Embed Size (px)
Citation preview

Insect Science 13, 251J256
Properties of phenoloxidase from Pieris rapae 251
Introduction
Phenoloxidase (PO) (EC 1.14.18.1), also known astyrosinase, a copper containing bifunctional enzyme, ishighly conserved, present in microorganisms, animals andplants (Sáchez-Ferrer et al., 1995; Chase et al., 2000). It isthe key enzyme involved in melanin formation inmelanocytes. Phenoloxdase catalyzes both hydroxylationof monophenols and oxidation of o-diphenols into o-quinones, and is involved in the formation of pigmentssuch as melanins (Chen et al., 2003). In insects, PO isuniquely associated with three different physiologicallyimportant biochemical processes, including sclerotizationof the insect cuticle, defensive encapsulation and mela-nization of foreign organisms and wound healing (Ashida
Correspondence: Wan-Chun Luo, College of Plant Protection,Shandong Agricultural University, Shandong, Tai�an 271018,China. Tel: +86 538 8242983; fax: +86 538 8242983; email:[email protected]
Enzymatic properties of phenoloxidase from Pieris rapae(Lepidoptera) larvae
www.blackwellpublishing.com/ins 251
Insect Science (2006) 13, 251J256
C H A O - B I N X U E 1, WA N - C H U N L U O 1, Q I N G - X I C H E N 2, Q I N WA N G 2 a n dL I - N A K E 2
1College of Plant Protection, Shandong Agricultural University, Shandong, Tai�an, 2The Key Lab of Ministry of Education forCell Biology and Tumor Cell Engineering, School of Life Sciences, Xiamen University, Fujian, Xiamen, China
& Brey, 1995). Production of o-quinones by PO has beenknown to be an initial step in the biochemical cascade of β-sclerotization, quinone tanning and melanin biosynthesis,processes that occupy several major roles in insect devel-opment and immunity. It is possible that inhibition of POcould lead to abrogation of insect defense mechanisms orabnormal body softening, both of which could be used inpest control. Therefore, PO inhibitors are potential candi-dates for developing novel insecticides.
Phenoloxidase inhibitors have increasingly been used inmedicinal and cosmetic products (Maeda & Fukuda, 1991;Friedman, 1996). Recently, copper chelators have beenfound to inhibit mushroom tyrosinase. Tetra-hexylresorci-nol and 4-dodecylresorcinol have been characterized as themushroom tyrosinase inhibitors of L-DOPA oxidation(Chen et al., 2004). Some flavor compounds from olive oiland some flavonol compounds from saffron flower havebeen reported to inhibit mushroom tyrosinase. Therefore,the use of PO inhibitors may enable the development ofsafe alternatives to environmentally hazardous insecticides.
The PO from some insects such as tobacco budworm,
Abstract The kinetic parameters of partially purified phenoloxidase (PO, EC. 1.14.18.1)from the 5th instar larvae of Pieris rapae (Lepidoptera) were determined, using L-3, 4-dihydroxyphenylalanine (L-DOPA) as substrate. The optimal pH and temperature of theenzyme for the oxidation of L-DOPA were determined to be at pH 7.0 and at 42oC,respectively. The enzyme was stable between pH 6.5 and 7.4 and at temperatures lower than37oC. At pH 6.8 and 37oC, the Michaelis constant (K
m) and maximal velocity (V
m) of the
enzyme for the oxidation of L-DOPA were determined to be 0.80 mmol/L and 1.84 µmol/L/min, respectively. Tetra-hexylresorcinol and 4-dodecylresorcinol effectively inhibitedactivity of phenoloxidase and this inhibition was reversible and competitive, with the IC
50
of 1.50 and 1.12 µmol/L, respectively. The inhibition constants were estimated to be 0.50 and0.47µmol/L, respectively.
