Embed Size (px)
Citation preview

Enrichment and characterization of three ammonia-oxidizing Archaea from freshwater 1
environments. 2
3
Elizabeth French 1, Jessica A. Kozlowski 1,3, Maitreyee Mukherjee 2, George Bullerjahn 2, and 4
Annette Bollmann 1* 5
6
1 Miami University 7
Department of Microbiology 8
32 Pearson Hall, 700 East High Street 9
Oxford, OH 45056 10
USA 11
12
2 Bowling Green State University 13
Department of Biological Sciences 14
516 Life Sciences 15
Bowling Green, OH 43403 16
USA 17
18
3 present address: 19
University of Alberta 20
Department of Biological Sciences 21
CW405, Biological Sciences Building 22
Edmonton, Alberta T6G 2E9 23
Canada 24
25
Copyright © 2012, American Society for Microbiology. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.00432-12 AEM Accepts, published online ahead of print on 8 June 2012
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

2
* Corresponding author: 26
email: [email protected] 27
phone: +1 513 529 0426 28
fax: +1 513 529 2431 29
30
31
Short title: Characterization of three AOA enrichments 32
33
Section: Microbial Ecology 34
35
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

3
Abstract 36
Aerobic biological ammonia oxidation is carried out by two groups of microorganisms, 37
ammonia-oxidizing bacteria (AOB), and the recently discovered ammonia-oxidizing archaea 38
(AOA). Here we present a study using cultivation-based methods to investigate the differences in 39
growth of three AOA cultures and one AOB culture enriched from freshwater environments. The 40
enriched AOA belong to the Thaumarchaeal group I.1a, with one enrichment culture having the 41
highest identity with Candidatus Nitrosoarchaeum koreensis and the other two representing a 42
new genus of AOA. The AOB enrichment culture was also obtained from freshwater and had the 43
highest identity to AOB from the Nitrosomonas oligotropha group (Nitrosomonas cluster 6a). 44
We investigated the influence of ammonium, oxygen, pH, and light on the growth of AOA and 45
AOB. The growth rates of the AOB increased with increasing ammonium concentrations while 46
the growth rates of the AOA decreased slightly. Increasing oxygen concentrations led to a 47
increase in the growth rate of the AOB, while the growth rates of AOA were almost oxygen 48
insensitive. Light exposure (white and blue wavelengths) inhibited the growth of AOA 49
completely, and the AOA did not recover when transferred to the dark. AOB were also inhibited 50
by blue light, however, growth recovered immediately after transfer to the dark. Our results show 51
that the tested AOB has an competitive advantage over the tested AOA under most investigated 52
conditions. Further experiments will elucidate the niches of AOA and AOB in more detail. 53
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

4
Introduction 54
Nitrification, the microbial oxidation of NH3 (ammonia) to NO3- (nitrate), is one of the key 55
processes of the global nitrogen cycle. The first and rate-limiting step of nitrification is the 56
oxidation of NH3 to NO2- (nitrite). Until recently, aerobic ammonia oxidation was attributed only 57
to a small subset of the Proteobacteria; most freshwater and terrestrial ammonia-oxidizing 58
bacteria (AOB) belong to a distinct group in the Betaproteobacteria, while a few marine AOB 59
species belong to the Gammaproteobacteria (30, 33, 34). The AOB have a 60
chemolithoautotrophic metabolism, oxidizing NH3 to NO2- via the intermediate NH2OH 61
(hydroxylamine), and fixing carbon from CO2 (carbon dioxide) via the Calvin cycle (1). 62
Recently, genes encoding ammonia monooxygenase (amoA), the first enzyme in the process of 63
ammonia oxidation, were discovered together with archaeal 16S rRNA genes in a metagenomic 64
study (62) and a soil fosmid library (59). At the same time, Nitrosopumilus maritimus, the first 65
archaeal ammonia oxidizer, was isolated into pure culture from a saltwater aquarium (31). 66
Ammonia-oxidizing archaea (AOA) in pure and enrichment cultures have essentially the same 67
metabolism as AOB; they oxidize NH3 stoichiometrically to NO2- and fix carbon from 68
bicarbonate (HCO3-) (15, 21, 31, 38, 45, 58). However, the genomes of N. maritimus and 69
Candidatus Nitrosoarchaeum limnia revealed differences between AOA and AOB, such as the 70
use of the 3-hydroxypropionate/4-hydroxybutyrate pathway for HCO3- fixation, the absence of 71
hydroxylamine oxidoreductase, and the presence of many copper-containing enzymes (5, 65). 72
AOA and AOB often co-occur in the same environment, but the contributions of AOA and AOB 73
to the total ammonia oxidation still need to be elucidated. Many previous studies focused on the 74
influence of environmental factors on niche differentiation between AOA and AOB using 75
cultivation-independent molecular methods. From those studies it can be concluded that AOA 76
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

5
are frequently found in environments with lower substrate (NH4+ and O2) availability, and AOB 77
in environments with higher substrate availability (4, 13, 17, 25, 44, 57 among others). However, 78
most of these studies were conducted using methods that target the abundance and/or expression 79
of the archaeal and bacterial amoA genes. Unfortunately, it is not possible to draw direct 80
conclusions about the activity of the AOA and AOB based on abundance and expression of the 81
amoA gene, because amoA mRNA has been detected in AOB for weeks and 16S rRNA 82
(ribosomes) for up to a year after the onset of starvation (8, 26, 27). The response of AOA 83
towards starvation and resuscitation has not yet been investigated. In addition it has been shown 84
that not all amoA-encoding Thaumarchaeota are autotrophic ammonia oxidizers (41, 66). While 85
studies focusing on the analysis of abundance and activity of microbes using molecular methods 86
give very valuable insights, it is also necessary to investigate the response of microbes to 87
environmental factors using cultivation based approaches, because these experiments will 88
demonstrate changes in physiological activity more conclusively. 89
Here we present a study that used a cultivation-dependent approach to investigate the responses 90
of AOA and AOB to environmental factors. Three phylogenetically distinct AOA cultures were 91
enriched from freshwater sediments in Ohio, USA and their growth was characterized under 92
different conditions and compared with that of a freshwater AOB enrichment culture. Factors of 93
interest include the NH4+ concentration, pH, O2 concentration, and light wavelength and 94
intensity. These factors have strong effects on the physiology and niche differentiation of AOB 95
(7, 24, 46, 56) and are, therefore, also very likely to influence the physiology and niche 96
differentiation between AOA and AOB. 97
98
99
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

6
Material and Methods 100
Sampling: Near shore sediment samples were taken from Lakes Acton (AC) (39°57’N, 101
84°74’W) and Delaware (DW) (40°39’N, 83°05’W) in Fall 2008. Additional sediment core 102
samples were collected from Lake Acton in Summer 2009. 103
Medium: Mineral salts medium (MS medium) used to enrich and cultivate AOA and AOB 104
contained 10 mM NaCl, 1 mM KCl, 1 mM CaCl22H2O, 0.2 mM MgSO47H2O, and 1ml l-1 105
trace elements solution (9, 63). HEPES buffer was added in a four-fold molar ratio to the NH4+ 106
concentration, and the pH was adjusted to 7.5 before autoclaving. After autoclaving, sterile 107
KH2PO4 solution was added to obtain a final concentration of 0.4 mM (9, 63). 108
Enrichment of the AOA (AOA-AC2, AOA-AC5 and AOA-DW): Sediment samples (1 g) 109
were inoculated into 50 ml MS medium with 0.25 mM NH4+ immediately upon arrival in the 110
laboratory. The enrichments were incubated at 27°C in the dark. NH4+ levels were monitored 111
weekly using a colorimetric assay (9, 29). When the cultures reached late logarithmic growth 112
phase (depletion of around 80% of the initial NH4+ concentration) they were transferred to fresh 113
medium using 10% v/v inoculum. The cultures were passed through 0.45 µm filters for the first 114
five to six transfers to exclude AOB (Annika Mosier, personal communication; 9). In addition to 115
filtration, the enrichment cultures from DW were also treated with 100 µg ml-1 streptomycin to 116
eliminate AOB. After several transfers, when the cultures depleted NH4+ in regular intervals, 20 117
ml was collected on 0.1 µm nitrocellulose filters for molecular characterization. The filters were 118
stored at -20°C. 119
AOB culture: We used the previously described AOB freshwater enrichment culture G5-7 120
(AOB-G5-7) to compare the growth of AOA to AOB (6, 7). The culture belongs to the 121
Nitrosomonas oligotropha cluster and is adapted to low NH4+ concentrations (6, 7). Members of 122
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

