Embed Size (px)
Citation preview

Endocrine Disrupting Chemicals and theDevelopmental Programming of Adipogenesisand Obesity
Amanda Janesick and Bruce Blumberg*
Obesity and related disorders are a burgeoning public health epidemic,particularly in the U.S. Currently 34% of the U.S. population is clinicallyobese (BMI [ 30) and 68% are overweight (BMI [ 25), more than dou-ble the worldwide average and 10-fold higher than Japan and SouthKorea. Obesity occurs when energy intake exceeds energy expenditure;however, individuals vary widely in their propensity to gain weight andaccrue fat mass, even at identical levels of excess caloric input. Clinical,epidemiological, and biological studies show that obesity is largely pro-grammed during early life, including the intrauterine period. The envi-ronmental obesogen hypothesis holds that prenatal or early life expo-sure to certain endocrine disrupting chemicals can predispose exposedindividuals to increased fat mass and obesity. Obesogen exposure can al-ter the epigenome of multipotent stromal stem cells, biasing them towardthe adipocyte lineage at the expense of bone. Hence, humans exposed toobesogens during early life might have an altered stem cell compartment,which is preprogrammed toward an adipogenic fate. This results in ahigher steady state number of adipocytes and potentially a lifelong strug-gle to maintain a healthy weight, which can be exacerbated by societalinfluences that promote poor diet and inadequate exercise. This reviewfocuses on the developmental origins of the adipocyte, the relationshipbetween adipocyte number and obesity, and how obesogenic chemicalsmay interfere with the highly efficient homeostatic mechanisms regulat-ing adipocyte number and energy balance. Birth Defects Research(Part C) 93:34–50, 2011. VC 2011 Wiley-Liss, Inc.
Key words: obesogen; endocrine disrupter; EDC; PPARc; nuclear re-ceptor; adipogenesis; obesity; stem cells; multipotent stromal cells;MSCs; epigenetics; transgenerational
INTRODUCTIONIn the 1940s, the American psy-chologist William Sheldon and col-leagues proposed that people couldbe grouped into three differentsomatotypes (Fig. 1), named afterthe three germ layers (Sheldon,1940; Sheldon et al., 1954). The‘‘mesomorph’’ and ‘‘endomorph’’had proportionally large muscle
and fat mass, respectively,whereas the ‘‘ectomorph’’ had nei-ther but allegedly had a moredeveloped nervous system. Thefurther assignment of psychologi-cal characteristics to these soma-totypes had about as much reli-ability as attributing personalitytraits to the signs of the zodiac.Nevertheless, the popularity of the
somatotypes has remainedbecause, like some stereotypes,there is a kernel of truth. This isthat the somatotype designationsrecognize that a person’s propen-sity toward building muscle orstoring fat is largely predeter-mined before birth. Investigationsinto how muscles and fat growrevealed that certain changes inbody composition are only associ-ated with early development andrarely persist into adulthood. Forexample, the number of skeletalmuscle fibers was found to bechiefly established during the pre-natal period and remained fixedthroughout life in mice, cattle,pigs, and chicken (Luff and Gold-spink, 1967; Rehfeldt et al., 1987;Wegner et al., 2000), whereas thediameter and length of fibers couldbe increased with exercise(reviewed in Pearson, 1990). Simi-larly, adipocyte numbers increasedduring early development thenreached a plateau in adulthood,after which adipose tissue growthbecame primarily hypertrophic innature (Hirsch and Knittle, 1970;Knittle, 1972; Salans et al., 1973;Hager et al., 1978; Knittle et al.,1979). In both muscle and fattissue, the number of cellsappeared to be largely pre-determined at birth, which partlyexplains the apparent implacabilityof somatotypes.
REVIE
W
VC 2011 Wiley-Liss, Inc.
