Embed Size (px)
Citation preview

ENVR 230 CHEMICAL CARCINOGENESIS
Instructor: Avram Gold
Office: McGavran-Greenberg 4114C
Office phone: 6 7304
Lab: McGavran-Greenberg 3221E
Lab phone: 6 7325
e-mail: [email protected]
2 exams: final, 60%; midterm, 30%; homework + class participation 10%.
Four problem sets during semester- more if current literature section is larger.
Course web site
To be established at: http//www.unc.edu/courses/2006spring/envr/230/001/

TEXTS
MOLECULAR BIOLOGY B. Lewin, Genes VIII, Pearson Prentice Hall 2004.
CALL NUMBER: QH430 .L4 2004
BASIC BIOCHEMISTYRY 1. J. Darnell, H. Lodish, D. Baltimore, Molecular Cell Biology (5th ed.) Freeman and Co. 2004.
CALL NUMBER: QH 581.2 D223m 2004 2. B. Alberts, D. Bray. J. Lewis, M. Raff, K. Roberts, J.D. Watson Molecular Biology of the Cell (4th ed.) Garland Publishing 2002. CALL NUMBER: QH581.2 .M64 2002, reserve 3. Christopher K. Mathews, K.E. van Holde, Kevin G. Ahern, BiochemistrySan Francisco, CA : Benjamin Cummings, 2000. CALL NUMBER: QU 4 M4294b 2000 4. L. Stryer, Biochemistry (4th ed.) New York : W.H. Freeman, 1995.
CALL NUMBER: QU 4 S928b 1995 JOURNALS Science, Nature, Cancer Research, Carcinogenesis, Chemical Research in Toxicology, Mutation Research

Oxidative stressApril 25, 27
DNA adducts, structure and activityApril 18, 20
P450 polymorphismsApril 11, 13
Readings in current literatureActivation of chemical carcinogensApr. 6
Ch. 30, sec. 30.3, sec. 30.6-30.11, (sec. 30.14-30.18 optional), 30.19-30.23, (sec. 30.25 and 30.26 optional)
Oncogenes/tumor suppressorsMar. 30, Apr. 4
Ch. 29, sec. 29.25-29.30ApoptosisMar. 28
Cell cycle regulationMar. 23
Ch. 29, sec. 29.1-29.18Cell cycle regulationMar. 21
Spring break, Mar. 10-20
Ch. 28, sec.28.1; sec. 28.5- 28.13 general; sec. 28.14-28.17 Ras pathway
Signal transduction; Ras oncoproteinsMar. 7, 9
Ch. 15, sec. 15.1-15.19 optional, details of recombination; sec. 15.20-15.30
Repair (enzymatic)Feb. 28, Mar. 2
Ch. 7, sec. 7.11-7.18 (suppressors)Repair (non-enzymatic)Feb. 23,
Ch. 11, 12 entirety Transcriptional controlFeb. 16, 21
Ch. 5 (mRNA + processing, rRNA, tRNA); Ch. 6, sec. 6.1, 6.2-6.8, 6.14, 6.15 other sec. optional); Ch. 7, sec. 7.1, 7.2, 7.4, 7.5, other optional)
Transcription/translationFeb. 14
Ch. 9, sec. 9.1-9.17, 9.20; Ch. 21, sec. 21.1-21.20 (promoters and enhancers)
Transcriptional processFeb. 7, 9
Ch. 13, sec. 13.1-13.6, 13.8; Ch. 14DNA replicationJan. 31, Feb. 2
Class notes or Biochem textThermodynamicsJan. 24, 26
Genes VIII, Ch. 1-2 through sec. 2.8Ch. 30, sec. 30.1-30.2
Introduction, chemistry overview, DNA structure.Jan. 12, 17, 19
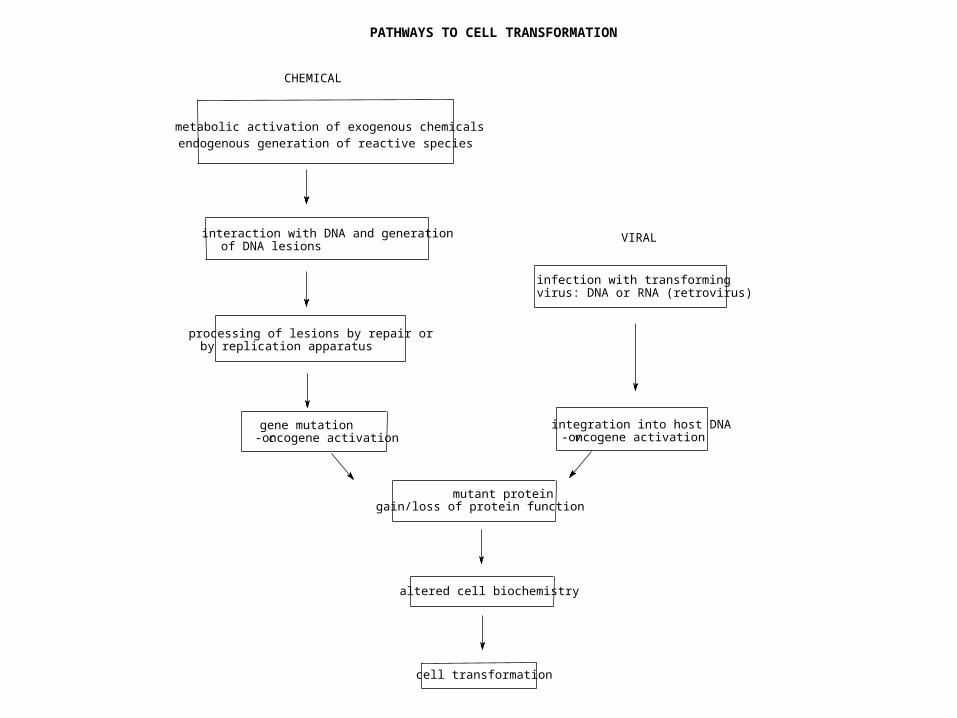
PATHWAYS TO CELL TRANSFORMATION
processing of lesions by repair orby replication apparatus
mutant proteingain/loss of protein function
altered cell biochemistry
cell transformation
infection with transformingvirus: DNA or RNA (retrovirus)
metabolic activation of exogenous chemicalsendogenous generation of reactive species
CHEMICAL
VIRALinteraction with DNA and generationof DNA lesions
gene mutationc-oncogene activation
integration into host DNAv-oncogene activation