Key words inhibitory mechanism, Pieris rapae, phenoloxidase, properties, 4-hexylresorcinol, 4-dodecylresorcinolDOI 10.1111/j.1744-7917.2006.00091.x

Insect Science 13, 251J256
252 C. B. Xue et al.
Musca domestica, Drosophila melanogaster, Aedes aegypti,Plodia interpunctella and Sarcophaga bullata has beeninvestigated (Yamaura et al., 1980; Ourth, 1988; Fujimotoet al., 1993; Burks & Fuchs, 1995; Chase et al., 2000;Wang et al., 2004; Hartzer et al., 2005). However, to ourknowledge, no reported study has focused on the inhibitionmechanism of PO from the larvae of Pieris rapae. Therefore,the properties of PO and inhibition mechanisms of 4-hexylresorcinol and 4-dodecylresorcinol on the enzymefrom the larvae of P. rapae were examined and are de-scribed here.
Materials and methods
Insects and reagents
Pieris rapae larvae were reared on Chinese cabbageBrassica parachinesi (Bailey) in a greenhouse at 25 � 1oCwith a 14:10 h (light: dark) photoperiod. The 5th instarlarvae were gathered for the experiments. L-3, 4-dihydroxyphenylalanine (L-DOPA) and dimethyl sulfox-ide (DMSO) were purchased from Aldrich Chemical Co.(Milwaukee, WI, USA). Tetra-hexylresorcinol and 4-dodecylresorcinol were purchased from Sigma ChemicalCo. (St. Louis, MO, USA). Sephadex G-100 was pur-chased from Amersham Pharmacia Biotech. (Uppsala,Sweden). All other reagents were local products of analyti-cal grade. The water was re-distilled and ion-free.
Purification of PO
All procedures were carried out at 4oC. The 5th instar P.rapae larvae were homogenized in a 5-fold weight of ice-cold 0.2 mol/L sodium phosphate buffer (pH 6.8). Thehomogenate was centrifuged for 30 min at 9 310 g afterextraction for 2 h. The supernatant under the fat layer wascollected as the crude enzyme extract, and was brought to35% saturation with solid ammonium sulfate. The precipi-tate was collected by centrifugation at 9 310 g for 30 minand then re-dissolved in a minimum volume of the samephosphate buffer, and was dialyzed against the 10 mmol/Lsodium phosphate buffer (pH 6.8). The dialyzed solutionwas loaded onto a Sephadex G-100 column (2.5 cm�60 cm),which was equilibrated with 10 mmol/L sodium phosphatebuffer (pH 6.8), and those fractions having high activitywere collected. The second process increased the specificactivity up to 6.22-fold of that of the crude enzyme extract.
Assays of PO activity
Phenoloxidase activity was determined by measuring
the rate of dopachrome formation at 475 nm with a molarabsorption coefficient of 3 700 (mol/L/cm) for the product(Jiménez et al., 2001) using a Beckman UV-650 spectro-photometer at 37oC. The linear increase in optical densityat 475 nm was monitored. The compositions of the assaymedia are given in the figure legends. The reaction wasstarted by enzyme addition.
Assay of the optimal pH and pH stability of PO
The optimum pH was determined by measuring theenzyme activity as described above at various pH condi-tions (range, 5.9J8.0) at 37oC. The pH stability of theenzyme was monitored after incubating the enzyme in thedifferent pH buffers for 30 min at 4oC. Some of the enzymemixture (0.5 mL) was used for an activity assay at 37oC, pH6.8.
Assay of the optimal temperature and thermal stability of PO
The optimum temperature of the enzyme was deter-mined by measuring the activity as described above atvarious temperatures (range, 22J62oC) at pH 6.8. Thethermal stability of the enzyme was monitored whileincubating the enzyme at different temperatures for 30min. Again, 0.5 mL of the enzyme mixture was used for anactivity assay at 37oC, pH 6.8.
Effect of 4-hexylresorcinol and 4-dodecylresorcinol on theactivity of PO
The inhibitor was first dissolved in DMSO (final DMSOconcentration in test solution was 3.3%) (Chen et al.,2004). The extent of inhibition was expressed as thechemical concentration to inhibit 50% of the enzymeactivity (IC
50). The inhibition type was determined by the
Lineweaver-Burk plot. The inhibition constant was deter-mined by the plot of the apparent Michaelis-Menten con-stant versus the concentration of the inhibitor.
