubation period and proportional to the amount of homogenate added. Enzyme activities have been calculated as pmol substrate transformed/min/g wet weight cotyledon.
Chemical assays The protein contents of tissue homogenates were determined by the method
of Lowry et al. [18] using crystallized and lyophilized bovine serum albumin as a standard. The DNA contents of tissue homogenates were determined using the method described by Burton [19]. The standard used was 2-deoxyribose and the concentrat ion of DNA in the sample was calculated by assuming a mean residue weight of 325 and one reacting deoxyribose molecule per two residues.
Results and Discussion
The results are presented as the mean values for the last three 31 day periods of pregnancy. During the period from 55--85 days the placenta may be con- sidered as being in the growth phase of development whilst during the two remaining periods the placenta may be considered as maturing. The results pre- sented in Table I for the DNA content of placental cotyledons support this view since they indicate that the number of cells per gram of tissue is signifi- cantly greater after 86 days gestation than in the previous month. There is no significant difference in the mean DNA content between 86--116 and 117--147

138
T A B L E I
T H E P R O T E I N C O N T E N T A N D D E O X Y R I B O N U C L E I C A C I D C O N T E N T OF P L A C E N T A L C O T Y L E -
D O N S O F S H E E P D U R I N G T H E L A S T T H R E E M O N T H S O F P R E G N A N C Y
E x p e r i m e n t a l deta i l s are g iven in t h e t e x t . Values are g iven as m e a n s ± S.E. w i t h the n u m b e r of a n ima l s in parenthese s . Resu l t s are e x p r e s s e d as m g / g we t we igh t .
D a y s f r o m c o n c e p t i o n
55--85 86--116 117--147
Prote in c o n t e n t 57 ± 3 (24) 96 +- 8 (12) 118 ± 6 (28)
D e o x y r i b o n u c l e i c acid 1.72 ± 0 .20 (24) 2 .35 +- 0 .24 (12) 2.51 +- 0 .16 (27) c o n t e n t
days of pregnancy. Since the weight of the sheep placenta is reported not to change after the third month of pregnancy [3] then it is apparent that the total number of placental cells does not alter markedly after 86 days. However, as indicated in Table I the protein content rises significantly by 67% between the third and fourth months of pregnancy and by 23% between the fourth and fifth months.
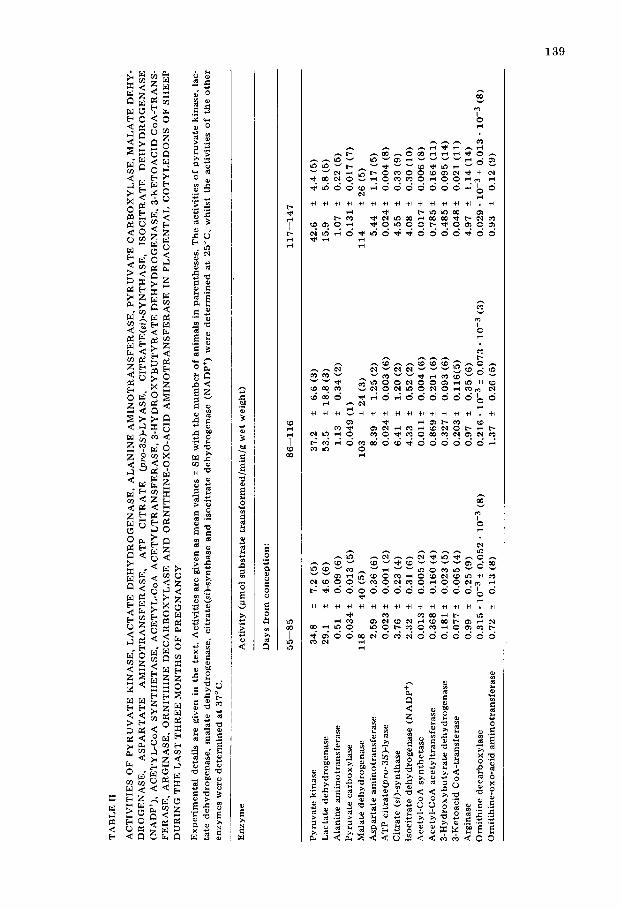
The activities of 16 enzymes are shown in Table II. No activity of either phosphoenolpyruvate carboxykinase (GTP) or ornithine carbamoyltransferase could be detected at any stage studied.
