Embed Size (px)
Citation preview

Endogenous Inhibitors of Angiogenesis
Pia Nyberg, Liang Xie, and Raghu Kalluri
Center for Matrix Biology, Department of Medicine, Beth Israel Deaconess Medical Center and Harvard Medical School,Boston, Massachusetts
Abstract
Angiogenesis, the formation of new blood vessels, is requiredfor many pathologic processes, including invasive tumorgrowth as well as physiologic organ/tissue maintenance.Angiogenesis during development and adulthood is likelyregulated by a balance between endogenous proangiogenicand antiangiogenic factors. It is speculated that tumor growthrequires disruption of such balance; thus, the angiogenicswitch must be turned ‘‘on’’ for cancer progression. If theangiogenic switch needs to be turned on to facilitate thetumor growth, the question remains as to what the physiologicstatus of this switch is in the adult human body; is it ‘‘off,’’ withinhibitors outweighing the stimulators, or maintained at afine ‘‘balance,’’ keeping the proangiogenic properties of manyfactors at a delicate ‘‘activity’’ balance with endogenousinhibitors of angiogenesis. The physiologic status of thisbalance is important to understand as it might determine anindividual’s predisposition to turn the switch on duringpathologic events dependent on angiogenesis. Conceivably, ifthe physiologic angiogenesis balance in human populationexists somewhere between off and even balance, an individ-ual’s capacity and rate to turn the switch on might reflect theirnormal physiologic angiogenic status. In this regard, althoughextensive knowledge has been gained in our understanding ofendogenous growth factors that stimulate angiogenesis, theactivities associated with endogenous inhibitors are poorlyunderstood. In this review, we will present an overview of theknowledge gained in studies related to the identification andcharacterization of 27 different endogenous inhibitors ofangiogenesis. (Cancer Res 2005; 65(10): 3967-79)
Introduction
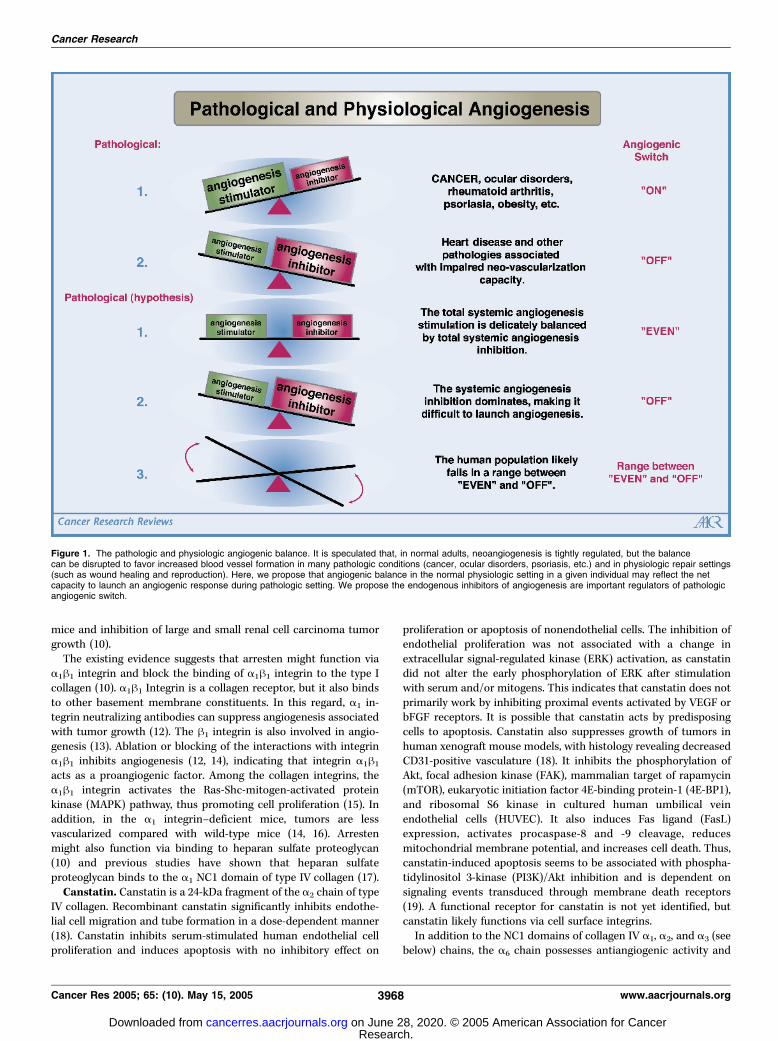
Angiogenesis, the formation of new capillaries, is among the keyevents in various tissue destructive pathologic processes, such astumor growth, metastasis, arthritis, etc., as well as in physiologicprocesses, like organ growth and development, wound healing, andreproduction (1). A hypothesis that tumor growth is angiogenesisdependent was first proposed by Folkman (2). The expansion oftumors is strictly dependent on the neoangiogenesis, and theinhibition of vascular supply to tumors can suppress their growth(3). Solid tumors cannot grow beyond 2 to 3 mm in diameterwithout being able to recruit their own blood supply. Angiogenesisis thought to depend on a delicate balance between endogenousstimulators and inhibitors (Fig. 1). Stimulators of angiogenesisinclude growth factors, such as vascular endothelial growth factor(VEGF), fibroblast growth factor (FGF), and platelet-derived growth
factor, and hypoxic conditions that activate hypoxia-induciblefactor-1, which itself can up-regulate angiogenic proteins, as well asangiogenic oncogenes, such as Ras, and tumor suppressors, such asp53. Endogenous inhibitors of angiogenesis include variousantiangiogenic peptides, hormone metabolites, and apoptosismodulators (Fig. 2; for reviews, see refs. 1, 4). To understand howsystemic angiogenesis balance in the body is regulated byendogenous inhibitors of angiogenesis requires an understandingof how many such molecules exist and their role in inhibitingangiogenesis. This review will highlight the important features ofeach of these molecules and address their role in the regulation ofangiogenesis balance.Vascular basement membrane components can modulate
endothelial cell behavior in addition to providing structural andfunctional support (5). A series of endogenous antiangiogenicfactors have been described, of which many are fragments ofnaturally occurring extracellular matrix (ECM) and basementmembrane proteins (6). In this review, we divide the endogenousinhibitors of angiogenesis into two major classes, matrix-derivedinhibitors and non-matrix-derived inhibitors (Fig. 2).The reader will notice that most studies dealing with
endogenous inhibitors of angiogenesis address therapeutic impactof these agents to control cancer progression when given atpharmacologic doses. Nevertheless, interesting studies are emerg-ing, which also highlight the role of these inhibitors in controllingcancer at physiologic concentrations.
Matrix-Derived Inhibitors of Angiogenesis
Arresten. The main component of vascular basement mem-branes is type IV collagen, forming a mesh-like structure with othermacromolecules, such as laminin, heparan sulfate proteoglycans,fibronectin, and entactin (7). Type IV collagen is composed of sixdistinct gene products (i.e., a1-a6; for a review, see ref. 8). It hasbeen observed earlier that a1 and a2 type IV collagen chainsisolated from the Engelbreth-Holm-Swarm sarcoma tumor areinhibitory to capillary endothelial cell proliferation (9).Arresten is a recently identified endogenous inhibitor of
angiogenesis. It is a 26-kDa molecule derived from the non-collagenous (NC1) domain of the a1 chain of type IV collagen (10).In endothelial cell proliferation assays, a dose-dependent inhibitionof basic FGF (bFGF)–stimulated endothelial cells proliferation isdetected with an ED50 value of as low as 10 nmol/L. Arrestenselectively inhibits endothelial cell tube formation (10). Normally,when mouse aortic endothelial cells are cultured on Matrigel(a solid gel of mouse basement membrane proteins), they rapidlyalign and form hollow tube-like structures (11), but arrestendisrupts with this process. It also inhibits the formation of newblood vessels in Matrigel plug assay in mice (10). Arresten inhibitsendothelial cell proliferation and migration (10). Arresten does nothave a significant effect on the proliferation of several cancer celllines even at very high doses. Arresten affects metastasis leading tosignificant reduction of pulmonary nodules in arresten-treated
Requests for reprints: Raghu Kalluri, Center for Matrix Biology, Department ofMedicine, Beth Israel Deaconess Medical Center and Harvard Medical School, 330Brookline Avenue, Dana 514, Boston, MA 02215. Phone: 617-667-0445; Fax: 617-975-5663; E-mail: [email protected].
I2005 American Association for Cancer Research.
www.aacrjournals.org 3967 Cancer Res 2005; 65: (10). May 15, 2005
Review
Research. on June 28, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from
mice and inhibition of large and small renal cell carcinoma tumorgrowth (10).The existing evidence suggests that arresten might function via
a1h1 integrin and block the binding of a1h1 integrin to the type Icollagen (10). a1h1 Integrin is a collagen receptor, but it also bindsto other basement membrane constituents. In this regard, a1 in-tegrin neutralizing antibodies can suppress angiogenesis associatedwith tumor growth (12). The h1 integrin is also involved in angio-genesis (13). Ablation or blocking of the interactions with integrina1h1 inhibits angiogenesis (12, 14), indicating that integrin a1h1
acts as a proangiogenic factor. Among the collagen integrins, thea1h1 integrin activates the Ras-Shc-mitogen-activated proteinkinase (MAPK) pathway, thus promoting cell proliferation (15). Inaddition, in the a1 integrin–deficient mice, tumors are lessvascularized compared with wild-type mice (14, 16). Arrestenmight also function via binding to heparan sulfate proteoglycan(10) and previous studies have shown that heparan sulfateproteoglycan binds to the a1 NC1 domain of type IV collagen (17).Canstatin. Canstatin is a 24-kDa fragment of the a2 chain of type
IV collagen. Recombinant canstatin significantly inhibits endothe-lial cell migration and tube formation in a dose-dependent manner(18). Canstatin inhibits serum-stimulated human endothelial cellproliferation and induces apoptosis with no inhibitory effect on
proliferation or apoptosis of nonendothelial cells. The inhibition ofendothelial proliferation was not associated with a change inextracellular signal-regulated kinase (ERK) activation, as canstatindid not alter the early phosphorylation of ERK after stimulationwith serum and/or mitogens. This indicates that canstatin does notprimarily work by inhibiting proximal events activated by VEGF orbFGF receptors. It is possible that canstatin acts by predisposingcells to apoptosis. Canstatin also suppresses growth of tumors inhuman xenograft mouse models, with histology revealing decreasedCD31-positive vasculature (18). It inhibits the phosphorylation ofAkt, focal adhesion kinase (FAK), mammalian target of rapamycin(mTOR), eukaryotic initiation factor 4E-binding protein-1 (4E-BP1),and ribosomal S6 kinase in cultured human umbilical veinendothelial cells (HUVEC). It also induces Fas ligand (FasL)expression, activates procaspase-8 and -9 cleavage, reducesmitochondrial membrane potential, and increases cell death. Thus,canstatin-induced apoptosis seems to be associated with phospha-tidylinositol 3-kinase (PI3K)/Akt inhibition and is dependent onsignaling events transduced through membrane death receptors(19). A functional receptor for canstatin is not yet identified, butcanstatin likely functions via cell surface integrins.In addition to the NC1 domains of collagen IV a1, a2, and a3 (see
below) chains, the a6 chain possesses antiangiogenic activity and
Figure 1. The pathologic and physiologic angiogenic balance. It is speculated that, in normal adults, neoangiogenesis is tightly regulated, but the balancecan be disrupted to favor increased blood vessel formation in many pathologic conditions (cancer, ocular disorders, psoriasis, etc.) and in physiologic repair settings(such as wound healing and reproduction). Here, we propose that angiogenic balance in the normal physiologic setting in a given individual may reflect the netcapacity to launch an angiogenic response during pathologic setting. We propose the endogenous inhibitors of angiogenesis are important regulators of pathologicangiogenic switch.