Results
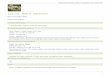
Determination of the kinetic parameters of PO
The kinetic behavior of the enzyme on L-DOPA oxida-tion was studied. Under the present conditions, L-DOPAoxidation by the enzyme followed Michaelis-Menten ki-netics (Fig. 1, inset). The enzyme�s kinetic parameters K
m
and Vmax
were 0.80 mmol/L and 1.84 µmol/L/min, respec-tively (Fig. 1).
Insect Science 13, 251J256
Properties of phenoloxidase from Pieris rapae 253
Effects of pH on enzyme activity and pH stability
At 37oC, the effect of pH on enzyme activity during L-DOPA oxidation was determined. The optimum oxidationreaction occurred at pH 7.0. When the tested pH conditionwas higher or lower than the optimum pH, enzyme activitydecreased rapidly (Fig. 2a). On the other hand, when theenzyme was incubated at various pH conditions for 30 minat 4oC and the mixture was used for activity assay in thestandard system (at 37oC, pH 6.8), activity of the enzymewas stable in the pH range of 6.5J7.4 (Fig. 2b).
Effects of temperature on enzyme activity and thermalstability
Enzyme activity results acquired at various temperaturessuggested that the optimal temperature was 42oC. Enzymeactivity decreased rapidly at temperatures higher or lowerthan the optimum (Fig. 3a). But the activity was relativelystable at temperatures below 37oC (Fig. 3b).Fig. 1 Lineweaver-Burk plot for the oxidation of L-DOPA by
PO from the 5th instar larvae of P. rapae. Assay conditions were0.2 mol/L Na
2HPO
4-NaH
2PO
4 buffer (pH 6.8) and different
concentrations of L-DOPA at 37oC. The addition of the enzymesample was 0.1 mL.
Fig. 2 Effects of pH on the PO activity. a. Optimum pH, enzymeactivity was assayed with 0.2 mol/L Na
2HPO
4-NaH
2PO
4 buffer,
the concentration of L-DOPA was 1 mol/L. b. pH stability, theenzyme sample (0.1 mL) was incubated in 0.2mol/L Na
2HPO
4-
NaH2PO
4 (pH 5.9J8.0) buffers (0.4 mL) for 30 min at 4oC.
Fig. 3 Effect of temperature on the PO activity. a. Optimumtemperature. b. The thermal stability of the enzyme, the enzymesample (0.1 mL) was incubated in 0.2 mol/L Na
2HPO
4-NaH
2PO
4
buffer (0.4 mL) at different temperatures (22 J 62oC) for 30 min,and enzyme activity was assayed at 37oC, pH 6.8.
Effects of inhibitor concentration on enzyme activity
The effects of 4-hexylresorcinol and 4-dodecylresorcinolon the L-DOPA oxidation by PO from the 5th instar P.