Birth Defects Research (Part C) 93:34–50 (2011)
Amanda Janesick and Bruce Blumberg are from Department of Developmental and Cell Biology, University of California, Irvine,California 92697-2300Bruce Blumberg is from Department of Pharmaceutical Sciences, University of California, Irvine, California 92697-2300
Supported by a grant from the NIH R01 ES015849.*Correspondence to: Prof. Bruce Blumberg, Department of Developmental and Cell Biology, University of California, Irvine, CA92697-2300. E-mail: [email protected]
View this article online at wileyonlinelibrary. DOI: 10.1002/bdrc.20197

Not surprisingly, such revela-tions about muscle growth engen-dered intense research in agri-cultural science aimed towarddeveloping particular properties ofmuscle fibers in livestock thatimproved the amount, texture,taste, and color of meat (Klontet al., 1998). Researchers in adi-pose biology still maintained ahigh level of interest in the factorsthat increase proliferation and dif-ferentiation of fat cells; however,this was seemingly uncoupledfrom the human predisposition tocertain somatotypes. Concurrentwith this shift, obesity researchbecame fairly detached from theadipocyte itself, focusing more onbroader subjects of human behav-ior, basal metabolic rate (as meas-ured by heat production and energyconsumption) and environment(nutrition and exercise). In the1990s, adipose tissue was recon-nected to human physiology,largely instigated by the discoveryof leptin (Zhang et al., 1994) and
the peroxisome proliferator-acti-vated receptor gamma (PPARc)(Tontonoz et al., 1994). It becamerecognized that adipose tissue isan endocrine/paracrine organ anda central regulator of metabolism(reviewed in Mohamed-Ali et al.,1998; Kershaw and Flier, 2004).Leptin is a protein hormone that isproduced in adipocytes and acts inthe ventromedial hypothalamus toregulate energy balance (reviewedin Ahima and Flier, 2000). Gener-ally speaking, each individual has a‘‘personal leptin threshold.’’ Whenleptin levels exceed this threshold,the brain senses energy sufficiencyand inhibits the fat storage pro-gram (reviewed in Lustig, 2006).Leptin decreases expression ofPPARc in adipocytes, whereas acti-vation of PPARc increases leptinexpression (reviewed in Rosenet al., 2000; Spiegelman and Flier,2001). Insulin increases both theexpression of PPARc and the gener-ation of endogenous PPARc ligandsin adipocytes, thereby increasing
triglyceride storage and adipocytesize (reviewed in Spiegelman andFlier, 2001). This new adipocyte-centric view fostered progress inthe understanding of signalingpathways underlying adipogenesis(Rosen and MacDougald, 2006;Kirchner et al., 2010) and how dys-regulation within the adipocyte leadsto metabolic disease (de Ferrantiand Mozaffarian, 2008; Vazquez-Vela et al., 2008).However, what remains missing
is an understanding of prenataland early life determinants of adi-pocyte number and morphology,as well as how these relate to met-abolic setpoints and the largelyunchangeable nature of body com-position. Adipose tissue is indeedan organ, albeit a distributed one(Wertheimer and Shapiro, 1948)and must be considered in thislight. Advances in stem cell biologyand epigenetics allow for a greaterunderstanding of how adipogenesisis regulated and may reveal themolecular basis underlying the
Figure 1. Depiction of the three classical somatotypes (from left to right): ectomorph, mesomorph, and endomorph.
ENDOCRINE DISRUPTORS AND OBESITY 35
Birth Defects Research (Part C) 93:34–50, (2011)

apparent human somatotypes inso-far as they exist. In this review, wediscuss current concepts about pre-natal and early life programming ofadipose tissue size and composi-tion, why adipose tissue tends to beresistant to persistent reductions inmass, and how prenatal exposureto obesogenic endocrine disruptingcompounds can program the fetustoward an obese, ‘‘endomorphic’’phenotype.