The constant and high activity of pyruvate kinase relative to most of the other enzymes studied suggests that the sheep placenta has a high glycolytic potential from 55 days onwards. High activity of pyruvate kinase has also been reported in the rat [20] and human [21] placentas but in these species, unlike the sheep, the activities decline as pregnancy advances. The high activity of lactate dehydrogenase in the sheep placenta will enable pyruvate formed from carbohydrate to be converted readily to lactate. Calculations based on the arterio-venous differences for lactate across the uterine and umbilical circula- tions [22] and the assumptions that uterine blood flow is 1.5 1/min [23] um- bilical blood flow is 200 ml/kg foetus/min [24] and foetal weight near term is 3.5 kg [9] reveal that there is a release of lactate by the uterus and placenta into both the maternal and foetal circulations at rates of 105 and 112 pmol/ min respectively. The activities of pyruvate kinase and lactate dehydrogenase when related to total placental weight (assumed to be 0.45 kg) are far in excess of the rate of catalysis required if the lactate released is derived from glucose. It is probable that most of the acetyl CoA required for citric acid cycle metabol- ism is derived from pyruvate but no measurement of pyruvate dehydrogenase (lipoate) activity has been made in the sheep placenta. In contrast to the out- put of lactate by the sheep placenta calculations based on arterio-venous differ- ences for plasma alanine [25] uterine and umbilical blood flows [23,24] and on the additional assumptions that maternal and foetal haematocrits are 30 and 35 per cent respectively [26] indicate that there is a net uptake of alanine by the placenta of 167 pmol/min. Alanine must either be utilized for protein syn- thesis or converted to pyruvate within placental tissue. The ability to convert alanine to pyruvate as indicated by the total activity of alanine aminotrans- ferase in the mature placenta is 481 pmol/min and this is approximately three
TA
BL
E
II
AC
TIV
ITIE
S
OF
P
YR
UV
AT
E
KIN
AS
E,
LA
CT
AT
E
DE
HY
DR
OG
EN
AS
E,
AL
AN
INE
A
MIN
OT
RA
NS
FE
RA
SE
, P
YR
UV
AT
E
CA
RB
OX
YL
AS
E,
MA
LA
TE
D
EH
Y-
DR
OG
EN
AS
E,
AS
PA
RT
AT
E
AM
INO
TR
AN
SF
ER
AS
E,
AT
P
CIT
RA
TE
(p
ro-3
S)-
LY
AS
E,
CIT
RA
TE
(si)
-SY
NT
HA
SE
, IS
OC
ITR
AT
E
DE
HY
DR
OG
EN
AS
E
(NA
DP
+),
A
CE
TY
L-C
oA
S
YN
TH
ET
AS
E,
AC
ET
YL
-Co
A
AC
ET
YL
TR
AN
SF
ER
AS
E,
3-H
YD
RO
XY
BU
TY
RA
TE
D
EH
YD
RO
GE
NA
SE
, 3
-KE
TO
AC
ID
Co
A-T
RA
NS
-
FE
RA
SE
, A
RG
INA
SE
, O
RN
ITH
INE
D
EC
AR
BO
XY
LA
SE
A
ND
O
RN
ITH
INE
-OX
O-A
CID
A
MIN
OT
RA
NS
FE
RA
SE
IN
P
LA
CE
NT
AL
C
OT
YL
ED
ON
S
OF
S
HE
EP
D
UR
ING
T
HE
L
AS
T
TH
RE
E
MO
NT
HS
O
F
PR
EG
NA
NC
Y
Ex
per
imen
tal
det
ails
ar
e g
iven
in
th
e te
xt.
A
ctiv
itie
s ar
e g
iven
as
mea
n
val
ues
+
SE
w
ith
th
e n
um
ber
o
f an
imal
s in
par
enth
eses
. T
he
acti
vit
ies
of
py
ruv
ate
kin
ase,
la
c-
tate
d
ehy
dro
gen
ase,
m
alat
e d
ehy
dro
gen
ase,
ci
trat
e(si
)-sy
nth
ase
and
is
oci
trat
e d
ehy
dro
gen
ase
(NA
DP
÷)
wer
e d
eter
min
ed
at
25
°C,
wh
ilst
th
e ac
tiv
itie
s o
f th
e o
ther
en
zym
es
wer
e d
eter
min
ed
at
37
°C.