Insect Science 13, 251J256
254 C. B. Xue et al.
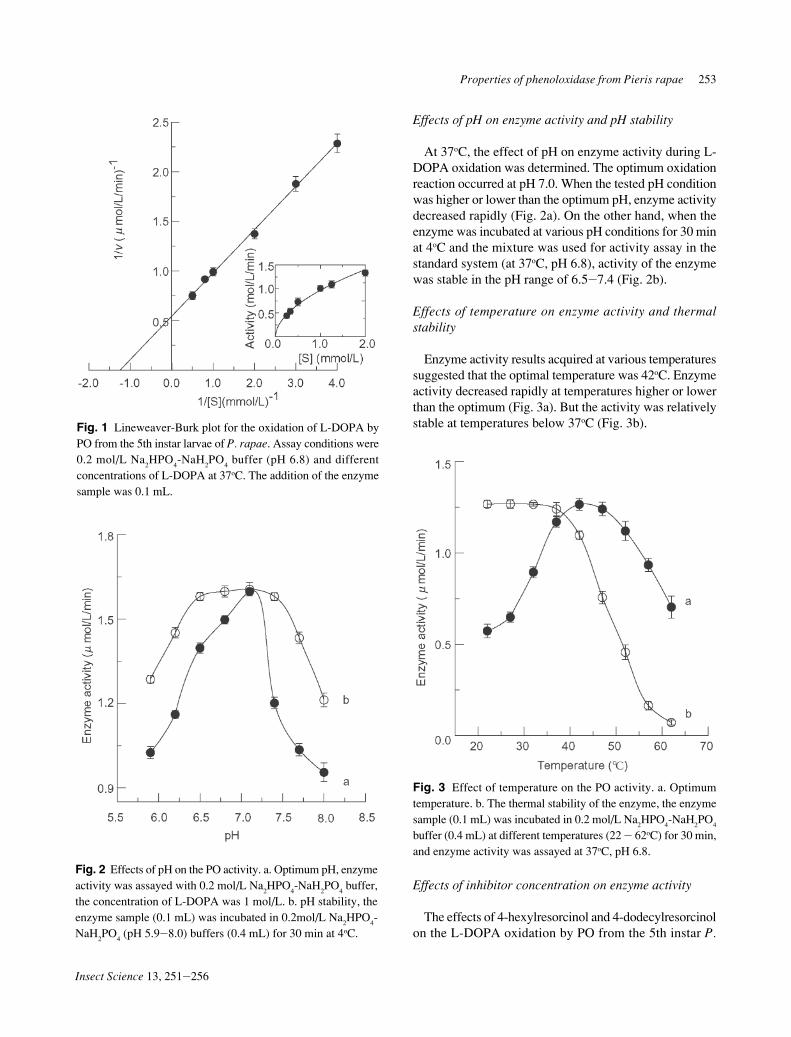
rapae larvae were studied first. The inhibitory course ofenzyme act ivi ty by 4-hexylresorcinol and 4-dodecylresorcinol was found to be concentration-depen-dent as shown in Fig. 4. With increasing concentrations of4-hexylresorcinol and 4-dodecylresorcinol, the residualenzyme activity decreased rapidly, although it was notcompletely suppressed at any concentration. The chemicalconcentration to inhibit 50% of the enzyme activity (IC
50)
was estimated to be 1.50 µmol/L for 4-hexylresorcinol and1.12 µmol/L for 4-dodecylresorcinol.
Fig. 4 Inhibitions of 4-hexylresorcinol (a) and 4-dodecylresorcinol(b) on the PO from the 5th larvae of P. rapae for the catalysis ofL-DOPA.
Reversible inhibition kinetics of two compounds on enzymeactivity
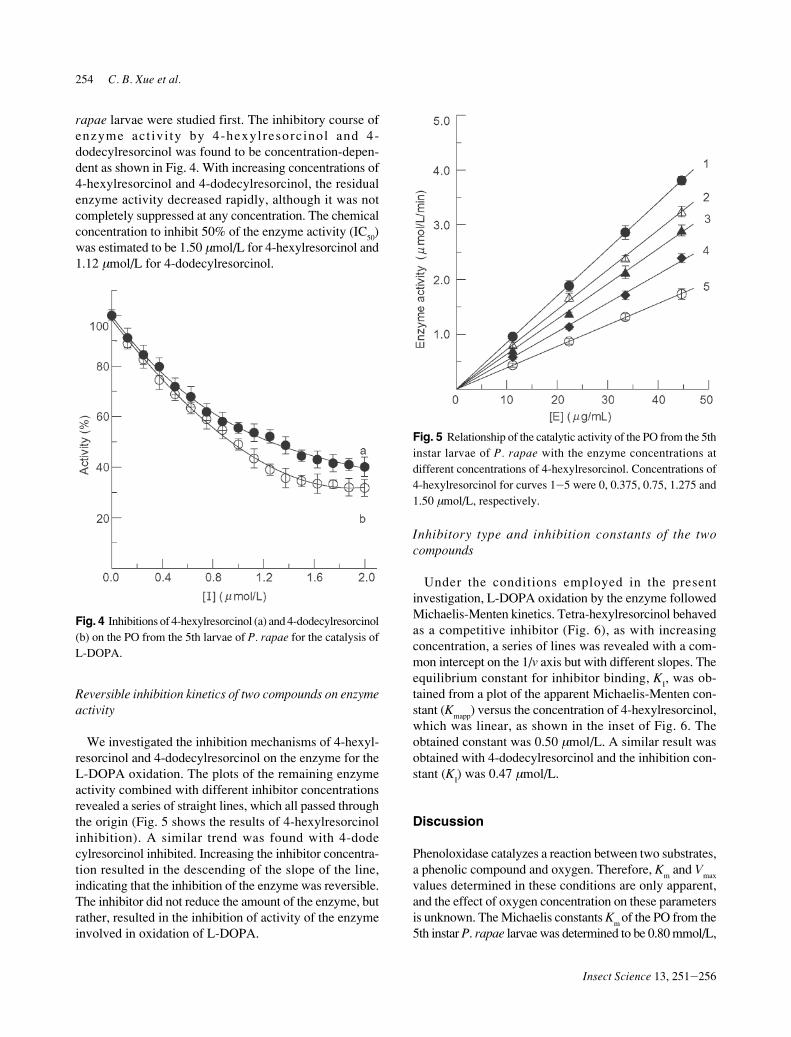
We investigated the inhibition mechanisms of 4-hexyl-resorcinol and 4-dodecylresorcinol on the enzyme for theL-DOPA oxidation. The plots of the remaining enzymeactivity combined with different inhibitor concentrationsrevealed a series of straight lines, which all passed throughthe origin (Fig. 5 shows the results of 4-hexylresorcinolinhibition). A similar trend was found with 4-dodecylresorcinol inhibited. Increasing the inhibitor concentra-tion resulted in the descending of the slope of the line,indicating that the inhibition of the enzyme was reversible.The inhibitor did not reduce the amount of the enzyme, butrather, resulted in the inhibition of activity of the enzymeinvolved in oxidation of L-DOPA.
Fig. 5 Relationship of the catalytic activity of the PO from the 5thinstar larvae of P. rapae with the enzyme concentrations atdifferent concentrations of 4-hexylresorcinol. Concentrations of4-hexylresorcinol for curves 1J5 were 0, 0.375, 0.75, 1.275 and1.50 µmol/L, respectively.
Inhibitory type and inhibition constants of the twocompounds