THE OBESOGEN
HYPOTHESIS
In 2006, we put forth the‘‘obesogen hypothesis’’ (Grun andBlumberg, 2006), which proposedthe existence of endocrine disrupt-ing chemicals (EDCs) that couldinfluence adipogenesis and obesityand that these might be important,yet hitherto unsuspected players inthe obesity epidemic. ‘‘Obesogens’’are defined functionally as chemi-cals (natural, pharmaceutical, orxenobiotic) that promote obesityby increasing the number of fatcells or the storage of fat intoexisting fat cells. Obesogens canalso act on adipocytes indirectly bychanging the basal metabolic rate,by shifting energy balance to favorthe storage of calories, and byaltering hormonal control of appe-tite and satiety (reviewed in Grunand Blumberg, 2009a,b; Janesickand Blumberg, 2011, in press;Blumberg, in press). Although theobesogen hypothesis was initiallycontroversial, many obesogenicchemicals have been identified inrecent years, underscoring therelevance of this novel model.Estrogens such as diethylstilbes-terol and genistein (Newbold et al.,2009), organotins such as tributyl-tin (Grun et al., 2006), perfluor-ooctanoates (Hines et al., 2009),and bisphenol A (Rubin et al.,2001) are known to be obesogenicin animals. A variety of chemicalshave been shown to increase adi-pogenesis in preadipocyte cell linessuch as murine 3T3-L1 cells andin primary multipotent mesenchy-mal stem cells (Lehmann et al.,1995; Grun et al., 2006; Feigeet al., 2007; Yang et al., 2007;Saito et al., 2009; Kirchner et al.,
2010; Park et al., 2010; Sargiset al., 2010; Styner et al., 2010;Zhang et al., 2010).Recent human epidemiological
studies have linked the presenceof xenobiotic chemicals withincreased body mass in humans.For example, the presence ofmono-benzyl and mono-ethyl-hexylphthalate metabolities in urine isassociated with increased waist di-ameter in men (Stahlhut et al.,2007; Hatch et al., 2008). Thepresence of hexachlorobenzene inumbilical cord blood, independentof whether the mother is over-weight, is associated withincreased BMI (Smink et al.,2008). Diethylstilbesterol in cordblood was associated withincreased BMI in young children,and this effect was exacerbated bymaternal smoking (Verhulst et al.,2009). This result was also sup-ported by evidence that higherserum concentration of DDE inmothers was linked to increasedweight and BMI in adult femaleoffspring (Karmaus et al., 2009).In addition to these xenobiotic
chemicals, several classes of phar-maceuticals are associated withincreased body mass in humans.These include atypical anti-psy-chotic drugs such as olanzapine(Nemeroff, 1997), selective sero-tonin reuptake inhibitors (Fava,2000), tricyclic antidepressants(Berken et al., 1984), and thia-zolidinedione anti-diabetic drugs(Larsen et al., 2003; Rubenstrunket al., 2007). Although, it shouldbe noted that most clinical and ep-idemiological studies are necessar-ily only correlational, animal andcell culture models have informedthese studies by providing mecha-nisms through which chemicalsmight act to create the obesity inhumans. For example, thiazolidi-nediones and phthalates are knownto act through the nuclear receptorperoxisome proliferator activatedreceptor gamma (PPARc), thedominant regulator of adipogenesis(Evans et al., 2004; Tontonoz andSpiegelman, 2008). Althoughdirect obesogenic effects of chemi-cals have been observed in animalsand humans, the more interestingpossibility is that EDC exposure
can exert effects during the prena-tal period that predispose humansto obesity later in life.
OBESOGENS AND THE
DEVELOPMENTAL ORIGINS
OF HEALTH AND DISEASE
Current ideas concerning obeso-gen actions on the endocrine sys-tem and hypothalamic-pituitary-adrenal axis have been reviewedrecently (Grun and Blumberg,2009a,b) and will not be empha-sized here. Instead, we will focuson the effects of EDCs (Diamanti-Kandarakis et al., 2009) known orsuspected to alter the develop-mental programming of adipogen-esis and obesity. EDC exposurecan, in principle, predispose anexposed individual to obesitythrough a variety of mechanismsthat act during the specification,differentiation, and maintenanceof adipocytes. EDCs can alter geneexpression both by modulating de-velopmental signaling pathwaysdirectly and by promoting epige-netic changes that produce stablyinherited changes in gene expres-sion. Clinical, epidemiological, andexperimental studies show thatsome of the origins of obesity canbe traced back to the intrauterineperiod. The developing fetusresponds to suboptimal conditionsduring sensitive developmental win-dows by producing structural andfunctional changes in cells, tissuesand organ systems—the concept offetal programming, or more appro-priately, the developmental originsof health and disease (DOHaD)(Gluckman and Hanson, 2004). Epi-genetic changes programmed dur-ing times of developmental plastic-ity in adipocyte development areheritable manifestations of envi-ronmental exposures whetherthey are nutritional, physiological,or chemical. Epigenetic changesresulting from EDC exposure couldfoster the type of swift change inthe metabolic profile of a populationthat has been observed in theUnited States in recent years (Flegalet al., 2010). Moreover, once anindividual becomes obese, it is diffi-cult to lose weight and sustain
36 JANESICK AND BLUMBERG
Birth Defects Research (Part C) 93:34–50, (2011)

weight loss due to highly efficienthomeostatic mechanisms regulatingenergy balance (Butte et al., 2007;Muhlhausler and Smith, 2009).