CHARACTERISTICS OF TRANSFORMED CELLS
(1) Immortalization and aneuploidy.
(2) Unrestricted growth; loss of density-dependent regulation (or contact inhibition), formation of foci.
(3) Loss of anchorage dependence for growth.
(4) Requirement for growth factor containing serum to sustain growth is absent or reduced.
(5) Cytoskeletal changes.
(6) Dedifferentiation - loss of cell function.
(7) Tumorigenic when injected into syngenetic host.
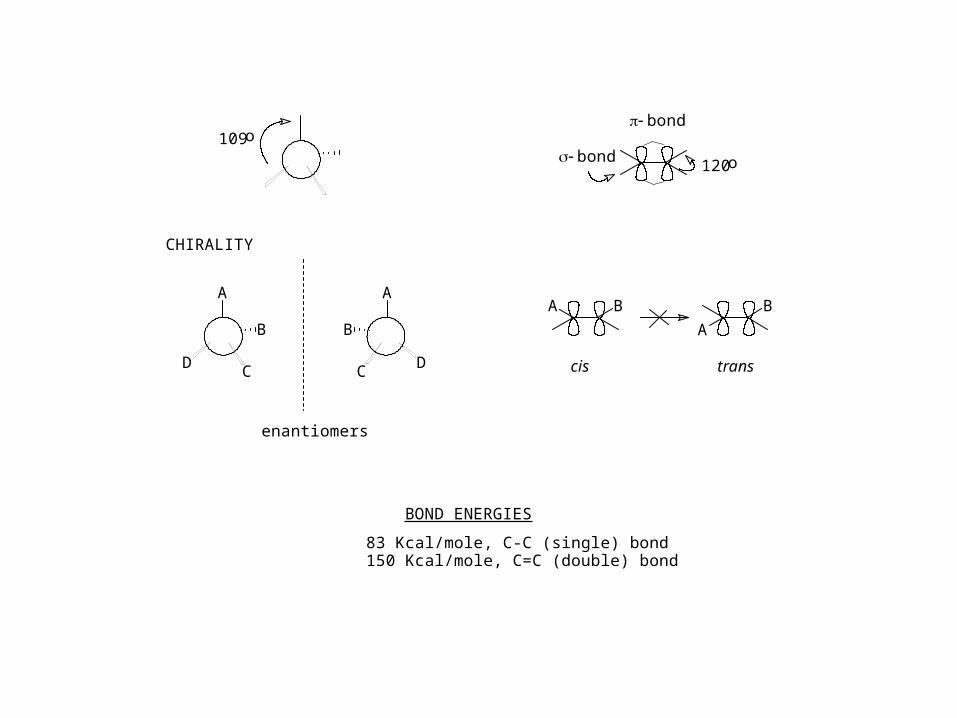
109o
A
B
CD
A
B
C D
B
A
BA
cis trans
bond
bond
120o
CHIRALITY
enantiomers
BOND ENERGIES
83 Kcal/mole, C-C (single) bond150 Kcal/mole, C=C (double) bond
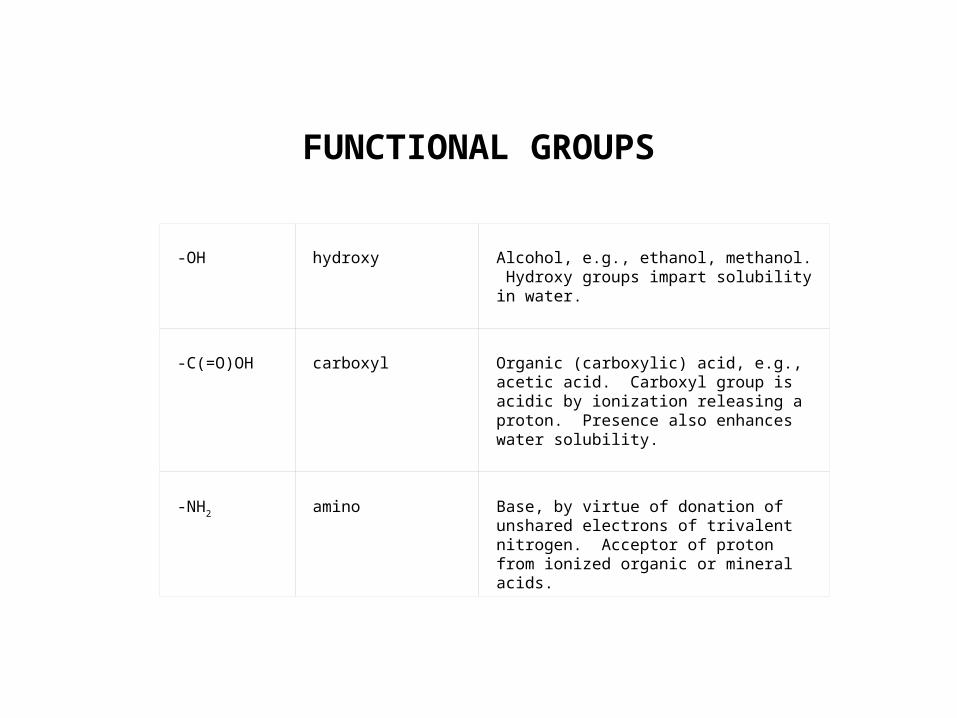
FUNCTIONAL GROUPS
-OH
hydroxy
Alcohol, e.g., ethanol, methanol. Hydroxy groups impart solubility in water.
-C(=O)OH
carboxyl
Organic (carboxylic) acid, e.g., acetic acid. Carboxyl group is acidic by ionization releasing a proton. Presence also enhances water solubility.
-NH2
amino
Base, by virtue of donation of unshared electrons of trivalent nitrogen. Acceptor of proton from ionized organic or mineral acids.
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
HH
OH
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
WATER LATTICE
-
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
HH
OH
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
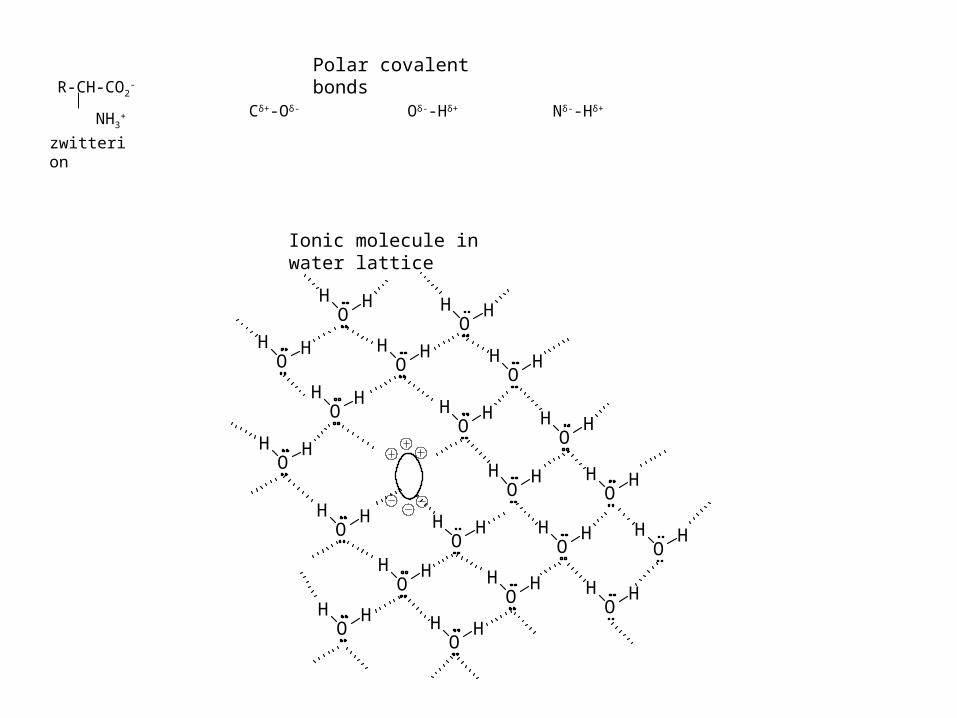
Cδ+-Oδ- Oδ--Hδ+ Nδ--Hδ+
Polar covalent bonds
Ionic molecule in water lattice
R-CH-CO2-
NH3+
zwitterion
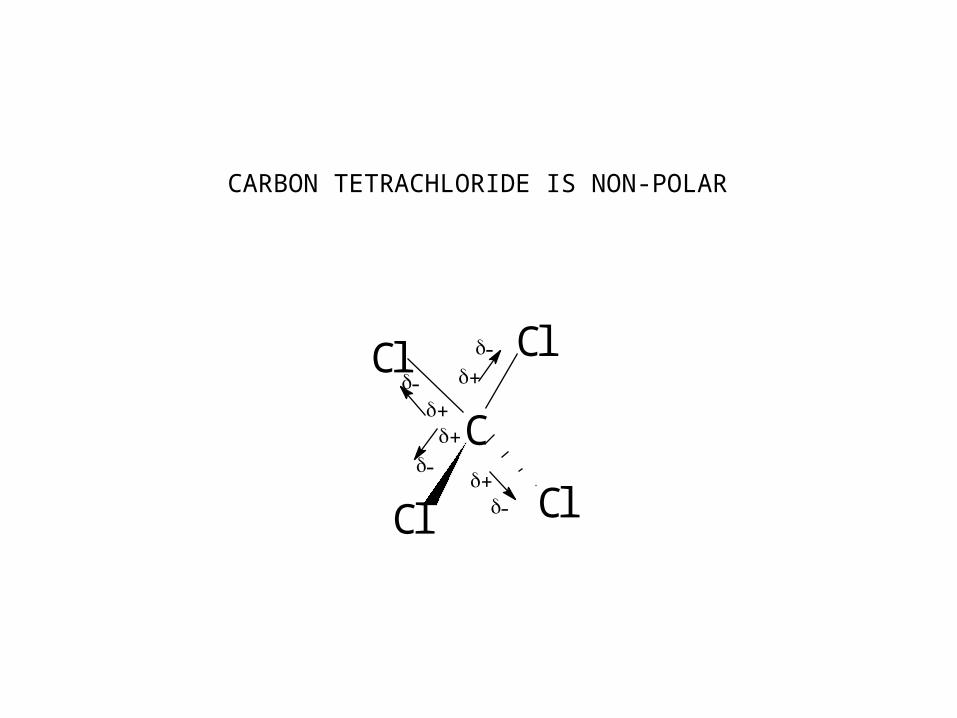
C
Cl Cl
ClCl
-+-
+
-+
-
+
CARBON TETRACHLORIDE IS NON-POLAR
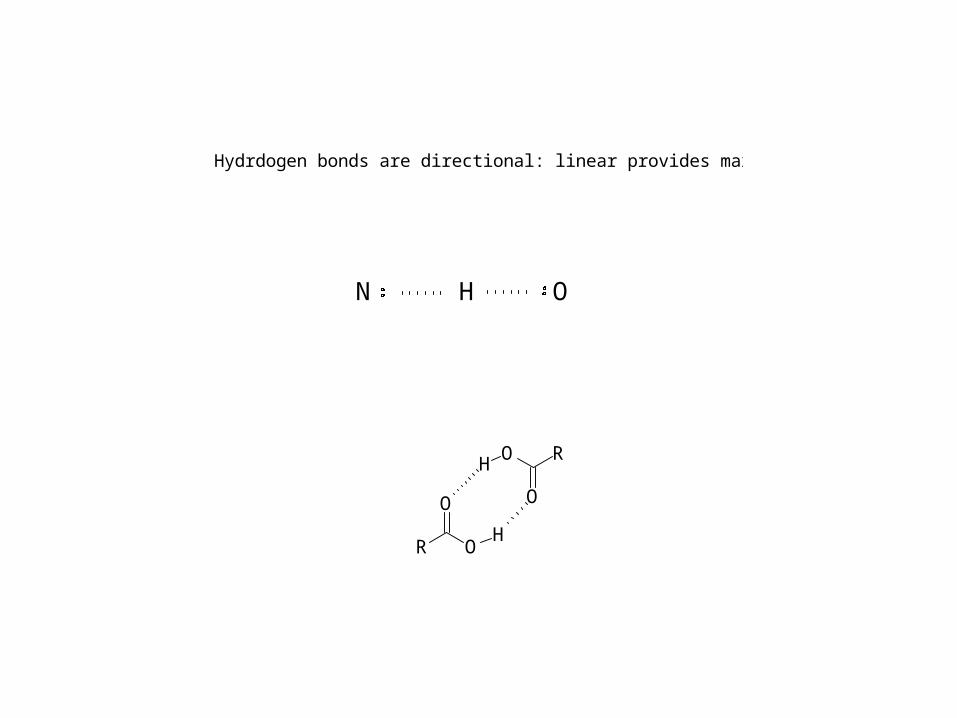
N H O
O
ORH
O
O RH
Hydrdogen bonds are directional: linear provides maximum overlap
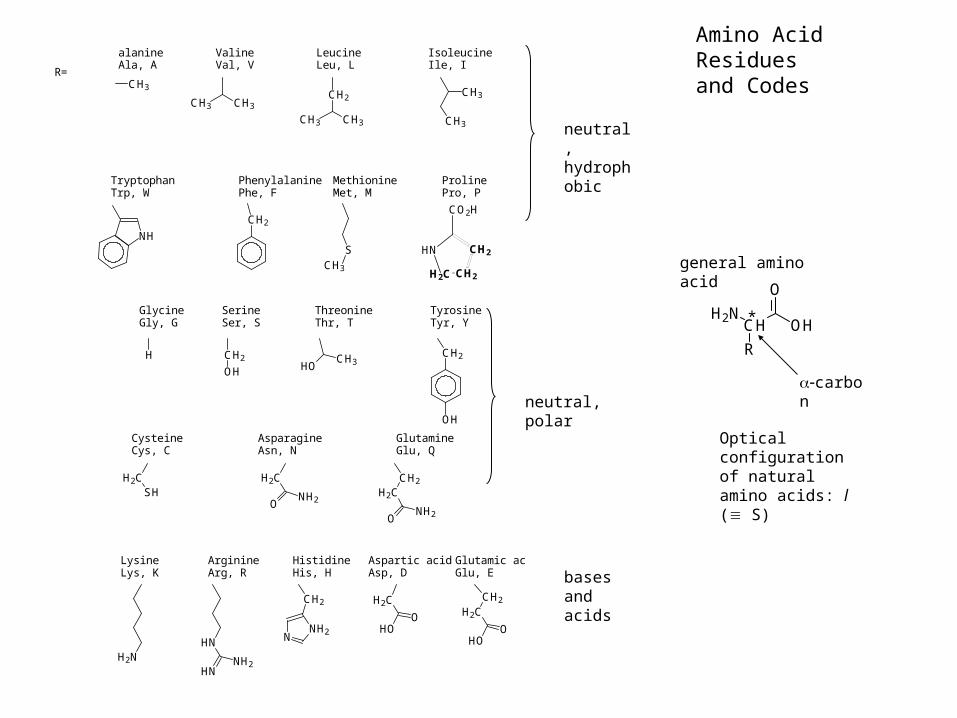
neutral, hydrophobic
neutral, polar
bases and acids
R=
GlycineGly, G
SerineSer, S
ThreonineThr, T
TyrosineTyr, Y
CH2
OHCH3HO
OH
CH2H
H2CSH
H2C
ONH2
CH2
H2C
ONH2
CysteineCys, C
AsparagineAsn, N
GlutamineGlu, Q
H2NHN
HNNH2
NNH2
CH2 H2C
HOO
CH2
H2C
HOO
LysineLys, K
ArginineArg, R
HistidineHis, H
Glutamic acidGlu, E
Aspartic acidAsp, D
CH3
CH3 CH3
CH3 CH3
CH2
CH3
CH3
alanineAla, A
ValineVal, V
LeucineLeu, L
IsoleucineIle, I
TryptophanTrp, W
PhenylalaninePhe, F
MethionineMet, M
ProlinePro, P
NH
CH2
SCH3
HN
H2C CH2
CH2
CO2H
Amino Acid Residues and Codes
OHCHH2N
O
R
*
general amino acid
carbon
Optical configuration of natural amino acids: l ( S)
N
H
O
O
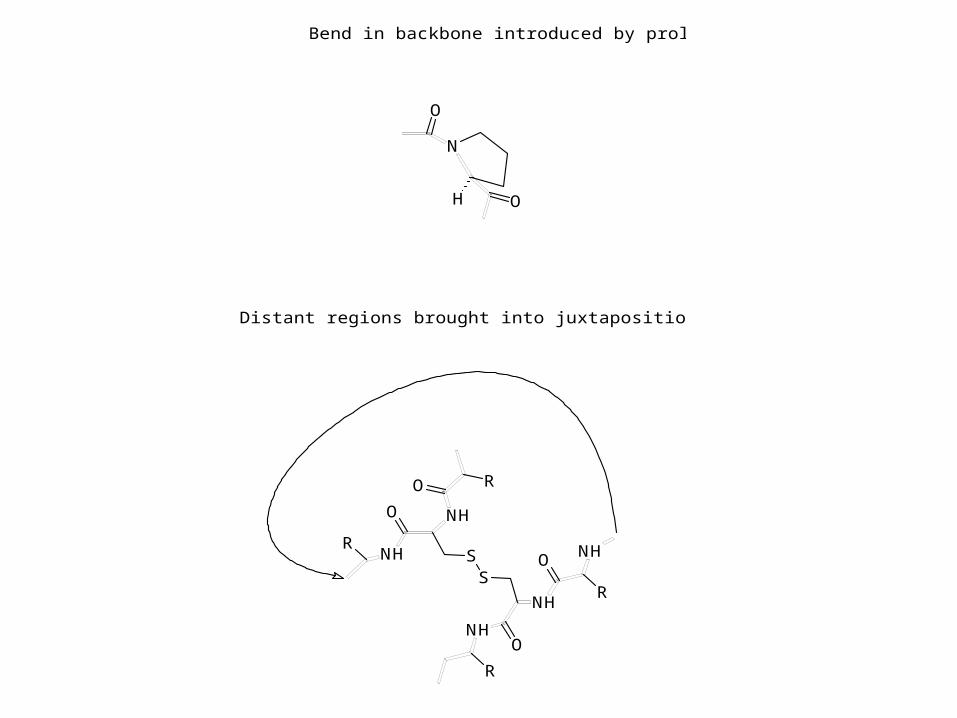
Bend in backbone introduced by proline
SS
NH
NH
NH
NH
NH
O
O
R
R
O
O R
R
Distant regions brought into juxtaposition by disulfide bond
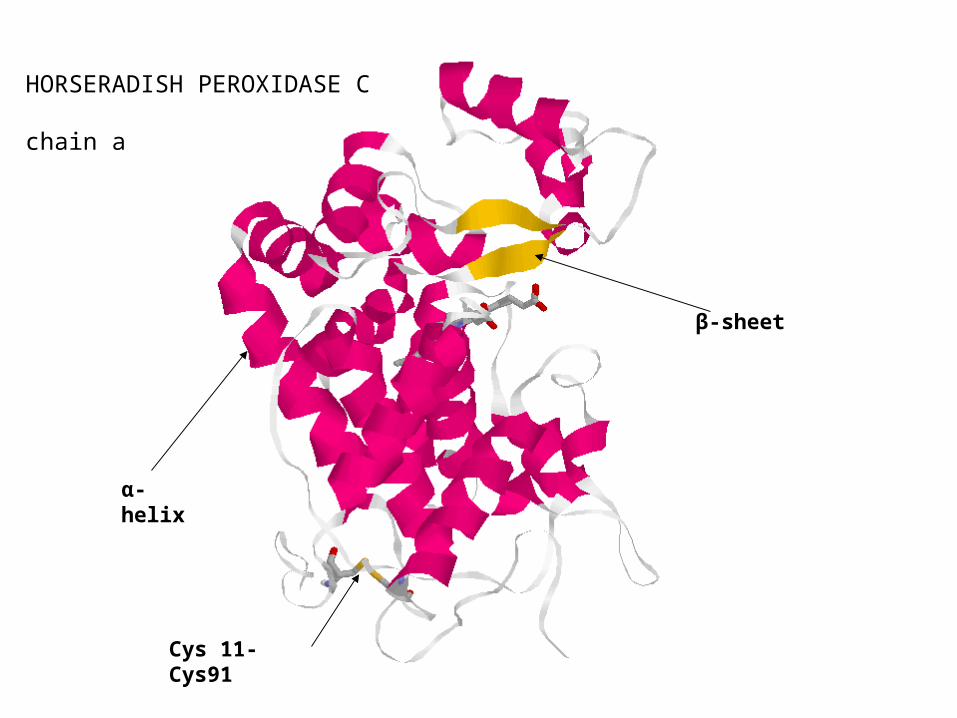
HORSERADISH PEROXIDASE C chain a
α-helix
β-sheet
Cys 11-Cys91
N
N
NH2
OO
HOOH
H N
N
O
H2 N N
N
O
H OOH
HO PO
O -O ][[ ] HO P
O
O -O ][[ ]
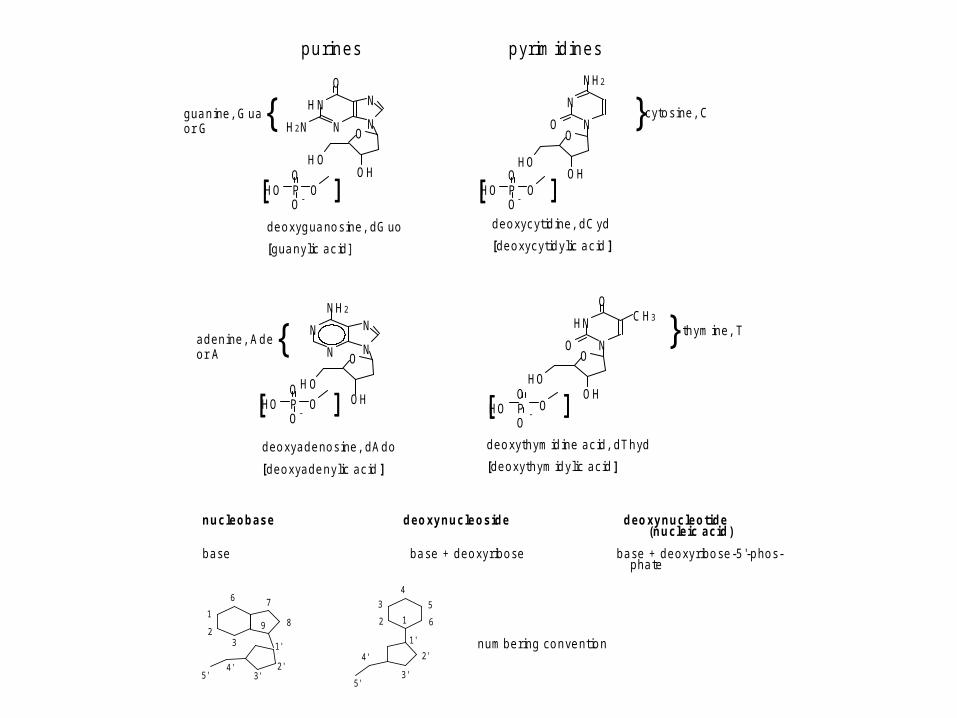
d e o xyg ua no s ine , d G uo
[g ua n y lic a c id ]
N
N
NH2
N
N
O
H OOHHO P
O
O -O ][[ ]
HN
N
O
O
C H3
O
HOOH
HO PO
O-
O ][[ ]
d e o xyc yt id ine , d C yd
[d e o xyc yt id y lic a c id ]
g ua n ine , G ua {o r G} c yto s ine , C
a d e n ine , A d e {o r A
d e o xya d e no s ine , d A d o
[d e o xya d e n y lic a c id ]
} th ym ine , T
d e o xyth ym id ine a c id , d T h yd
[d e o xyth ym id y lic a c id ]
n u c le o b a s e d e o xy n u c le o s id e d e o xy n u c le o tid e(n u c le ic a c id )
b a se b a se + d e o x y r ib o se b a se + d e o xy r ib o se -5 '-p ho s-p ha te
1
23
6 7
89
1'
2'3'
4'5'
12
3
4
5
6
1'
2'
3'
4'
5'
n um b e r ing c o n ve nt io n
p u r in e s p yr im id in e s
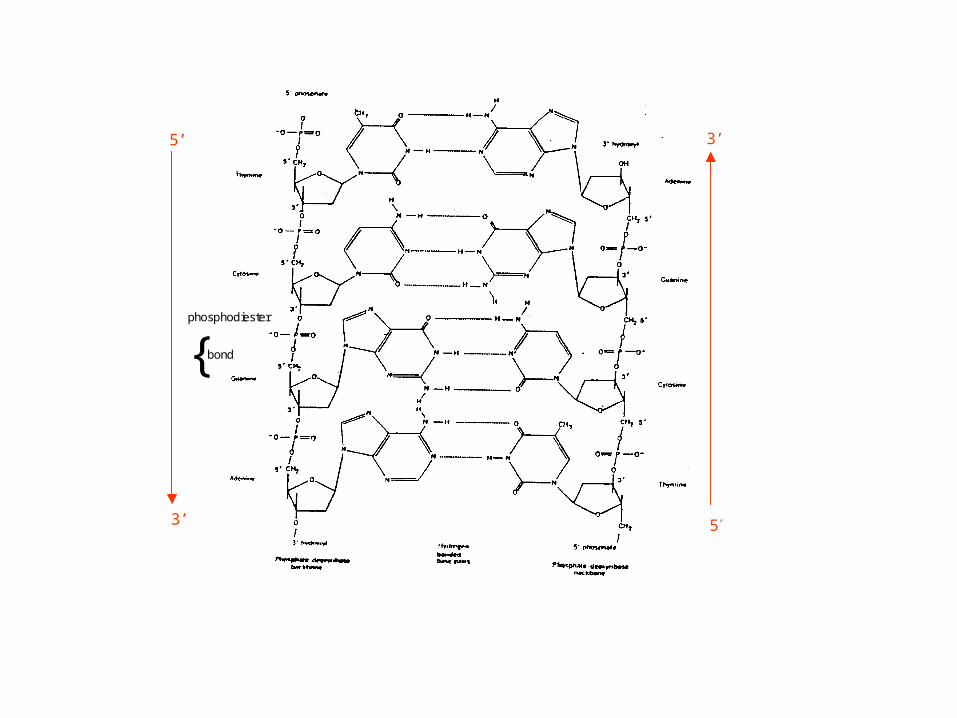
phosphodiester
{ bond
3’
53’
5’
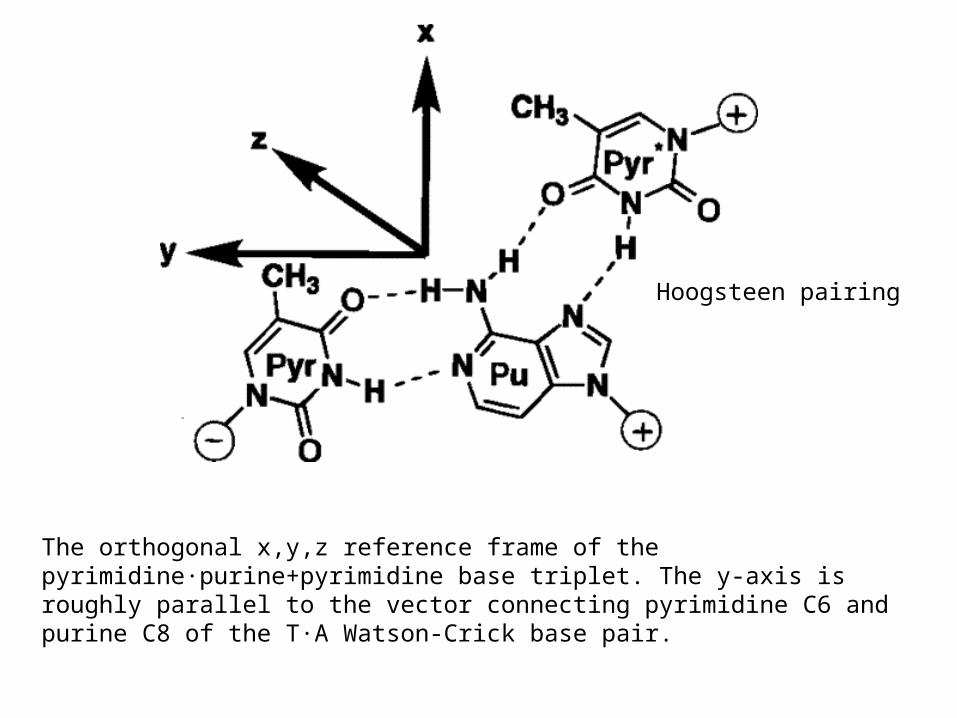
The orthogonal x,y,z reference frame of the pyrimidine·purine+pyrimidine base triplet. The y-axis is roughly parallel to the vector connecting pyrimidine C6 and purine C8 of the T·A Watson-Crick base pair.
Hoogsteen pairing
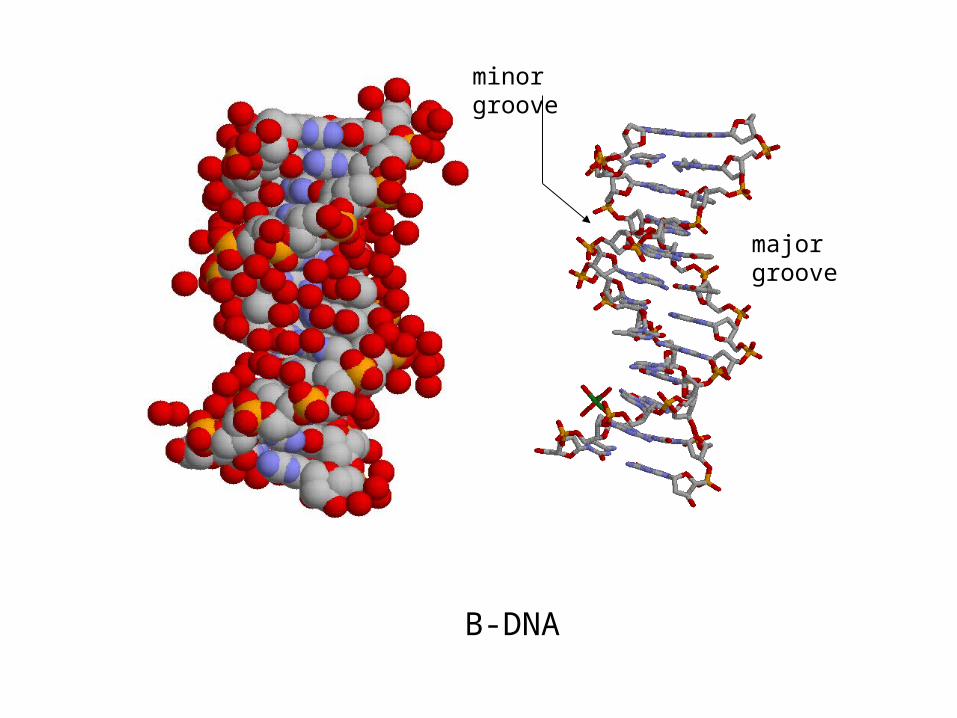
major groove
minor groove
B-DNA
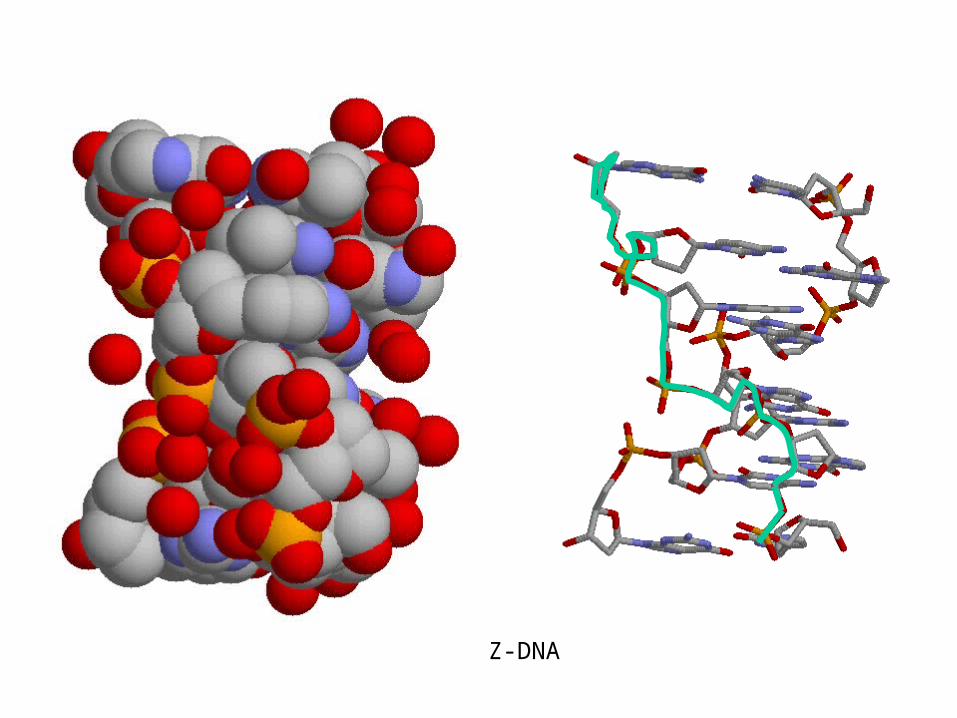
Z-DNA
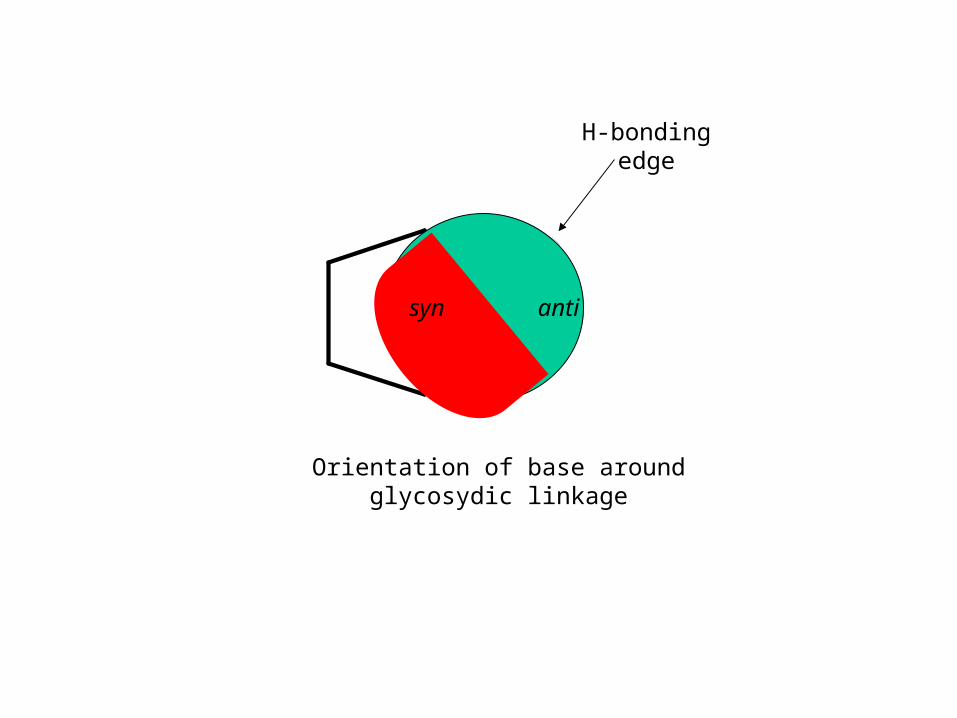
H-bonding edge
antisyn
Orientation of base around glycosydic linkage
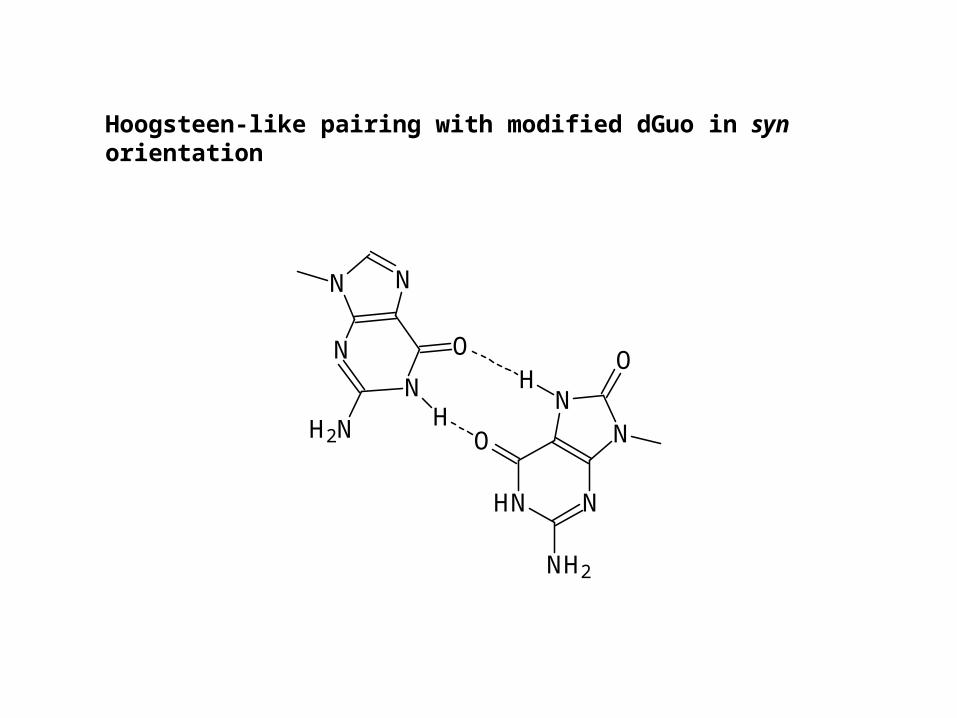
Hoogsteen-like pairing with modified dGuo in syn orientation
N
N
NN
O
H2N H
HO
NH2
O NN
NHN
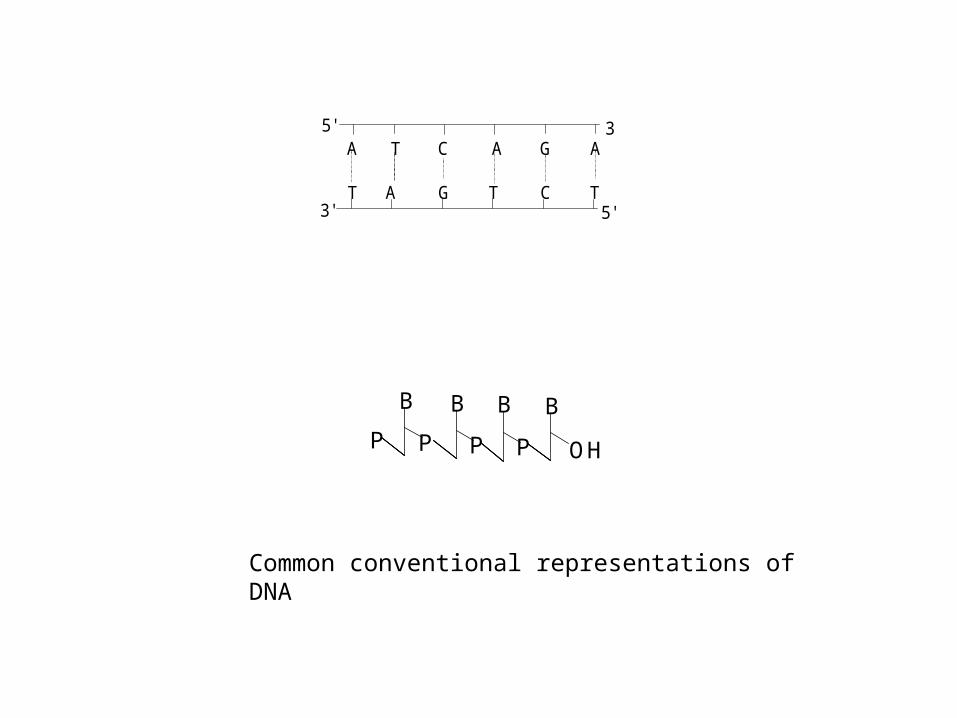
A T C A G A
T A G T C T
5' 3'
3' 5'
B
PP
B
P
B
P
B
OH
Common conventional representations of DNA
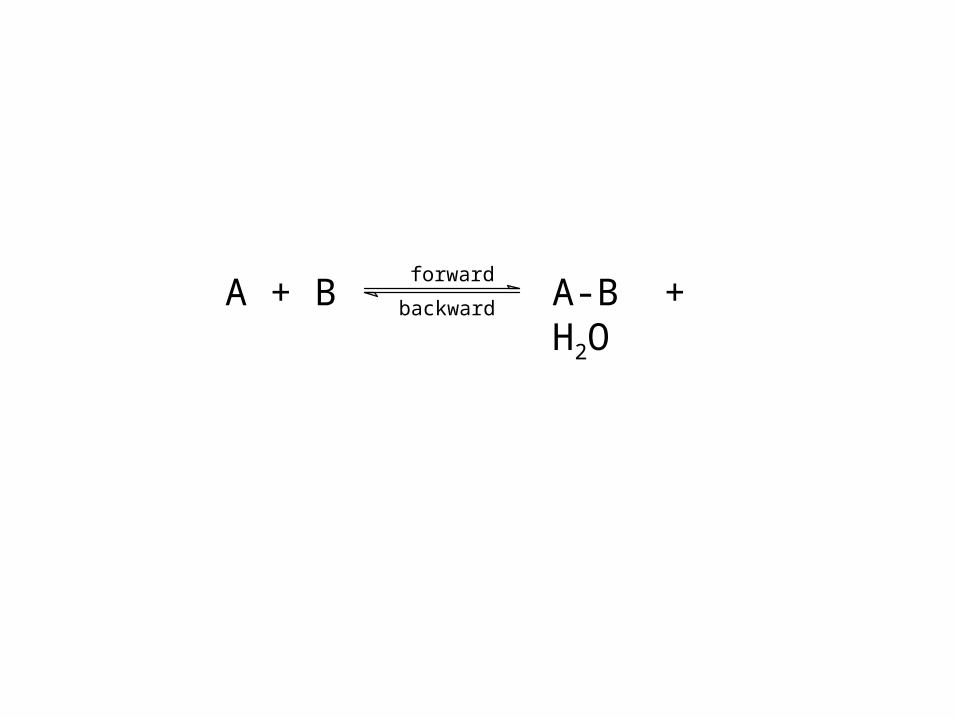
forward
backwardA + B A-B + H2O