Cancer Research
Cancer Res 2005; 65: (10). May 15, 2005 3968 www.aacrjournals.org
Research. on June 28, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

inhibits tumor growth. Soluble NC1 domain of the a6 chainregulates endothelial cell adhesion and migration (20).Endorepellin. Perlecan is basement membrane heparan sulfate
proteoglycan that plays key roles in vascular growth (21). TheCOOH-terminal end of perlecan, called endorepellin or perlecandomain V, potently inhibits several aspects of angiogenesis:endothelial cell migration, collagen-induced endothelial tubemorphogenesis, and blood vessel growth in the chicken chorioal-lantoic membrane assay and in mouse Matrigel plug assays.Endorepellin is active at nanomolar concentrations in the in vitroassays and blocks endothelial cell adhesion to fibronectin and typeI collagen without directly binding to these matrix proteins (22).Endorepellin binds to endothelial cells as well as to squamous cellcarcinoma cells and breast carcinoma cells via high-affinityreceptors. Interestingly, endorepellin binds endostatin, anothermatrix-derived inhibitor of angiogenesis, and counteracts itsantiangiogenic effects (22). The exact mechanism of endorepellinis not yet known. In this regard, h1 integrin and a-dystroglycanhave been shown to interact with the COOH-terminal domain ofperlecan (a region containing endorepellin; refs. 23–25).Endostatin. Endostatin is an endogenous collagen XVIII–derived
angiogenesis inhibitor identified and purified from murine heman-gioendothelioma cell line (26) and later characterized in mice (27).It corresponds to a 20-kDa fragment derived from the COOH-terminal NC1 domain of type XVIII collagen (26, 28, 29).Recombinant endostatin efficiently blocks angiogenesis and
suppresses primary tumor growth and metastasis in experimentalanimal models without any apparent side effects, toxicity, ordevelopment of drug resistance (26, 30, 31).New insights into the molecular mechanisms associated with
endostatin for inhibition of tumor growth are emerging. Recentstudies have reported that endostatin interferes with FGF-2-induced signal transduction, blocking endothelial cell motility(32), inducing apoptosis (33), causing G1 arrest of endothelial cellsthrough inhibition of cyclin D1 (34), blocking VEGF-mediatedsignaling via direct interaction with the VEGF-R2/KDR/Flk-1receptor tyrosine kinase in HUVECs (35), and blocking tumornecrosis factor–induced activation of c-Jun NH2-terminal kinaseand c-Jun NH2-terminal kinase–dependent proangiogenic geneexpression (36). Endostatin rapidly down-regulates many genes ingrowing endothelial cells, including immediate-early responsegenes, cell cycle–related genes, and genes regulating apoptosisinhibitors, MAPKs, FAKs, and G-protein-coupled receptors medi-ating endothelial cell growth, mitogenic factors, adhesionmolecules, and cell structure components (37). Recently, it isshown that endostatin down-regulates many signaling pathwaysin human microvascular endothelium associated with proangio-genic activity and at the same time up-regulating manyantiangiogenic genes. Endostatin also affects signaling events thatare not associated with angiogenesis, demonstrating the impor-tance of interpathway communications in an intricate signalingnetwork (38).
Figure 2. The endogenous angiogenesis inhibitors.
Endogenous Angiogenesis Inhibitors
www.aacrjournals.org 3969 Cancer Res 2005; 65: (10). May 15, 2005
Research. on June 28, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

Rehn et al. showed that recombinant ‘‘immobilized’’ humanendostatin interacts with a5 and av integrins on the surface ofhuman endothelial cells. Furthermore, this endostatin-integrininteraction is of functional significance in vitro , as immobilizedendostatin promotes integrin-dependent endothelial cell functions(39). Sudhakar et al. have further studied the receptors for solubleendostatin and the downstream signaling events. Endostatin bindsto the a5h1 integrin and inhibits the migration of endothelial cellsby blocking signaling pathways via Ras and Raf and furtherdownstream via ERK1 or p38 (Fig. 3; ref. 40). In the endothelialcells, endostatin induces rapid clustering of a5h1 integrinassociated with actin stress fibers and causes colocalization withthe membrane anchor protein caveolin-1, which couples integrinsto cytoplasmic signaling cascades (41). In these experiments, it wasshown that endostatin binds to both a5h1 and caveolin-1 and thatendostatin treatment induces phosphatase-dependent activation ofcaveolin-associated Src family kinases (42). The disassembly ofactin stress fibers/focal adhesions by endostatin effects cell-matrixinteraction and cell motility via activation of Src and in a tyrosylphosphatase–dependent manner (42). On the other hand, Erikssonet al. conclude that endostatin inhibits chemotaxis, withoutaffecting intracellular pathways known to regulate endothelial cellmigration and proliferation/survival, because they did not find anyeffect of endostatin on phospholipase C-g, Akt/protein kinase B,
p44/42 MAPK, p38 MAPK, and p21-activated kinase activity (43).Moreover, a recent study shows that endostatin action isdependent on expression of E-selectin on the endothelial cells,although a direct binding of endostatin to E-selectin was notobserved (44). Therefore, more work needs to be done to sort outthe exact mechanism of action associated with endostatin.Endostatin binds to heparin (45) and with low affinity to all
surface heparan sulfate proteoglycans that are involved in growthfactor signaling (46–48). The antiangiogenic activity of endostatinseems to depend on the interactions with heparan sulfateproteoglycans possibly by an interaction between discontinuoussulfated domains in heparan sulfate proteoglycans and arginineclusters at the endostatin surface (49). Recent work has shown theidentity of a specific arginine-rich sequence motif of humanendostatin that interacts with endothelial cell surface h1 integrinand heparin and inhibits endothelial cell migration and tubeformation. This Arg-Gly-Asp (RGD)–independent sequence is likelyto be the motif responsible for the antiangiogenic activity ofendostatin (50).Endostatin inhibits the activation and activity of certain matrix
metalloproteinases (MMP; i.e., MMP-2, -9, and -13 and MT1-MMP)and it binds directly to at least MMP-2 and -9 (51–53). However,endostatin does not inhibit all MMPs, because it had no effect onthe activation of MMP-8 (53). In addition to MMPs, endostatin has
Figure 3. Interactions of vascular endothelial cell–associated integrins with matrix-derived angiogenic inhibitors. Basement membrane collagen-derivedendogenous inhibitors of angiogenesis are novel cryptic ligands for various integrins on proliferating endothelial cell surface. It is interesting to note that endostatin andtumstatin, both basement membrane collagen-derived inhibitors, function via distinct integrins to negatively regulate proliferating endothelial cells through uniquesignaling pathways. The functional receptor/s for canstatin are still unknown, although it is known to bind avh3 and a3h1 integrins.
Cancer Research
Cancer Res 2005; 65: (10). May 15, 2005 3970 www.aacrjournals.org
Research. on June 28, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

been shown to interfere with the actions of other proteases, like theplasminogen activator system (54). Interestingly, certain MMPs cangenerate endostatin-containing peptides differing in molecular size(20-30 kDa) from human type XVIII collagen (55). These fragmentsinhibit the proliferation and migration of HUVECs in a similarfashion as native 20-kDa endostatin.1
Endostatin has been shown to significantly reduce invasion ofnot only endothelial cells but also tumor cells into reconstitutedbasement membrane, Matrigel (51). Although the main focus hasbeen on the antiangiogenic activity of endostatin, it should also benoted that endostatin inhibits tumor growth not only by acting viaendothelial cells but also by directly decreasing carcinoma cellmigration. Furthermore, intravasation (the key step of carcinomaprocess leading to metastasis, where tumor cells invade throughblood vessel walls into the bloodstream) of oral carcinoma cells isinhibited by endostatin (53).Endostatin can exist in two forms: as a monomer or as a trimer
that exerts different or even opposite effects (e.g., trimericendostatin in the NC1 domain is needed for endothelial cellmigration, but monomeric endostatin inhibits the migratoryactivity; refs. 56, 57). In addition, it has been shown recently thatendostatin can exist as a soluble globular form or as an insolubleform with abundant cross-h-sheets aggregating into amyloiddeposits. Such different endostatin conformations have distincteffects on plasminogen activation. Insoluble endostatin stimulatesplasminogen activation, whereas soluble endostatin has no effecton plasminogen activation (58).The physiologic levels of circulating endostatin in the serum are
f40 to 100 ng/mL compared with the concentrations ofendostatin (0.2-20 mg/mL) that are effective in the inhibition oftumor growth in various experiments. It has been shown that someof the antiangiogenic and antitumor effects of endostatin might bein fact represent pharmacologic effects at high doses and notnecessarily related to the physiologic function of endostatin (59). Inthis regard, the physiologic levels of endostatin have no effect onthe growth of fibrosarcomas and melanomas in collagen XVIII/endostatin knockout mice (60).Endostatin-like fragment from type XV collagen. Based on a
homology search with endostatin, a 22-kDa fragment of collagen XVwas found with 70% homology to endostatin. It inhibits the migrationof endothelial cells but has no effect on proliferation. Systemicadministration of endostatin-like fragment from type XV collagen(EFC-XV) suppresses the growth of tumors in a xenograft renalcarcinoma model (61). Both NC1 domains of collagen XV and XVIIIcontain a trimerization domain, a hinge region that is more sensitiveto proteolysis in collagen XVIII than collagen type XV. Unlikeendostatin, EFCXV does not bind to zinc or heparin. Both endostatinand EFC-XV inhibit FGF-2-induced or VEGF-induced angiogenesisassociated with chicken chorioallantoic membrane angiogenesis, butthere are striking differences depending onwhich cytokine is used andwhether free EFC-XV or XV-NC1 domains are used (62). Endostatinand EFC-XV show a similar binding repertoire for ECM proteins.However, differences are detected using immunohistologic localiza-tion in vessel walls and basement membrane zones (62). Although itseems that the functions of endostatin andEFC-XVsomewhat overlap,double knockout mice show no additional defects compared with thesingle knockout mice (63).
Anastellin—a fibronectin fragment. Incubation of solublefibronectin with a small fibronectin-derived fragment, calledanastellin, results in a polymeric form of fibronectin that isstrongly antimetastatic in tumor-bearing mice (64). Both anastellinand polymeric fibronectin reduce tumor growth in mice, and thetumors are less vascularized (65). Anastellin is unable to inhibitMatrigel plug angiogenesis in mice that lack plasma fibronectin,but it is fully active in mice that are null for vitronectin, which likefibronectin is a major endothelial cell adhesion protein. Interest-ingly, the activity of endostatin is impaired in both fibronectin-deficient and vitronectin-deficient mice. This suggests a sharedmechanism of action for antiangiogenic factors derived from ECMand plasma proteins (66).Fibulins. Proteolytic digestion of basement membrane prepara-
tions by elastases and cathepsins releases fragments, which possessantiangiogenic activity (67). Two such fragments were COOH-terminal fragments corresponding to fibulin 1D and the domain IIIof fibulin 5.2 Recently, fibulin 5 has been shown to antagonize VEGFsignaling and inhibit endothelial angiogenic sprouting (68). Fibulin5 also binds to extracellular superoxide dismutase (69). In contrast,another report recently shows that fibulin 5 promotes woundhealing in vivo (70). Considering that wound healing is dependenton angiogenesis, this report opposes the notion that fibulin 5 is aninhibitor of angiogenesis. Future studies will hopefully shed morelight on this opposing action of fibulin 5.Thrombospondins. Thrombospondin-1 (TSP-1) was the first
protein to be recognized as a naturally occurring inhibitor ofangiogenesis (71). It is a large multifunctional ECM glycoproteinthat regulates various biological events, like cell adhesion,angiogenesis, cell proliferation and survival, transforming growthfactor-h (TGF-h) activation, and protease activation (ref. 63; for areview, see ref. 72). Some studies suggest that TSP-1 may possessdual activity (proangiogenic and antiangiogenic) depending onproteases that generate fragments of TSP-1 (73, 74). It has beenshown to inhibit tumor growth and metastasis, thus making it apotent inhibitor of in vivo neovascularization and tumorigenesis.Overexpression of TSP-1 in mice suppresses wound healing andtumorigenesis, whereas the lack of functional TSP-1 results inincreased vascularization of selected tissues (75–77). Expression ofTSP-1 has been inversely correlated with malignant progression inbreast and lung carcinomas and melanomas (78). To evaluate theimportance of TSP-1 for the progression of naturally arising tumorsin vivo , Lawler et al. have crossed TSP-1-deficient mice with p53-deficient mice. In the p53-null mice, the absence of TSP-1decreased survival. They also determined more directly whetherhost TSP-1 inhibited tumor growth by implanting melanoma andtesticular teratocarcinoma cells into the TSP-1-null mice. Thetumors grew faster in the TSP-1-null background and exhibited anincrease in vascular density, a decrease in the rate of tumor cellapoptosis, and an increase in the rate of tumor cell proliferation(79). The antiangiogenic activity of TSP-1 has been mapped to thetype 1 repeats and within the NH2-terminal portion of the moleculewithin the procollagen-like domain. TSP-1 and peptides from thetype 1 repeat region (tryptophan-rich, heparin-binding sequencesand TGF-h1 activation sequences) were evaluated in two models ofretinal angiogenesis. TSP-1 inhibited angiogenesis in both exper-imental models, but peptides from the native TSP-1 sequence