7
this AOB cluster have been found in many freshwater environments around the world (11, 12, 123
14, 18, 23, 55). 124
Growth experiments: All growth experiments were conducted in MS medium with 0.5 mM 125
NH4+ at pH 7.5 in 125 ml Erlenmeyer flasks with cotton stoppers unless otherwise noted. We 126
tested the influence of different factors (NH4+ concentration, O2 concentration, pH, and light) on 127
the rate of NO2-/NO3
- production of the three AOA enrichment cultures (AOA-AC2, AOA-AC5 128
and AOA-DW) and the AOB enrichment culture (AOB-G5-7). All cultures were inoculated with 129
10% v/v of a conditioned late log phase culture and incubated in the dark at 27°C. Samples (1 130
ml) were taken at regular intervals and centrifuged at 16,000 rpm for 20 min. The supernatant 131
was stored at -20°C for further chemical analysis. To investigate the influence of different NH4+ 132
concentrations, media with 15 µM - 5 mM NH4+ were prepared with the corresponding HEPES 133
concentrations. The influence of pH was investigated by adjusting the initial pH in the medium 134
to values between 6 and 9. The influence of O2 concentration was investigated by equilibrating 135
the medium in serum bottles under anaerobic conditions overnight. After equilibration the bottles 136
were sealed with rubber stoppers. Different calculated O2 concentrations in the headspace were 137
achieved by exchanging the corresponding volume of the headspace with sterile filtered air. The 138
influence of light was investigated by incubating the cultures 18 cm above LED panels emitting 139
30 µmol photons m-2 s-1 at the wavelength 5000 - 7000 K (white light), 623 ± 3 nm (red light), 140
and 470 ± 5 nm (blue light); and 3 µmol photons m-2 s-1 at the wavelength 470 ± 5 nm (blue 141
light). The light intensity inside the glass bottles was 25 µmol photons m-2 s-1 (high light) and 2.5 142
µmol photons m-2 s-1 (low light conditions) as measured with a LI-250A light meter (LI-COR 143
Biosciences, Lincoln, NE) indicating that the glass filtered approximately 15% of the light. To 144
investigate the influence of light-to-dark and dark-to-light transitions on the growth of AOA and 145
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

8
AOB, cultures were incubated in the dark until 50% of the NH4+ was consumed and then 146
transferred to the light. At the same time, cultures that were incubated in the light were 147
transferred from the light to the dark. Controls were incubated for the complete cycle in the dark. 148
Evaluation of growth experiments: NO2- and NO3
- concentrations were determined in the 149
supernatants using colorimetric assays (9, 51). NO2-/NO3
- concentrations were log transformed 150
and plotted against time (Fig. S1). Growth rates were calculated from the linear increase (slope) 151
of the log-transformed NO2-/NO3
- concentrations over time, assuming that NO2-/NO3
- production 152
in the cultures is correlated with the growth of AOA and AOB (3, 9, 31). The increase in NO2-153
/NO3- production was linear for several days to one week and the correlation coefficients were 154
always ≥ 0.97 but in most cases even ≥ 0.99. 155
156
Molecular analysis 157
DNA isolation from the AOA enrichment cultures: DNA was isolated from the nitrocellulose 158
filters using the Qiagen DNeasy Blood and Tissue Kit (Valencia, CA) with the following 159
modifications. Acid-washed zirconium beads (1g) and 500 µl high salt buffer (1 M NaCl, 5 mM 160
MgCl22H2O, 10 mM Tris, pH 8) (2) were added to the nitrocellulose filters. The filters were 161
homogenized using a bead beater (Biospec Products, Bartlesville, OK) at 4800 rpm for 30 s. This 162
was repeated three times and the samples were stored in between cycles on ice for 10 min. After 163
bead beating, 500 µl Qiagen Buffer AL and 50 µl proteinase K were added and the mixture was 164
incubated at 56°C for 30 min. The reaction mixture was spun down at 8000 rpm for 1 min and 165
transferred to spin columns supplied by the manufacturer. The spin columns were treated 166
according to the manufacturer’s recommendations and the DNA was eluted with 100 µl elution 167
buffer AE. 168
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

9
PCR: GoTaq Green Master Mix (Promega, Madison WI) was used for all standard PCR 169
according to the manufacturer’s recommendations using the primers and protocols summarized 170
in Table S1. 171
Cloning and sequencing: PCR products were cleaned using the Wizard SV Gel and PCR 172
product Clean up system (Promega, Madison, WI) and cloned into the pGEM-T easy vector 173
system (Promega, Madison, WI). Transformants were screened for inserts using PCR with M13 174
primers and the PCR products were cleaned up and sequenced using the BigDye Terminator 175
V3.1 cycle sequencing kit (Life Technology Corporation, Carlsbad, CA) on an Applied 176
Biosystems 3730x1 DNA analyzer (Life Technology Corporation). 177
DNA sequence analysis: All sequences were edited with 4Peaks (A. Griekspoor and T. 178
Groothuis, The Netherlands Cancer Institute). The sequences were aligned using ARB (37). 179
Phylogenetic trees were constructed using the neighbor-joining algorithm in ARB, and 180
parsimony and maximum likelihood methods using PHYLIP (16). Trees constructed with all 181
three methods showed the same overall grouping, therefore only the tree constructed with 182
neighbor-joining method has been presented. All sequences were deposited in Genbank under 183
the numbers: JQ669389-JQ669394. 184
Fluorescence in-situ Hybridization (FISH): The CARD-FISH protocol (47, 50) was used with 185
the following modifications: the hybridization temperature was 46°C, the first wash was 186
performed at 48°C, followed by an amplification step at 46°C. All probes (Table S2) were 187
labeled at their 5’ end with horseradish peroxidase and used at a final concentration of 50 ng µl-1. 188
All filters were counterstained with DAPI for total cell counts. Direct microscopic counts by 189
fluorescence microscopy (Zeiss Axiophot HB0100, Carl Zeiss Inc, North America) were 190
performed at 1000X magnification. 191
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