En
zym
e A
ctiv
ity
(p
mo
l su
bst
rate
tr
ansf
orm
ed/m
in/g
w
et
wei
gh
t)
Day
s fr
om
co
nce
pti
on
:
55
--8
5
86
--1
16
1
17
--1
47
Py
ruv
ate
kin
ase
34
.8
+ 7
.2
(5)
37
.2
± 6
.6
(3)
42
.6
± 4
.4
(5)
Lac
tate
d
ehy
dro
gen
ase
29
.1
± 4
.6
(6)
53
.5
+- 1
8.8
(3
) 1
5.9
±
5.8
(5
) A
lan
ine
amin
otr
ansf
eras
e 0
.51
+
0.0
9
(6)
1.1
3
± 0
.34
(2
) 1
.07
±
0.2
2
(5)
Py
ruv
ate
carb
ox
yla
se
0.0
34
+-
0
.01
3
(5)
0.0
49
(1
) 0
.13
1
± 0
.01
7
(7)
Ma]
ate
deh
yd
rog
enas
e 1
18
±
40
(5
) 1
03
±
24
(3
) 1
14
±
26
(5
)
Asp
arta
te
amin
otr
ansf
eras
e 2
.59
±
0.3
6
(6)
8.3
9
± 1
.25
(2
) 5
.44
±
1.1
7
(5)
AT
P ci
trat
e(pr
o-3S
)-ly
ase
0.0
23
±
0.0
01
(2
) 0
.02
4
± 0
.00
3
(6)
0.0
24
±
0.0
04
(8
)
Cit
rate
(s
i)-s
yn
thas
e 3
.76
±
0.2
3
(4)
6.4
1
± 1
.20
(2
) 4
.55
+-
0
.33
(9
) Is
oci
trat
e d
ehy
dro
gen
ase
(NA
DP
+)
2.3
2
+-
0.3
1
(6)
4.3
3
± 0
.52
(2
) 4
.08
+
0.3
0
(10
) A
cety
l-C
oA
sy
nth
etas
e 0
.01
3
± 0
.00
5
(2)
0.0
11
±
0.0
04
(6
) 0
.01
7
± 0
.00
6
(8)
Ace
tyl-
Co
A
acet
ylt
ran
sfer
ase
0.3
68
±
0.1
60
(4
) 0
.86
9
± 0
.20
1
(6)
0.7
85
±
0.1
64
(1
1)
3-H
yd
rox
yb
uty
rate
d
ehy
dro
gen
ase
0.1
81
±
0.0
23
(5
) 0
.32
7
± 0
.09
3
(6)
0.4
85
±
0.0
95
(1
4)
3-K
eto
acid
C
oA
-tra
nsf
eras
e 0
.07
7
± 0
.06
5
(4)
0.2
03
±
0.1
16
(5)
0.0
48
±
0.0
21
(1
1)
Arg
inas
e 0
.99
*-
0
.25
(9
) 0
.97
±
0.3
5
(6)
4.9
7
± 1
.14
(1
4)
Orn
ith
ine
dec
arb
ox
yla
se
0.3
15
•
10
-3
± 0
.05
2
• 1
0 -
3 (
8)
0.2
16
•
10
-3
±
0.0
73
•
10
-3
(3
) 0
.02
9
• 1
0 -
3 ±
0
.01
3
• 1
0 -
3
(8)
Orn
itih
ine-
ox
o-a
cid
am
ino
tran
sfer
ase
0.7
2
+-
0.1
3
(8)
1.3
7
± 0
.26
(5
) 0
.93
±
0.1
2
(9)
~D

140
times the calculated alanine uptake. The activity of alanine aminotransferase per gramme of tissue is significantly less during the growth phase than that found in the mature placenta.
One fate of pyruvate formed from either carbohydrate or alanine is its con- version to oxaloacetate. That this process assumes more importance as the placenta ages is indicated by the fact that the activity of pyruvate carboxylase is four times greater near term compared with that found during the third month of pregnancy.