Under the conditions employed in the presentinvestigation, L-DOPA oxidation by the enzyme followedMichaelis-Menten kinetics. Tetra-hexylresorcinol behavedas a competitive inhibitor (Fig. 6), as with increasingconcentration, a series of lines was revealed with a com-mon intercept on the 1/v axis but with different slopes. Theequilibrium constant for inhibitor binding, K
I, was ob-
tained from a plot of the apparent Michaelis-Menten con-stant (K
mapp) versus the concentration of 4-hexylresorcinol,
which was linear, as shown in the inset of Fig. 6. Theobtained constant was 0.50 µmol/L. A similar result wasobtained with 4-dodecylresorcinol and the inhibition con-stant (K
I) was 0.47 µmol/L.
Discussion
Phenoloxidase catalyzes a reaction between two substrates,a phenolic compound and oxygen. Therefore, K
m and V
max
values determined in these conditions are only apparent,and the effect of oxygen concentration on these parametersis unknown. The Michaelis constants K
m of the PO from the
5th instar P. rapae larvae was determined to be 0.80 mmol/L,
Insect Science 13, 251J256
Properties of phenoloxidase from Pieris rapae 255
which is lower than that of other insects, such as thehousefly (Musca domestica) pupae with 3.93 mmol/L andthe blowfly (Sarcophaga bullata) pupae with 1.54 mmol/L(Wang et al., 2004). A pH of 7.0 was optimal for the L-DOPA catalysis reaction with P. rapae PO. Similar pHactivity profiles have been reported: the optimal pH ofhousefly (Musca domestica) PO aged pupae was deter-mined to be between 6.5 and 7.5, and an optimal pH of6.5 was reported in an examination of blowfly pupae(Sarcophaga bullata) (Wang et al., 2004). The optimaltemperature was 42oC for the L-DOPA catalysis reactionwith PO activity. The optimal temperature for PO fromother pupae, such as blowfly (Sarcophaga bullata), was40oC (Wang et al., 2004), and that of the pink shrimp(Panaeus duorarum) was 40oC. The PO was not extremelyheat stable, and was, in most cases, partially or totallydestroyed after short exposures to temperatures above 70oC.