10
Results 192
Enrichment of AOA: AOA were enriched from the sediment of Lakes Acton (AOA-AC2 and 193
AOA-AC5) and Delaware (AOA-DW) under autotrophic conditions with NH4+ as the sole 194
electron donor in the medium. Based on the AOA amoA sequences, all enrichment cultures 195
belong to the water column/sediment group I.1a of the Thaumarchaeota (Fig. 1). AOA-AC2 was 196
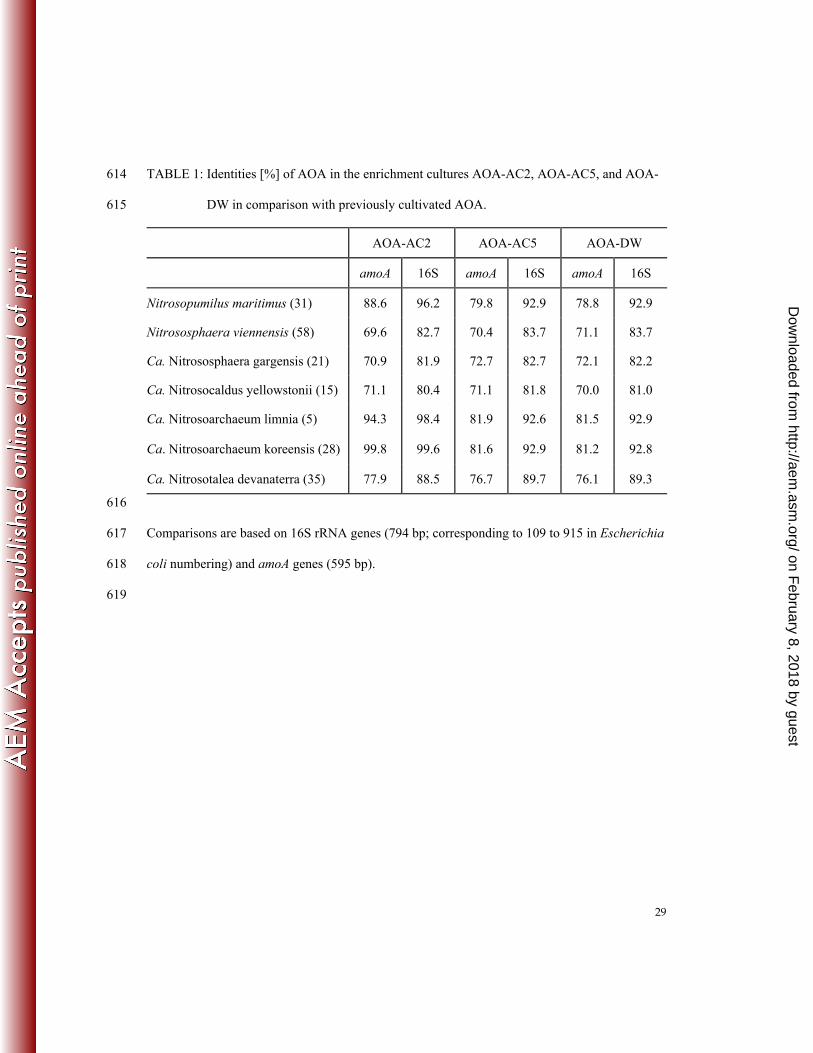
81-81.7% (amoA) and 92.8-93.1% (16S rDNA) identical to the other two enrichment cultures, 197
while AOA-AC5 and AOA-DW were 87.1% (amoA) and 97.9% (16S rDNA) identical to each 198
other. The amoA sequences of AOA-DW were 98.2-98.5% identical to clones from the sediment 199
of Lakes Acton, Delaware and Pleasant Hill (36), 98.5% to clones from the freshwater sediment 200
in the San Francisco Bay (40), and 98.1% to clones from a drinking water distribution system in 201
the Netherlands (61). The amoA sequences of AOA-AC5 were 99% identical to a clone from a 202
paddy soil in Japan (19). The third enrichment culture AOA-AC2 is closely related to Ca. 203
Nitrosoarchaeum koreensis (99.8% identity on amoA basis and 99.6% identity on 16S rDNA 204
basis) and Ca. Nitrosoarchaeum limnia (94.3% identity on amoA basis and 98.5% identity on 205
16S rDNA basis) (5, 28). In contrast to AOA-AC2, AOA-AC5 and AOA-DW were not closely 206
related to described AOA isolates or enrichment cultures such as N. maritimus and 207
Nitrososphaera viennensis among others (70-82% identity on amoA basis and 81-93% identity 208
on 16S rDNA basis) (Table 1). 209
Fluorescent in situ hybridization (CARD-FISH) was used to determine the proportion of AOA in 210
the enrichment cultures at the end of the logarithmic growth phase. AOA-DW contained 85% 211
AOA, AOA-AC2 91% and AOA-AC5 81% (Table 2). AOB and NOB (nitrite-oxidizing bacteria) 212
were not detected as tested by PCR amplification with AOB-specific 16S rRNA and amoA 213
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

11
primers (Table S1) (results not shown) and FISH using AOB- and NOB-specific 16S rRNA 214
probes (Table 2 and Table S2). 215
Influence of NH4+ concentration on the growth rates of AOA and AOB. During stratification 216
in the summer the NH4+ concentration in Lake Acton increases up to 400 µM (43), which falls 217
within the tested range of NH4+ concentrations of 15 µM and 2 mM NH4
+. Increasing NH4+ 218
concentrations up to 1 mM NH4+ doubled the growth rate of AOB-G5-7, while the growth rates 219
of the AOA enrichment cultures decreased or remained constant (Fig. 2). The growth rate of 220
AOA-DW at the lowest NH4+ concentration (15 µM) was significantly higher than the growth 221
rate at higher NH4+ concentrations (Table S3). The same tendency was observed for the other 222
two cultures, although the statistical support was less strong (Fig. 2; Table S3). The AOA 223
enrichment cultures exhibited different tolerances to high NH4+ concentrations; AOA-DW grew 224
at NH4+ concentrations up to 1 mM, AOA-AC2 up to 2 mM and AOA-AC5 up to 5 mM (Fig. 2). 225
The lag phase of AOA and AOB differed; AOB-G5-7 became active 1-3 days after inoculation at 226
all tested NH4+ concentrations, whereas the lag phase of the AOA cultures increased with 227
increasing initial NH4+ concentrations up to more than two weeks before logarithmic growth 228
could be detected at NH4+ concentrations between 1 mM and 5 mM NH4
+ (Fig. S2; Table S4). 229
Influence of O2 concentration on the growth of AOA and AOB: Lake Acton, stratifies during 230
the summer and has an anaerobic zone as well as a zone with low oxygen availability (1 mg l-1 231
O2) (43). We therefore investigated the response of our enrichment cultures to 0.5-2% O2 232
(calculated) in the headspace, which corresponded to 0.2-0.8 mg l-1 O2 in the medium. The 233
growth rate of AOB-G5-7 decreased with decreasing O2 concentration and the growth rates at all 234
different O2 concentrations were significantly different from each other (Fig. 3; Table S5). The 235
AOA enrichment cultures grew at all O2 concentrations in the headspace with the exception of 236
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