2 Xie and Kalluri, unpublished data.1 Heljasvaara et al., unpublished observations.
Endogenous Angiogenesis Inhibitors
www.aacrjournals.org 3971 Cancer Res 2005; 65: (10). May 15, 2005
Research. on June 28, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

containing both the tryptophan-rich repeat and the TGF-h1activation sequence or containing only the tryptophan-rich,heparin-binding sequence had distinct efficiencies in the twomodels. These results suggest that the type 1 repeats of TSP-1contain two subdomains that might independently influence theprocess of neovascularization (80). The existence of two sub-domains also explains how TSP-1 may block FGF-2 and VEGFangiogenic signals by two independent pathways (81). TSP-1 is ableto distinguish pathologic neovascularization from preexistingvasculature due to the dependence of proliferating endothelialcells on Fas/FasL-mediated apoptosis. TSP-1 up-regulates FasLexpression on endothelial cells. Expression for the receptor of FasL,was low on quiescent endothelial cells but greatly enhanced byinducers of angiogenesis, thereby specifically sensitizing thestimulated cells to apoptosis by inhibitor-generated FasL (82).TSP-2 also shows antiangiogenic activity. Injection of TSP-2-
transfected squamous cell carcinoma cells into the dermis of nudemice resulted in inhibition of tumor growth that was even strongerthan the inhibition observed with TSP-1-transfected cells. Thecombined overexpression of TSP-1 and TSP-2 completely pre-vented tumor formation. Extensive areas of necrosis were observedin TSP-2-expressing tumors, and both the density and the size oftumor vessels were significantly reduced (83). Furthermore, tumorangiogenesis was significantly enhanced in TSP-2-deficient mice.Although TSP-2 deficiency did not affect tumor differentiation orproliferation, tumor cell apoptosis was significantly reduced (84).The antiangiogenic role of TSP-2 was further confirmed with animplant system that continuously produces TSP-2. Fibroblasts,which overexpress TSP-2 and transplanted into nude miceresulting in increased levels of circulating TSP-2, inhibited tumorgrowth and angiogenesis of human squamous cell carcinomas,malignant melanomas, and Lewis lung carcinomas implanted at adistant site (85). It has been shown recently that the antiangiogenicregion of TSP-2 lies approximately within the 80-kDa fragment ofthe NH2-terminal globular region (86). Daily injections of TSP-2resulted in a significant inhibition of the growth of humansquamous cell carcinomas in vivo and reduced tumor vasculari-zation. Possible mechanisms for this antiangiogenic activity areinhibition of VEGF-induced endothelial cell migration, tubeformation, and increased endothelial cell–specific apoptosis (86).Tumstatin. Synthetic peptides derived from the NC1 domain of
a3 chain of type IV collagen, residues 183 to 205, have been shown toinhibit the proliferation of melanoma and other epithelial tumor celllines in vitro (87, 88) andbind to theCD47/avh3 integrin complex (89).(89). The entire 28-kDa fragment of a3 chain of NC1 domain of typeIV collagen was named tumstatin (90, 91). In vivo overexpression oftumstatin domains by tumor cells inhibits their invasive propertiesin a mouse melanoma model (92). Tumstatin inhibits formation ofnew blood vessels in Matrigel plug assays and suppresses tumorgrowth of human renal cell carcinoma and prostate carcinoma inmouse xenograft models. This is associated with in vivo endothelialcell–specific apoptosis. The antiangiogenic activity is localized to adistinct region of the tumstatin molecule that is separate from theregion responsible for the antitumor cell activity (90). Tumstatin hastwo binding sites for avh3 integrin, one in the NH2-terminal end ofthe molecule (containing amino acids 54-132) that is associatedwith the antiangiogenic properties and the other in the COOH-terminal end (containing amino acids 185-203) that is associatedwith the antitumor cell activity (89, 91, 93). The tumstatin fragmentcontaining amino acids 54 to 132 binds to both endothelial cells andmelanoma cells but only inhibits proliferation of endothelial cells,
with no effect on tumor cell proliferation. On the other hand, thefragment of tumstatin containing amino acids 185 to 203 binds alsoto both endothelial cells and melanoma cells but only inhibits theproliferation of melanoma cells. The presence of cyclic RGDpeptides do not compete for the avh3 integrin–mediated activityof tumstatin, although this peptide shows significant inhibition ofendothelial cell binding to vitronectin and fibronectin. These twodistinct binding sites on tumstatin suggest unique avh3 integrin–mediated mechanisms governing the two distinct antitumoractivities (91). A third proximal site, RGD site, capable of bindingto avh3 integrin, outside the NC1 domain sequence of tumstatin,has also been proposed recently (94). How these different avh3
integrin sites regulate the antiangiogenic activity of tumstatin is notunderstood completely.The antiangiogenic domain of tumstatin within the 54– to 132–
amino acid region was named Tum-5. Recombinant Tum-5 inhibitsendothelial cell tube formation on Matrigel and induces G1
endothelial cell cycle arrest. Tum-5 possesses antiangiogenicactivity and inhibits human prostate cancer growth in associationwith a decrease in CD31-positive vasculature in nude mice (95).More specifically, the antiangiogenic activity of tumstatin islocalized to a 25–amino acid region within the 69– to 98–aminoacid area as shown with overlapping synthetic peptides. Both ofthese peptides are antiangiogenic and function via avh3 integrin ina similar fashion as the full-length tumstatin (96).The antiangiogenic and proapoptotic activity of tumstatin is
specific for endothelial cells. Maeshima et al. show that tumstatinfunctions as an endothelial cell–specific inhibitor of proteinsynthesis. Through an interaction with avh3 integrin, tumstatininhibits activation of FAK, PI3K, protein kinase B/Akt, and mTORand prevents the dissociation of eukaryotic initiation factor 4Eprotein from 4E-BP1 leading to the inhibition of Cap-dependentprotein synthesis. Furthermore, these results establish a role forintegrins in mediating cell-specific inhibition of Cap-dependentprotein synthesis and suggest a potential mechanism for theselective effects of tumstatin on endothelial cells. Consequently,tumstatin causes endothelial cell apoptosis by inhibiting signalingpathways involved in protein synthesis via interaction with avh3
integrin (97). It is known that endothelial cells adhere toimmobilized VEGF mediated by avh3, a3h1, as well as other av
integrins but not by VEGF receptors (VEGFR). This adhesionalmost totally abolishes endothelial cell apoptosis through theinteraction with integrins. Tumstatin can inhibit the adhesionbetween endothelial cells and immobilized VEGF via the interac-tion with avh3 integrin; thus, it induces apoptosis (98).Although tumstatin and endostatin are both antiangiogenic
molecules derived from the NC1 domains of basement membranecollagens, they share only 14% amino acid homology and theiractivities seem to be mediated via distinct signaling pathways.Human tumstatin prevents angiogenesis via inhibition of endothe-lial cell proliferation and promotion of apoptosis with no effect onmigration, whereas human endostatin prevents endothelial cellmigration with no effect on VEGF-induced proliferation. Thereason for these distinct functions of endostatin and tumstatin canbe explained partly by different integrin-binding capabilities andinduction of different signaling pathways. The activity of tumstatinis mediated via binding to avh3 integrin and inhibition of proteinsynthesis via FAK/PI3K/Akt/mTOR/4E-BP1 pathway, whereas theactivity of endostatin is mediated by a5h1 integrin and inhibition ofFAK/c-Raf/MAPK-ERK kinase-1/2/p38/ERK1 MAPK pathway, withno effect on Cap-dependent protein synthesis (40).
Cancer Research
Cancer Res 2005; 65: (10). May 15, 2005 3972 www.aacrjournals.org
Research. on June 28, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

Does the physiologic level of endogenous circulating tumstatin(f300-360 ng/mL in mice) have an effect on angiogenesis andtumor growth? Tumors in collagen IV a3 chain–deficient mice (thatare concomitantly deficient in tumstatin as well) grow faster thanin wild-type mice. Furthermore, administration of exogenoustumstatin (missing physiologic levels) decreases the growth backto the wild-type level (99). However, physiologic angiogenesisassociated with tissue repair was not affected in these tumstatin-deficient mice. As mentioned before, the antiangiogenic activity oftumstatin is mediated by integrin avh3 (40, 91, 96). This was furtherconfirmed by showing that tumstatin has no effect on theproliferation of mouse lung endothelial cells deficient in integrinh3 or on neovascularization of Matrigel plugs in the integrin h3–null mice (99). A clinical correlation study between levels oftumstatin and tumor progression of human bronchopulmonarycarcinoma was done recently, which highlights the potentialsignificance of tumstatin as an endogenous angiogenesis inhibitor(100). A clear correlation between tumstatin expression and mildlydeveloped vascular networks was observed (100).MMPs are capable of degrading type IV collagen and liberating
fragments containing tumstatin (101). MMP-9 is the most effectivein cleaving tumstatin-containing fragments from type IV collagen,but MMP-2, -3, and -13 also can release tumstatin. MMP-9-deficientmice have decreased levels of tumstatin in their blood, and thetumors in MMP-9-null mice grow faster than the tumors in wild-type mice (99).Recent studies also show that tumstatin peptide can prevent
glomerular hypertrophy in the early stages of diabetic nephropathy(102). Increase in the renal expression of VEGF, Flk-1, and Ang-2was inhibited by tumstatin in mice with diabetic nephropathy(102).
Non-Matrix-Derived Inhibitors of Angiogenesis
Angiostatin. Cleavage of plasminogen (contains five kringles)by proteases results in the formation of 38- to 45-kDaantiangiogenic peptides that contain homologous triple-disulfidebridged kringle domains: kringle-1 to -4 or kringle-1 to -3. Theyare collectively called angiostatin (103–105). Subsequent studieshave also shown that plasminogen kringle-5 by itself alsoexhibits antiangiogenesis activity (106). Angiostatin is a crypticfragment of plasminogen that possesses antiangiogenic proper-ties, a property not shared by the parent molecule (plasmino-gen). Angiostatin was originally purified from serum and urine ofmice bearing s.c. Lewis lung carcinoma, where the growth ofmetastases was inhibited by tumor-generated angiostatin (107).Several members of the human MMP family, including matrilysin(MMP-7) and gelatinase B and A (MMP-9 and -2), metalloelastase(MMP-12), and stromelysin-1 (MMP-3), hydrolyze human plas-minogen to generate angiostatin fragments (104, 108–110) afterplasminogen is converted to plasmin by plasminogen activator,which is followed by reduction by phosphoglycerate kinase (111).Angiostatineen inhibits endothelial cell proliferation and migra-tion (107). It is suggested that the different kringle domains maycontribute to the overall antiangiogenic function of angiostatinby their distinct antimigratory activities (e.g., kringle-4, thepotent fragment in inhibiting endothelial cell migration, whichhas only marginal antiproliferative activity). In contrast, kringle-1to -3, which is equivalent to angiostatin in inhibiting endothelialcell proliferation, manifests only a modest antimigratory effect(112).
There are several targets proposed for the action of angiostatin( for review, see ref. 113). It binds directly to the ATP synthase onthe surface of endothelial cells, which might play a role inallowing the intracellular pH to drop, thus triggering apoptoticevents in endothelial cells (114). Both angiostatin (which containsthe RGD motif) and plasmin specifically bind to avh3 integrin,and angiostatin significantly inhibits plasmin-induced cellmigration. This suggests that that binding of plasmin to avh3
integrin is required for its activity, and angiostatin may interferewith such activity (115, 116). Angiomotin is also a potentialtarget for angiostatin, as it has been shown that cells thatcontained angiomotin are able to bind and internalize angios-tatin, leading to an induction of FAK activity (117). In addition,Claesson-Welsh et al. found that treatment with angiostatin hasno effect on growth factor–induced signal transduction but leadsto a RGD-independent induction of FAK activity. They also showthat angiostatin treatment of endothelial cells in the absenceof growth factors results in increased apoptosis, whereasproliferation does not change. Angiostatin also inhibits migrationand tube formation associated with proliferating endothelialcells (118).Cleaved antithrombin III and prothrombin kringle-2.