EQUATIONS FOR THERMODYNAMICS H ≡ enthalpyE ≡ internal energyP ≡ pressureV ≡ volumeChange in enthalpy: ΔH = ΔE + P ΔV S ≡ entropy Change in free energy: ΔG = ΔH – TΔS For the reaction as written: ΔG < 0, spontaneousΔG > 0, not spontaneous- work must be put into the system to drive it in the forward directionΔG = 0, the system is in equilibrium K ≡ equilibrium constant, ratio of concentrations of products to reactants:
ΔG = ΔGo + RTln KR ≡ gas constant (= 1.98 cal/mole-oK = 0.00198 kcal/mole-oK)T in oKΔGo = ΣGo
products - ΣGoreactants at Pstd = 1 atm, Tstd = 25o C (biochem.) or 0o C (physical chem.)
At equilibrium, ΔG = 0, the expression becomes:0 = ΔGo + RTln K or ΔGo = -RT ln K
Superscript “o” is dropped, the relationship written as:ΔG = -RT ln K
]][[
]][[ 2
BA
OHBAK
forward
backwardA + B A-B + H2O

ΔG = -RT ln K ΔG = +6 kcal/moleR = 0.00198 kcal/mole-oKT = (25 + 273) o K = 298 oK 6 kcal/mole = -(0.00198kcal/mole-oK)(298 oK)ln K ln K = -6/(1.98 x 10-3)(298) = -10.2K = e-10.2 = 3.83 x 10-5
K= 3.83 x 10-5 = [p-dN-p-dN][H2O]/[p-dN][p-dN]
Initial dinucleotide concentration [p-dN-p-dN1 x 10-3 MVirtually all the dimer will disappear; therefore, approximate the product nucleotides as [p-dN] = [p-dN] 1 x 10-3 M Exact expression is [p-dN] = [p-dN] = (1 x 10-3 –x) [dimer] = x
[H20] ≈ constant = 55.6 M
[x][55.6]/[1 x 10-3][1 x 10-3] = 3.8 x 10-5
[x] = (3.8 x 10-5)(1 x 10-3)2/55.6 = 6.8 x 10-13 M
p-dN + p-dN p-dN-p-dN + H2O ΔG = +6 kcal/mole
Dinucleotide from 5-deoxynucleotide phosphates
Q: What is the equilibrium constant for the formation of a dinucleotide from 5-phosphates?
Q: What is the equilibrium concentration of dinucleotide from a 1 x 10-3 M initial concentration?