12
AOA-AC2 at 0.5% O2. The decrease of the growth rates with decreasing O2 concentration in the 237
AOA cultures was less steep than the decrease of the growth rates in the AOB enrichment 238
culture. However, the growth rates at low O2 concentrations in the AOA-AC2 and AOA-AC5 239
were significantly lower than the growth rates at 21% O2 (Fig. 3; Table S5). 240
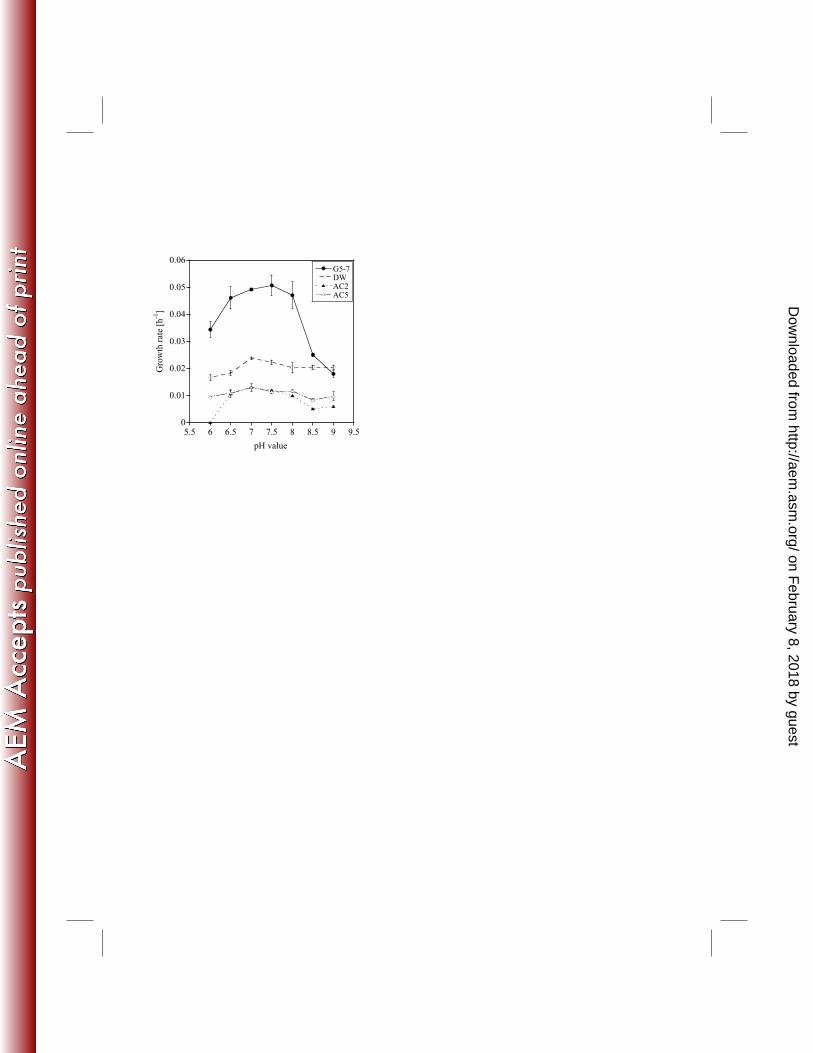
Influence of pH on the growth of AOA and AOB: We investigated the growth of all cultures 241
at pH 6-9, the range at which non-acidophilic ammonia oxidizers grow (32, 33, 34). The growth 242
rates of all cultures showed bell-shaped curves in relation to the pH with maximum growth rates 243
at pH 7-7.5 (Fig. 4). AOA-AC2 did not grow at pH 6, while the other AOA and AOB cultures 244
did. The growth rates of AOA-AC5 and AOA-DW at pH 9 were similar to the growth rates at pH 245
7.5, while the growth rates of AOA-AC2 and AOB-G5-7 differed significantly to their respective 246
rates at pH 7.5 (Fig. 4; Table S6). 247
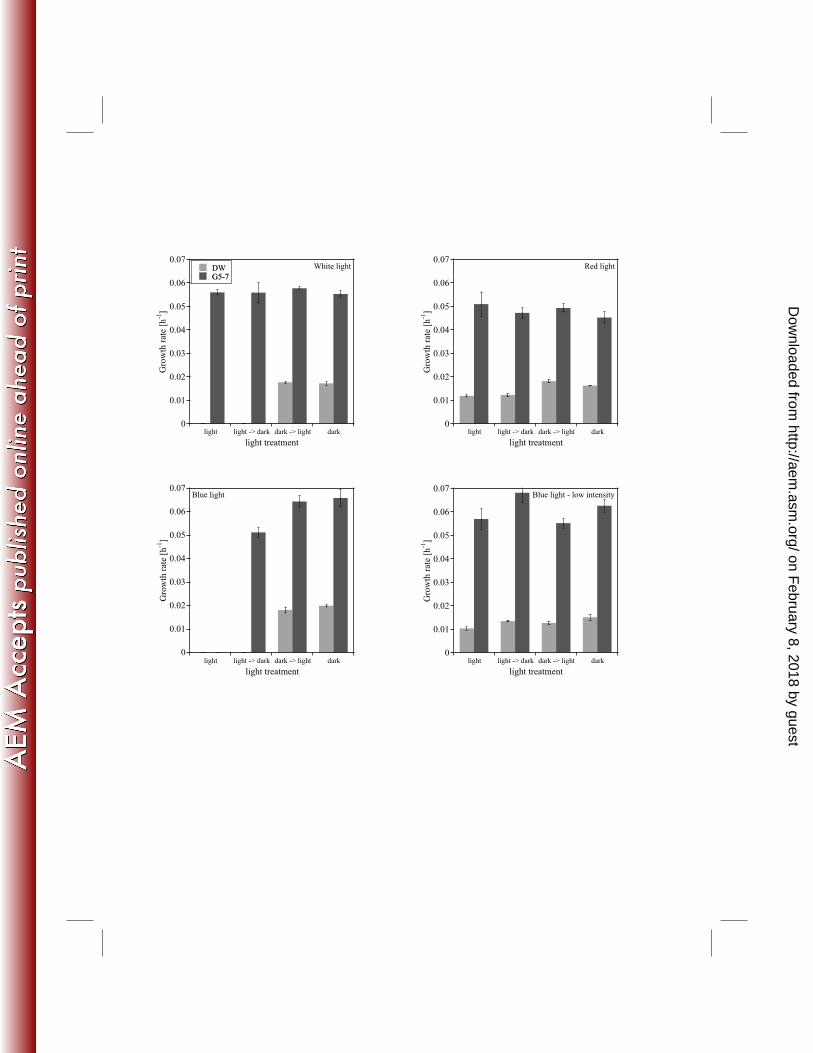
Influence of light on the growth of AOA and AOB: The investigated intensities represent a 248
range at which phytoplankton in freshwater systems is able to grow, but below light saturation 249
(53). White light (30 µmol photons m-2 s-1) strongly inhibited the growth of AOA-DW, but had 250
no effect on AOB-G5-7 (Fig. 5). The AOA did not grow in white light and did not begin to grow 251
after being transferred from the light to the dark. However, growth continued when the AOA 252
cultures were transferred from the dark to the light. To get a better insight into which wavelength 253
of light had the strongest influence on the growth of AOA and AOB, we conducted similar 254
experiments with red (623±3 nm) and blue (470±5 nm) light. Both cultures grew in the red light, 255
but while the growth of AOB-G5-7 was not influenced by the red light, the growth rates of 256
AOA-DW were significantly lower in the red light and after transfer from the light to the dark 257
(Fig. 5; Table S7 and S8). Blue light at 30 µmol photons m-2 s-1 had the strongest effect on the 258
growth of both cultures (Fig. 5). In the blue light none of the cultures grew, and growth of AOA-259
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

13
DW did not recover after transfer from the light to the dark. In contrast, AOB-G5-7 recovered 260
immediately after transfer from the light to the dark, but the growth rate was significantly lower 261
than the growth rate in the continuous dark (Table S7). Transfer of the cultures from the dark 262
into blue light stopped growth immediately. Both cultures grew in the less intense blue light (3 263
µmol photons m-2 s-1) but the growth rate of AOA-DW was significantly lower in the low blue 264
light than in the dark (Fig. 5; Table S8). 265
266
Discussion 267
Enrichment of AOA cultures AOA-DW, AOA-AC2, and AOA-AC5: We enriched and 268
characterized the growth of three different freshwater AOA enrichment cultures belonging to the 269
Thaumarchaeal group I.1a within the newly described phylum Thaumarchaeota (10, 54). One of 270
the cultures, AOA-AC2, is closely related to Ca. Nitrosoarchaeum koreensis, while the other two 271
strains, AOA-AC5 and AOA-DW, are only 70-82% (amoA) and 81-93% (16S rDNA) identical 272
to other cultivated isolates and enrichment cultures such as N. maritimus and Nitrososphaera 273
viennensis (Table 1). This finding indicates that these two enriched AOA belong to a new genus 274
of the ammonia-oxidizing Thaumarchaeota, assuming that the identity between two genera is on 275
average 96.4% based on the 16S rRNA gene (68). This new genus/group includes many 276
ribotypes from non-salt water systems such as freshwater (36, 40) and drinking water systems 277
(61) as well as soil and hot spring environments (69), as indicated by highly identical clones (Fig 278
1). 279
Pure cultures: In this study no pure cultures of the AOA have been obtained thus far. It is safe 280
to assume that the heterotrophic satellite community is providing some compound that enabled 281
the AOA to grow in the enrichment culture. Similar observations have been made with other 282
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

14
AOA as well as for AOB. Potential compounds that positively influence the growth of AOA 283
could be small organic compounds such as pyruvate, which improved growth and enabled 284
isolation of N. viennensis (58). However, the addition of pyruvate during serial dilution did not 285
lead to isolation of any of these strains to date, indicating that different compounds might be 286
important for different AOA. Further research will be necessary to elucidate the interactions 287
between AOA (and AOB) and the heterotrophic satellite bacteria in ammonia oxidizing 288
enrichment cultures. 289
Growth of AOA and AOB: Overall the growth experiments showed that the growth rates of the 290
AOA were almost always lower than the growth rates of the AOB. All our experiments have 291
been conducted under strict chemolithoautotrophic conditions. The results indicate that AOB-292
G5-7 had an advantage over the three tested AOA strains under the conditions investigated. In 293
nature, however, conditions are often less defined with respect to energy generating processes. It 294
has been suggested that not all Thaumarchaeota are chemolithoautotrophic ammonia oxidizers; 295
some carry the amoA gene but are not actively oxidizing NH4+ and others utilize mixotrophic or 296
heterotrophic lifestyles in pure and enrichment cultures (41, 58, 67). Based on these observations 297
and our data one could speculate that AOA in natural samples utilize mixotrophic and/or 298
heterotrophic rather than a completely autotrophic life style, which could explain their success in 299
nature compared to the laboratory. 300
Increasing NH4+ concentrations have different influence on the growth rates and lag phases of 301
AOA and AOB with AOB growing faster and having shorter lag phases than AOA (Fig. 2; Fig. 302
S2; Table S3 and S4). After comparing these results with data provided by other studies that 303
determined the Km of AOA for NH3/NH4+ to be approximately 1000 times lower than Km of AOB 304
(28, 38, 45) we suggest that AOB have an advantage over AOA at higher NH4+ concentrations (> 305
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