Tetra-hexylresorcinol has been used to prevent shrimpand crab melanosis, and is considered safe for use inbrowning control of fresh and hot air-dried foods such asapple slices, potatoes, avocados, and grape juices (McEvilyet al., 1991). The present paper reported the effect of 4-hexylresorcinol or 4-dodecylresorcinol on the oxidation ofL-DOPA by PO from the larvae of P. rapae. The resultsshowed that 4-hexylresorcinol and 4-dodecylresorcinolare competitive inhibitors during the reaction with theenzyme, meaning that the inhibitor (I) is only bound withthe free enzyme (E) to form the EI (enzyme and inhibitor)
complex rather than binding with the ES (enzyme andsubstrate) complex. This finding is consistent with theresults reported for the mushroom tyrosinase (Chen et al.,2004).
The binding of diphenols with PO involves severalmolecular processes, including diffusion to the active siteand orientation of the substrate, followed by coordinationof a phenolic oxygen atom to one of the copper ions, andelectron transfer from the substrate to the copper center.This reaction may be facilitated or retarded by interactionswith amino acid side chains near the binuclear coppercenter (Rodríguez-López et al., 2000). These previousfindings indicate that the copper center is important for thecatalytic activity of PO. However, the present findingssuggest that the inhibitor reacted with Cu2+ to form a newcomplex, and the inhibitor seemed to affect the free en-zyme directly. Therefore, the inhibitory mechanism of 4-hexylresorcinol or 4-dodecylresorcinol could, at least inpart, involve an interaction with the copper molecules atthe active site of the enzyme. However, elucidation of thisinteraction will await further work, as the structure of POfrom the larvae of P. rapae has not yet been clearlyestablished.
In normal insect development PO plays a key role. Thusit may be possible to control pests by inhibiting or disturb-ing this enzyme. This may be a useful basis for the devel-opment of novel insecticides to replace traditional ones,many of which currently pose serious environmental threatsand/or have been associated with resistance, toxic residues,and/or resurgence in targeted pest populations. Recentstudies on PO from agricultural pests have illuminatedsome of their enzymatic properties, and some efficientinhibitors such as kojic acid and apigenin have beenidentified. This study showed that 4-hexylresorcinol and 4-dodecylresorcinol are effective PO inhibitors, and revealedsome mechanistic details of PO activity. This work mayserve as a useful basis for designing effective, selective andenvironmentally friendly PO inhibitors and in the develop-ment of novel candidate insecticides.
Acknowledgments
The present investigation was supported in part by grantNO. 30270887, 30571237 of the National Natural ScienceFoundation of China for W. C. Luo.
References
Ashida, M. and Brey, P. (1995) Role of the integument in insectdefense: prophenoloxidase cascade in the cuticular matrix.
Fig. 6 Lineweaver-Burk plot for inhibition of 4-hexylresorcinolon the PO from the 5th instar larvae of P. rapae for the catalysis ofL-DOPA at 37oC, pH 6.8. Concentrations of 4-hexylresorcinol forcurves 1J5 were 0, 0.375, 0.75, 1.275 and 1.50 µmol/L, respectively.The inset represents the plot of K
mapp versus the 4-hexylresorcinol
concentration for determining the inhibition constants (KI).

Insect Science 13, 251J256
256 C. B. Xue et al.
Proceedings of the National Academy of Science USA, 92,10698J10702.