Circulating clotting factors in the blood seem to play an importantrole in angiogenesis. In addition to angiostatin (see above), TSPs(see above), and platelet factor-4 (PF-4; see below), the antiangio-genic forms of antithrombin III (119) and prothrombin kringle-2(120) have been shown to posses antiangiogenic properties.Cleavage of the COOH-terminal loop of antithrombin induces aconformational change in the molecule, and the cleaved confor-mation has potent antiangiogenic and antitumor activity in mousemodels. In addition, the latent form of intact antithrombin, whichis similar in conformation to the cleaved molecule, also inhibitsangiogenesis and tumor growth (119). In this regard, prothrombinkringle-2 domain also exhibits anti–endothelial cell proliferativeactivity (120).Chondromodulin-I. Although cartilage contains many angio-
genic factors during ossification, in adults it is an avascular tissue.The 25-kDa cartilage-specific NC1 matrix protein chondromodulin-Iis a strong inhibitor of angiogenesis (121). The expression patternof the gene suggests a role for chondromodulin-I in themorphogenesis during embryonic development (122). The levelof chondromodulin-I transcripts is substantially reduced inchondrosarcomas or in other benign cartilage tumors. Humanchondrosarcoma cells in nude mice produce tumors withcartilaginous matrix. Local administration of recombinant humanchondromodulin-I almost completely blocks vascular invasionand tumor growth in vivo . Furthermore, it inhibits the growth ofcolon adenocarcinoma in vivo , implying therapeutic potential forother solid tumors (123).Soluble Fms-like tyrosine kinase 1. The soluble version of
VEGFR-1 [soluble Fms-like tyrosine kinase 1 (sFlt-1)] was identifiedby Kendall et al. (124, 125). In addition to the full-length receptor,VEGFR-1 gene encodes for a small soluble form (sFlt-1) carryingonly six Ig domain via an attenuate splicing (124, 126, 127). sFlt-1has a strong affinity for VEGF and placental growth factor (124,128, 129). In human, VEGF circulates at 75 pg/mL and sFlt-1circulates at a concentration of 23 ng/mL (130). Therefore, VEGFcan circulate in a bound state (bound to sFlt-1) as well as in freestate depending on the amount of sFlt-1 in the circulation. In thisregard, several reports have suggested that sFlt-1 can serve as anantitumor agent by inhibiting VEGF (130–132). The relevance for
Endogenous Angiogenesis Inhibitors
www.aacrjournals.org 3973 Cancer Res 2005; 65: (10). May 15, 2005
Research. on June 28, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

sFlt-1 at the normal physiologic concentration in the regulation ofcancer progression is not understood.Interferons. IFNs, pleiotropic cytokines that regulate antiviral,
antitumor, apoptotic, and cellular immune responses, were the firstendogenous antiangiogenic regulators identified. IFNs inhibitangiogenesis induced by tumor cells in mice (133). IFN-a or IFN-h has biological activity against squamous cell carcinomas andinhibits angiogenesis in tumor-bearing nude mice. When tumorcells are treated in vitro with low-dose IFN-a, there is a significantdrop in their secretion of interleukin (IL)-8, the major angiogenicfactor produced by tumors (134). IFN-a reduces urokinase-typeplasminogen activator and plasminogen activator inhibitor-1activity (135) and significantly inhibits MMP-9 enzymatic activityand protein expression (136). IFN-a and IFN-h treatmentalso inhibits angiogenesis by down-regulation of bFGF expression(137–139), but other studies suggest that the action of IFN-a is notmediated by bFGF or VEGF (140–142).Interleukins. ILs are a family of leukocyte-derived proteins with
broad-ranging effects on multiple physiologic properties, includingangiogenesis. ILs bearing a NH2-terminal Glu-Leu-Arg (ERL) motif,such as IL-8, tend to display proangiogenic properties, whereasthose lacking this motif have been found to inhibit angiogenesis(143). IL-1, mainly secretable IL-1h, is involved in inflammation,tumor growth, and metastasis. It is shown that IL-1h inhibits FGF-stimulated angiogenesis by an autocrine pathway (144). However,recent study found that the IL-1 receptor antagonist reducedinflammatory responses and inhibited tumor development inmice (145). IL-4 inhibits bFGF-induced angiogenesis (146). IL-12and IL-18 are IFN-g-inducing cytokines. IL-12 can inhibitangiogenesis through the downstream chemokines, such as IFN-inducible protein-10 and monokine induced by IFN-g (147–149).Tumors in mice treated with IL-12 are significantly smaller andhave extensive necrosis compared with untreated tumors (150).The combination of IL-12 and IL-2 mediated synergistic antitumoractivity in preclinical tumor models (151). The IL-12 plasmid DNAgene transfer significantly prevented the growth and vasculariza-tion of highly angiogenic sarcoma and TS/A murine mammarycarcinoma tumors in nude and/or syngeneic mice (152). IL-18inhibits FGF-stimulated endothelial cell proliferation in vitro andsuppresses the FGF-induced corneal neovascularization by sys-temic administration in mice (153). IL-18 can also inhibitembryonic angiogenesis.2-Methoxyestradiol. 2-Methoxyestradiol (2-ME), an endogenous
estradiol metabolite, is an inhibitor of angiogenesis, with directeffect on cancer cells (154–157). 2-ME is orally active molecule withlow affinity for estrogen receptors (158). Its mechanism of action isattributed to its ability to bind to the colchicine binding site oftubulin and the inhibition of superoxide dismutase enzymaticactivity (154, 159–161). Recently, another novel mechanism for2-ME was proposed (162). 2-ME-mediated destabilization ofmicrotubules was associated with a block in nuclear accumulationand activity of hypoxia-inducible factor-1a (oxygen and protea-some–independent pathway), leading to significant reduction inthe VEGF levels (162). The endogenous physiologic role for 2-ME asan inhibitor of angiogenesis is not yet understood.Pigment epithelium-derived factor. Because pigment epithe-
lium-derived factor (PEDF), a noninhibitory member of the serpinsuperfamily, was identified to be responsible for the avascularity ofocular compartments in 1999, it is the most potent inhibitor ofangiogenesis in the mammalian eye and is involved in thepathogenesis of angiogenic eye diseases, such as proliferative
diabetic retinopathy (82, 163–167). It also has neurotrophic activityboth in retina and in the central nervous system. PEDF expressionis suppressed by hypoxia, and changes in PEDF correlate with thedevelopment of retinal neovascularization in animal models ofhypoxic eye disease.PEDF possesses several physiologic properties that make it a
potentially important protein in the regulation of angiogenesis, inthe neuronal cell survival, and in the protection of neurons fromneurotoxic agents. Its antiangiogenic activity is selective, in thatPEDF targets only new vessel growth but spares existing ones, andit is reversible. It is a protein that is highly up-regulated in the Go
phase of early-passage cells compared with rapidly proliferating orsenescent cells and thus is also linked to both cell cycle and cellsenescence (168, 169).Recent study highlights two beneficial effects of PEDF treatment
on tumor growth and expansion. One is the suppression of tumorangiogenesis. Overexpression of PEDF was found to significantlyinhibit melanoma growth and vessel formation in G361 nude micexenografts (170). PEDF is also expressed strongly in normal murinekidney, and the loss of angioinhibitory activity may contribute topathologic angiogenesis in Wilms’ tumor (171). In addition, PEDFmay serve as a multifunctional antitumor agent in neuroblastomas,inhibiting angiogenesis while promoting the numbers of Schwanncells and differentiated tumor cells that in turn produce PEDF(172). The loss of PEDF expression was also detected in gliomaprogression (173). In PEDF-deficient mice, stromal vessels wereincreased and associated with epithelial cell hyperplasia (174). It isa key inhibitor of stromal vasculature and epithelial tissue growthin mouse prostate and pancreas. The other activity of PEDF isinduction of FasL-dependent apoptosis in tumor cells. PEDF up-regulates FasL on endothelial cells. Expression of the essentialpartner of FasL, Fas/CD95 receptor, was low on quiescentendothelial cells but greatly enhanced by inducers of angiogenesis,thereby specifically sensitizing the stimulated cells to apoptosis byinhibitor-generated FasL. The antiangiogenic activity of PEDF, bothin vitro and in vivo , was dependent on this dual induction of Fasand FasL and the resulting apoptosis (82).PEDF can be given therapeutically as a soluble protein or by
viral-mediated gene transfer (175, 176). It is stable and nontoxicwhen injected systemically. Gene transfer of PEDF suppressestumor vascularization and growth while prolonging survival insyngeneic murine models of thoracic malignancies. Gene transferof PEDF using adeno-associated viral vectors also inhibitedischemia-induced neovascularization (177).A recent study suggests that, in physiologic conditions, a critical
balance between PEDF and VEGF exists, and PEDF may counteractthe angiogenic potential of VEGF. Under oxidative stress, PEDFdecreases, disrupting the angiogenic balance (178). This criticalbalance between PEDF and VEGF is important to prevent thedevelopment of choroidal neovascularization (163, 179). In addition,bone angiogenesis and matrix modeling may also be mediated bothby dynamic intraplay between PEDF and by VEGF (180).PEX. It has been shown that PEX, a noncatalytic COOH-
terminal hemopexin-like domain of MMP-2, prevents binding ofMMP-2 to integrin avh3, thus inhibiting proteolytic activity on thecell surface and disrupting angiogenesis (181). Delivery of PEX byviral vectors resulted in suppressed endothelial cell invasion andformation of capillary-like structures, blocking of bFGF-inducedMMP-2 activation and angiogenesis, and inhibition of tumor-induced angiogenesis and tumor growth in nude mice (182). PEXformation seems to depend on the stage of angiogenesis, reaching
Cancer Research
Cancer Res 2005; 65: (10). May 15, 2005 3974 www.aacrjournals.org
Research. on June 28, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

the maximum level during the late stage when the vessels arematurating (181).Platelet factor-4. PF-4 is a protein released from platelet
a-granules during platelet aggregation that has been shown to haveantiangiogenic properties both in vitro and in vivo (183).Recombinant human PF-4 inhibits blood vessel proliferation inthe chicken chorioallantoic membrane assay in a dose-dependentmanner. PF-4 inhibits angiogenesis by associating directly withFGF-2, inhibiting its dimerization and blocking FGF-2 binding toendothelial cells. The inhibitory activities are associated with theCOOH-terminal, heparin-binding region of the molecule (183) andmore specifically in the short COOH-terminal segment (amino acids47-70). A minor modification of this active segment of PF-4, anaddition of a ERL, a critical domain present in proangiogenicchemokines, surprisingly elicits several times greater antiangiogenicpotential than the original peptide. Thus, tailored PF-4 peptidesrepresent a new class of antiangiogenic agents with a defined modeof action and a strong in vivo activity (184). Protein assays show thatPF-4 and FGF-2 bind to adjacent or overlapping sites togethercovering a 12-kDa stretch of heparan sulfate, suggesting that thesethree components may form a ternary complex. This enables PF-4to inhibit FGF-2-stimulated endothelial cell proliferation throughheparan sulfate–dependent mechanism (185).Prolactin fragment. The intact prolactin (23 kDa) is enzymat-
ically cleaved in several different tissues to generate a 16-kDa (16KPRL) and a 8-kDa fragment (186). Although the intact prolactin hasactivities consistent with proangiogenesis, the generation of the16K PRL fragment exposes a cryptic antiangiogenic activity (187,188). The opposing angiogenesis activities of intact prolactin andNH2-terminal 16K PRL offers an insight into the body’s mechanismto regulate angiogenesis (189). The mechanism by which suchopposing action is regulated can in part be explained by differentialregulation of MAPK signaling pathway (190). Additionally, the 16KPRL inhibits VEGF-induced activation of Ras in capillaryendothelial cells (191).Tissue inhibitors of matrix metalloproteinases. Tissue
inhibitors of matrix metalloproteinases (TIMP) suppress MMPactivity and ECM turnover. In addition to their MMP inhibitoryactivity, TIMPs have pluripotent effects on cell growth, apoptosis,and differentiation (192–194). TIMP-2 inhibits angiogenic factor–induced endothelial cell proliferation in vitro and angiogenesisin vivo , independent of MMP inhibition. These effects requirea3h1 integrin–mediated binding of TIMP-2 to endothelial cells.Further, TIMP-2 induces a decrease in total protein tyrosinephosphatase activity associated with h1 integrin subunit as wellas dissociation of the phosphatase SHP-1 from h1 integrin. TIMP-2 treatment also results in an increase in protein tyrosinephosphatase activity associated with tyrosine kinase receptorsFGF receptor-1 and KDR (195).Troponin I. Troponin I (Tn I) is a novel cartilage-derived
angiogenesis inhibitor, which inhibits endothelial cell proliferationand angiogenesis in both in vivo and in vitro model systems. Tn Ialso inhibits metastasis of a wide variety of tumors in vivo . Tn I is asubunit of the troponin complex, which along with tropomyosin isresponsible for the calcium-dependent regulation of striatedmuscle contraction (196). Interestingly, Tn I is capable of inhibitingactomyosin ATPase, which might constitute for its angiogenesis,tumor growth, and metastasis inhibitory activity (196). Tn I inhibitsboth bFGF-stimulated and basal levels of endothelial cellproliferation probably via an interaction of Tn I with the cellsurface bFGF receptor on capillary endothelial cells (197).