ORIGIN AND
ESTABLISHMENT OF THE
ADIPOCYTE COMPARTMENT
Adipogenesis is a differentiationevent in the mesodermal lineagewherein multipotent stromal stemcells (MSCs, otherwise known asmesenchymal stem cells) or theirmore lineage-restricted derivativesgive rise to adipocytes (reviewed inRosen and MacDougald, 2006).Until recently, the identity of adipo-cyte progenitors has been elusive.Most investigations into adipogene-sis used cell lines (e.g., 3T3-L1,3T3-F442A), or primary cells thatwere already committed to the adi-pocyte fate and would not easily dif-ferentiate into any other lineage (likebone, cartilage, muscle, or evenbrown fat) (Cornelius et al., 1994;Timmons et al., 2007; Park et al.,2008; Seale et al., 2008). Therefore,the origins of the adipocyte compart-ment could only be studied from thepoint of lineage commitment towardthe mature adipocyte. An importantbreakthrough occurred when MSCswere recharacterized as pericytes,cells that surround vasculaturethroughout the body (Crisan et al.,2008). Those who study these cellsintently have demonstrated thatwhile MSC-pericytes have enormouscapacity for multipotency in vitro,they exhibit a much more limited tis-sue-specific capacity to differentiatein vivo, (Fischbach et al., 2004).MSC-pericytes are known to expressdifferent markers, depending ontheir tissue of origin (Rosenbaumet al., 2008); therefore, MSC-peri-cytes are probably not a single classof cells. This argument is supportedby the observation that pericytes canhave different developmental ori-gins. For example, neural crest cellsgive rise to pericytes in the brain,whereas mesodermal cells give riseto pericytes in the adipose, muscle,and cartilage (da Silva Meirelleset al., 2008).Tissue-specific, committed pro-
genitors occupy this perivascular
niche of any organ. Although it iswell established that adipocytesare derived from MSC-pericytes,until recently there was very littleknown about the process andintermediates through which MSCsbecome committed to the adipocytelineage. Friedman and coworkerspurified candidate progenitor cellsfrom the stromal-vascular fractionof white adipose tissue (WAT) andverified that they were bona fideadipocyte progenitors by showingthat they could generate an entireadipose depot in lipodystrophicmice (Rodeheffer et al., 2008). Inadipose tissue, this progenitor is aPPARc-expressing cell (Tang et al.,2008) displaying the following suiteof markers: lin2, CD291, CD341,Sca11, and CD241 (Rodehefferet al., 2008). In support of the vas-cular niche argument, these cellsare also positive for pericytemarkers (e.g., SMA, PDGFRb andNG2) (Tang et al., 2008). More-over, adipose tissue is highly vascu-larized, and anti-angiogenic agentsreduce adipose mass (Rupnicket al., 2002; Kahn, 2008). Armedwith new knowledge of the originsof the adipose compartment, wecan now understand how the devel-opment of adipose (from MSC-peri-cyte to the mature adipocyte) canbe dysregulated by EDCs.
ADIPOCYTE NUMBER IS
REFRACTORY TO CHANGE
ONCE ESTABLISHED
It was once generally believedthat the root cause of obesity wasa lack of will-power (Newburghand Johnston, 1930) that was ei-ther due to individual personalitydefects or perhaps a disruption ofthe hypothalamus that interferedwith satiety (Hetherington andRanson, 1942). In May 2009, JohnBirkbeck, a professor at MasseyUniversity in New Zealand, re-energized this notion that obeseindividuals are wholly responsiblefor their condition. Noting thatthere were no ‘‘fattys’’ in Nazi con-centration camps, he suggestedthat obesity is the singular resultof a positive energy balance thatany obese individual will lose
weight under coercion and thatgovernments are too merciful withthe obese populus (Johns andLeask, 2009). This logic is flawedon many levels, but mostlybecause it conflates capacity withreality and assumes that obesityonly results from disordered eatingand inactivity. Indeed, as Birkbeckargues, obese individuals can losean appreciable amount of weightgiven a forced, calorie restricteddiet; however, the metabolism ofobese people is fundamentallyderanged compared with normalindividuals.Jules Hirsch and coworkers first