Burks, C.S. and Fuchs, M.S. (1995) Partial purification of plasmaphenoloxidase of the yellow fever mosquito Aedes aegypti L.(Diptera: Culicidae). Comparative Biochemistry andPhysiology, 110B, 641J647.
Chase, M.R., Raina, K., Bruno, J. and Sugumaran, M. (2000)Purification, characterization and molecular cloning ofprophenoloxidase from Sarcophaga bullata. Insect Biochem-istry and Molecular Biology, 30, 953J967.
Chen, Q.X., Ke, L.N., Song, K.K., Huang, H. and Liu, X.D.(2004) Inhibitory effects of hexylresorcinol anddodecylresorcinol on mushroom (Agaricus bisporus)tyrosinase. Journal of Protein Chemistry, 23, 135J141.
Chen, Q.X., Liu, X.D. and Huang, H. (2003) Inactivation kineticsof mushroom tyrosinase in the dimethyl sulfoxide solution.Biochemistry (Moscow), 68, 644J649.
Friedman, M. (1996) Food browning and its prevention: anoverview. Journal of Agricultural and Food Chemistry, 44,631J653.
Fujimoto, K., Masuda, K.-I., Asada, N. and Ohnishi, E. (1993)Purification and characterization of prophenoloxidase frompupae of Drosophila melanogaster. Journal of Biochemistry,113, 285J291.
Hartzer, K.L., Zhu, K.Y. and Baker, J.E. (2005) Phenoloxidase inlarvae of Plodia interpunctella (Lepidoptera: Pyralidae): mo-lecular cloning of the proenzyme cDNA and enzyme activityin larvae paralyzed and parasitized by Habrobracon hebetor(Hymenoptera: Braconidae). Archives of Insect Biochemistryand Physiology, 59, 67J79.
Jiménez, M., Chazarra, S., Escribano, J., Cabanes, J. and Garcia-Carmína, F. (2001) Competitive inhibition of mushroom tyro-sinase by 4-substituted benzaldehydes. Journal of Agricul-
tural and Food Chemistry, 49, 4060J4063.Maeda, K. and Fukuda, M. (1991) In vitro effectiveness of several
whitening cosmetic components in human melanocytes. Jour-nal of Society Cosmetic Chemistry, 42, 361J368.
McEvily, A.J., Iyengar, R. and Otwell, W.S. (1991) Sulfitealternative prevents shrimp melanosis. Food Technology, 45,80J86.
Ourth, D.D. (1988) Phenoloxidase activity, lack of bactericidalimmunity and oral susceptibility of Tobacco budworm(Lepidoptera: Noctuidae) larvae to Serratia marcescens. Jour-nal of Economic Entomology, 81, 148J151.
Rodríguez-López, J.N., Fenoll, L.G., García-Ruiz, P.A., Varón,R., Tudela, J., Thoneley, R.N. and García-Cánovas, F. (2000)Stopped-flow and steady-state study of the dipenolase activityof mushroom tyrosinase. Biochemistry, 39, 10497J10506.
Sánchez-Ferrer, A., Rodríguez-López, J.N., García-Cánovas, F.and García-Carmona, F. (1995) Review tyrosinase:acomprehensive review of its mechanism. Biochimica etBiophysica Acta, 1247, 1J11.
Solomon, E.I. and Lowery, M.D. (1993) Structure contributions tofunction in bioinorganic chemistry. Science, 259, 1575J1581.
Wang, Q., Chen, Q.X., Huang, X.H., Ke, L.N., Shi, Y. andWang, J. (2004) Studies on the enzymatic characterizationand functional groups of polyphenoloxidase from pupae ofblowfly (Sarcophaga bullata). Biochemistry (Moscow), 69,918J920.
Yamaura, I., Yonekura, M., Katsura, Y., Ishiguro, M. and Funatsu,M. (1980) Purification and some physico-chemical propertiesof phenoloxidase from the larvae of housefly. AgriculturalBiological Chemistry, 44, 55J59.
Accepted March 26, 2006