ATP + H2O ADP + Pi ΔG = -7 kcal/mole
ADP = adenosine diphosphate
Pi = inorganic phosphate group
ATP is sometimes written as ADP~P to emphasize high energy of the phosphate bond
The first stage in polynucleotide synthesis is the transfer of a high-energy bond to p-dN in two steps:
ATP + p-dN ADP + dNDP
ATP + dNDP ADP + dNTP ΔG ~< 0
p-dN′ + p3-dN p-dN′-p-dN + p-p ΔG = +0.5 kcal/mole
p-p + H2O 2Pi ΔG = -7 kcal/mole
p-dN′ + p3-dN + H2O p-dN′-p-dN + 2Pi ΔG = (+0.5 - 7.0)kcal/mole = -6.5 kcal/mole
N
N
N
N
O
OHOH
O
NH2
P
O
O-
OP
O
O-
OP
O
O-
O-
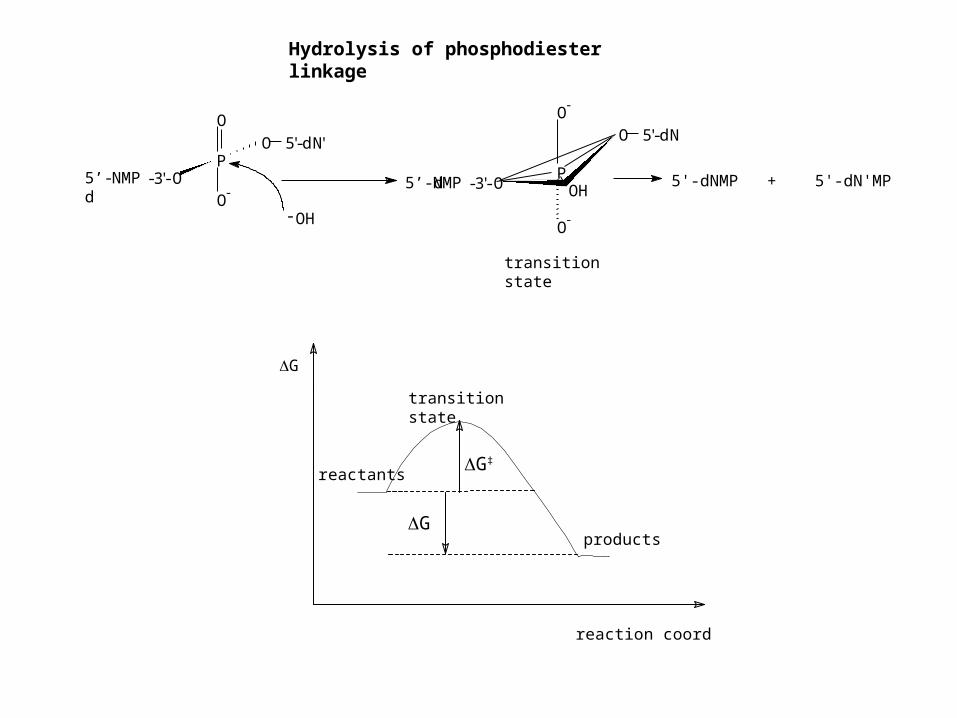
O-
P
O
O
5’-dNMP-3'-O
5'-dN'
P
O-
O-
O 5'-dN
5’-dNMP-3'-O OH
-OH
5'-dNMP + 5'-dN'MP
transition state
Hydrolysis of phosphodiester linkage
products
reactants
reaction coordinate
G
G‡
G
transition state
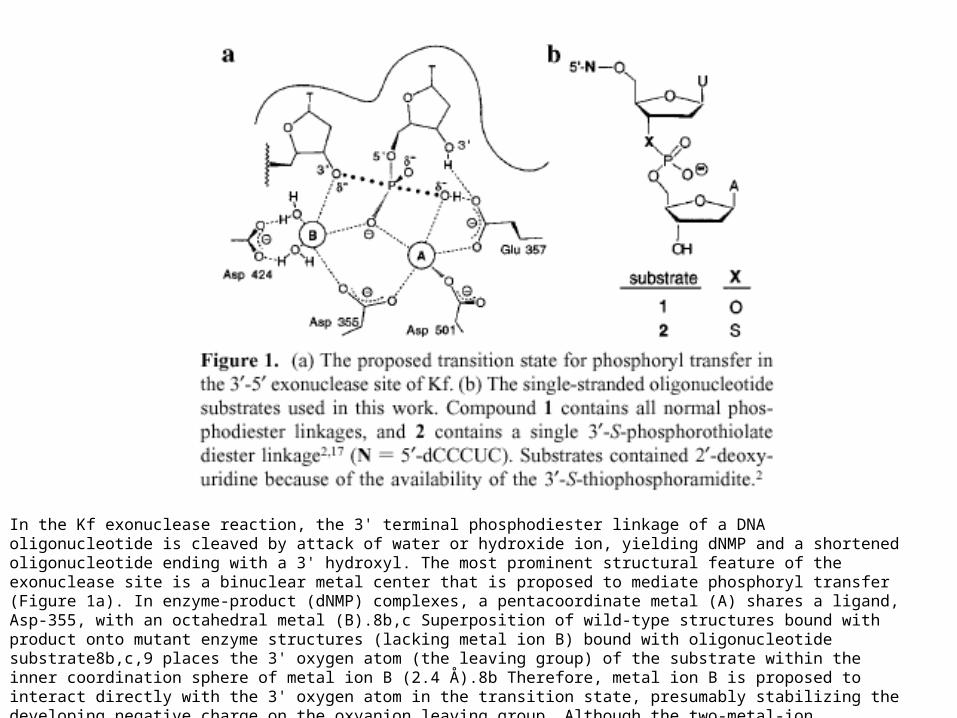
In the Kf exonuclease reaction, the 3' terminal phosphodiester linkage of a DNA oligonucleotide is cleaved by attack of water or hydroxide ion, yielding dNMP and a shortened oligonucleotide ending with a 3' hydroxyl. The most prominent structural feature of the exonuclease site is a binuclear metal center that is proposed to mediate phosphoryl transfer (Figure 1a). In enzyme-product (dNMP) complexes, a pentacoordinate metal (A) shares a ligand, Asp-355, with an octahedral metal (B).8b,c Superposition of wild-type structures bound with product onto mutant enzyme structures (lacking metal ion B) bound with oligonucleotide substrate8b,c,9 places the 3' oxygen atom (the leaving group) of the substrate within the inner coordination sphere of metal ion B (2.4 Å).8b Therefore, metal ion B is proposed to interact directly with the 3' oxygen atom in the transition state, presumably stabilizing the developing negative charge on the oxyanion leaving group. Although the two-metal-ion mechanism of Kf is thought to be a general strategy by which many protein enzymes and ribozymes catalyze phosphoryl transfer,8a,10 there is no direct biochemical evidence that the 3'-5' exonuclease employs a metal ion in this role.
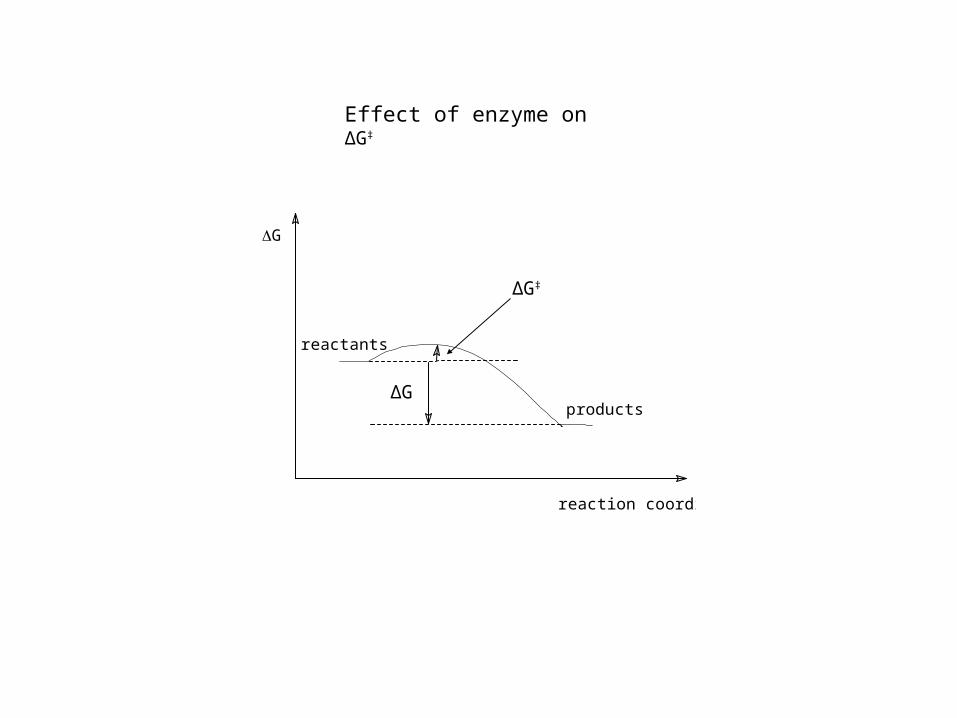
G
reaction coordinate
reactants
products
ΔG‡
ΔG
Effect of enzyme on ΔG‡
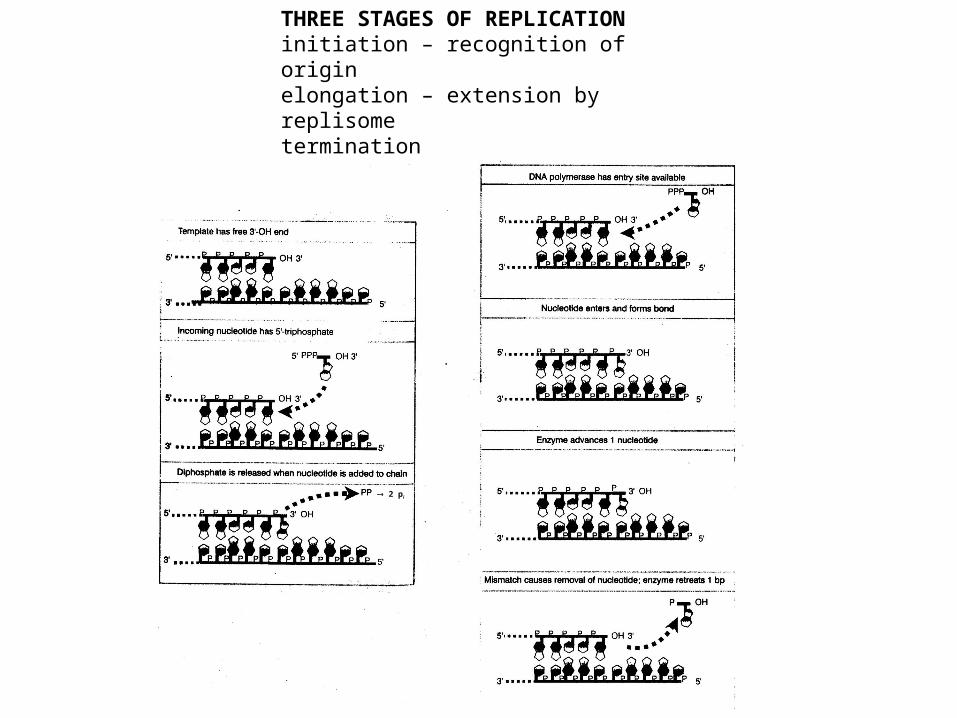
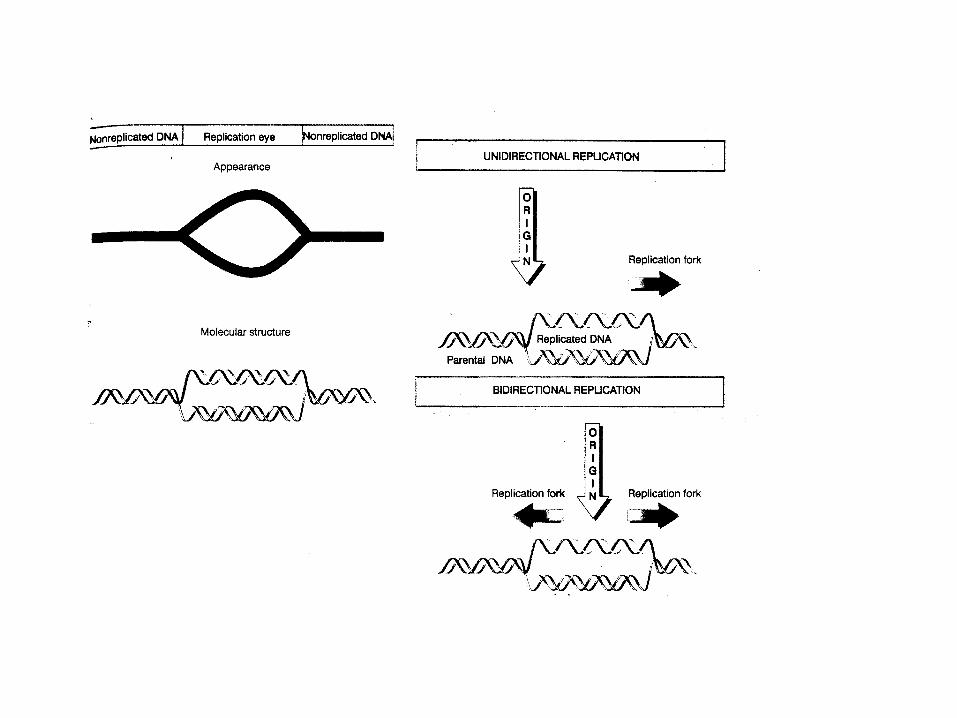
THREE STAGES OF REPLICATIONinitiation – recognition of originelongation – extension by replisometermination
2 pi
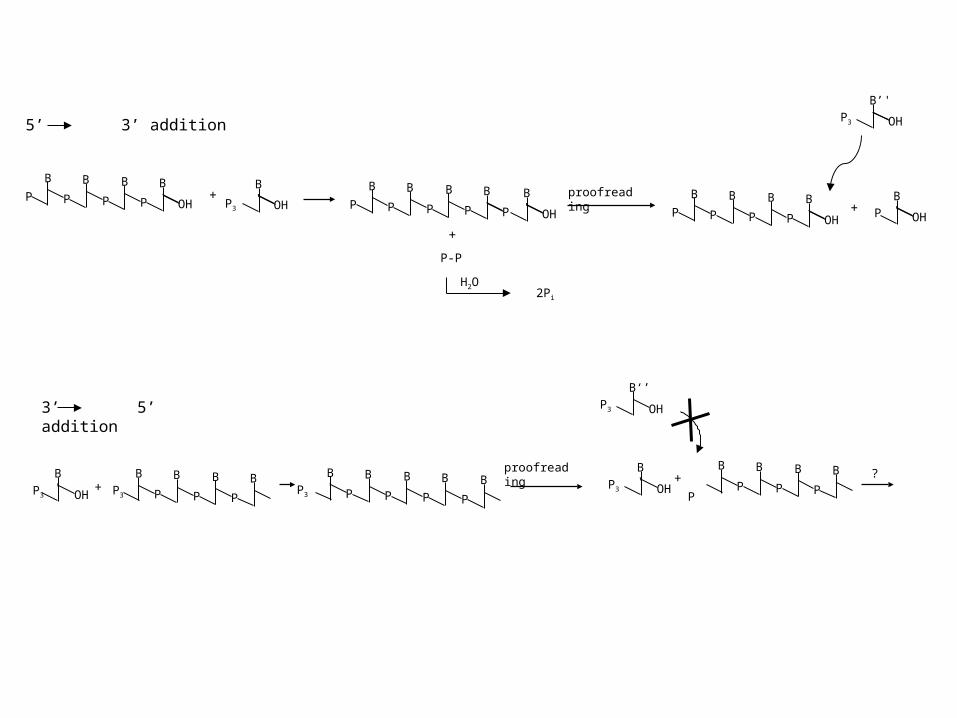
+OH
B'
P3
proofreading
OH
B'
P3+ ?
B
P P
B
P
B B
OHP
P3
B'
OH+
B
P P
B
P
B B
PP
B'
OH
proofreading B
P P
B
P
B B
OHP +
B'
OHP
5’ 3’ addition
3’ 5’ addition
+
P-P
H2O2Pi
P P
B B
P
B B P3 P
BP P
B' B
P
B BP3
P P
B B
P
B B P
B’'
OHP3
OH
B‘’
P3