There is an uptake of aspartate from both the uterine and umbilical circula- tions [25]. In addition to probable roles in protein and nucleic acid synthesis, aspartate may be converted to oxaloacetate at a greater rate in the mature placenta than in the growing organ since aspartate aminotransferase activity is greater in the mature organ. Oxaloacetate formed is likely to be in equilibrium with malate at all times since the activity of malate dehydrogenase is substan- tiaUy higher than that of any other enzyme determined in the study. The pro- vision of extramitochondrial oxaloacetate by way of the reaction catalysed by ATP citrate(pro-3S)lyase would not appear to be of any great quantitative significance. It is unlikely that oxaloacetate derived from any source could be converted to glucose in the sheep placenta since phosphoenolpyruvate carboxy- kinase (GTP) activity could not be detected in this tissue in contrast to the rat [20] and human [21]. That phosphoenolpyruvate carboxykinase activity is not absent from all foetal lamb tissues is indicated by the results of our studies in foetal lamb liver in which similar methods for preparation of tissue homog- enates and assay of enzyme activity were used [9]. Hence gluconeogenesis from substrates such as alanine, pyruvate and lactate is unlikely to occur in the sheep placenta.
There are no changes in the activity of citrate(si)-synthase per g tissue and hence the potential for energy production by way of the citric acid cycle does not change during the maturation of the placenta when it might be expected that the energy requirements would be increased to enable active transport to the growing foetus to proceed more rapidly. The potential utilization of citrate as a source of reduced NADP ÷ by way of the isocitrate dehydrogenase reaction increases significantly during the growth phase but remains unchanged after 86 days. An important role of reduced NADP in the placenta is in the interconver- sion of steroid hormones resulting in the formation of progesterone and oestro- gens [27].
Acetate is unlikely to be metabolised rapidly by placental tissue since acetyl- CoA synthetase activity is relatively low. Calculations based on the arterioven- ous differences for acetate [28] and on blood flows [23,24] show that there is a net uptake of acetate by the uterus and placenta of 111 pmol/min. Assuming a mature placental weight of 0.45 kg the total placental activity of acetyl-CoA synthetase near term is not sufficient to account for the disappearance of ace- tate and therefore it must be assumed that acetate is being utilised by tissues other than the placenta which are drained by the uterine veins.
Acetoacetate production from an acetyl-CoA generating system was found to be very variable and there were no significant differences found between the three 31 day periods studied. The mean rate of production in nineteen placen- tae was 0.270 + 0.128 pmol/min/g wet weight. Therefore, the rate of ketone

141
body product ion b y a 0.45 g placenta near term would not be greater than 121 pmol/min. In the mature placenta the activity of 3-hydroxybutyra te dehydro- genase is significantly higher than in the growth phase and is sufficient to con- vert any acetoacetate synthesised to 3-hydroxybutyrate . The calculated release of acetoacetate plus 3-hydroxyburyrate into the foetal circulation based on the studies of Morriss et al. [29] is only 6 #mol/min. If acetoacetate is synthesised in vivo at 121 pmol/min/placenta then approximately 95% might be expected to be released into the maternal circulation and hence contr ibute to the ketosis found in pregnant ewes. The presence of placental 3-hydroxybutyra te dehydro- genase activity may play a role in the maintenance of the low acetoacetate con- centration found in the foetal circulation when the maternal acetoacetate con- centration is raised by starvation [29]. The placenta has the capacity to oxidise ketone bodies since 3-ketoacid CoA-transferase activity is present. There are no significant differences be tween the mean values during the last three months of pregnancy. The activity of this enzyme was found to be considerably less than that of acetyl-CoA acetyltransferase an enzyme believed to be concerned not only with ketone body metabolism but also with the oxidation of long chain fat ty acids. Since the activity of the acetyltransferase enzyme is only between 10--20% of the activity of citrate synthase it seems unlikely that either ketone bodies or fa t ty acids make a substantial contr ibut ion to the oxidative metabol- ism of the placenta by way of the citric acid cycle. The presence of the ketone body oxidative pathway in the placenta suggests that the production of aceto- acetate from acetyl-CoA need not proceed by way of 3-hydroxy-3-methyl- glutaryl-CoA. It could occur by reversal of the oxidative pathway coupled with non-specific deacylat ion as discussed by Bush and Milligan [30].