15
10µM). This assumption is supported by the detection of high abundances of AOB environments 306
with higher NH4+ input due to fertilization and other processes, while AOA are more abundant in 307
low NH4+ and unfertilized environments (17, 22, 25, 64, 66). 308
The enrichment cultures AOA-DW and AOA-AC5 showed lower tolerance to high NH4+ 309
concentrations than AOA-AC2, with the highest concentrations supporting growth at 1 mM 310
NH4+ (AOA-DW) and 2 mM NH4
+ (AOA-AC5). These concentrations are lower than the highest 311
tolerances towards NH4+ observed for N. viennensis (15 mM), Ca. Nitrosoarchaeum koreensis 312
(10 mM) and enrichment AOA-AC2 (5 mM), a strain closely related to Ca. Nitrosoarchaeum 313
koreensis (28, 58). These results indicate that AOA-DW and AOA-AC5 are less tolerant to high 314
NH4+ concentrations when compared with other AOA isolates and enrichment cultures. Similar 315
observations have been made for AOB; members of the Nitrosomonas oligotropha cluster, which 316
are also commonly found in freshwater environments, are less tolerant to high NH4+ 317
concentrations and better adapted to low NH4+ concentrations, while members of the 318
Nitrosomonas europaea/eutropha cluster are found primarily in environments with high NH4+ 319
concentrations (6, 7, 32, 33, 34). 320
AOA and AOB responded differently when cultured over a range of O2 concentrations. AOA-321
AC5 and AOA-DW grew at all tested O2 concentrations at the same rate, while AOA-AC2 did 322
not grow at 0.5% O2, and the growth rate of AOB-G5-7 decreased with decreasing O2 323
concentrations (Fig. 3; Table S5). Environmental surveys often detected AOA at the oxic-anoxic 324
interface (4, 13, 17, 49) indicating an adaptation to low oxygen conditions. The low Km for O2 325
found for N. maritimus as well as other AOA (28, 38, 45) and the environmental data support the 326
hypothesis that AOA are very likely better adapted to low O2 than AOB and may therefore have 327
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

16
a competitive advantage at the oxic-anoxic interface while AOB are active under more aerobic 328
conditions. 329
AOA and AOB grew at most of the tested pH values, with AOA growing at almost the same rate 330
over a wide pH range and AOB showing a more bell-shaped curve with the highest growth rate 331
at pH 7-7.5 (Fig. 4; Table S6). AOA are found over a wide pH range in different environments 332
such as soils and hot springs (15, 20, 22, 42, 48), but most cultivated AOA such as N. maritimus, 333
N. viennensis, Ca. Nitrosoarchaeum koreensis and Ca. Nitrosotalea devanaterra have rather 334
narrow pH ranges for growth and activity compared with the tested AOA enrichment cultures 335
(28, 31, 58, 60). 336
AOB-G5-7 was more tolerant to light than AOA-DW and also recovered faster after exposure, 337
while AOA-DW did not fully recover from light exposure (Fig. 5; Table S7 and S8). In the 338
environment, maxima of Thaumarchaeal amoA and 16S rRNA copies have been detected at 339
levels below where photosynthetically active radiation (PAR) in the water column dropped to 340
zero, indicating that no light was penetrating to this depth (4, 49). In the same study, AOB and 341
AOA were detected in low abundance in more shallow waters of the Pacific, indicating that AOB 342
as well as some AOA strains could be more tolerant to light than those that are most abundant in 343
the lower parts of the water column (49). The light response of AOA and AOB could be due to 344
differences in the reaction of the copper-containing enzymes to light. AOB are very sensitive to 345
blue near UV light (24, 52). The authors discussed that this inhibition could be attributed to the 346
absorption of light by the oxygenated state of the copper- containing ammonia monooxygenase, 347
which leads to inactivation of the enzyme (52). Genome studies of AOA showed a large number 348
of copper containing enzymes such as multi-copper oxidases and blue copper proteins (5, 65), 349
suggesting that some of the copper-containing enzymes in AOA could be sensitive to light as 350
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

17
well, leading to inhibition of overall metabolism in AOA by light. During the preparation of this 351
manuscript Merbt et al. (2012) published a study investigating the response of two AOB 352
(Nitrosomonas europaea and Nitrosospira multiformis) and two AOA (N. maritimus and C. 353
Nitrosotalea devanaterra) to white light (39). The study confirmed our findings. 354
Conclusion: The results of this study show that AOB are able to outcompete AOA under almost 355
all tested conditions. These findings are in accordance with other cultivation-based studies, as 356
well as observations made in the environment using molecular approaches. Further investigation 357
must be done using other cultivation-based experiments such as continuous cultures, which 358
enable us to cultivate AOA and AOB under more stringently controlled conditions, and in situ 359
incubations which enable us to investigate the response of AOA and AOB to environmental 360
changes in conditions which allow AOA and AOB to utilize metabolic functions as they would 361
naturally in the environment. 362
363
Acknowledgements: We thank Annika Mosier (UC Berkeley) for helpful discussions at the 364
beginning of the project; Michael Vanni and Beth Mette (Department of Zoology, Miami 365
University) for support with sampling; Lynn Johnson (Instrumentation Laboratory, Miami 366
University) for construction of the light installations; Anne Bernhard (Connecticut College, New 367
London) for providing her AOA amoA ARB alignment file, and Anne Morris Hooke 368
(Department of Microbiology; Miami University) for critical reading of the manuscript. This 369
work was supported by startup funds of Miami University and by the National Science 370
Foundation grants no: DEB-1120443 to AB and OCE-0927277 to GB. This paper is dedicated to 371
the memory of Dr. John W. Hawes (Center for Bioinformatics and Functional Genomics and 372
Department of Chemistry and Biochemistry, Miami University). 373
374
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

18
References: 375
1. Arp, D. J., L. A. Sayavedra-Soto, and N. G. Hommes. 2002. Molecular biology and 376
biochemistry of ammonia oxidation by Nitrosomonas europaea. Arch. Microbiol. 377
178:250-255. 378
2. Bateson, M. M. and D. M. Ward. 2008. Methods for extracting DNA from microbial 379
mats and cultivated micro-organisms: high molecular weight DNA from French press 380
lysis. p. 53-60. In G. A. Kowalchuk, F. J. de Bruijn, I. M. Head, A. D. L. Akkermans and 381
J. D. van Elsas (ed.), Molecular Microbial Ecology Manual, Vol 1. Springer: Dordrecht, 382
The Netherlands. 383
3. Belser, L. W. and E. L. Schmidt. 1980. Growth and oxidation kinetics of 3 genera of 384
ammonia-oxidizing nitrifiers. FEMS Microbiol. Lett. 7:213-216. 385
4. Beman, J. M., B. N. Popp, and C. A. Francis. 2008. Molecular and biogeochemical 386
evidence for ammonia oxidation by marine Crenarchaeota in the Gulf of California. 387
ISME Journal 2:429-441. 388
5. Blainey, P. C., A. C. Mosier, A. Potanina, C. A. Francis, and S. R. Quake. 2011. 389
Genome of a Low-salinity ammonia-oxidizing archaeon determined by single-cell and 390
metagenomic analysis. Plos One 6:e16626. 391
6. Bollmann, A. and H. J. Laanbroek. 2001. Continuous culture enrichments of ammonia-392
oxidizing bacteria at low ammonium concentrations. FEMS Microbiol. Ecol. 37:211-221. 393
7. Bollmann, A., M. J. Bär-Gilissen, and H. J. Laanbroek. 2002. Growth at low 394
ammonium concentrations and starvation response as potential factors involved in niche 395
differentiation among ammonia-oxidizing bacteria. Appl. Environ. Microbiol. 68:4751-396
4757. 397
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