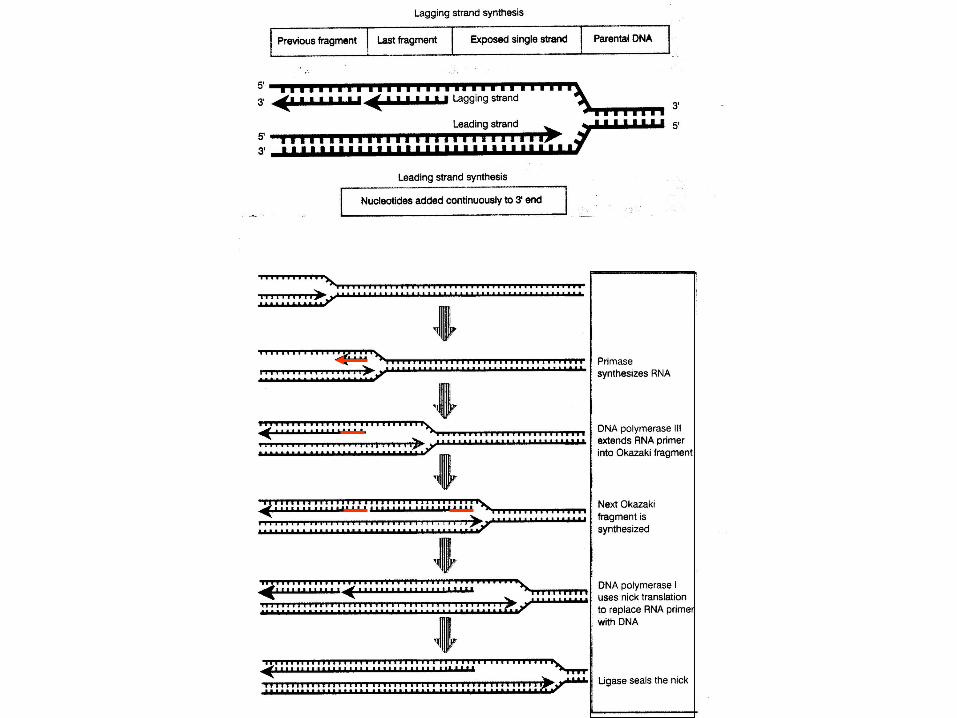
pol I, 5'3' synthesis + 3'5' exonuclease, unique 5'3' exonuclease capability. Pol I responsible for repair, since 5'3' exonuclease activity allows pol I to extenda strand from a nick in DNA. (Nick: strand break caused by hydrolysis of phosphodiester
bond.) pol II, 5'3' synthesis + 3'5' exonuclease, also is involved in repair. pol III, large multi-unit enzyme 5'3' synthesis + 3'5' exonuclease, primarily involved in strand
extension during replication.
α, 5'3' synthesis but no 3'5' exonucleaseβ, 5'3' synthesis with no 3'5' exonucleaseδ, 5'3' synthesis + 3'5' exonucleaseε, 5'3' synthesis + 3'5' exonucleaseγ, 5'3' synthesis + 3'5' exonucleaseα -ε are located in the nucleus, and γ in mitochondria.
α initiates strand synthesis, δ is responsible for strand extension, ε and β are involved in repairwhile γ is responsible for replication of mitochondrial DNA
PROKARYOTIC POLYMERASES
EUKARYOTIC POLYMERASES
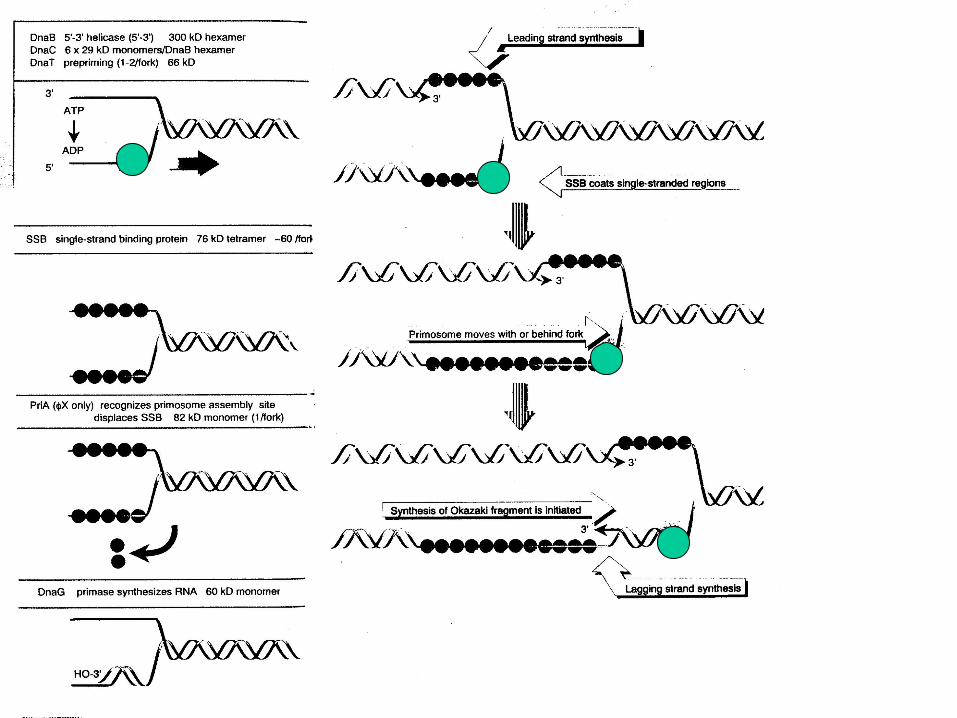
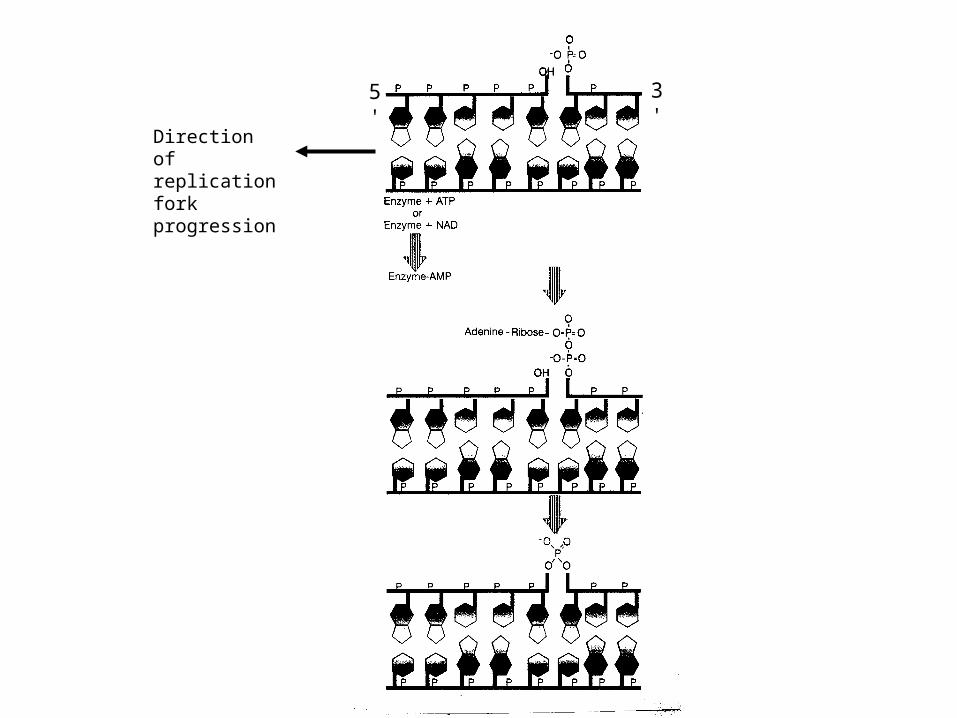
5' 3'
Direction of replication fork progression
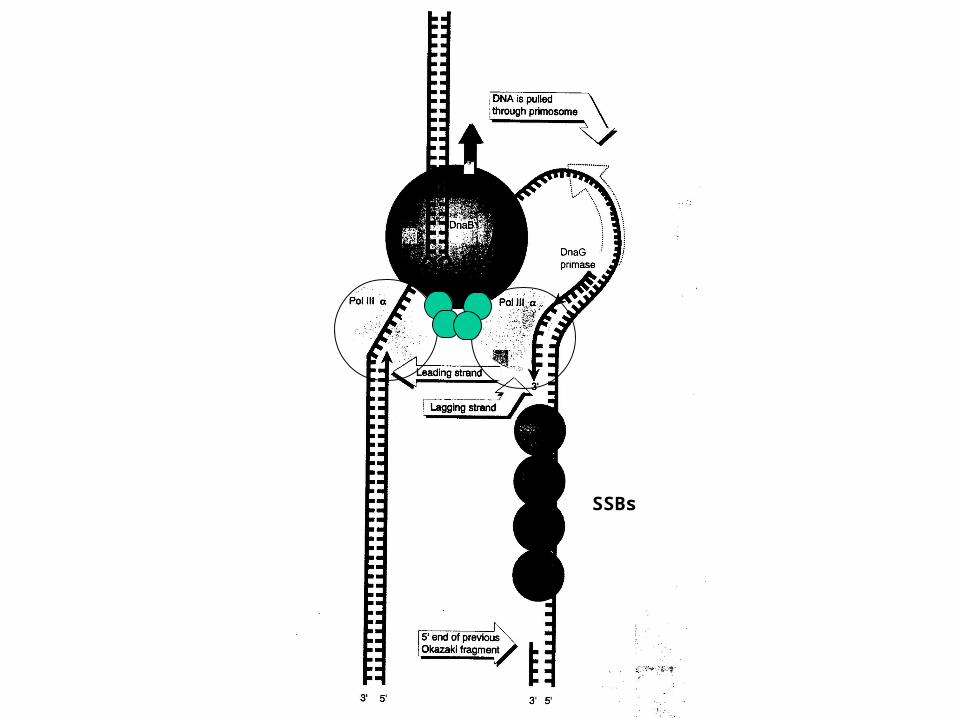
SSBs
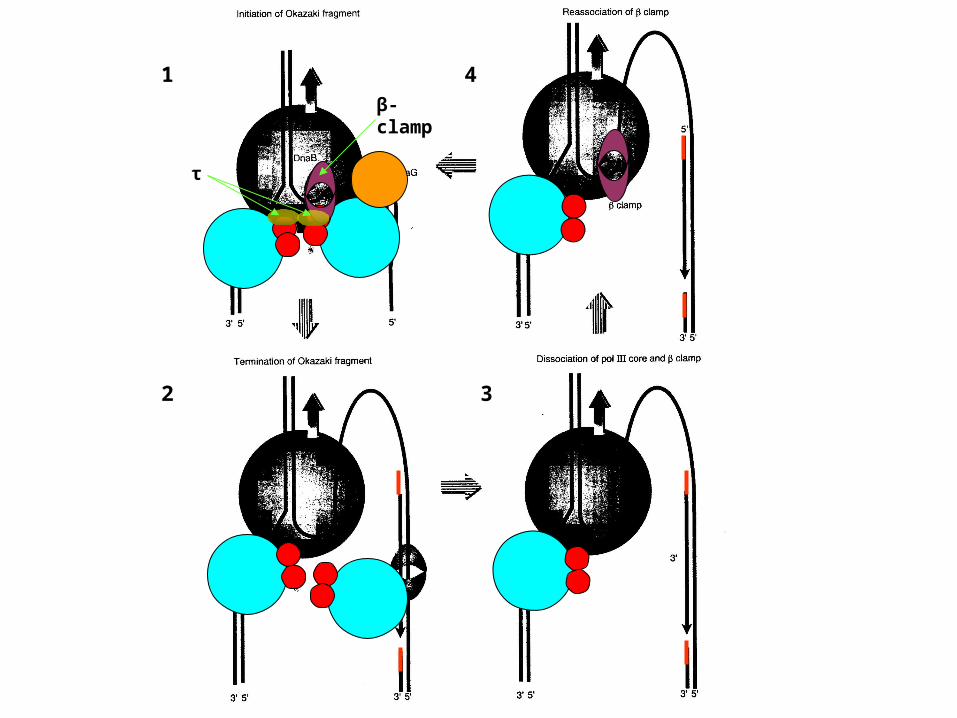
1
2 3
4
τ
β-clamp

DNA pol α RNA priming + short 3 – 4 base DNA extension (iDNA; i = initiation)
DNA pol δ Strand extensionPCNA (proliferating cell nuclear antigen) Processivity (equivalent function to β-clamp)RFC (replication factor C) Loads pol δ and PCNA at end of iDNAFEN1, Dna2 (5 3 exonuclease) Removal of RNA primerDNA ligase I Seal nicksRPA Single strand binding proteinsMCM Helicase function
Some Eukaryotic Replication Proteins
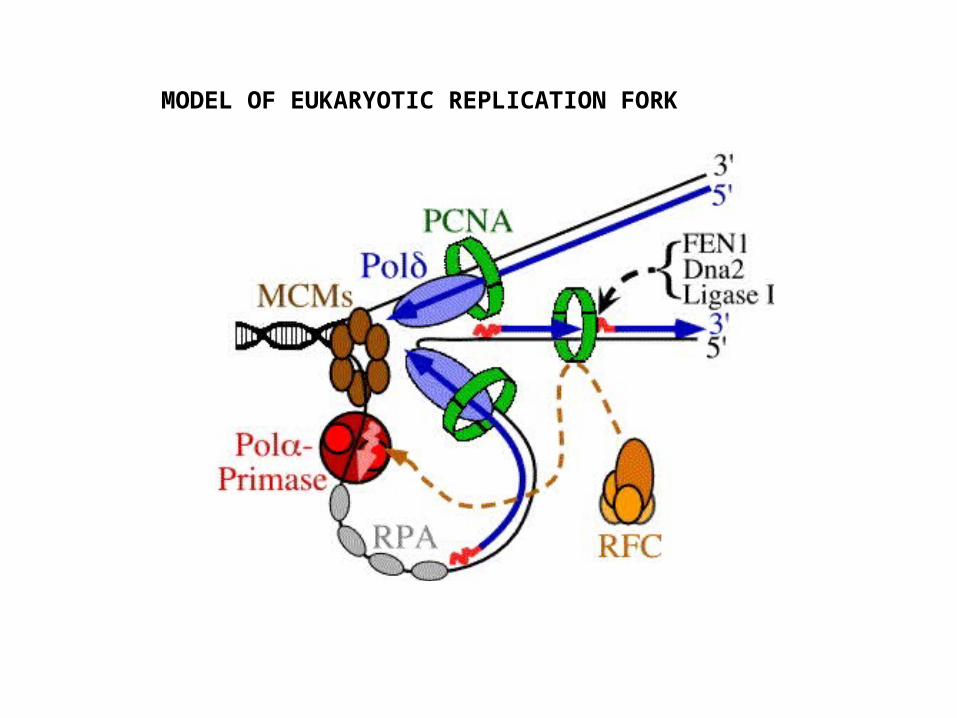
MODEL OF EUKARYOTIC REPLICATION FORK
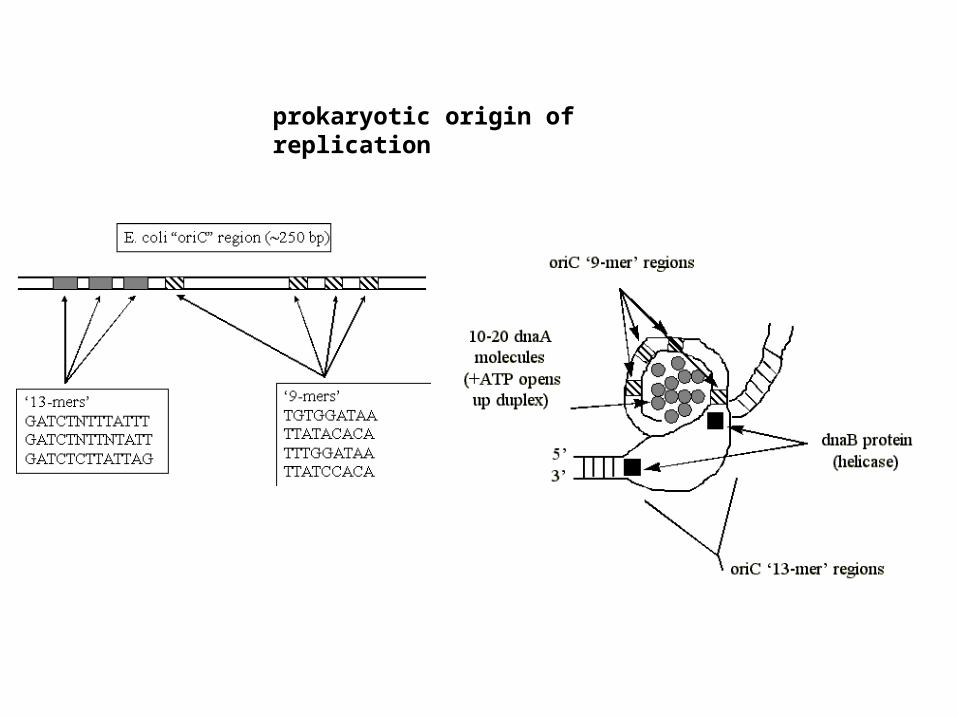
prokaryotic origin of replication