Vasostatin. Vasostatin, a NH2-terminal domain of humancalreticulin inclusive of amino acids 1,180, is a potent angiogenesisinhibitor. It selectively inhibits endothelial cell proliferation andangiogenesis in response to stimulation from growth factors andsuppresses tumor growth (198, 199). Burkitt, colon, and ovariantumors in mice treated with vasostatin were smaller than tumors incontrol mice (150). Combination of vasostatin and IP-10 reducedtumor growth more effectively than each agent alone, but completeregression was not observed (200). Vasostatin specifically inhibitsendothelial cell attachment to laminin and reduces subsequentendothelial cell growth induced by bFGF (201). Furthermore, genetherapy experiments with intramuscular delivery of vasostatin DNAis effective in the inhibition of angiogenesis and tumor growth inmurine tumor models (202).
Summary and Future Perspective
Relative systemic levels of proangiogenic and antiangiogenicfactors likely govern tumor progression by regulating the‘‘angiogenic balance.’’ Conversion of dormant carcinomas to aninvasive malignant carcinomas is considered to involve a shift infavor of enhanced angiogenesis potential. Influenced by oncogenesand tumor suppressor genes, disruption of the ‘‘angiogeniccheckpoint’’ via increase in angiogenic factors, such as VEGF, ordecrease in the physiologic levels of endogenous inhibitors ofangiogenesis, like TSP-1, tumstatin, and endostatin, could representan important step in the progression of cancer. Currently, at least27 different protein and small molecules are known to exist in thebody that function as inhibitors of angiogenesis. Therefore, it isquite possible that genetic control of the physiologic levels ofendogenous inhibitors of angiogenesis might constitute a criticalline of defense against the conversion of dormant neoplastic eventsinto a malignant phenotype of cancer.How these endogenous inhibitors orchestrate the formation of a
barrier to counteract the effects of physiologic levels of proangio-genic growth factors and cytokines is not yet understood. Forendogenous inhibitors that need to be generated via proteolysis oflarger precursor proteins, regulation of enzymatic activity alsolikely plays a key role in the formation of the angiogenic barrier.Further studies are needed to address the function of physiologiclevels of these 27 inhibitors in controlling cancer progression. It isconceivable that genetic defects reflected by loss-of-function singlenucleotide polymorphisms in the sequence of the protein inhibitorsor enzymes that generate angiogenesis inhibitors could determinehow fast a set of neoplastic events switch from a nonlethal lesion toa malignant and angiogenic tumor.Lastly, the potential therapeutic application for these endoge-
nous inhibitors of angiogenesis is being considered in the clinicand early results from clinical trials with angiostatin, endostatin,TSP-1 (ABT-510), and 2-ME (Panzam) suggest that more laboratorystudies are required to better understand the mechanism of actionassociated with each of these drug candidates (203–209). Suchcontinuing efforts will enable us to design more relevant andtargeted clinical trial guided by the inherent antiangiogenicmechanism of each particular inhibitor.
Acknowledgments
Received 7/13/2004; revised 12/1/2004; accepted 2/24/2005.Grant support: NIH grants DK 55001 and DK 62987, Center for Matrix Biology at
the Beth Israel Deaconess Medical Center research funds, Emil Aaltonen Foundation,Ann L. and Herbert J. Siegel Philantropic Jewish Communal Fund, and Joseph LubranoMemorial Goldfield Family Charitable trust fund.
Endogenous Angiogenesis Inhibitors
www.aacrjournals.org 3975 Cancer Res 2005; 65: (10). May 15, 2005
Research. on June 28, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

References1. Folkman J. Angiogenesis in cancer, vascular, rheuma-toid and other disease. Nat Med 1995;1:27–31.
2. Folkman J. Tumor angiogenesis: therapeutic implica-tions. N Engl J Med 1971;285:1182–6.
3. Hanahan D, Folkman J. Patterns and emergingmechanisms of the angiogenic switch during tumori-genesis. Cell 1996;86:353–64.
4. Folkman J. Angiogenesis and apoptosis. Semin CancerBiol 2003;13:159–67.
5. Darland DC, D’Amore PA. Blood vessel maturation:vascular development comes of age. J Clin Invest 1999;103:157–8.
6. Cao Y. Endogenous angiogenesis inhibitors and theirtherapeutic implications. Int J Biochem Cell Biol 2001;33:357–69.
7. Timpl R. Macromolecular organization of basementmembranes. Curr Opin Cell Biol 1996;8:618–24.
8. Prockop DJ, Kivirikko KI. Collagens: molecular biology,diseases, and potentials for therapy. Annu Rev Biochem1995;64:403–34.
9. Madri JA. Extracellular matrix modulation of vascularcell behaviour. Transpl Immunol 1997;5:179–83.
10. Colorado PC, Torre A, Kamphaus G, et al. Anti-angiogenic cues from vascular basement membranecollagen. Cancer Res 2000;60:2520–6.
11. Grant DS, Kibbey MC, Kinsella JL, Cid MC,Kleinman HK. The role of basement membrane inangiogenesis and tumor growth. Pathol Res Pract1994;190:854–63.
12. Senger DR, Claffey KP, Benes JE, Perruzzi CA,Sergiou AP, Detmar M. Angiogenesis promoted byvascular endothelial growth factor: regulation througha1h1 and a2h1 integrins. Proc Natl Acad Sci U S A1997;94:13612–7.
13. Bloch W, Forsberg E, Lentini S, et al. h1 Integrin isessential for teratoma growth and angiogenesis. J CellBiol 1997;139:265–78.
14. Pozzi A, Moberg PE, Miles LA, Wagner S, Soloway P,Gardner HA. Elevated matrix metalloprotease andangiostatin levels in integrin a1 knockout mice causereduced tumor vascularization. Proc Natl Acad Sci U S A2000;97:2202–7.
15. Pozzi A, Wary KK, Giancotti FG, Gardner HA. Integrina1h1 mediates a unique collagen-dependent prolifera-tion pathway in vivo . J Cell Biol 1998;142:587–94.
16. Pozzi A, LeVine WF, Gardner HA. Low plasma levelsof matrix metalloproteinase 9 permit increased tumorangiogenesis. Oncogene 2002;21:272–81.
17. Keller KM, Keller JM, Kuhn K. The C-terminus of typeI collagen is a major binding site for heparin. BiochimBiophys Acta 1986;882:1–5.
18. Kamphaus GD, Colorado PC, Panka DJ, et al.Canstatin, a novel matrix-derived inhibitor of angioge-nesis and tumor growth. J Biol Chem 2000;275:1209–15.
19. Panka DJ, Mier JW. Canstatin inhibits Akt activationand induces Fas-dependent apoptosis in endothelialcells. J Biol Chem 2003;278:37632–6.
20. Petitclerc E, Boutaud A, Prestayko A, et al. Newfunctions for non-collagenous domains of humancollagen type IV. Novel integrin ligands inhibitingangiogenesis and tumor growth in vivo . J Biol Chem2000;275:8051–61.
21. Yurchenco PD, O’Rear JJ. Basal lamina assembly.Curr Opin Cell Biol 1994;6:674–81.
22. Mongiat M, Sweeney SM, San Antonio JD, Fu J,Iozzo RV. Endorepellin, a novel inhibitor of angio-genesis derived from the C terminus of perlecan. J BiolChem 2003;278:4238–49.
23. Hayashi K, Madri JA, Yurchenco PD. Endothelial cellsinteract with the core protein of basement membraneperlecan through h1 and h3 integrins: an adhesionmodulated by glycosaminoglycan. J Cell Biol 1992;119:945–59.
24. Brown JC, Sasaki T, Gohring W, Yamada Y, Timpl R.The C-terminal domain V of perlecan promotes h1
integrin-mediated cell adhesion, binds heparin, nidogenand fibulin-2 and can be modified by glycosaminogly-cans. Eur J Biochem 1997;250:39–46.
25. Talts JF, Andac Z, Gohring W, Brancaccio A, Timpl R.Binding of the G domains of laminin a1 and a2 chainsand perlecan to heparin, sulfatides, a-dystroglycan and
several extracellular matrix proteins. EMBO J 1999;18:863–70.
26. O’Reilly MS, Boehm T, Shing Y, et al. Endostatin: anendogenous inhibitor of angiogenesis and tumorgrowth. Cell 1997;88:277–85.
27. Standker L, Schrader M, Kanse SM, Jurgens M,Forssmann WG, Preissner KT. Isolation and character-ization of the circulating form of human endostatin.FEBS Lett 1997;420:129–33.
28. Rehn M, Hintikka E, Pihlajaniemi T. Primary structureof the a1 chain of mouse type XVIII collagen, partialstructure of the corresponding gene, and comparison ofthe a1(XVIII) chain with its homologue, the a1(XV)collagen chain. J Biol Chem 1994;269:13929–35.
29. Oh SP, Kamagata Y, Muragaki Y, Timmons S, OoshimaA, Olsen BR. Isolation and sequencing of cDNAs forproteins with multiple domains of Gly-Xaa-Yaa repeatsidentify a distinct family of collagenous proteins. ProcNatl Acad Sci U S A 1994;91:4229–33.
30. Marneros AG, Olsen BR. The role of collagen-derivedproteolytic fragments in angiogenesis. Matrix Biol 2001;20:337–45.
31. Boehm T, Folkman J, Browder T, O’Reilly MS.Antiangiogenic therapy of experimental cancer doesnot induce acquired drug resistance. Nature 1997;390:404–7.
32. Dixelius J, Cross M, Matsumoto T, Sasaki T, Timpl R,Claesson-Welsh L. Endostatin regulates endothelial celladhesion and cytoskeletal organization. Cancer Res 2002;62:1944–7.
33. Dhanabal M, Volk R, Ramchandran R, Simons M,Sukhatme VP. Cloning, expression, and in vitro activityof human endostatin. Biochem Biophys Res Commun1999;258:345–52.
34. Hanai J, Dhanabal M, Karumanchi SA, et al.Endostatin causes G1 arrest of endothelial cellsthrough inhibition of cyclin D1. J Biol Chem 2002;277:16464–9.
35. Kim YM, Hwang S, Pyun BJ, et al. Endostatin blocksvascular endothelial growth factor-mediated signalingvia direct interaction with KDR/Flk-1. J Biol Chem 2002;277:27872–9.
36. Yin G, Liu W, An P, et al. Endostatin gene transferinhibits joint angiogenesis and pannus formation ininflammatory arthritis. Mol Ther 2002;5:547–54.
37. Shichiri M, Hirata Y. Antiangiogenesis signals byendostatin. FASEB J 2001;15:1044–53.
38. Abdollahi A, Hahnfeldt P, Maercker C, et al. Endo-statin’s antiangiogenic signaling network. Mol Cell 2004;13:649–63.
39. Rehn M, Veikkola T, Kukk-Valdre E, et al. Interactionof endostatin with integrins implicated in angiogenesis.Proc Natl Acad Sci U S A 2001;98:1024–9.
40. Sudhakar A, Sugimoto H, Yang C, Lively J, Zeisberg M,Kalluri R. Human tumstatin and human endostatinexhibit distinct antiangiogenic activities mediated byavh3 and a5h1 integrins. Proc Natl Acad Sci U S A 2003;100:4766–71.
41. Wary KK, Mariotti A, Zurzolo C, Giancotti FG. Arequirement for caveolin-1 and associated kinase Fyn inintegrin signaling and anchorage-dependent cellgrowth. Cell 1998;94:625–34.
42. Wickstrom SA, Alitalo K, Keski-Oja J. Endostatinassociates with integrin a5h1 and caveolin-1, andactivates Src via a tyrosyl phosphatase-dependentpathway in human endothelial cells. Cancer Res 2002;62:5580–9.
43. Eriksson K, Magnusson P, Dixelius J, Claesson-WelshL, Cross MJ. Angiostatin and endostatin inhibit endo-thelial cell migration in response to FGF and VEGFwithout interfering with specific intracellular signaltransduction pathways. FEBS Lett 2003;536:19–24.
44. Yu Y, Moulton KS, Khan MK, et al. E-selectin isrequired for the antiangiogenic activity of endostatin.Proc Natl Acad Sci U S A 2004;101:8005–10.
45. Dixelius J, Larsson H, Sasaki T, et al. Endostatin-induced tyrosine kinase signaling through the Shbadaptor protein regulates endothelial cell apoptosis.Blood 2000;95:3403–11.
46. Karumanchi SA, Jha V, Ramchandran R, et al. Cellsurface glypicans are low-affinity endostatin receptors.Mol Cell 2001;7:811–22.