19
8. Bollmann, A., I. Schmidt, A. Saunders, and M. Nicolaisen. 2005. Influence of 398
starvation on potential ammonia-oxidizing activity and amoA mRNA levels of 399
Nitrosospira briensis. Appl. Environ. Microbiol. 71:1276-1282. 400
9. Bollmann, A., E. French, and H. J. Laanbroek. 2011. Isolation, cultivation, and 401
characterization of ammonia-oxidizing bacteria and archaea adapted to low ammonium 402
concentrations. Methods in Enzymology 486: 55-88. 403
10. Brochier-Armanet, C., B. Boussau, S. Gribaldo, and P. Forterre. 2008. Mesophilic 404
crenarchaeota: proposal for a third archaeal phylum, the Thaumarchaeota. Nat. Rev. 405
Microbiol. 6: 245-252. 406
11. Chen, G. Y., S. L. Qui, and Y. Y. Zhou. 2009. Diversity and abundance of ammonia-407
oxidizing bacteria in eutrophic and oligotrophic basins of a shallow Chinese lake (Lake 408
Donghu). Res. Microbiol. 160: 173-178. 409
12. Coci, M., P. L. E. Bodelier, and H. J. Laanbroek. 2008. Epiphyton as a niche for 410
ammonia-oxidizing bacteria: Detailed comparison with benthic and pelagic 411
compartments in shallow freshwater lakes. Appl. Environ. Microbiol. 74:1963-1971. 412
13. Coolen, M. J. L., B. Abbas, J. van Bleijswijk, E. C. Hopmans, M. M. M. Kuypers, S. 413
G. Wakeham, and J. S. S. Damste. 2007. Putative ammonia-oxidizing Crenarchaeota in 414
suboxic waters of the Black Sea: a basin-wide ecological study using 16S ribosomal and 415
functional genes and membrane lipids. Environ. Microbiol. 9:1001-1016. 416
14. De Bie, M. J. M., A. G. C. L. Speksnijder, G. A. Kowalchuk, T. Schuurman, G. 417
Zwart, J. R. Stephen, O. E. Diekmann, and H. J. Laanbroek. 2001. Shifts in the 418
dominant populations of ammonia-oxidizing b-subclass proteobacteria along the 419
eutrophic Schelde estuary. Aquat. Microb. Ecol. 23: 225-236. 420
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

20
15. de la Torre, J. R., C. B. Walker, A. E. Ingalls, M. Könneke and D. A. Stahl. 2008. 421
Cultivation of a thermophilic ammonia oxidizing archaeon synthesizing crenarchaeol. 422
Environ. Microbiol. 10:810-818. 423
16. Felsenstein, J. 2005. PHYLIP (Phylogenetic Inference Package) version 3.6. Distributed 424
by the author. Department of Genome Science, University of Washington, Seattle, USA. 425
17. Francis, C. A., K. J. Roberts, J. M. Beman, A. E. Santoro and B. B. Oakley. 2005. 426
Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of 427
the ocean. Proc. Natl. Acad. Sci. 102:14683-14688. 428
18. French, E. and A. Bollmann. Unpublished. 429
19. Fujii, C., T. Nakagawa, Y. Onodera, N. Matsutani, K. Sasada, R. Takahashi, and T. 430
Tokuyama. 2010. Succession and community composition of ammonia-oxidizing 431
archaea and bacteria in bulk soil of a Japanese paddy field. Soil Sci. Plant Nutr. 56:212-432
219. 433
20. Hansel, C. M., S. Fendorf, P. M. Jardine, and C. A. Francis. 2008. Changes in 434
bacterial and archaeal community structure and functional diversity along a 435
geochemically variable soil profile. Appl. Environ. Microbiol. 74:1620-1633. 436
21. Hatzenpichler, R., E. V. Lebedeva, E. Spieck, K. Stoecker, A. Richter, H. Daims, 437
and M. Wagner. 2008. A moderately thermophilic ammonia-oxidizing crenarchaeote 438
from a hot spring. Proc. Natl. Acad. Sci. 105:2134-2139. 439
22. He, J. Z., J. P. Shen, L. M. Zhang, Y. G. Zhu, Y. M. Zheng, M. G. Xu, and H. Di. 440
2007. Quantitative analyses of the abundance and composition of ammonia-oxidizing 441
bacteria and ammonia-oxidizing archaea of a Chinese upland red soil under long-term 442
fertilization practices. Environ. Microbiol. 9:2364-2374. 443
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

21
23. Herrmann, M., A. M. Saunders, and A. Schramm. 2009. Effect of lake trophic status 444
and rooted macrophytes on community composition and abundance of ammonia-445
oxidizing prokaryotes in freshwater sediments. Appl. Environ. Microbiol. 75:3127-3136. 446
24. Hooper, A. B. and K. R. Terry. 1974. Photoinactivation of ammonia oxidation in 447
Nitrosomonas. J. Bacteriol. 119:899-906. 448
25. Jia, Z. and R. Conrad. 2009. Bacteria rather than Archaea dominate microbial ammonia 449
oxidation in an agricultural soil. Environ. Microbiol. 11:1658-1671. 450
26. Johnstone, B. H. and R. D. Jones. 1988. Recovery of a marine chemolithotrophic 451
ammonium-oxidizing bacterium from long-term energy-source deprivation. Can. J. 452
Microbiol. 34:1347-1350. 453
27. Jones, R. D. and R. Y. Morita. 1985. Survival of a marine ammonium oxidizer under 454
energy-source deprivation. Marine Ecol. Prog. Ser. 26:175-179. 455
28. Jung, M.-Y., S.-J. Park, D. Min, J.-S. Kim, W. I. C. Rijpstra, J. S. Sinninghe 456
Damste, G.-J. Kim, E. L. Madsen, and S.-K. Rhee. 2011. Enrichment and 457
characterization of an autotrophic ammonia-oxidizing archaeon of mesophilic 458
crenarchaeal group I.1a from an agricultural soil. Appl. Environ. Microbiol. 77:8635-459
8647. 460
29. Kandeler, E. and H. Gerber. 1988. Short-term assay of soil urease activity using 461
colorimetric determination of ammonium. Biol. Fertil. Soils 6:68-72. 462
30. Klotz, M. G., D. J. Arp, P. S. G. Chain, A. F. El-Sheikh, L. J. Hauser, N. G. 463
Hommes, F. W. Larimer, S. A. Malfatti, J. M. Norton, A. T. Poret-Peterson, L. M. 464
Vergez, and B. B. Ward. 2006. Complete genome sequence of the marine, 465
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

22
chemolithoautotrophic, ammonia-oxidizing bacterium Nitrosococcus oceani ATCC 466
19707. Appl. Environ. Microbiol. 72:6299-6315. 467
31. Könneke, M., A. E. Bernhard, J. R. de la Torre, C. B. Walker, J. B. Waterbury, and 468
D. A. Stahl. 2005. Isolation of an autotrophic ammonia-oxidizing marine archaeon. 469
Nature 437:543-546. 470
32. Koops, H. P. and A. Pommerening-Roeser. 2001. Distribution and ecophysiology of 471
the nitrifying bacteria emphasizing cultured species. FEMS Microbiol. Ecol. 37:1-9. 472
33. Koops, H. P., U. Purkhold, A. Pommerening-Röser, G. Timmermann, and M. 473
Wagner. 2007. The lithoautotrophic Ammonia-oxidizing bacteria, p. 778-811. In M. 474
Dworkin, S. Falkow, E. Rosenberg, K.-H. Schleifer, E. Stackebrandt (ed.). The 475
Prokaryotes 5. Springer New York, United States. 476
34. Kowalchuk, G. A. and J. R. Stephen. 2001. Ammonia-oxidizing bacteria: A model for 477
molecular microbial ecology. Ann. Rev. Microbiol. 55:485-529. 478
35. Lehtovirta-Morley, L. E., K. Stoecker, A. Vilcinskas, J. I. Prosser, and G. W. Nicol. 479
2011. Cultivation of an obligate acidophilic ammonia oxidizer from a nitrifying acid soil. 480
Proc. Natl. Acad. Sci. 108:15892-15897. 481
36. Li, C. and A. Bollmann. Unpublished. 482
37. Ludwig, W., O. Strunk, R. Westram, L. Richter, H. Meier, Yadhukumar, A. 483
Buchner, T. Lai, S. Steppi, G. Jobb, W. Förster, I. Brettske, S. Gerber, A. W. 484
Ginhart, O. Gross, S. Grumann, S. Hermann, R. Jost, A. König, T. Liss, R. 485
Lüssmann, M. May, B. Nonhoff, B. Reichel, R. Strehlow, A. Stamatakis, N. 486
Stuckmann, A. Vilbig, M. Lenke, T. Ludwig, A. Bode, and K.-H. Schleifer. 2004. 487
ARB: a software environment for sequence data. Nucleic Acids Res. 32:1363-1371. 488
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