G (*A) T C
C T (*A) G
G (*A) T C
C T (A) G
control of replication at prokaryotic origins
parent duplex parent + daughter duplex
*A =
fully methylated hemi-methylated
N
NNH
N
HN
CH3
N6-MeAde
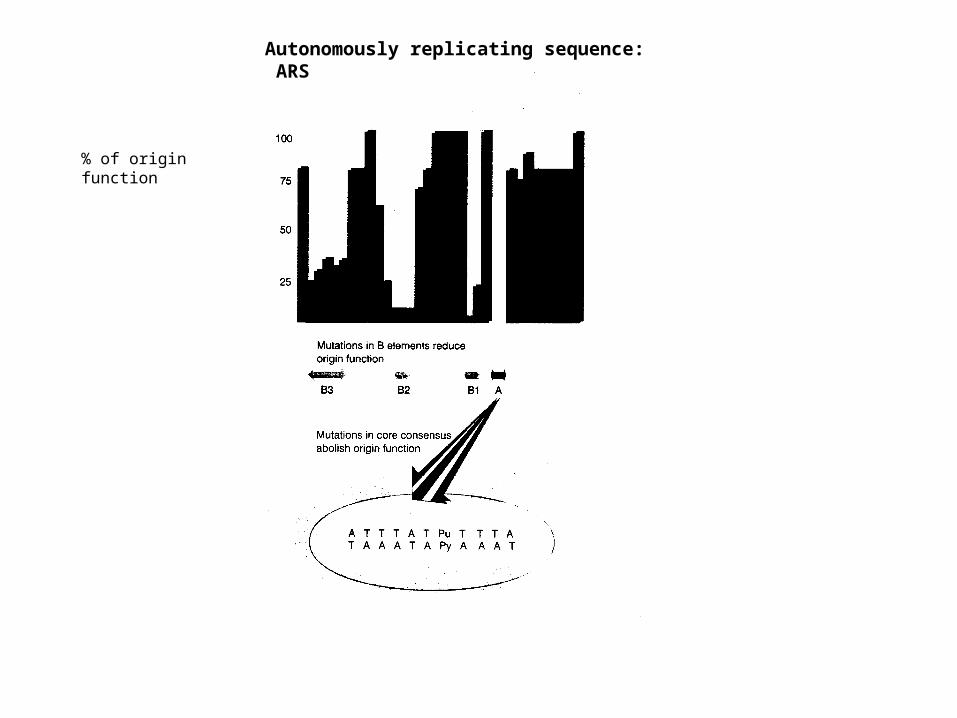
% of origin function
Autonomously replicating sequence: ARS
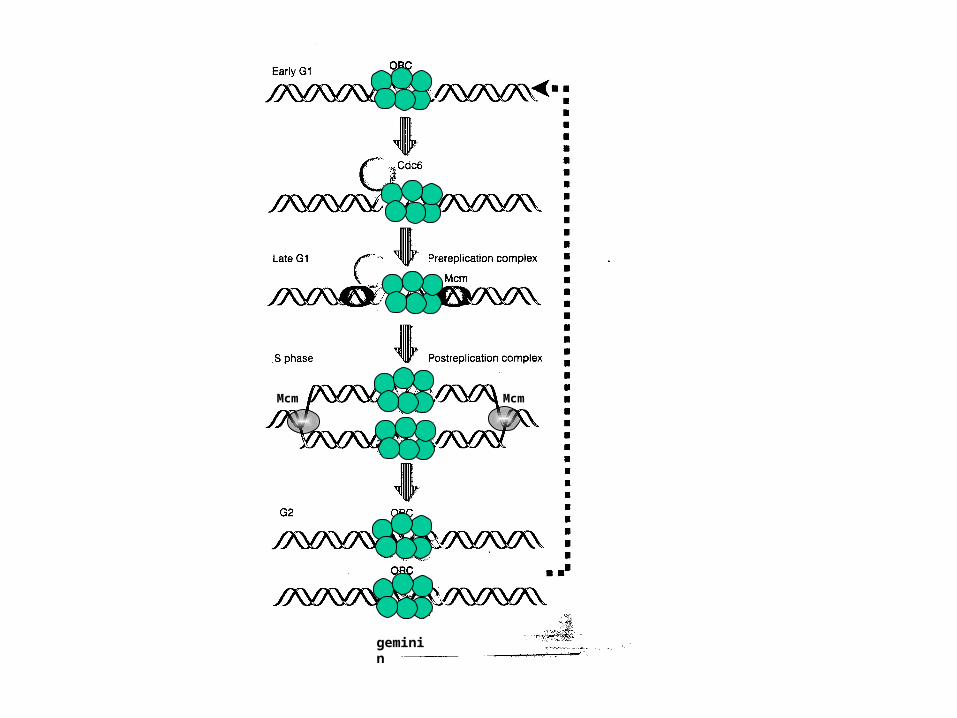
McmMcm
geminin
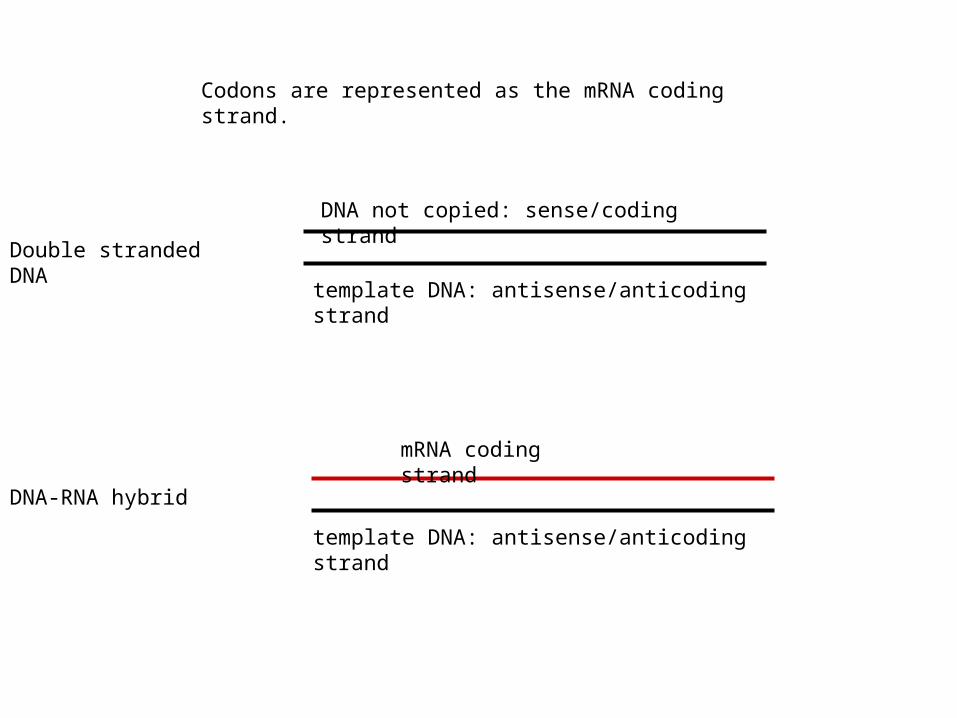
Double stranded DNA
template DNA: antisense/anticoding strand
DNA not copied: sense/coding strand
DNA-RNA hybrid
mRNA coding strand
template DNA: antisense/anticoding strand
Codons are represented as the mRNA coding strand.
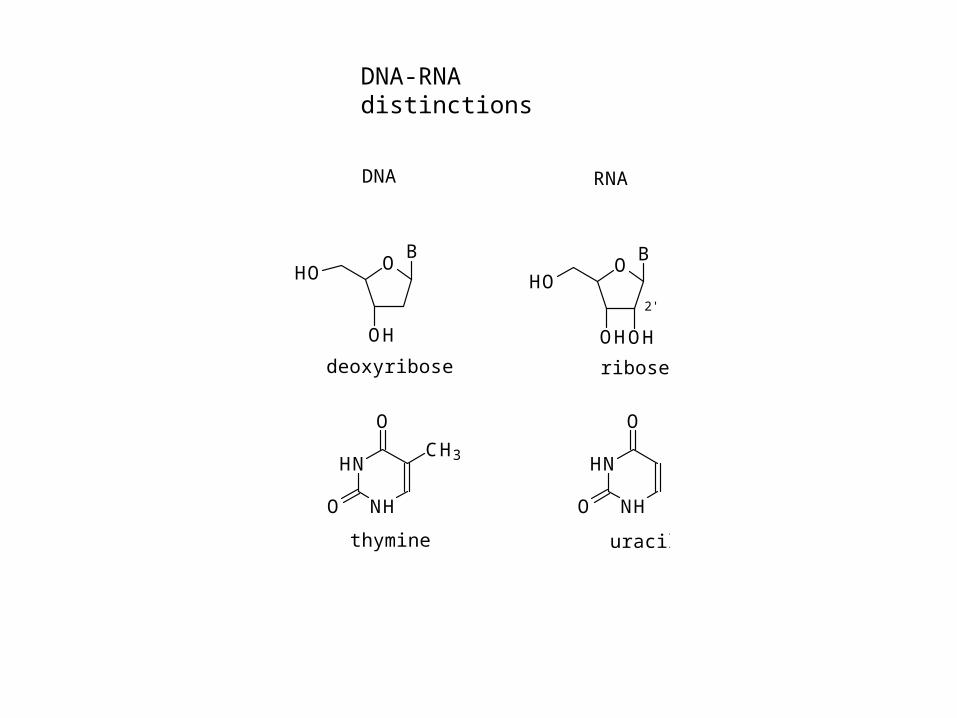
OB
OH
HO
DNA RNA
HN
NH
O
O
CH3HN
NH
O
O
thymine uracil
deoxyribose ribose
OB
OHOH
HO2'
DNA-RNA distinctions
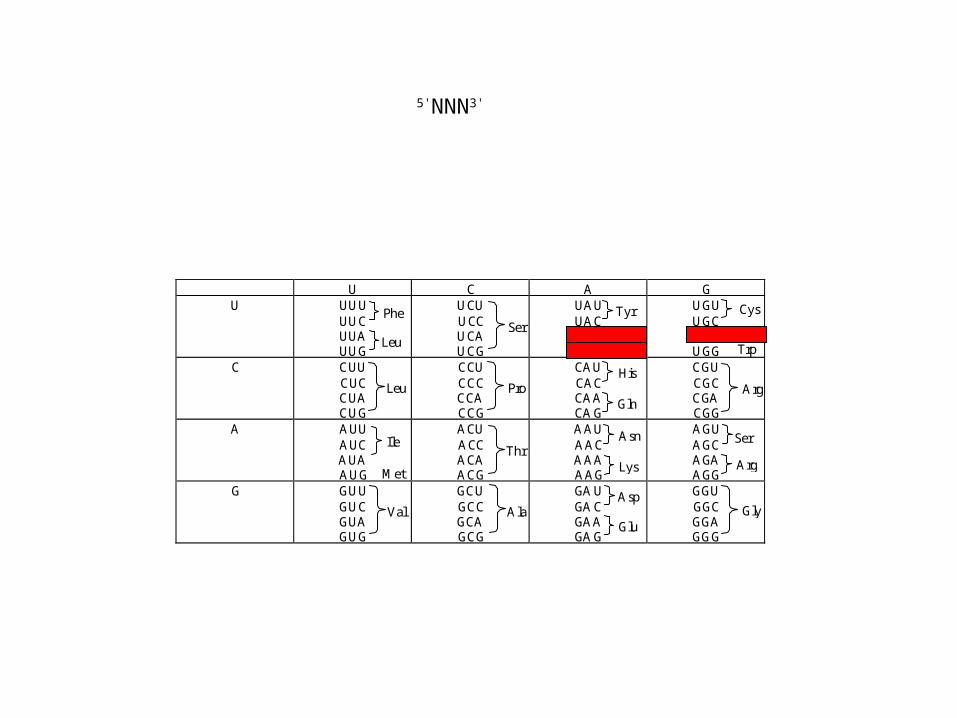
U C A GU UUU
UUCUUAUUG
UCUUCCUCAUCG
UAUUACUAAUAG
UGUUGCUGAUGG
C CUUCUCCUACUG
CCUCCCCCACCG
CAUCACCAACAG
CGUCGCCGACGG
A AUUAUCAUAAUG
ACUACCACAACG
AAUAACAAAAAG
AGUAGCAGAAGG
G GUUGUCGUAGUG
GCUGCCGCAGCG
GAUGACGAAGAG
GGUGGCGGAGGG
Phe
Leu
Leu
Ile
Met
Val
Ser
Ser
Ala
Pro
Thr
Tyr
STOP
His
Gln
Asn
Lys
Asp
Glu
Cys
Trp
Arg
Arg
Gly
STOP
5'NNN3'
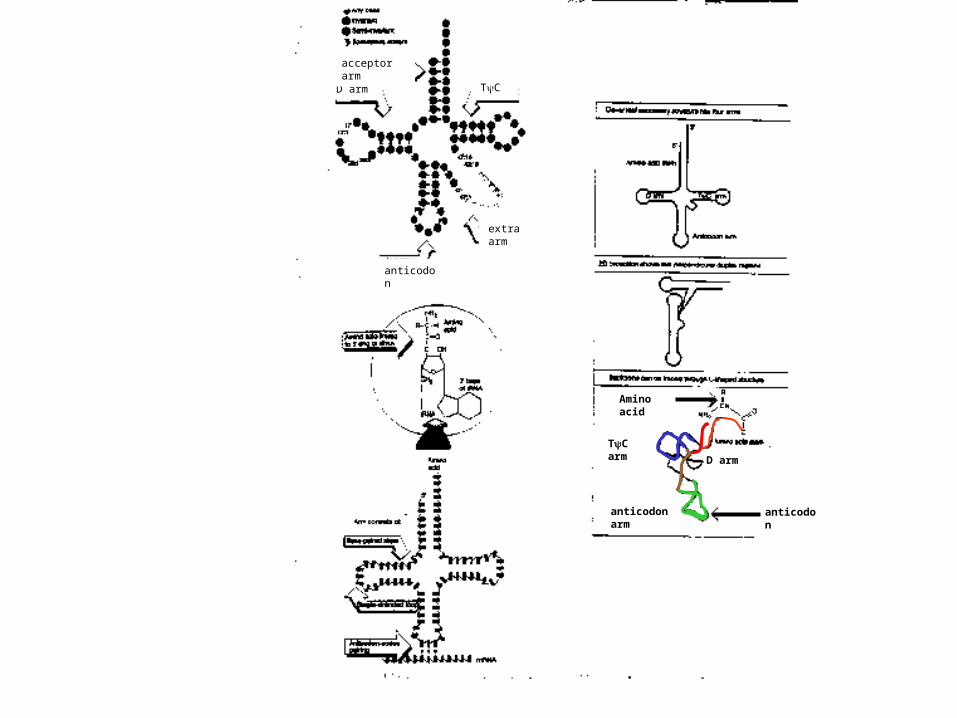
anticodon
D arm
anticodon arm
TC arm
Amino acid
TCD arm
anticodon
acceptor arm
extra arm
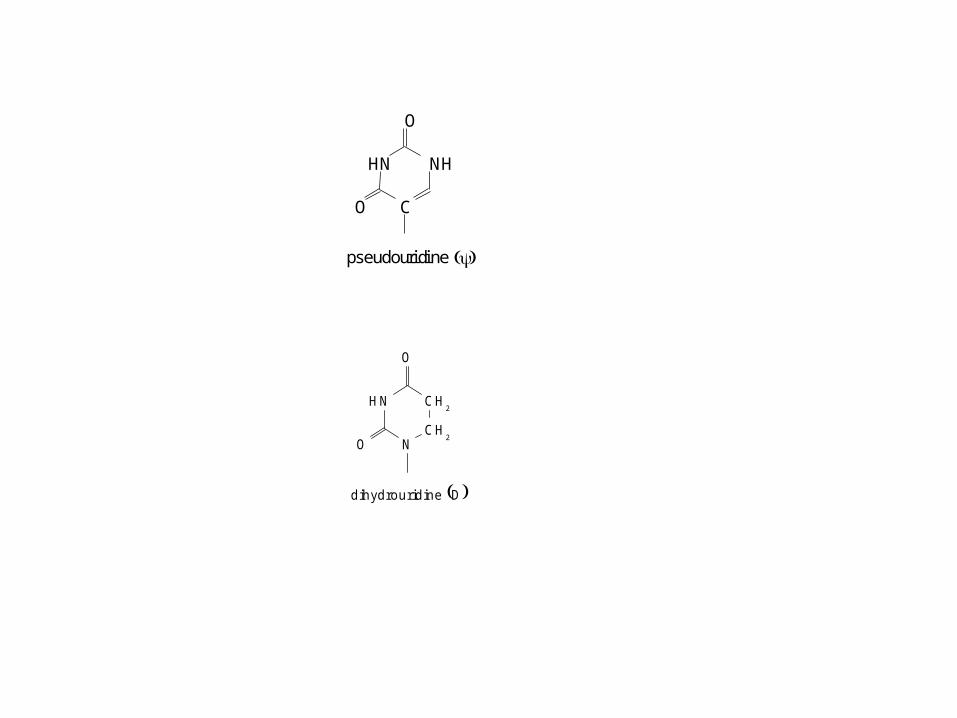
dihydrouridine D
HN CH2
CH2N
O
O
pseudouridine
HN NH
C
O
O

Yeast phe tRNA
(not charged with aa)
3-terminus
5-terminus
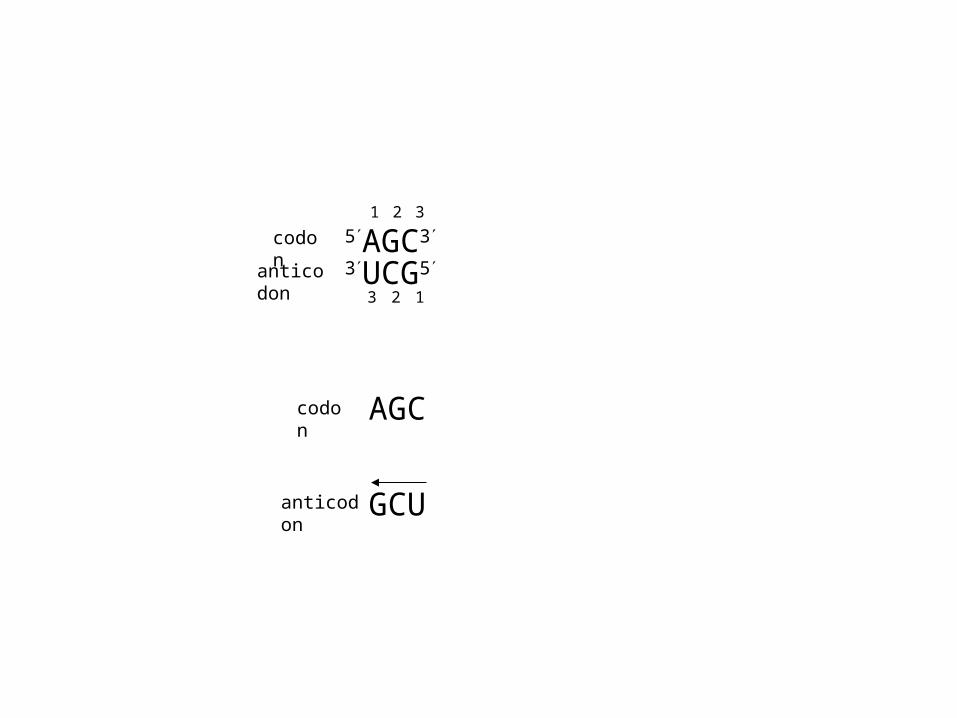
5AGC3
3UCG5
AGC
GCU
1 2 3
codon
anticodon
codon
anticodon
123

U in position 1 of the anticodon pairs with A or G in position 3 of codon
C G only
A U only
G C or U
Wobble hypothesis: rules for codon/anticodon pairing
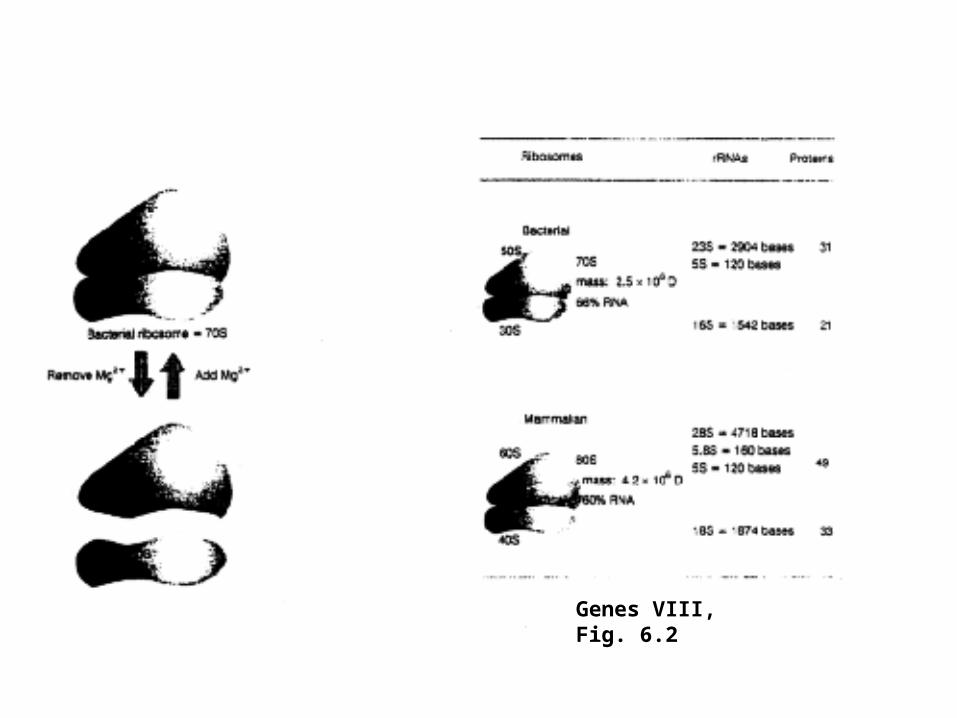
Genes VIII, Fig. 6.2