47. Hohenester E, Sasaki T, Olsen BR, Timpl R. Crystal
structure of the angiogenesis inhibitor endostatin at 1.5A resolution. EMBO J 1998;17:1656–64.
48. Sasaki T, Larsson H, Kreuger J, et al. Structural basisand potential role of heparin/heparan sulfate binding tothe angiogenesis inhibitor endostatin. EMBO J 1999;18:6240–8.
49. Kreuger J, Matsumoto T, Vanwildemeersch M, et al.Role of heparan sulfate domain organization in endo-statin inhibition of endothelial cell function. EMBO J2002;21:6303–11.
50. Wickstrom SA, Alitalo K, Keski-Oja J. An endostatin-derived peptide interacts with integrins and regulatesactin cytoskeleton and migration of endothelial cells.J Biol Chem 2004.
51. Kim YM, Jang JW, Lee OH, et al. Endostatin inhibitsendothelial and tumor cellular invasion by blocking theactivation and catalytic activity of matrix metallopro-teinase. Cancer Res 2000;60:5410–3.
52. Lee SJ, Jang JW, Kim YM, et al. Endostatin binds tothe catalytic domain of matrix metalloproteinase-2.FEBS Lett 2002;519:147–52.
53. Nyberg P, Heikkila P, Sorsa T, et al. Endostatininhibits human tongue carcinoma cell invasion andintravasation and blocks the activation of matrixmetalloprotease-2, 9 and -13. J Biol Chem 2003.
54. Wickstrom SA, Veikkola T, Rehn M, PihlajaniemiT, Alitalo K, Keski-Oja J. Endostatin-induced modu-lation of plasminogen activation with concomitantloss of focal adhesions and actin stress fibers incultured human endothelial cells. Cancer Res 2001;61:6511–6.
55. Ferreras M, Felbor U, Lenhard T, Olsen BR, Delaisse J.Generation and degradation of human endostatinproteins by various proteinases. FEBS Lett 2000;486:247–51.
56. Ackley BD, Crew JR, Elamaa H, Pihlajaniemi T, Kuo CJ,Kramer JM. The NC1/endostatin domain of Caenorhab-ditis elegans type XVIII collagen affects cell migrationand axon guidance. J Cell Biol 2001;152:1219–32.
57. Kuo CJ, LaMontagne KR Jr, Garcia-Cardena G, et al.Oligomerization-dependent regulation of motility andmorphogenesis by the collagen XVIII NC1/endostatindomain. J Cell Biol 2001;152:1233–46.
58. Kranenburg O, Bouma B, Kroon-Batenburg LM, et al.Tissue-type plasminogen activator is a multiligandcross-h structure receptor. Curr Biol 2002;12:1833–9.
59. Olsen BR. From the editor’s desk. Matrix Biol 2002;21:309–10.
60. Fukai N, Eklund L, Marneros AG, et al. Lack ofcollagen XVIII/endostatin results in eye abnormalities.EMBO J 2002;21:1535–44.
61. Ramchandran R, Dhanabal M, Volk R, et al.Antiangiogenic activity of restin, NC10 domain ofhuman collagen XV: comparison to endostatin. Bio-chem Biophys Res Commun 1999;255:735–9.
62. Sasaki T, Larsson H, Tisi D, Claesson-Welsh L,Hohenester E, Timpl R. Endostatins derived fromcollagens XV and XVIII differ in structural and bindingproperties, tissue distribution and anti-angiogenicactivity. J Mol Biol 2000;301:1179–90.
63. Ylikarppa R, Eklund L, Sormunen R, et al. Doubleknockout mice reveal a lack of major functionalcompensation between collagens XV and XVIII. MatrixBiol 2003;22:443–8.
64. Pasqualini R, Bourdoulous S, Koivunen E, Woods VLJr, Ruoslahti E. A polymeric form of fibronectin hasantimetastatic effects against multiple tumor types. NatMed 1996;2:1197–203.
65. Yi M, Ruoslahti E. A fibronectin fragment inhibitstumor growth, angiogenesis, and metastasis. Proc NatlAcad Sci U S A 2001;98:620–4.
66. Yi M, Sakai T, Fassler R, Ruoslahti E. Antiangiogenicproteins require plasma fibronectin or vitronectin forin vivo activity. Proc Natl Acad Sci U S A 2003;100:11435–8.
67. Kalluri R. Discovery of type IV collagen non-collagenous domains as novel integrin ligands andendogenous inhibitors of angiogenesis. Cold SpringHarb Symp Quant Biol 2002;67:255–66.
68. Albig AR, Schiemann WP. Fibulin-5 antagonizesvascular endothelial growth factor (VEGF) signalingand angiogenic sprouting by endothelial cells. DNA CellBiol 2004;23:367–79.
Cancer Research
Cancer Res 2005; 65: (10). May 15, 2005 3976 www.aacrjournals.org
Research. on June 28, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

69. Nguyen AD, Itoh S, Jeney V, et al. Fibulin-5 is a novelbinding protein for extracellular superoxide dismutase.Circ Res 2004;95:1067–74.
70. Lee MJ, Roy NK, Mogford JE, Schiemann WP,Mustoe TA. Fibulin-5 promotes wound healing in vivo .J Am Coll Surg 2004;199:403–10.
71. Good DJ, Polverini PJ, Rastinejad F, et al. A tumorsuppressor-dependent inhibitor of angiogenesis isimmunologically and functionally indistinguishablefrom a fragment of thrombospondin. Proc Natl AcadSci U S A 1990;87:6624–8.
72. Chen H, Herndon ME, Lawler J. The cell biology ofthrombospondin-1. Matrix Biol 2000;19:597–614.
73. Iruela-Arispe ML, Luque A, Lee N. Thrombospondinmodules and angiogenesis. Int J Biochem Cell Biol 2004;36:1070–8.
74. Agah A, Kyriakides TR, Lawler J, Bornstein P. Thelack of thrombospondin-1 (TSP1) dictates the course ofwound healing in double-TSP1/TSP2-null mice. Am JPathol 2002;161:831–9.
75. Streit M, Velasco P, Brown LF, et al. Overexpression ofthrombospondin-1 decreases angiogenesis and inhibitsthe growth of human cutaneous squamous cellcarcinomas. Am J Pathol 1999;155:441–52.
76. Rodriguez-Manzaneque JC, Lane TF, Ortega MA,Hynes RO, Lawler J, Iruela-Arispe ML. Thrombospon-din-1 suppresses spontaneous tumor growth andinhibits activation of matrix metalloproteinase-9 andmobilization of vascular endothelial growth factor. ProcNatl Acad Sci U S A 2001;98:12485–90.
77. Stellmach V, Volpert OV, Crawford SE, Lawler J,Hynes RO, Bouck N. Tumour suppressor genes andangiogenesis: the role of TP53 in fibroblasts. Eur JCancer 1996;32A:2394–400.
78. Zabrenetzky V, Harris CC, Steeg PS, Roberts DD.Expression of the extracellular matrix molecule throm-bospondin inversely correlates with malignant progres-sion in melanoma, lung and breast carcinoma cell lines.Int J Cancer 1994;59:191–5.
79. Lawler J, Miao WM, Duquette M, Bouck N, BronsonRT, Hynes RO. Thrombospondin-1 gene expressionaffects survival and tumor spectrum of p53-deficientmice. Am J Pathol 2001;159:1949–56.
80. Shafiee A, Penn JS, Krutzsch HC, Inman JK,Roberts DD, Blake DA. Inhibition of retinal angiogen-esis by peptides derived from thrombospondin-1 [inprocess citation]. Invest Ophthalmol Vis Sci 2000;41:2378–88.
81. Iruela-ArispeML, LombardoM, KrutzschHC, Lawler J,Roberts DD. Inhibition of angiogenesis by thrombo-spondin-1 is mediated by 2 independent regions withinthe type 1 repeats. Circulation 1999;100:1423–31.
82. Volpert OV, Zaichuk T, Zhou W, et al. Inducer-stimulated Fas targets activated endothelium fordestruction by anti-angiogenic thrombospondin-1 andpigment epithelium-derived factor. Nat Med 2002;8:349–57.
83. Streit M, Riccardi L, Velasco P, et al. Thrombospon-din-2: a potent endogenous inhibitor of tumor growthand angiogenesis. Proc Natl Acad Sci U S A 1999;96:14888–93.
84. Hawighorst T, Velasco P, Streit M, et al. Thrombo-spondin-2 plays a protective role in multistep carcino-genesis: a novel host anti-tumor defense mechanism.EMBO J 2001;20:2631–40.
85. Streit M, Stephen AE, Hawighorst T, et al. Systemicinhibition of tumor growth and angiogenesis bythrombospondin-2 using cell-based antiangiogenic genetherapy. Cancer Res 2002;62:2004–12.
86. Noh YH, Matsuda K, Hong YK, et al. An N-terminal 80kDa recombinant fragment of human thrombospondin-2 inhibits vascular endothelial growth factor inducedendothelial cell migration in vitro and tumor growthand angiogenesis in vivo . J Invest Dermatol 2003;121:1536–43.
87. Monboisse JC, Garnotel R, Bellon G, et al. The a3 chainof type IV collagen prevents activation of humanpolymorphonuclear leukocytes. J Biol Chem 1994;269:25475–82.
88. Han J, Ohno N, Pasco S, Monboisse JC, Borel JP,Kefalides NA. A cell binding domain from the a3 chainof type IV collagen inhibits proliferation of melanomacells. J Biol Chem 1997;272:20395–401.
89. Shahan TA, Ziaie Z, Pasco S, et al. Identification ofCD47/integrin-associated protein and a(v)h3 as tworeceptors for the a3(IV) chain of type IV collagen ontumor cells. Cancer Res 1999;59:4584–90.
90. Maeshima Y, Colorado PC, Torre A, et al. Distinctanti-tumor properties of a type IV collagen domainderived from basement membrane. J Biol Chem 2000;275:21340–8.
91. Maeshima Y, Colorado PC, Kalluri R. Two RGD-independent avh3 integrin binding sites on tumstatinregulate distinct anti-tumor properties. J Biol Chem2000;275:23745–50.
92. Pasco S, Ramont L, Maquart FX, Monboisse JC.Control of melanoma progression by various matrikinesfrom basement membrane macromolecules. Crit RevOncol Hematol 2004;49:221–33.
93. Floquet N, Pasco S, Ramont L, et al. The antitumorproperties of the a3(IV)-(185-203) peptide from the NC1domain of type IV collagen (tumstatin) are conforma-tion-dependent. J Biol Chem 2004;279:2091–100.
94. Imagawa W, Pedchenko VK, Helber J, Zhang H.Hormone/growth factor interactions mediating epithe-lial/stromal communication in mammary gland deve-lopment and carcinogenesis. J Steroid Biochem MolBiol 2002;80:213–30.
95. Maeshima Y, Manfredi M, Reimer C, et al. Identifi-cation of the anti-angiogenic site within vascularbasement membrane derived tumstatin. J Biol Chem2001;276:15240–8.
96. Maeshima Y, Yerramalla UL, Dhanabal M, et al.Extracellular matrix-derived peptide binds to a(v)h(3)integrin and inhibits angiogenesis. J Biol Chem 2001;276:31959–68.
97. Maeshima Y, Sudhakar A, Lively JC, et al. Tumstatin,an endothelial cell-specific inhibitor of protein synthe-sis. Science 2002;295:140–3.
98. Hutchings H, Ortega N, Plouet J. Extracellular matrix-bound vascular endothelial growth factor promotesendothelial cell adhesion, migration, and survivalthrough integrin ligation. FASEB J 2003;17:1520–2.
99. Hamano Y, Zeisberg M, Sugimoto H, et al. Physio-logical levels of tumstatin, a fragment of collagen IV a3chain, are generated by MMP-9 proteolysis andsuppress angiogenesis via aVh3 integrin. Cancer Cell2003;3:589–601.
100. Caudroy S, Cucherousset J, Lorenzato M, et al.Implication of tumstatin in tumor progression ofhuman bronchopulmonary carcinomas. Hum Pathol2004;35:1218–22.
101. McCawley LJ, Matrisian LM. Matrix metalloprotei-nases: they’re not just for matrix anymore! Curr OpinCell Biol 2001;13:534–40.
102. Yamamoto Y, Maeshima Y, Kitayama H, et al.Tumstatin peptide, an inhibitor of angiogenesis, pre-vents glomerular hypertrophy in the early stage ofdiabetic nephropathy. Diabetes 2004;53:1831–40.