23
38. Martens-Habbena, W., P. M. Berube, H. Urakawa, J. R. de la Torre and D. A. Stahl. 489
2009. Ammonia oxidation kinetics determine niche separation of nitrifying Archaea and 490
Bacteria. Nature 461:976-981. 491
39. Merbt, S. N., D. A. Stahl, E. O. Casamayor, E. Marti, G. W. Nicol, and J. I. Prosser. 492
2012. Differential photoinhibition of bacterial and archaeal ammonia oxidation. FEMS 493
Microbiol. Lett. 327:41-46. 494
40. Mosier, A. C. and C. A. Francis. 2008. Relative abundance and diversity of ammonia-495
oxidizing archaea and bacteria in the San Francisco Bay estuary. Environ. Microbiol. 496
10:3002-3016. 497
41. Mussmann, M., I. Brito, A. Pitcher, J. S. Sinninghe Damste, R. Hatzenpichler, A. 498
Richter, J. L. Nielsen, P. H. Nielsen, A. Mueller, H. Daims, M. Wagner, and I. M. 499
Head. 2011. Thaumarchaeotes abundant in refinery nitrifying sludges express amoA but 500
are not obligate autotrophic ammonia oxidizers. Proc. Natl. Acad. Sci. 108:16771-16776. 501
42. Nicol, G. W., S. Leininger, C. Schleper, and J. I. Prosser. 2008. The influence of soil 502
pH on the diversity, abundance and transcriptional activity of ammonia oxidizing archaea 503
and bacteria. Environ. Microbiol. 10:2966-2978. 504
43. Nowlin, W. H., J. L. Evarts, and M. J. Vanni. 2005. Release rates and potential fates of 505
nitrogen and phosphorus from sediments in a eutrophic reservoir. Freshwater Biology 506
50:301-322. 507
44. Offre, P., J. I. Prosser, and G. W. Nicol. 2009. Growth of ammonia-oxidizing archaea 508
in soil microcosms is inhibited by acetylene. FEMS Microbiol. Ecol. 70:99-108. 509
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

24
45. Park, B. J., S. J. Park, D. N. Yoon, S. Schouten, J. S. S. Damste, and S. K. Rhee. 510
2010. Cultivation of Autotrophic Ammonia-Oxidizing Archaea from Marine Sediments 511
in Coculture with Sulfur-Oxidizing Bacteria. Appl. Environ. Microbiol. 76:7575-7587. 512
46. Park, H. D. and D. R. Noguera. 2007. Characterization of two ammonia-oxidizing 513
bacteria isolated from reactors operated with low dissolved oxygen concentrations. J. 514
Appl. Microbiol. 102:1401-1417. 515
47. Pernthaler, A., J. Pernthaler, and R. Amann. 2002. Fluorescence in situ hybridization 516
and catalyzed reporter deposition for the identification of marine bacteria. Appl. Environ. 517
Microbiol. 68:3094-3101. 518
48. Reigstad, L. J., A. Richter, H. Daims, T. Urich, L. Schwark, and C. Schleper. 2008. 519
Nitrification in terrestrial hot springs of Iceland and Kamchatka. FEMS Microbiol. Ecol. 520
64:167-174. 521
49. Santoro, A. E., K. L. Casciotti, and C. A. Francis. 2010. Activity, abundance and 522
diversity of nitrifying archaea and bacteria in the central California Current. Environ. 523
Microbiol. 12:1989-2006. 524
50. Sekar, R., A. Pernthaler, J. Pernthaler, F. Warnecke, T. Posch, and R. Amann. 525
2003. An improved protocol for quantification of freshwater Actinobacteria by 526
fluorescence in situ hybridization. Appl. Environ. Microbiol. 69:2928-2935. 527
51. Shand, C. A., B. L. Williams, and G. Coutts. 2008. Determination of N-species in soil 528
extracts using microplate techniques. Talanta 74:648-654. 529
52. Shears, J. H. and P. M. Wood. 1985. Spectroscopic evidence for a photosensitive 530
oxygenated state of ammonia monooxygenase. Biochem. J. 226:499-507. 531
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

25
53. Sigee, D.C. 2005. Freshwater microbiology: biodiversity and dynamic interactions of 532
microorganisms in the aquatic environment. J. Wiley & Sons. Chichester, Great Britain. 533
54. Spang, A., R. Hatzenpichler, C. Brochier-Armanet, T. Rattei, P. Tischler, E. Spieck, 534
W. Streit, D. A. Stahl, M. Wagner, and C. Schleper. 2010. Distinct gene set in two 535
different lineages of ammonia-oxidizing archaea supports the phylum Thaumarchaeota. 536
Trends Microbiol. 18:331-340. 537
55. Speksnijder, A. G. C. L., G. A. Kowalchuk, K. Roest, and H. J. Laanbroek. 1998. 538
Recovery of a Nitrosomonas-like 16S rDNA sequence group from freshwater habitats. 539
Sys. Appl. Microbiol. 21: 321-330. 540
56. Suzuki, I., U. Dular, and S. C. Kwok. 1974. Ammonia or ammonium ion as substrate 541
for oxidation by Nitrosomonas europaea cells and extracts. J. Bacteriol. 120:556-558. 542
57. Tourna, M., T. E. Freitag, G. W. Nicol, and J. I. Prosser. 2008. Growth, activity and 543
temperature responses of ammonia-oxidizing archaea and bacteria in soil microcosms. 544
Environ. Microbiol. 10:1357-1364. 545
58. Tourna, M., M. Stieglmeier, A. Spang, M. Könneke, A. Schintlmeister, T. Urich, M. 546
Engel, M. Schloter, M. Wagner, A. Richter, and C. Schleper. 2011. Nitrososphaera 547
viennensis, an ammonia oxidizing archaeon from soil. Proc. Natl. Acad. Sci. 108:8420-548
8425. 549
59. Treusch, A. H., S. Leininger, A. Kletzin, S. C. Schuster, H. P. Klenk, and C. 550
Schleper. 2005. Novel genes for nitrite reductase and Amo-related proteins indicate a 551
role of uncultivated mesophilic crenarchaeota in nitrogen cycling. Environ. Microbiol. 552
7:1985-1995. 553
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

26
60. Urakawa, H., W. Martens-Habbena, and D. A. Stahl. 2011. Physiology and genomics 554
of ammonia-oxidizing archaea, p. 117-155. In B. B. Ward, D. J. Arp, M. G. Klotz MG 555
(ed.), Nitrification. ASM Press Washington DC, United States. 556
61. Van der Wielen, P. W. J. J., S. Voost, and D. van der Kooij. 2009. Ammonia-557
Oxidizing Bacteria and Archaea in Groundwater Treatment and Drinking Water 558
Distribution Systems. Appl. Environ. Microbiol. 75:4687-4695. 559
62. Venter, J. C., K. Remington, J. F. Heidelberg, A. L. Halpern, D. Rusch, J. A. Eisen, 560
D. Wu, I. Paulsen, K. E. Nelson, W. Nelson, D. E. Fouts, S. Levy, A. H. Knap, M. W. 561
Lomas, K. Nealson, O. White, J. Peterson, J. Hoffman, R. Parsons, H. Baden-562
Tillson, C. Pfannkoch, Y.-H. Rogers, and H. O. Smith. 2004. Environmental genome 563
shotgun sequencing of the Sargasso Sea. Science 304 no5667:66-74. 564
63. Verhagen, F. J. M. and H. J. Laanbroek. 1991. Competition for ammonium between 565
nitryfying and heterotrophic bacteria in dual energy-limited chemostats. Appl. Environ. 566
Microbiol. 57:3255-3263. 567
64. Verhamme, D. T., J. I. Prosser, and G. W. Nicol. (2011). Ammonia concentration 568
determines differential growth of ammonia-oxidizing archaea and bacteria in soil 569
microcosms. ISME Journal 5:1067-1071. 570
65. Walker, C. B., J. R. de la Torre, M. G. Klotz, H. Urakawa, N. Pinel, D. J. Arp, C. 571
Brochier-Armanet, P. S. G. Chain, P. P. Chan, A. Gollabgir, J. Hemp, M. Hügler, E. 572
A. Karr, M. Könneke, M. Shin, T. J. Lawton, T. Lowe, W. Martens-Habbena, L. A. 573
Sayavedra-Soto, D. Lang, S. M. Sievert, A. C. Rosenzweig, G. Manning, and D. A. 574
Stahl. 2010. Nitrosopumilus maritimus genome reveals unique mechanisms for 575
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