Genes VIII, Fig. 6.3
Genes VIII, Fig. 6.7
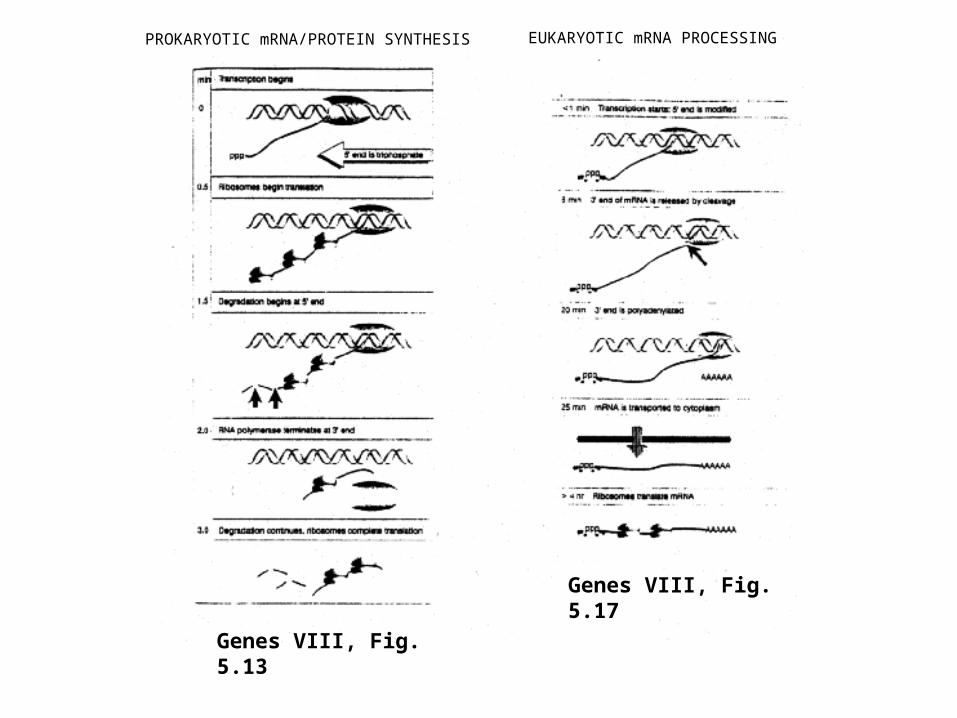
PROKARYOTIC mRNA/PROTEIN SYNTHESIS EUKARYOTIC mRNA PROCESSING
Genes VIII, Fig. 5.13
Genes VIII, Fig. 5.17
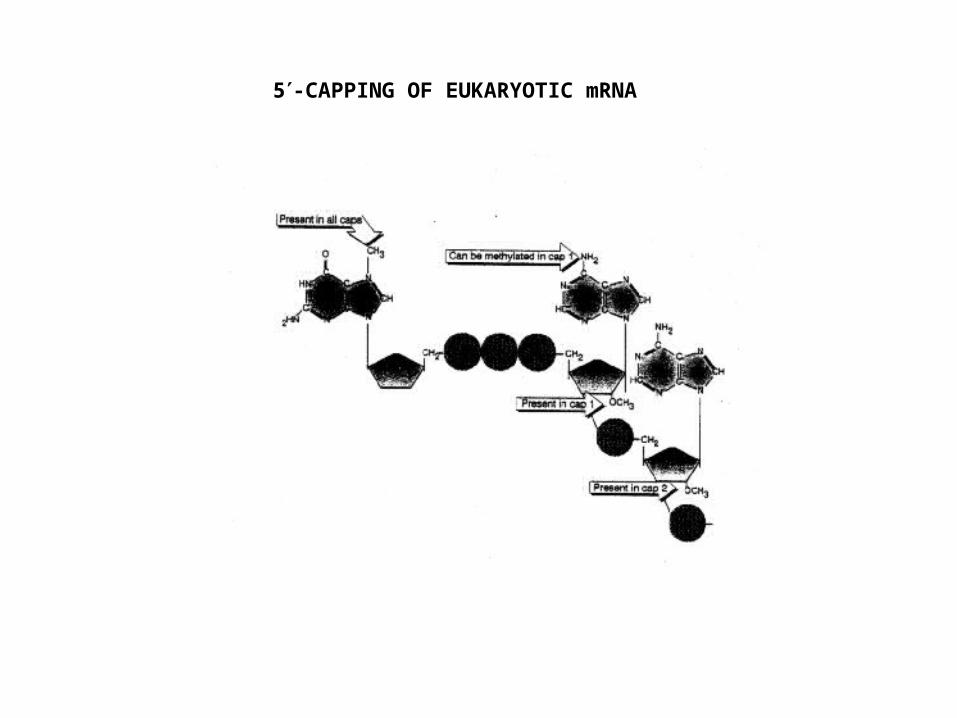
5-CAPPING OF EUKARYOTIC mRNA
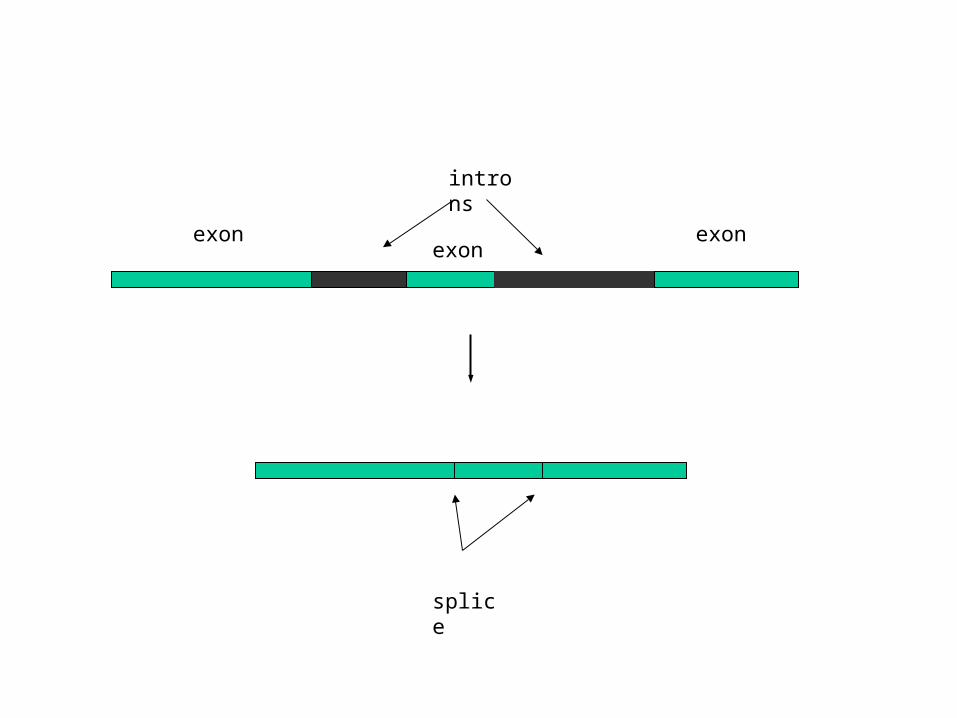
splice
exon
introns
exonexon
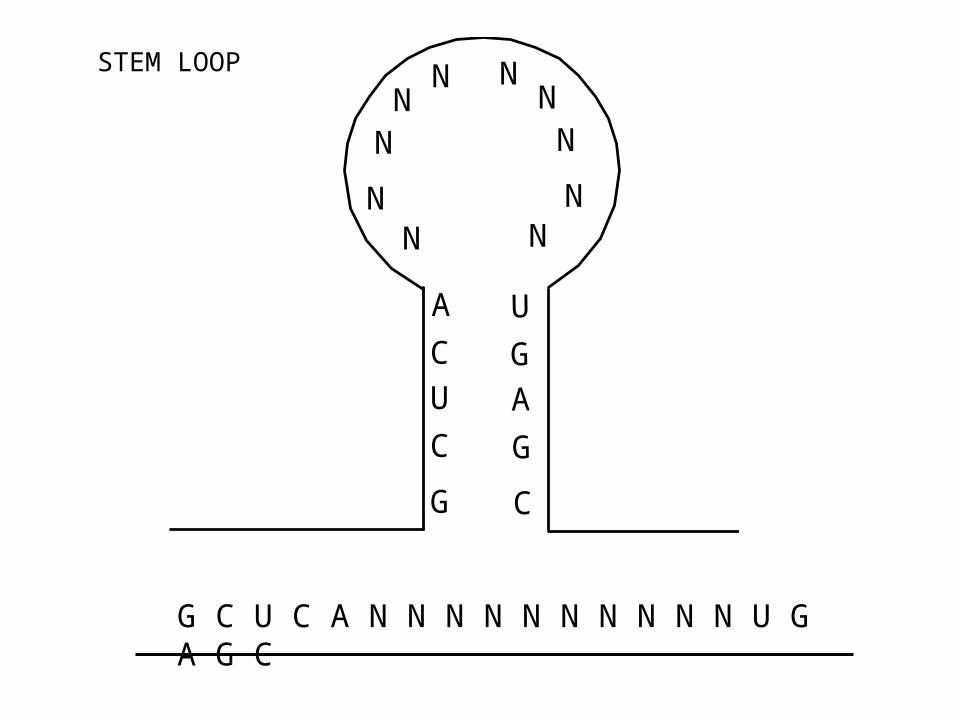
G
C
U
C
A
C
G
A
G
U
NN
NN
N
NN
NN
N
G C U C A N N N N N N N N N N U G A G C
STEM LOOP

Subunit (molecular weight) Function
2 x (40 kD) enzyme assembly, promoter recognition
(155 kD) catalytic center
(160 kD) catalytic center
(32-90 kD) promoter specificity
Subunits of prokaryotic RNA polymerase
Cat
alyt
ic c
ore
2′= holoenzyme
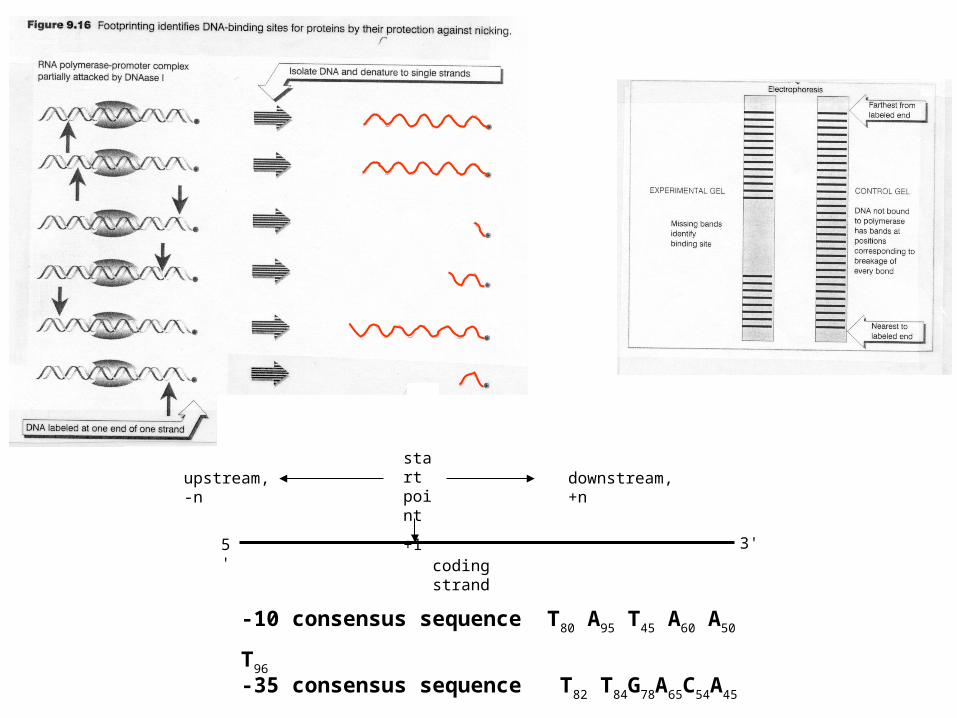
-10 consensus sequence T80 A95 T45 A60 A50 T96
-35 consensus sequence T82 T84G78A65C54A45
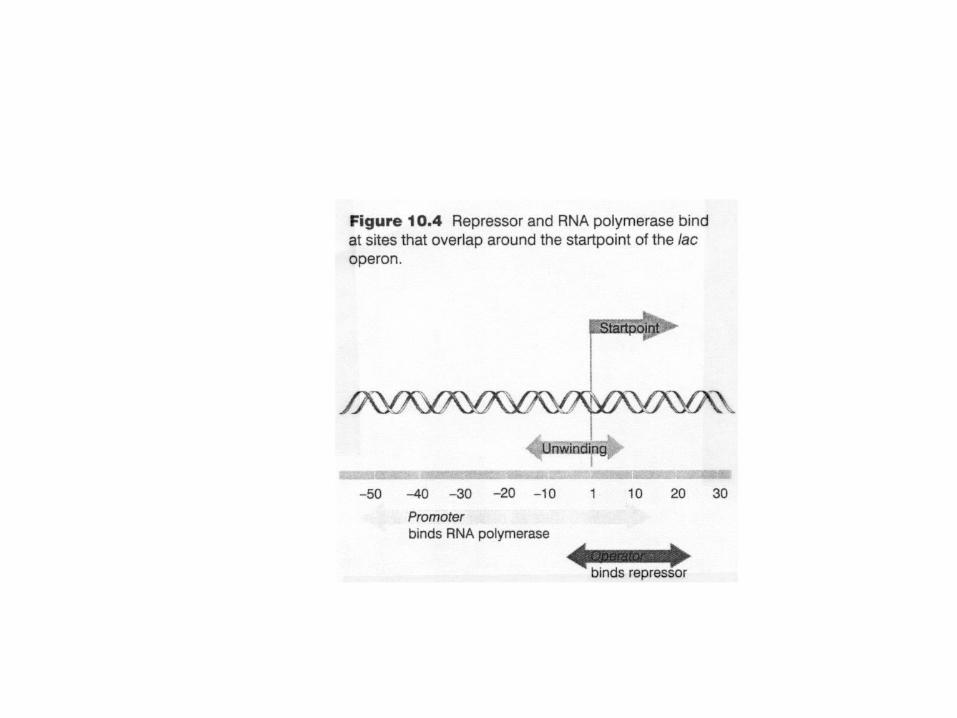
coding strand
start point
+1
5' 3'
upstream, -n downstream, +n


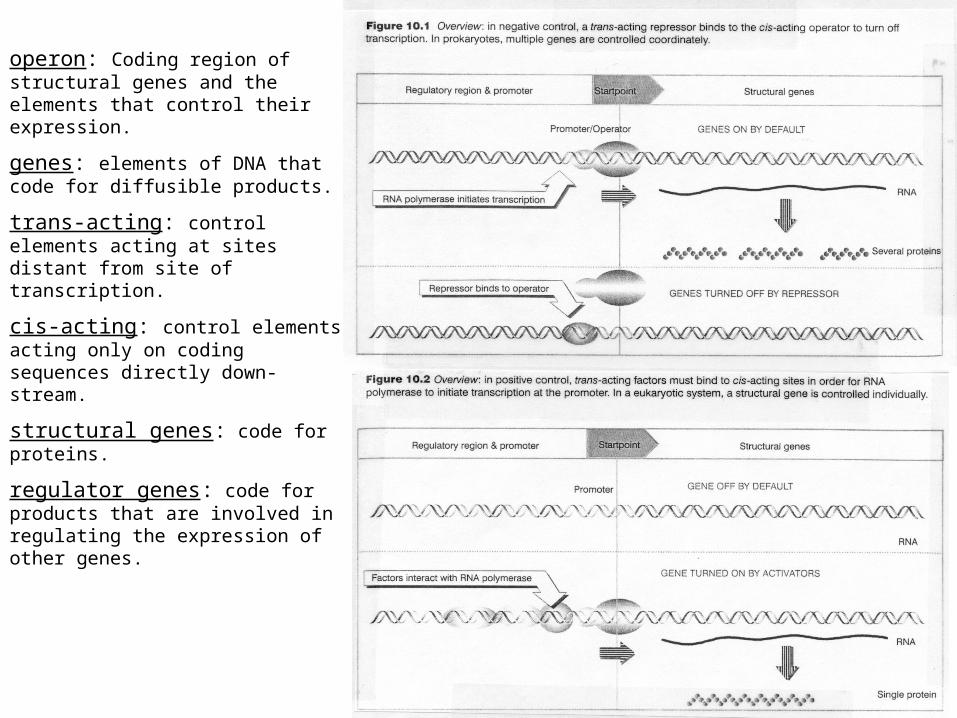
operon: Coding region of structural genes and the elements that control their expression.
genes: elements of DNA that code for diffusible products.
trans-acting: control elements acting at sites distant from site of transcription.
cis-acting: control elements acting only on coding sequences directly down-stream.
structural genes: code for proteins.
regulator genes: code for products that are involved in regulating the expression of other genes.