If the uptake of alanine and aspartate by the placenta is followed by conver- sion to the corresponding ketoacids, the fate of the amino group must be con- sidered. Release of this nitrogen from the placenta in the form of urea is extremely unlikely since urea formation could not be demonstrated in placen- tal slices incubated with ammonium salts and ornithine. Since amino-nitrogen is not released from the placenta in the form of alanine the possibility exists that amino groups may be released in the form of glutamine provided that glutamate is available. There is an uptake of glutamate by the sheep placenta from both maternal and foetal circulations [25] but no information is avail- able on the concentrat ion of glutamine in either the uterine or umbilical circu- lations. The inability of the placenta to synthesise urea is in contrast to the finding that slices of foetal ovine liver incubated in similar conditions can syn- thesise urea at a mean rate of 0.310 pmol/min/g wet weight of liver at 37°C [31]. The absence of detectable ornithine carbamoyltransferase activity in pla- centa and its presence in foetal liver can account for these differences [32]. How- ever, the placenta could produce urea from arginine since arginase activity is present. There is a five-fold increase in arginase activity during the last month of pregancy. From the data of Hopkins et al. [25] and the consideration of relative plasma flows it is evident that there is a net uptake of arginine by the sheep uterus and placenta and since the activity of placental arginase far exceeds this uptake it is likely that any arginine not incorporated into protein will be converted to ornithine and urea.
Ornithine is taken up from maternal and foetal circulations [25]. This orni-

1 4 2
thine together with any derived from arginine must be metabolised. The high activity of placental ornithine-oxo-acid aminotransferase compared with that found in sheep liver and kidney [31] suggests that the conversion of ornithine to A-pyrroline 5-carboxylate may be important in the placenta. This may account for the failure of raised maternal plasma ornithine concentrations to be reflected in foetal plasma [33]. In mammalian tissues A-pyrroline 5-carboxy- late can be converted to either glutamate or proline. Since there is no release of glutamate it is suggested that A-pyrroline 5-carboxylate may be converted into proline and released from the placenta.
It is unlikely that decarboxylation and subsequent polyamine synthesis is quantitatively an important route of ornithine metabolism since ornithine decarboxylase activity is very much lower than that of the aminotransferase. The activity of ornithine decarboxylase has been related to nucleic acid synthe- sis and organ growth [34]. The fact that the activity of this enzyme declines 10-fold during the maturation stage is notable since by this stage the placenta has ceased to grow and its DNA content is constant (Table I).
It must be emphasised that the activities of enzymes in vivo are dependent not only on the amount of enzyme protein present but also on the concen- tration of substrates, products and possible allosteric effectors. Even so it might be expected that the enzyme profile of the sheep placenta would be markedly altered during the last month of pregnancy to allow placental metabolism to keep pace with the requirements of the growing foetus. The present study shows that maximal activities of most of the enzymes studied are achieved by the time placental weight has reached its maximum. Only arginase and possibly pyruvate carboxylaseaclfivity increases in proportion to foetal weight during placental maturation whilst ornithine decarboxylase activity declines during this time.
Acknowledgements
This investigation was supported by a grant from the Agricultural Research Council. E.M.E., J.M.R. and G.C.E.V. were recipient of Postgraduate Student- ships from the Ministry of Agriculture, Fisheries and Food. We thank Mr. I.A.N. Wilson of the Grassland Research Institute, Hurley, Berks. who carried out the radiological examination of the pregnant sheep.