27
nitrification and autotrophy in globally distributed marine crenarchaea. Proc. Natl. Acad. 576
Sci. 107:8818-8823. 577
66. Wells, G. F., H. D. Park, C. H. Yeung, B. Eggleston, C. A. Francis, and C. S. 578
Criddle. 2009. Ammonia-oxidizing communities in a highly aerated full-scale activated 579
sludge bioreactor: betaproteobacterial dynamics and low relative abundance of 580
Crenarchaea. Environ. Microbiol. 11:2310-2328. 581
67. Xu, M., J. Schnorr, B. Keibler, and H. M. Simon. 2012. Comparative analysis of 16S 582
rRNA and amoA genes from Archaea selected with organic and inorganic amendments in 583
enrichment culture. Appl. Environ. Microbiol. 78:2137-2146. 584
68. Yarza, P., M. Richter, J. Peplies, J. Euzeby, R. Amann, K.-H. Schleifer, W. Ludwig, 585
F. O. Glöckner, and R. Rosselló-Móra. 2008. The All-Species Living Tree project: A 586
16S rRNA-based phylogenetic tree of all sequenced type strains. Syst. Appl. Microbiol. 587
31:241-250. 588
69. Zhang, C. L., Q. Ye, Z. Huang, W. J. Li, J. Chen, Z. Song, W. Zhao, C. Bagwell, W. 589
P. Inskeep, C. Ross, L. Gao, J. Weigel, C. S. Romanek, E. L. Shock, and B. P. 590
Hedlund. 2008. Global occurence of archaeal amoA genes in terrestrial hot springs. 591
Appl. Environ. Microbiol. 74:6417-6426. 592
593
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

28
Figure legends: 594
Figure 1: Neighbor-joining phylogenetic tree of the AOA enrichment cultures based on amoA 595
gene sequences (595bp). Bootstrap values > 50 of 100 replicates are shown at the 596
nodes. 597
598
Figure 2: Influence of NH4+ concentration on the growth rates of the enrichment cultures AOA-599
AC2; AOA-AC5, AOA-DW, and AOB-G5-7 (mean ± SD; n=3). A: NH4+ 600
concentration linear scale; B: NH4+ concentration logarithmic scale 601
602
Figure 3: Influence of the calculated O2 concentration in the headspace of the bottle on the 603
growth rate of the enrichment cultures AOA-AC2; AOA-AC5, AOA-DW, and AOB-604
G5-7 (mean ± SD; n=3) 605
606
Figure 4: Influence of the pH of the medium on the growth rates of the enrichment cultures 607
AOA-AC2, AOA-AC5, AOA-DW, and AOB-G5-7 (mean ± SD; n=3) 608
609
Figure 5: Influence of white, red, and blue light with the intensity of 30 µmol photons m-2 s-1 and 610
blue light with the intensity of 3 µmol photons m-2 s-1 on the growth rates of the 611
enrichment cultures AOA-DW and AOB-G5-7 (mean ± SD; n=3) 612
613
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from
29
TABLE 1: Identities [%] of AOA in the enrichment cultures AOA-AC2, AOA-AC5, and AOA-614
DW in comparison with previously cultivated AOA. 615
AOA-AC2 AOA-AC5 AOA-DW
amoA 16S amoA 16S amoA 16S
Nitrosopumilus maritimus (31) 88.6 96.2 79.8 92.9 78.8 92.9
Nitrososphaera viennensis (58) 69.6 82.7 70.4 83.7 71.1 83.7
Ca. Nitrososphaera gargensis (21) 70.9 81.9 72.7 82.7 72.1 82.2
Ca. Nitrosocaldus yellowstonii (15) 71.1 80.4 71.1 81.8 70.0 81.0
Ca. Nitrosoarchaeum limnia (5) 94.3 98.4 81.9 92.6 81.5 92.9
Ca. Nitrosoarchaeum koreensis (28) 99.8 99.6 81.6 92.9 81.2 92.8
Ca. Nitrosotalea devanaterra (35) 77.9 88.5 76.7 89.7 76.1 89.3
616
Comparisons are based on 16S rRNA genes (794 bp; corresponding to 109 to 915 in Escherichia 617
coli numbering) and amoA genes (595 bp). 618
619
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

30
TABLE 2: Quantitative analysis of the composition of the enrichment cultures AOA-AC2; 620
AOA-AC5 and AOA-DW. 621
AOA-AC2 AOA-AC5 AOA-DW
Crenarchaeota 91.0 81.2 85.4
Bacteria 9.5 3.3 9.2
Nitrospira (NOB) ND* ND ND
AOB ND ND ND
The cell numbers were determined using CARD-FISH [% of DAPI counts] (n=1). Samples were 622
taken at the end of the logarithmic phase. 623
*ND: not detected. 624
625
626
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

Enrichment AOA-AC2
Candidatus Nitrosoarchaeum koreensis MY1DQ278585 (wastewater)EU651166 (freshwater sediment -SF bay)HQ317041 (wastewater - oil refinery)HQ317037 (wastewater - oil refinery)
EU651167 (freshwater sediment - SF bay)Candidatus Nitrosoarchaeum limnia
Clone BO-D07Clone BO-22
EU650806 (estuarine sediment - SF bay)EU651121 (estuarine sediment - SF bay)
Nitrosopumilus maritimus
Enrichment marine sediment Arctic (FJ656552)Enrichment marine sediment South Korea (FJ656572)
DQ148692 (Black sea)Enrichment marine water CN150 (JF521547)Enrichment marine water CN25 (JF521548)
EU651017 (freshwater sediment - SF bay)
EU852677 (drinking water distribution system)
Clone AC−30
Clone DW−41Clone PH−30
Enrichment AOA-DW
FJ543353 (groundwater)Clone PH−46
FJ543354 (groundwater)
AB516244 (paddy soil)Enrichment AOA-AC5
EU553412 (hot spring)EU553410 (hot spring)
Candidatus Nitrosotalea devanaterraDQ148879 (soil, non-contaminated)
DQ312267 (soil with high nitrogen)FJ227868 (estuarine sediment)
DQ501047 (estuarine sediment)Candidatus Nitrososphaera gargensis
EU651130 (estuarine sediment - SF bay)
EU239968 (hot spring - Yellowstone)Candidatus Nitrosocaldus yellowstonii
Nitrososphaera viennensis
0.1
54
59
61
>99
59
>99
98
92
>99
65
>99
>99
97
54
93
>99
>99
>99
59
63
76
96
73
>99
>99
98
67
>99
>99
soil/sed
imen
tw
ater colu
mn
/sedim
ent
]
]
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
0 1 2 3 4 5
G5-7DWAC2AC5
Gro
wth
rat
e [h
-1]
NH4
+ concentration [mM]
A
0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
0.01 0.1 1 10
G5-7DWAC2AC5
Gro
wth
rat
e [h
-1]
NH4
+ concentration [mM]
B
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

0
0.01
0.02
0.03
0.04
0.05
0.1 1 10 100
G5-7DWAC2AC5
Gro
wth
rat
e [h
-1]
O2 concentration in gas phase [%]
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from
0
0.01
0.02
0.03
0.04
0.05
0.06
5.5 6 6.5 7 7.5 8 8.5 9 9.5
G5-7DWAC2AC5
Gro
wth
rat
e [h
-1]
pH value
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from
0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
light light -> dark dark -> light dark
DWG5-7
Gro
wth
rat
e [h
-1]
light treatment
DWG5-7
White light
0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
light light -> dark dark -> light darkG
row
th r
ate
[h-1
]
light treatment
Red light
0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
light light -> dark dark -> light dark
Gro
wth
rat
e [h
-1]
light treatment
Blue light - low intensity
0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
light light -> dark dark -> light dark
Gro
wth
rat
e [h
-1]
light treatment
Blue light
on February 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from





