characterized the ‘‘reduced obese’’state (Glucksman and Hirsch,1968; 1969; Glucksman et al.,1968) where a small group ofobese patients were given strictly600 kcal per day, for three months.On average, these individuals,achieved and maintained a �30%reduction in weight, but this wasnot a homeostatic state (Glucksmanand Hirsch, 1968; 1969; Glucks-man et al., 1968). They exhibiteddepression, lethargy, and a manic,obsessive craving for food, indica-tive of a starvation response. Con-versely, a study involving healthymale prisoners in a Vermont peni-tentiary demonstrated that gainingweight through overconsumption ofcalories could not ‘‘be undertakenas a secondary occupation’’ (Simsand Horton, 1968). Although gain-ing weight was difficult, almost allparticipants were able to return totheir former weight quite easily, af-ter the period of overfeeding wasover. These studies suggest thatwhile the potential for weight loss(or gain) always exists, it is notnecessarily the case that forcingobese patients to lose weight with astrict dietary regiment improvestheir quality of life. The implicationis that obese and normal individualsrespond differently to increased ordecreased caloric intake and thatforcing lean people to gain weightor obese people to drastically loseweight is leading people to struggleagainst their basic nature.A key finding in Hirsch’s studies
was the observation that weightloss was primarily due to a reduc-tion in fat cell size, not number. In
ENDOCRINE DISRUPTORS AND OBESITY 37
Birth Defects Research (Part C) 93:34–50, (2011)

the reduced obese patients, theamount of triglycerides per cellwas reduced about 25% whereasthe number of adipocytes didnot change (Stern et al., 1972).The ‘‘reduced obese’’ population’sweight loss was hypotrophic,whereas the prisoner’s weight gainwas hypertrophic in nature. Theseresults were confirmed by subse-quent studies showing that weightloss after bariatric surgery orcancer-related cachexia reducesadipocyte size but not number(Spalding et al., 2008; Dahlmanet al., 2010). Thus, adults are lim-ited mostly to changes they canmake in adipocyte volume, and itis this component of adipose massthat can be altered most easily bydisciplined diet and exercise. Incontrast, for the developing fetus,child, and adolescent, the numberand distribution of adipocytes isnot at all constant. Human babieshave about 15% body fat (mostlywhite adipose) at birth (Kuzawa,1998), which nearly doubles dur-ing early development (to about28%), then declines during earlychildhood (Hager et al., 1977).From about 8 years of age untilthe end of puberty, adipocytenumber increases as the result ofhyperplastic expansion (Spaldinget al., 2008). Obese children pos-sess more fat cells, which multiplyat a faster rate, than nonobese chil-dren (Knittle et al., 1979) and obe-sity, particularly early-onset obe-sity, is linked to hypercellularity ofadipose tissue (Hirsch and Knittle,1970; Knittle, 1972; Salans et al.,1973). Adolescent girls experienc-ing a relatively large increase inadipose cell number over a periodof 1.5 years were the most resist-ant to treatment (e.g., restrictivediet and exercise regime) (Hageret al., 1978).Hence, it is likely that obese indi-
viduals either acquire more adipo-cytes before reaching adulthood,which may be causal for theirobesity, or create more adipocytesand associated obesity as adultsthrough poor diet and exercise.Although diet may be the drivingforce for adipose hypertrophy, it isunlikely to cause the hypercellular-ity observed in obesity under all
but the most extraordinary levelsof dietary excess. Adipocytes arecapable of reaching a certain tri-glyceride limit (e.g., in the caseof a high fat diet) and spawningthe generation of more fat cellsthrough paracrine signaling, butthis only occurs when adipocytecapacity for lipid storage is com-pletely exhausted (Shillabeeret al., 1989; Lau et al., 1990; Mar-ques et al., 1998). In general,once early adulthood is reached,the total number of adipocytesbecomes largely stable in bothmales and females (Spalding et al.,2008). Using the same [14C]-label-ing method used to prove thatadults have the capacity to producenew brain cells, it was found thatadipocytes are continually under-going apoptosis and being replen-ished (Spalding et al., 2008). Thegeneration of new adipocytes wasgreater for the obese population,because they were replenishing analready-established, larger pool ofcells (Spalding et al., 2008). Whencomparing the turnover rate of adi-pocytes between normal individualsand those with early onset obesity,no significant difference wasobserved (Spalding et al., 2008).Taken together, increased adipo-cyte number is the primary featureshared among obese individuals,particularly those who developedthe disease early or during adoles-cence, compared with those ofnormal weight. This suggests thatobese individuals possess a pool ofMSCs that is intrinsically biasedtoward the fat cell lineage. Morefat cells need to be generatedfrom precursors to maintain asteady state level of adipocytesthat is higher than in nonobesepeople. We argue (below) thatsuch a bias in the stem cell com-partment could be regulated byepigenetic changes due to expo-sure to environmental cues experi-enced during critical developmen-tal windows.