References
1 K u l h a n e k , J . F . , Meschia , G. , M a k o w s k i , E.L. a n d Ba t t agHa , F .C. ( 1 9 7 4 ) Am. J . Phys io l . 2 2 6 , 1 2 5 7 - - 1 2 6 3
2 Dawes , G.S. ( 1 9 6 8 ) F o e t a l a n d Neonata l Phys io logy , p. 25 , Yeaz B o o k Medical Publishers Inc. , C h i c a g o
3 Ever i t t , C.G. ( 1 9 6 4 ) N a t u r e 2 0 1 , 1 3 4 1 - - 1 3 4 2 4 B r i t t o n , H.G. , H u g g e t t , A. S t .G . a n d N i x o n , D .A . ( 1 9 6 7 ) B i o c h i m . B i o p h y s . A c t a 136 , 4 2 6 - - 4 4 0 5 S tave , U. ( 1 9 7 0 ) Phys io logy of the Per ina t a l Per iod (Stave, U., ed.) , pp . 5 5 9 - - 5 9 4 , Appleton-Century-
Crofts, N e w Y o r k 6 Hall , L.M. ( 1 9 6 2 ) A n a l y t i c a l B i o c h e m i s t r y 3, 7 5 - - 8 0 7 S i m o n , E.J . a n d S h e m i n , D. ( 1 9 5 3 ) J . Am- Chem~ Soc . 75, 2 5 2 0 8 S te rn , J .R . ( 1 9 5 6 ) J . Biol. C h e m . 2 2 1 , 3 3 - - 4 4 9 E d w a r d s , E.M., D h a n d , U.K. , J e a c o c k , M.K. a n d S h e p h e r d , D . A . L . ( 1 9 7 5 ) B i o c h i m . B i o p h y s . A c t a
399 , 2 1 7 - - 2 2 7

143
10 Krebs, H.A. and Henseleit, K. (1932) Hoppe-Zeyler 's Z. Physiol. Chem. 210, 33--66 11 Moore, R.B. and Kauffman, N.J. (1970) AnaL Biochem. 33, 263--272 12 Williamson" D.H., Bates, M.W. and Krebs, H.A. (1968) Biocherrh J. 108, 353--361 13 Wllliamson" D.H., Mella-nby, J. and Krebs, H.A. (1962) Biochem. J. 82, 90--96 14 Dhand, U.K., Jeacock, MJ£., Shepherd, D.A.L., Smith, E.M. and Varnam, G.C.E. (1970) Biochim.
Biophy~ Acta 222, 216--218 15 Williamson, D.H., Bates, M.W., Page, M.A. and Krebs, H.A. (1971) Biochem. J. 121, 41--47 16 Brown" G.W. and Cohen" P.P. (1959) J. Biol. Chem. 234, 1769--1774 17 Peralno, C. and Pitot, H.C. (1963) Biochim. Biophys. Acta 73, 222--231 18 Lowry, O.H., Rosebrough, N.J., Farr, A.L. and Randall, R.J. (1951) J. Biol. Chem. 193, 265--275 19 Burton, K. (1956) Biochem. J. 62, 315--323 20 Diamant, Y.Z. and ShafrJx, E. (1972) Biochim. Biophys. Acta 279, 424--430 21 Diamant, Y.Z., Mayorek, N., Neuman" S. and Shafrix, E. (1975) Am. J. Obstet. Gynecol. 121, 58--61 22 Burd, L.I., Jones, M.D., Simmons, M.A., Makowski~ E.L., Meschia, G. and BattagHa, F.C. (1975)
Nature 254, 710--711 23 Rosenfeld, C.R., Morriss, F.H., Makowski, E .L, Meschia, G. and Battaglia, F.C. (1974) Gynecol.
Invest. 5, 252--268 24 Stevens, D.H. (1975) Comparative Placentation. Essays in Structure and Function, p. 166, Academic
Press, London 25 Hopkins, L., MeFadyen" I.R. and Young, M. (1971) J. Physiol (London) 215, 9--10P 26 Shelley, H.J. (1973) in Foetal and Neonatal Physiology (Comline, K., Cross, K.W., Dawes, G.S. and
Nathanielsz, P.W., eds.), pp. 360--381, Cambridge 27 Pierrepoint, C.G., Anderson, A.B.M., Turnbull, A.C. and Griffiths, K. (1973) in The Endocrinology of
Pregnancy and Parturit ion. (Pierrepoint, C.G., ed.), pp. 40--53, Alpha Omega Alpha Publishing, Cardiff
28 Char, V.C. and Creasy, R.K. (1976) Am. J. Physiol. 230, 357--261 29 Morriss, F.H., Boyd, R.D.H., Makowski, E.L., Meschia, G. and Ba~aglia, F.C. (1974) l~oc. Soc. Exp.
Biol. Med. 145, 879--883 30 Bush, R.S. and Milligan, L.P. (1971) Can. J. Anim. Sci. 51, 129--133 31 Rat tenbury, J.M. (1975) Ph.D. Thesis, University of Reading 32 Rat tenbury, J.M., Jeacock, M.K. and Shepherd, D.A.L. (1972) Biochem. J. 128, 143--144P 33 Hopkins, L., McFadyen" I.R. and Young, M. (1971) J. Physiol (London) 215, l l - - 1 2 P 34 Cohen, S.S. (1971) In t roduct ion to the Polyamines, pp. 28--63, Prentice-Hall Inc. New Jersey