103. Sang QX. Complex role of matrix metalloprotei-nases in angiogenesis. Cell Res 1998;8:171–7.
104. Patterson BC, Sang QA. Angiostatin-convertingenzyme activities of human matrilysin (MMP-7) andgelatinase B/type IV collagenase (MMP-9). J Biol Chem1997;272:28823–5.
105. Cornelius LA, Nehring LC, Harding E, et al. Matrixmetalloproteinases generate angiostatin: effects onneovascularization. J Immunol 1998;161:6845–52.
106. Zhang D, Kaufman PL, Gao G, Saunders RA, Ma JX.Intravitreal injection of plasminogen kringle 5, anendogenous angiogenic inhibitor, arrests retinal neo-vascularization in rats. Diabetologia 2001;44:757–65.
107. O’Reilly MS, Holmgren L, Shing Y, et al. Angiostatin:a novel angiogenesis inhibitor that mediates thesuppression of metastases by a Lewis lung carcinoma[see comments]. Cell 1994;79:315–28.
108. Lijnen HR, Ugwu F, Bini A, Collen D. Generation ofan angiostatin-like fragment from plasminogen bystromelysin-1 (MMP-3). Biochemistry 1998;37:4699–702.
109. Dong Z, Kumar R, Yang X, Fidler IJ. Macrophage-derived metalloelastase is responsible for the genera-tion of angiostatin in Lewis lung carcinoma. Cell 1997;88:801–10.
110. O’Reilly MS, Wiederschain D, Stetler-Stevenson WG,Folkman J, Moses MA. Regulation of angiostatinproduction by matrix metalloproteinase-2 in a model
of concomitant resistance. J Biol Chem 1999;274:29568–71.
111. Stathakis P, Fitzgerald M, Matthias LJ, ChestermanCN, Hogg PJ. Generation of angiostatin by reductionand proteolysis of plasmin. Catalysis by a plasminreductase secreted by cultured cells. J Biol Chem 1997;272:20641–5.
112. Ji WR, Castellino FJ, Chang Y, et al. Characterizationof kringle domains of angiostatin as antagonists ofendothelial cell migration, an important process inangiogenesis. FASEB J 1998;12:1731–8.
113. Geiger JH, Cnudde SE. What the structure ofangiostatin may tell us about its mechanism of action.J Thromb Haemost 2004;2:23–34.
114. Moser TL, Stack MS, Asplin I, et al. Angiostatinbinds ATP synthase on the surface of human endo-thelial cells. Proc Natl Acad Sci U S A 1999;96:2811–6.
115. Tarui T, Majumdar M, Miles LA, Ruf W, Takada Y.Plasmin-induced migration of endothelial cells. Apotential target for the anti-angiogenic action ofangiostatin. J Biol Chem 2002;277:33564–70.
116. Tarui T, Miles LA, Takada Y. Specific interaction ofangiostatin with integrin a(v)h(3) in endothelial cells.J Biol Chem 2001;276:39562–8.
117. Troyanovsky B, Levchenko T, Mansson G,Matvijenko O, Holmgren L. Angiomotin: an angiostatinbinding protein that regulates endothelial cell migra-tion and tube formation. J Cell Biol 2001;152:1247–54.
118. Claesson-Welsh L, Welsh M, Ito N, et al. Angiostatininduces endothelial cell apoptosis and activation offocal adhesion kinase independently of the integrin-binding motif RGD. Proc Natl Acad Sci U S A 1998;95:5579–83.
119. O’Reilly MS, Pirie-Shepherd S, Lane WS, Folkman J.Antiangiogenic activity of the cleaved conformation ofthe serpin antithrombin. Science 1999;285:1926–8.
120. Lee TH, Rhim T, Kim SS. Prothrombin kringle-2domain has a growth inhibitory activity against basicfibroblast growth factor-stimulated capillary endo-thelial cells. J Biol Chem 1998;273:28805–12.
121. Kusafuka K, Hiraki Y, Shukunami C, Kayano T,Takemura T. Cartilage-specific matrix protein, chon-dromodulin-I (ChM-I), is a strong angio-inhibitor inendochondral ossification of human neonatal vertebraltissues in vivo : relationship with angiogenic factors inthe cartilage. Acta Histochem 2002;104:167–75.
122. Shukunami C, Yamamoto S, Tanabe T, Hiraki Y.Generation of multiple transcripts from the chickenchondromodulin-I gene and their expression duringembryonic development. FEBS Lett 1999;456:165–70.
123. Hayami T, Shukunami C, Mitsui K, et al. Specific lossof chondromodulin-I gene expression in chondrosar-coma and the suppression of tumor angiogenesis andgrowth by its recombinant protein in vivo . FEBS Lett1999;458:436–40.
124. Kendall RL, Wang G, Thomas KA. Identificationof a natural soluble form of the vascular endothelialgrowth factor receptor, FLT-1, and its heterodimeri-zation with KDR. Biochem Biophys Res Commun1996;226:324–8.
125. Kendall RL, Thomas KA. Inhibition of vascularendothelial cell growth factor activity by an endo-genously encoded soluble receptor. Proc Natl Acad SciU S A 1993;90:10705–9.
126. Kondo K, Hiratsuka S, Subbalakshmi E, MatsushimeH, Shibuya M. Genomic organization of the flt-1 geneencoding for vascular endothelial growth factor (VEGF)receptor-1 suggests an intimate evolutionary relation-ship between the 7-Ig and the 5-Ig tyrosine kinasereceptors. Gene 1998;208:297–305.
127. He Y, Smith SK, Day KA, Clark DE, Licence DR,Charnock-Jones DS. Alternative splicing of vascularendothelial growth factor (VEGF)-R1 (FLT-1) pre-mRNAis important for the regulation of VEGF activity. MolEndocrinol 1999;13:537–45.
128. Sawano A, Takahashi T, Yamaguchi S, Aonuma M,Shibuya M. Flt-1 but not KDR/Flk-1 tyrosine kinase is areceptor for placenta growth factor, which is related tovascular endothelial growth factor. Cell Growth Differ1996;7:213–21.
129. Barleon B, Totzke F, Herzog C, et al. Mapping ofthe sites for ligand binding and receptor dimerizationat the extracellular domain of the vascular endothelial
Endogenous Angiogenesis Inhibitors
www.aacrjournals.org 3977 Cancer Res 2005; 65: (10). May 15, 2005
Research. on June 28, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

growth factor receptor FLT-1. J Biol Chem 1997;272:10382–8.
130. Belgore FM, Blann AD, Lip GY. sFlt-1, a potentialantagonist for exogenous VEGF. Circulation 2000;102:E108–9.
131. Yoshimura I, Mizuguchi Y, Miyajima A, Asano T,Tadakuma T, Hayakawa M. Suppression of lungmetastasis of renal cell carcinoma by the intramusculargene transfer of a soluble form of vascular endothelialgrowth factor receptor I. J Urol 2004;171:2467–70.
132. Ohtani K, Egashira K, Hiasa K, et al. Blockade ofvascular endothelial growth factor suppresses experi-mental restenosis after intraluminal injury by inhibitingrecruitment of monocyte lineage cells. Circulation 2004;110:2444–52.
133. Mitsuyasu RT. Interferon a in the treatment ofAIDS-related Kaposi’s sarcoma. Br J Haematol 1991;79:69–73.
134. LingenMW, Polverini PJ, BouckNP. Retinoic acid andinterferon a act synergistically as antiangiogenic andantitumor agents against human head and necksquamous cell carcinoma. Cancer Res 1998;58:5551–8.
135. Pepper MS, Vassalli JD, Wilks JW, Schweigerer L,Orci L, Montesano R. Modulation of bovine micro-vascular endothelial cell proteolytic properties byinhibitors of angiogenesis. J Cell Biochem 1994;55:419–34.
136. Ma Z, Qin H, Benveniste EN. Transcriptionalsuppression of matrix metalloproteinase-9 gene expres-sion by IFN-g and IFN-h: critical role of STAT-1a.J Immunol 2001;167:5150–9.
137. Dinney CP, Bielenberg DR, Perrotte P, Reich R,Eve BY, Bucana CD, Fidler IJ. Inhibition of basicfibroblast growth factor expression, angiogenesis, andgrowth of human bladder carcinoma in mice bysystemic interferon-a administration. Cancer Res1998;58:808–14.
138. Ribatti D, Crivellato E, Candussio L, et al. Angio-genic activity of rat mast cells in the chick embryochorioallantoic membrane is down-regulated by treat-ment with recombinant human a-2a interferon andpartly mediated by fibroblast growth factor-2. Haema-tologica 2002;87:465–71.
139. Lopez-Ocejo O, Perea SE, Bequet-Romero M, AranaMJ, Lopez Saura P. Impaired angiogenic balance andsuppression of tumorigenicity in HeLa cells chronicallyexposed to interferon-a. Biochem Biophys Res Com-mun 2000;277:410–6.
140. Nilsson A, Janson ET, Eriksson B, Larsson A. Levelsof angiogenic peptides in sera from patients withcarcinoid tumours during a-interferon treatment.Anticancer Res 2001;21:4087–90.
141. Sasamura H, Takahashi A, Miyao N, et al. Inhibitoryeffect on expression of angiogenic factors by antiangio-genic agents in renal cell carcinoma. Br J Cancer2002;86:768–73.
142. Wang L, Wu WZ, Sun HC, et al. Mechanism ofinterferon a on inhibition of metastasis and angio-genesis of hepatocellular carcinoma after curativeresection in nude mice. J Gastrointest Surg 2003;7:587–94.
143. Strieter RM, Polverini PJ, Kunkel SL, et al. Thefunctional role of the ELR motif in CXC chemokine-mediated angiogenesis. J Biol Chem 1995;270:27348–57.
144. Cozzolino F, Torcia M, Aldinucci D, et al. Interleukin1 is an autocrine regulator of human endothelial cellgrowth. Proc Natl Acad Sci U S A 1990;87:6487–91.
145. Bar D, Apte RN, Voronov E, Dinarello CA, Cohen S.A continuous delivery system of IL-1 receptor antago-nist reduces angiogenesis and inhibits tumor develop-ment. FASEB J 2004;18:161–3.
146. Volpert OV, Fong T, Koch AE, et al. Inhibitionof angiogenesis by interleukin 4. J Exp Med 1998;188:1039–46.
147. Voest EE, Kenyon BM, O’Reilly MS, Truitt G,D’Amato RJ, Folkman J. Inhibition of angiogenesisin vivo by interleukin 12. J Natl Cancer Inst 1995;87:581–6.
148. Sgadari C, Angiolillo AL, Tosato G. Inhibition ofangiogenesis by interleukin-12 is mediated by theinterferon-inducible protein 10. Blood 1996;87:3877–82.
149. Yao L, Sgadari C, Furuke K, Bloom ET, Teruya-Feldstein J, Tosato G. Contribution of natural killer cells
to inhibition of angiogenesis by interleukin-12. Blood1999;93:1612–21.
150. Yao L, Pike SE, Setsuda J, et al. Effective targetingof tumor vasculature by the angiogenesis inhibitorsvasostatin and interleukin-12. Blood 2000;96:1900–5.
151. Johansson M, Henriksson R, Bergenheim AT,Koskinen LO. Interleukin-2 and histamine in combina-tion inhibit tumour growth and angiogenesis inmalignant glioma. Br J Cancer 2000;83:826–32.
152. Morini M, Albini A, Lorusso G, et al. Prevention ofangiogenesis by naked DNA IL-12 gene transfer:angioprevention by immunogene therapy. Gene Ther2004;11:284–91.
153. Cao R, Farnebo J, Kurimoto M, Cao Y. Interleukin-18acts as an angiogenesis and tumor suppressor. FASEBJ 1999;13:2195–202.
154. D’Amato RJ, Lin CM, Flynn E, Folkman J, Hamel E.2-Methoxyestradiol, an endogenous mammalian me-tabolite, inhibits tubulin polymerization by interactingat the colchicine site. Proc Natl Acad Sci U S A 1994;91:3964–8.
155. Yue TL, Wang X, Louden CS, et al. 2-Methoxyestra-diol, an endogenous estrogen metabolite, inducesapoptosis in endothelial cells and inhibits angiogenesis:possible role for stress-activated protein kinase signal-ing pathway and Fas expression. Mol Pharmacol1997;51:951–62.
156. Bu S, Blaukat A, Fu X, Heldin NE, Landstrom M.Mechanisms for 2-methoxyestradiol-induced apoptosisof prostate cancer cells. FEBS Lett 2002;531:141–51.
157. Li L, Bu S, Backstrom T, Landstrom M, Ulmsten U,Fu X. Induction of apoptosis and G2-M arrest by2-methoxyestradiol in human cervical cancerHeLaS3 cells. Anticancer Res 2004;24:873–80.