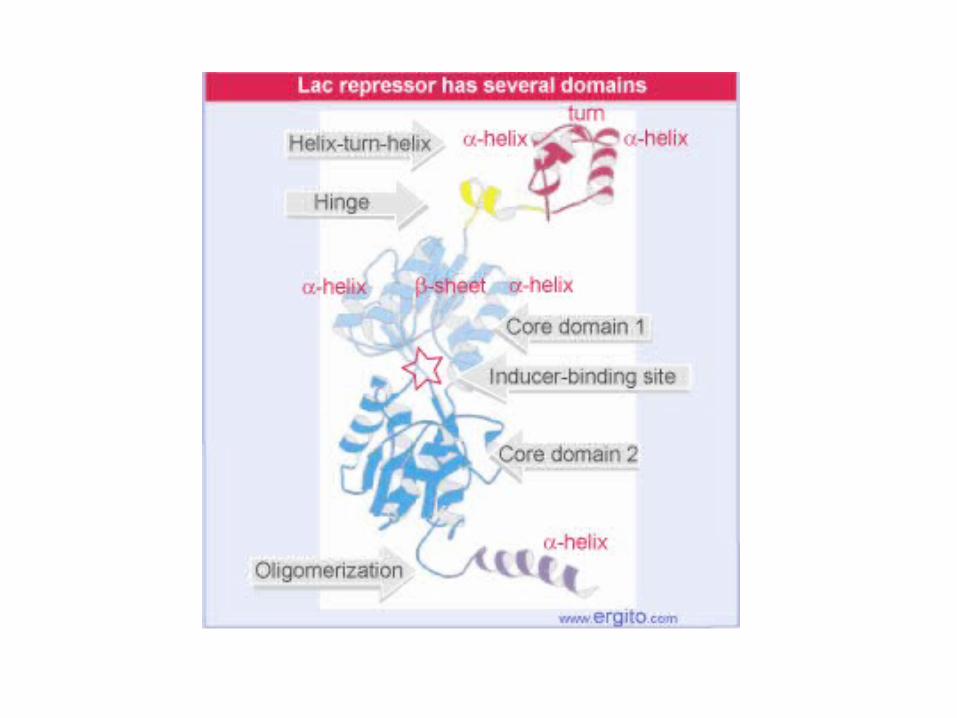
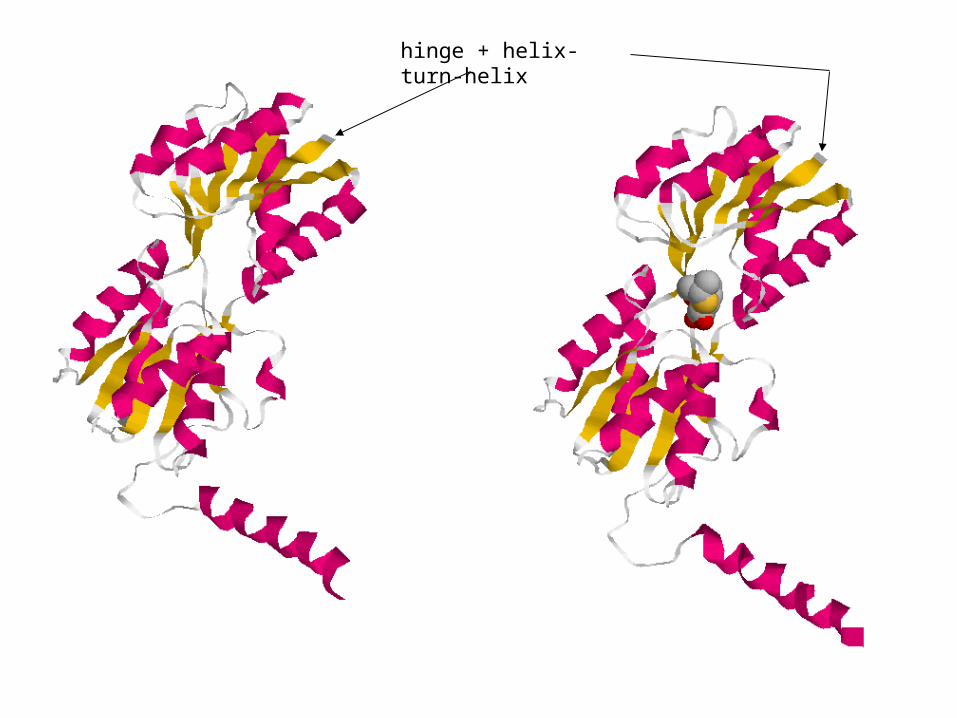
hinge + helix-turn-helix
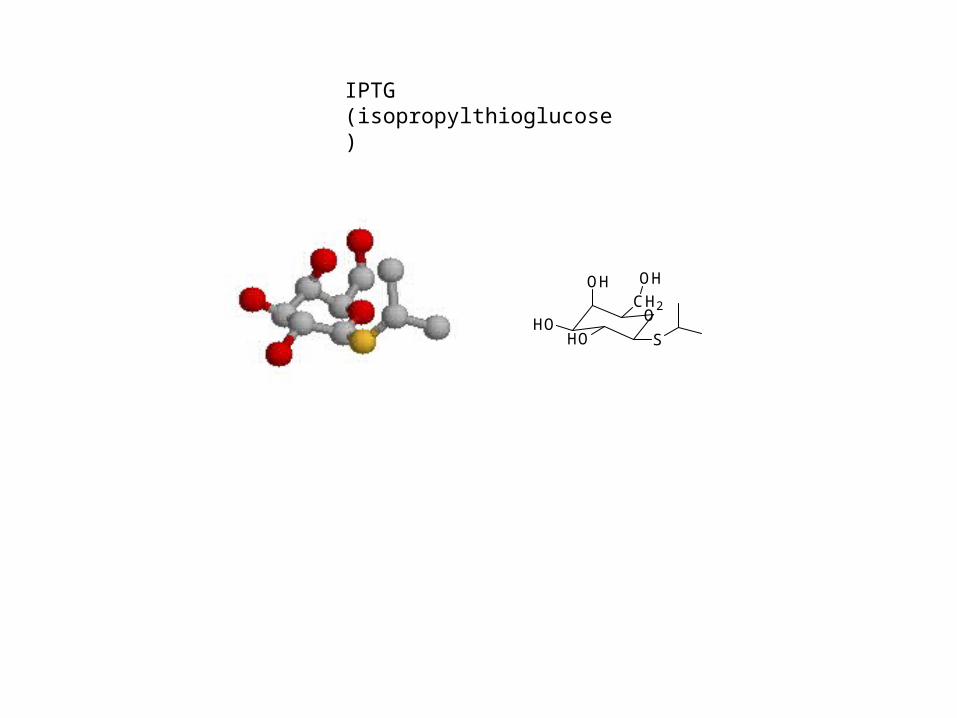
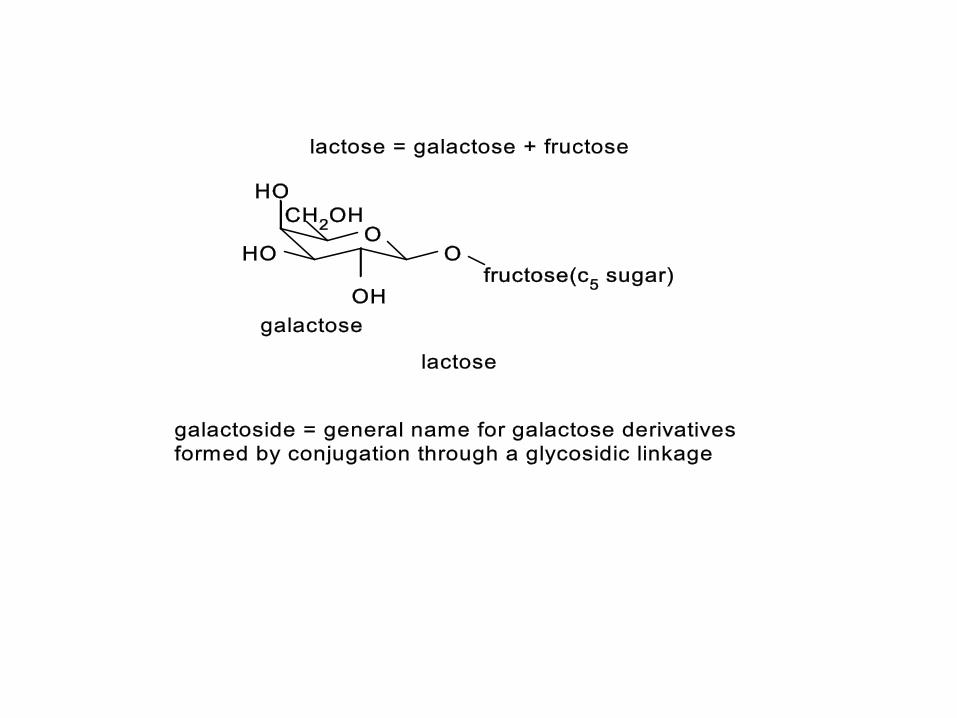
IPTG (isopropylthioglucose)
CH2
S
OH
HOHO
OH
O
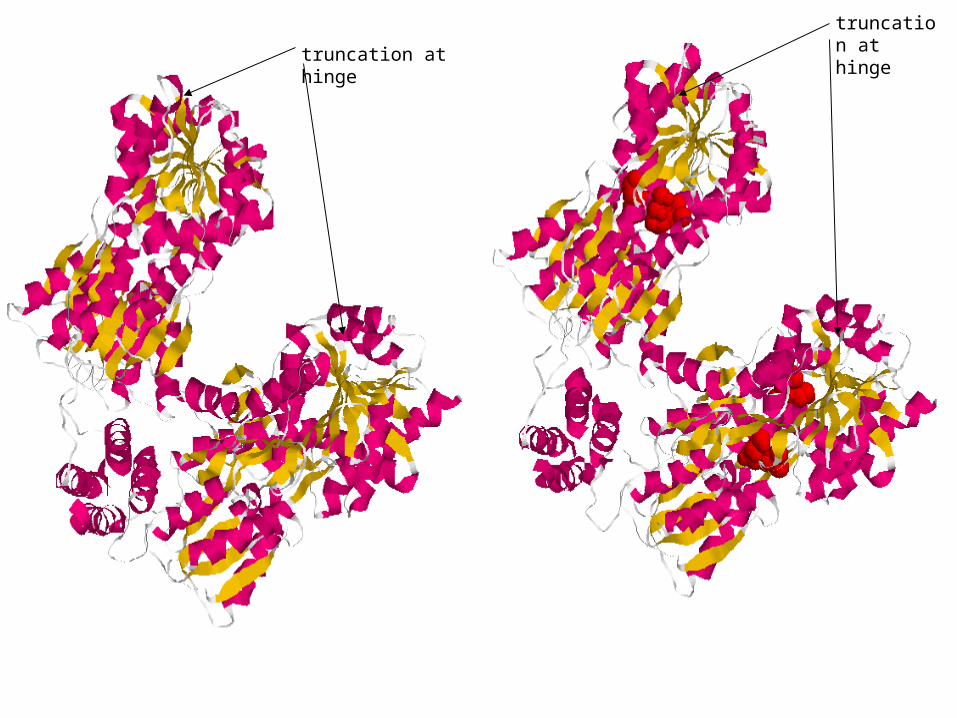
truncation at hinge
truncation at hinge
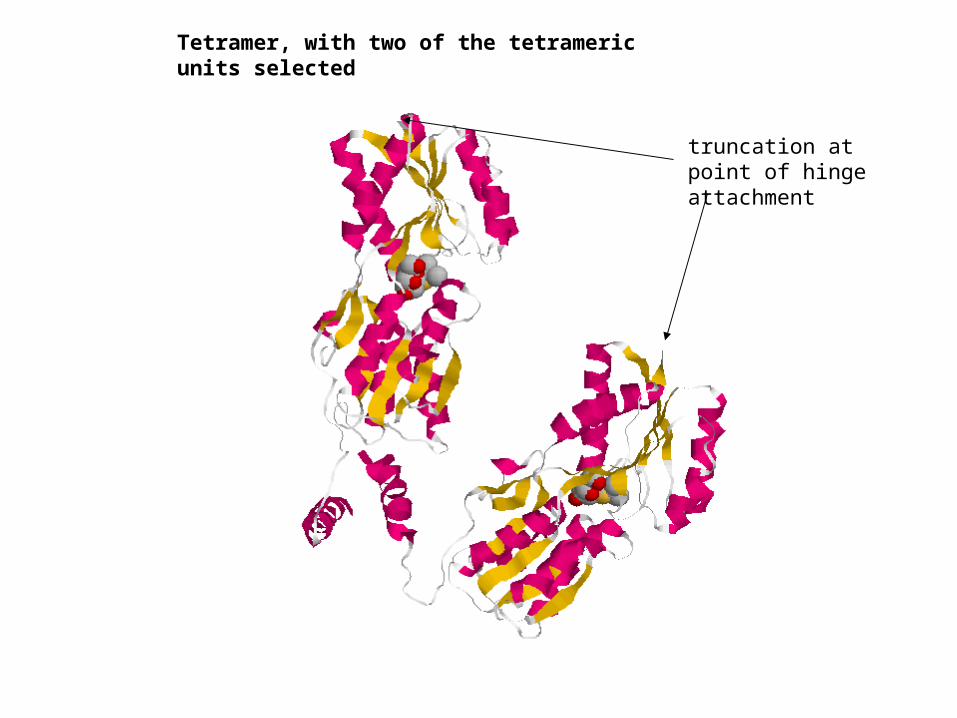
truncation at point of hinge attachment
Tetramer, with two of the tetrameric units selected
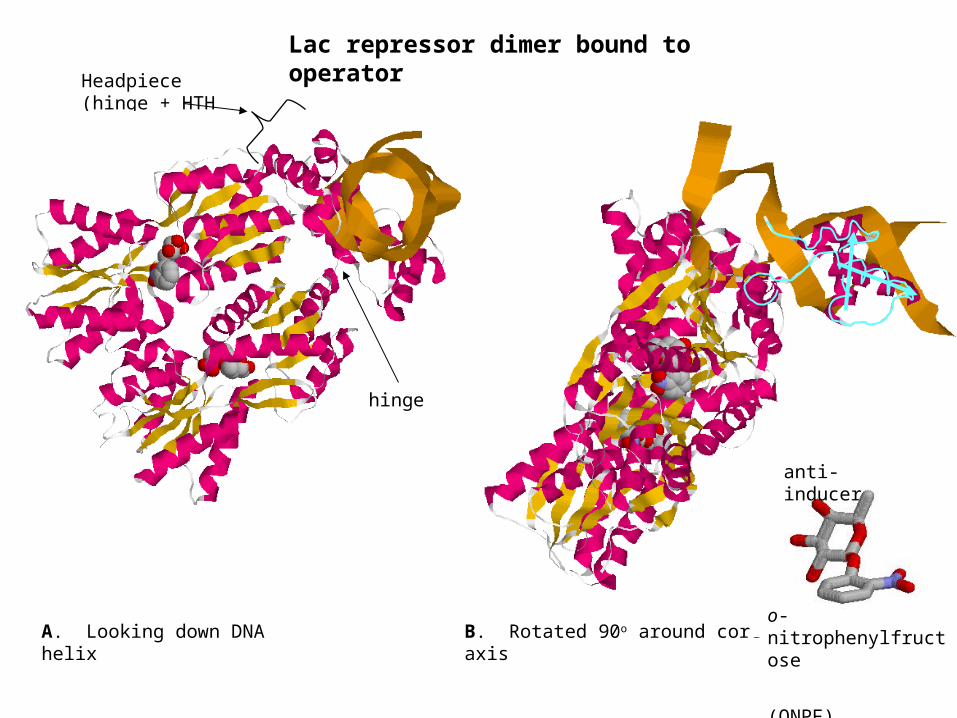
B. Rotated 90o around core axis
Headpiece (hinge + HTH motif)
A. Looking down DNA helix
Lac repressor dimer bound to operator
anti-inducer
o-nitrophenylfructose
(ONPF)
hinge
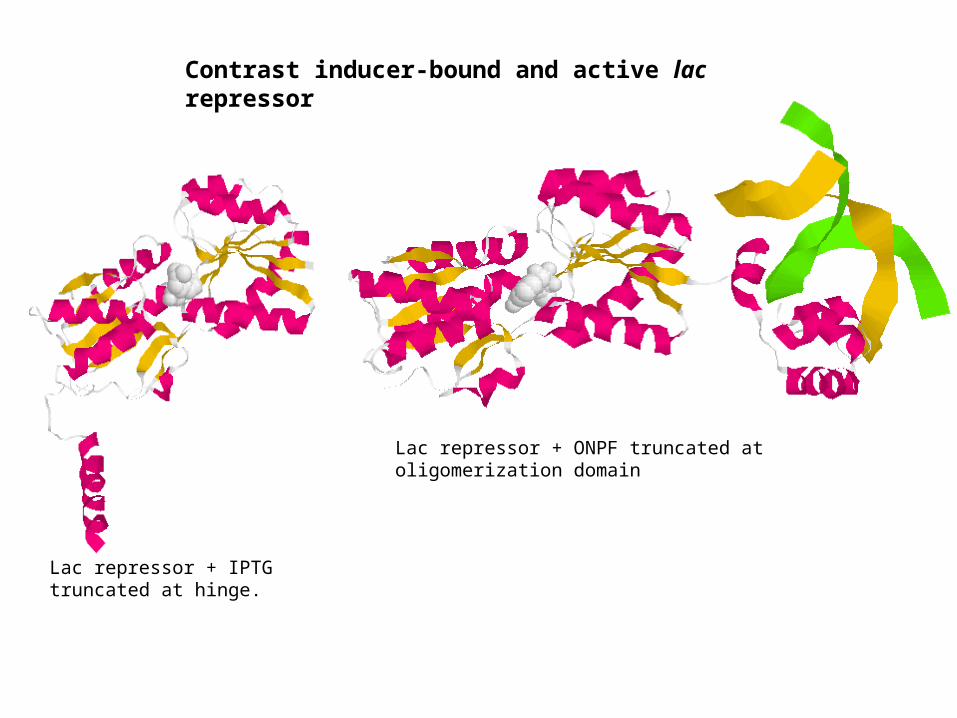
Lac repressor + IPTG truncated at hinge.
Lac repressor + ONPF truncated at oligomerization domain
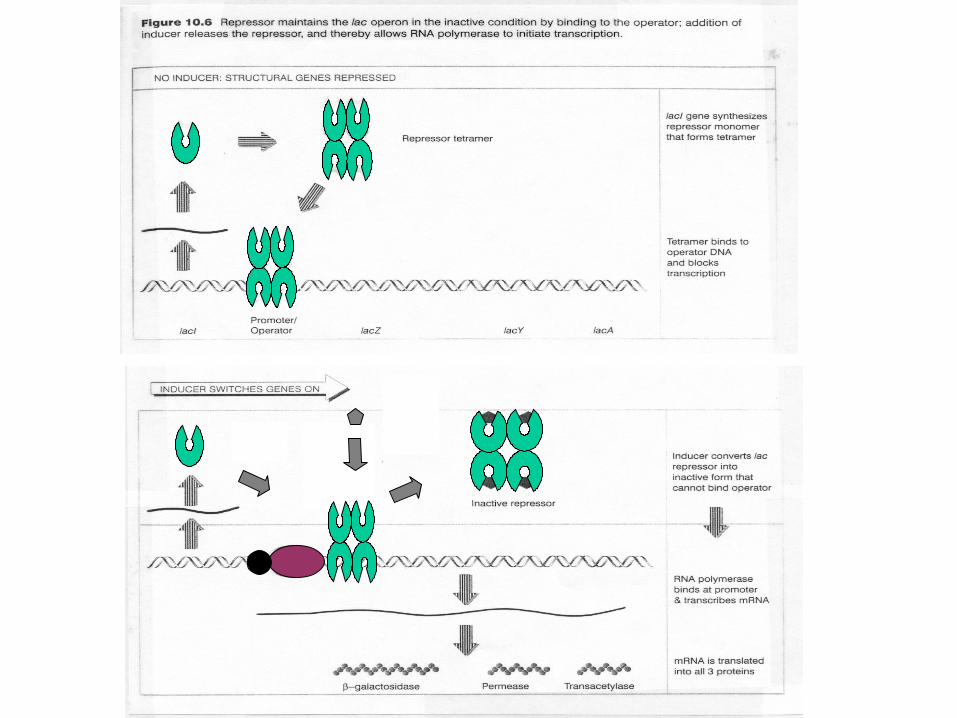
Contrast inducer-bound and active lac repressor
OO
OOH
P
O
N
N
NH2
N
N
O
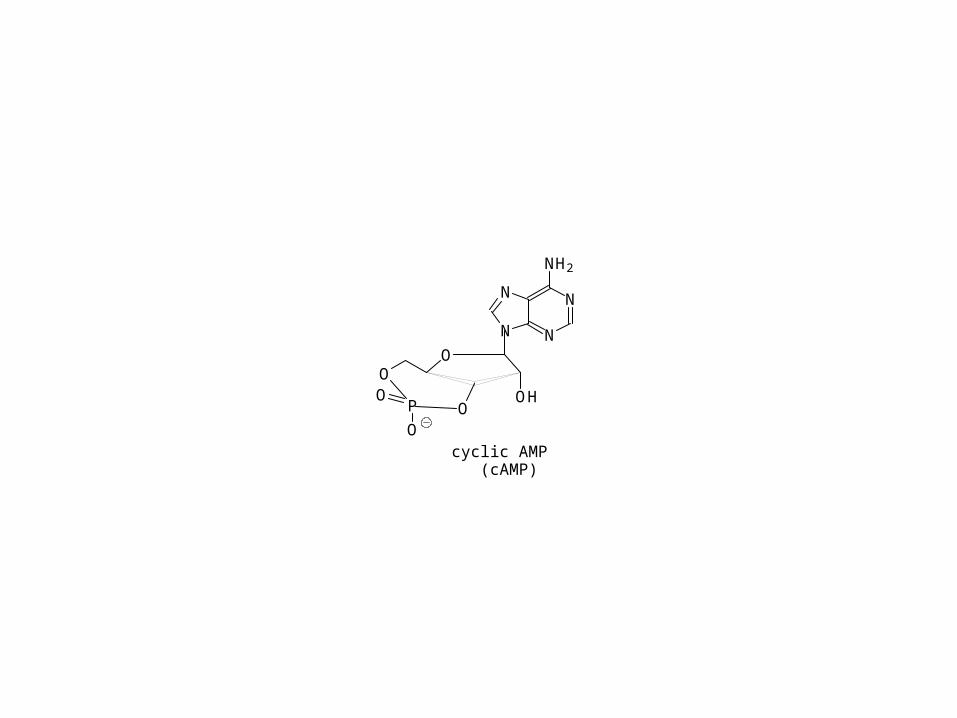
cyclic AMP (cAMP)





