158. Pribluda VS, Gubish ER Jr, Lavallee TM, Treston A,Swartz GM, Green SJ. 2-Methoxyestradiol: an endoge-nous antiangiogenic and antiproliferative drug candi-date. Cancer Metastasis Rev 2000;19:173–9.
159. Hileman EO, Liu J, Albitar M, Keating MJ, Huang P.Intrinsic oxidative stress in cancer cells: a biochemicalbasis for therapeutic selectivity. Cancer ChemotherPharmacol 2004;53:209–19.
160. Huang CY, Fujimura M, Noshita N, Chang YY, ChanPH. SOD1 down-regulates NF-nB and c-Myc expressionin mice after transient focal cerebral ischemia. J CerebBlood Flow Metab 2001;21:163–73.
161. Cushman M, He HM, Katzenellenbogen JA, Lin CM,Hamel E. Synthesis, antitubulin and antimitotic activity,and cytotoxicity of analogs of 2methoxyestradiol, anendogenous mammalian metabolite of estradiol thatinhibits tubulin polymerization by binding to thecolchicine binding site. J Med Chem 1995;38:2041–9.
162. Mabjeesh NJ, Escuin D, LaVallee TM, et al. 2ME2inhibits tumor growth and angiogenesis by disruptingmicrotubules and dysregulating HIF. Cancer Cell2003;3:363–75.
163. Duh EJ, Yang HS, Haller JA, et al. Vitreous levels ofpigment epithelium-derived factor and vascular endo-thelial growth factor: implications for ocular angio-genesis. Am J Ophthalmol 2004;137:668–74.
164. Yamagishi S, Inagaki Y, Amano S, Okamoto T,Takeuchi M, Makita Z. Pigment epithelium-derivedfactor protects cultured retinal pericytes from advancedglycation end product-induced injury through itsantioxidative properties. Biochem Biophys ResCommun 2002;296:877–82.
165. Ogata N, Tombran-Tink J, Nishikawa M, et al.Pigment epithelium-derived factor in the vitreous islow in diabetic retinopathy and high in rhegmatogenousretinal detachment. Am J Ophthalmol 2001;132:378–82.
166. Ogata N, Nishikawa M, Nishimura T, Mitsuma Y,Matsumura M. Unbalanced vitreous levels of pigmentepithelium-derived factor and vascular endothelialgrowth factor in diabetic retinopathy. Am J Ophthalmol2002;134:348–53.
167. Bouck N. PEDF: anti-angiogenic guardian of ocularfunction. Trends Mol Med 2002;8:330–4.
168. Gettins PG, Simonovic M, Volz K. Pigmentepithelium-derived factor (PEDF), a serpin with potentanti-angiogenic and neurite outgrowth-promoting prop-erties. Biol Chem 2002;383:1677–82.
169. Coljee VW, Rotenberg MO, Tresini M, Francis MK,Cristofalo VJ, Sell C. Regulation of EPC-1/PEDF in
normal human fibroblasts is posttranscriptional. J CellBiochem 2000;79:442–52.
170. Abe R, Shimizu T, Yamagishi S, et al. Over-expression of pigment epithelium-derived factordecreases angiogenesis and inhibits the growth ofhuman malignant melanoma cells in vivo . Am J Pathol2004;164:1225–32.
171. Abramson LP, Stellmach V, Doll JA, Cornwell M,Arensman RM, Crawford SE. Wilms’ tumor growth issuppressed by antiangiogenic pigment epithelium-derived factor in a xenograft model. J Pediatr Surg2003;38:336–42; discussion 336–42.
172. Crawford SE, Stellmach V, Ranalli M, et al. Pigmentepithelium-derived factor (PEDF) in neuroblastoma: amultifunctional mediator of Schwann cell antitumoractivity. J Cell Sci 2001;114:4421–8.
173. Guan M, Pang CP, Yam HF, Cheung KF, Liu WW,Lu Y. Inhibition of glioma invasion by overexpression ofpigment epithelium-derived factor. Cancer Gene Ther2004.
174. Doll JA, Stellmach VM, Bouck NP, et al. Pigmentepithelium-derived factor regulates the vasculatureand mass of the prostate and pancreas. Nat Med2003;9:774–80.
175. Wang L, Schmitz V, Perez-Mediavilla A, Izal I, Prieto J,Qian C. Suppression of angiogenesis and tumor growthby adenoviral-mediated gene transfer of pigmentepithelium-derived factor. Mol Ther 2003;8:72–9.
176. Mahtabifard A, Merritt RE, Yamada RE, Crystal RG,Korst RJ. In vivo gene transfer of pigment epithelium-derived factor inhibits tumor growth in syngeneicmurine models of thoracic malignancies. J ThoracCardiovasc Surg 2003;126:28–38.
177. Auricchio A, Behling KC, Maguire AM, et al.Inhibition of retinal neovascularization by intraocularviral-mediated delivery of anti-angiogenic agents. MolTher 2002;6:490–4.
178. Ohno-Matsui K, Morita I, Tombran-Tink J, et al.Novel mechanism for age-related macular degeneration:an equilibrium shift between the angiogenesis factorsVEGF and PEDF. J Cell Physiol 2001;189:323–33.
179. Ohno-Matsui K, Yoshida T, Uetama T, Mochizuki M,Morita I. Vascular endothelial growth factor upregulatespigment epithelium-derived factor expression viaVEGFR-1 in human retinal pigment epithelial cells.Biochem Biophys Res Commun 2003;303:962–7.
180. Tombran-Tink J, Barnstable CJ. Osteoblasts andosteoclasts express PEDF, VEGF-A isoforms, and VEGFreceptors: possible mediators of angiogenesis andmatrix remodeling in the bone. Biochem Biophys ResCommun 2004;316:573–9.
181. Brooks PC, Silletti S, von Schalscha TL,Friedlander M, Cheresh DA. Disruption of angiogenesisby PEX, a noncatalytic metalloproteinase fragment withintegrin binding activity. Cell 1998;92:391–400.
182. Pfeifer A, Kessler T, Silletti S, Cheresh DA,Verma IM. Suppression of angiogenesis by lentiviraldelivery of PEX, a noncatalytic fragment of matrixmetalloproteinase 2. Proc Natl Acad Sci U S A 2000;97:12227–32.
183. Maione TE, Gray GS, Petro J, et al. Inhibition ofangiogenesis by recombinant human platelet factor-4and related peptides. Science 1990;247:77–9.
184. Hagedorn M, Zilberberg L, Wilting J, et al. Domainswapping in a COOH-terminal fragment of plateletfactor 4 generates potent angiogenesis inhibitors.Cancer Res 2002;62:6884–90.
185. Chadderton NS, Stringer SE. Interaction of plateletfactor 4 with fibroblast growth factor 2 is stabilisedby heparan sulphate. Int J Biochem Cell Biol 2003;35:1052–5.
186. Ferrara N, Clapp C, Weiner R. The 16K fragment ofprolactin specifically inhibits basal or fibroblast growthfactor stimulated growth of capillary endothelial cells.Endocrinology 1991;129:896–900.
187. Pan H, Nguyen NQ, Yoshida H, et al. Moleculartargeting of antiangiogenic factor 16K hPRL inhibitsoxygen-induced retinopathy in mice. Invest OphthalmolVis Sci 2004;45:2413–9.
188. Corbacho AM, Martinez De La Escalera G, Clapp C.Roles of prolactin and related members of theprolactin/growth hormone/placental lactogen familyin angiogenesis. J Endocrinol 2002;173:219–38.
Cancer Research
Cancer Res 2005; 65: (10). May 15, 2005 3978 www.aacrjournals.org
Research. on June 28, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

189. Struman I, Bentzien F, Lee H, et al. Opposingactions of intact and N-terminal fragments of thehuman prolactin/growth hormone family memberson angiogenesis: an efficient mechanism for theregulation of angiogenesis. Proc Natl Acad Sci U S A1999;96:1246–51.
190. D’Angelo G, Struman I, Martial J, Weiner RI.Activation of mitogen-activated protein kinases byvascular endothelial growth factor and basic fibroblastgrowth factor in capillary endothelial cells is inhibitedby the antiangiogenic factor 16-kDa N-terminal frag-ment of prolactin. Proc Natl Acad Sci U S A 1995;92:6374–8.
191. D’Angelo G, Martini JF, Iiri T, Fantl WJ, Martial J,Weiner RI. 16K human prolactin inhibits vascularendothelial growth factor-induced activation of Rasin capillary endothelial cells. Mol Endocrinol 1999;13:692–704.
192. Baker EA, Stephenson TJ, Reed MW, Brown NJ.Expression of proteinases and inhibitors in humanbreast cancer progression and survival. Mol Pathol2002;55:300–4.
193. Brew K, Dinakarpandian D, Nagase H. Tissueinhibitors of metalloproteinases: evolution, structureand function. Biochim Biophys Acta 2000;1477:267–83.
194. Jiang Y, Goldberg ID, Shi YE. Complex roles of tissueinhibitors of metalloproteinases in cancer. Oncogene2002;21:2245–52.
195. Seo DW, Li H, Guedez L, et al. TIMP-2 mediatedinhibition of angiogenesis: an MMP-independent mech-anism. Cell 2003;114:171–80.
196. Moses MA, Wiederschain D, Wu I, et al. Troponin Iis present inhuman cartilage and inhibits angiogenesis.Proc Natl Acad Sci U S A 1999;96:2645–50.
197. Feldman L, Rouleau C. Troponin I inhibits capillaryendothelial cell proliferation by interaction with thecell’s bFGF receptor. Microvasc Res 2002;63:41–9.
198. Pike SE, Yao L, Setsuda J, et al. Calreticulin andcalreticulin fragments are endothelial cell inhibitorsthat suppress tumor growth. Blood 1999;94:2461–8.
199. Pike SE, Yao L, Jones KD, et al. Vasostatin, acalreticulin fragment, inhibits angiogenesis and sup-presses tumor growth. J Exp Med 1998;188:2349–56.
200. Yao L, Pike SE, Pittaluga S, et al. Anti-tumor activitiesof the angiogenesis inhibitors interferon-inducibleprotein-10 and the calreticulin fragment vasostatin.Cancer Immunol Immunother 2002;51:358–66.
201. Yao L, Pike SE, Tosato G. Laminin binding to thecalreticulin fragment vasostatin regulates endothelialcell function. J Leukoc Biol 2002;71:47–53.
202. Xiao F, Wei Y, Yang L, et al. A gene therapy forcancer based on the angiogenesis inhibitor, vasostatin.Gene Ther 2002;9:1207–13.
203. Ryan DP, Penson RT, Ahmed S, Chabner BA, LynchTJ Jr. Reality testing in cancer treatment: the phase Itrial of endostatin. Oncologist 1999;4:501–8.
204. Thomas JP, Arzoomanian RZ, Alberti D, et al. PhaseI pharmacokinetic and pharmacodynamic study ofrecombinant human endostatin in patients withadvanced solid tumors. J Clin Oncol 2003;21:223–31.
205. Davis DW, Shen Y, Mullani NA, et al. Quantitativeanalysis of biomarkers defines an optimal biologicaldose for recombinant human endostatin in primaryhuman tumors. Clin Cancer Res 2004;10:33–42.
206. Bukowski RM. AE-941, a multifunctional antiangio-genic compound: trials in renal cell carcinoma. ExpertOpin Investig Drugs 2003;12:1403–11.
207. Chamberlain J. Transforming growth factor-h: apromising target for anti-stenosis therapy. CardiovascDrug Rev 2001;19:329–44.
208. Westphal JR. Technology evaluation: ABT-510,Abbott. Curr Opin Mol Ther 2004;6:451–7.
209. Lakhani NJ, Sarkar MA, Venitz J, Figg WD. 2-Methoxyestradiol, a promising anticancer agent.Pharmacotherapy 2003;23:165–72.
Endogenous Angiogenesis Inhibitors
www.aacrjournals.org 3979 Cancer Res 2005; 65: (10). May 15, 2005
Research. on June 28, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

2005;65:3967-3979. Cancer Res Pia Nyberg, Liang Xie and Raghu Kalluri Endogenous Inhibitors of Angiogenesis
Updated version
http://cancerres.aacrjournals.org/content/65/10/3967
Access the most recent version of this article at:
Cited articles
http://cancerres.aacrjournals.org/content/65/10/3967.full#ref-list-1
This article cites 200 articles, 91 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/65/10/3967.full#related-urls
This article has been cited by 53 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://cancerres.aacrjournals.org/content/65/10/3967To request permission to re-use all or part of this article, use this link
Research. on June 28, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from