DO ADIPOCYTES CRAVE TO
BE FILLED?
The observation that obese indi-viduals have more adipocytesleads naturally to the hypothesis
that obesity is a consequence ofmore adipocytes. Although it isunclear at present whether such acausal connection exists betweenadipocyte number and obesity, itis known that mice with hyper-plastic obesity become morbidlyobese (Kim et al., 2007), and thathumans whose diabetes is beingtreated with rosiglitazone (a drugthat activates PPARc) developmore adipocytes and gain weight(Shim et al., 2006). Obese sub-jects (in particular, those withearly onset obesity (Salans et al.,1973)), are also predisposed tohave more adipocytes. Althoughthe evidence is incomplete, thereappears to be a critical minimumof triglycerides that adipocytes areprogrammed (or ‘‘want’’) to main-tain. In normal weight individuals,there is an inverse relationshipbetween the number of fat cellswithin an adipose depot and thesize of each cell (Arner et al.,2010). Thus, a reduction in cellvolume can only be carried out toa point. If an obese individual pos-sesses both adipose hypertrophyand hyperplasia, he can comfort-ably consume fewer calories toreduce cell size somewhat; how-ever, to achieve further perma-nent weight reduction, the adipo-cyte population must be reduced.Since adipocyte number is refrac-tory to change (as stated above)and adipocytes are resistant to ap-optosis (Sorisky et al., 2000), theobese individual can reach a pointof physiological starvation whilestill being fat. The sensation ofstarvation results when smaller fatcells secrete less leptin (Van Har-melen et al., 1998), a hormonethat plays a role in informing thebrain about how much fat isstored, which in turn suppressesfood intake and upregulates thesympathetic nervous system.These data all converge on the
conclusion that a hypercellularadipose mass, in essence,‘‘craves’’ to be filled with a mini-mum amount of lipid, per cell. Thisargues in favor of therapeuticstrategies that aim to alleviateadipose hyperplasia. It has beensuggested that altering the ‘‘birth–death balance’’ of adipose tissue
38 JANESICK AND BLUMBERG
Birth Defects Research (Part C) 93:34–50, (2011)

might be an effective treatmentfor obesity (Spalding et al., 2008;Arner et al., 2010). Of course,encouraging the death of adipo-cytes or increasing the triglycerideload on existing adipocytes mightcounterproductively generate aninflammatory response (Gustafsonet al., 2009; Alkhouri et al.,2010). Nevertheless, an obeseindividual who has successfullyreduced adipocyte volume throughdiet and exercise but reaches abarrier due to his fat-craving,hypercellular adipose mass, mightbenefit from therapeutic strategiesthat target the mechanisms regu-lating adipocyte turnover (Arner,2010). Alternatively, from a pre-ventative standpoint, one mightattempt to understand early de-velopmental events that createdthe hypercellular adipose mass.For example, factors, which en-courage the commitment of MSCsto the adipocyte lineage or pro-mote differentiation of progenitorsinto mature adipocytes, could cre-ate such a hyperplastic adiposemass that an individual might bestuck with for life. We discuss(below) how EDCs have the poten-tial to disrupt both the commit-ment and differentiation phase ofadipogenesis to create hyperplas-tic obesity.
ENDOCRINE DISRUPTION
DURING THE COMMITMENT
PHASE OF ADIPOGENESIS
Mature adipocytes are generatedfrom multipotent stromal cells(MSCs) found in almost all fetaland adult tissues (da Silva Meir-elles et al., 2006). What remainslargely unknown is the transcrip-tional program that turns MSCsinto preadipocytes, a subset ofstem cells that retains somedegree of multipotency but whichtypically gives rise to adipocytes,in vivo. One important player inthe process of lineage commit-ment is bone morphogenic protein(BMP) signaling. BMP signaling iswell known for its importance inskeletal development and BMP4was also shown to divert the fibro-blastic cell line C3H10T1/2 from
an MSC-like state into preadipo-cytes (Tang et al., 2004). Thissuggests that BMP signaling mightbe important for commitment tothe preadipocyte lineage. In sup-port of this idea, it was recentlyshown that the zinc finger tran-scription factor, Zfp423, a down-stream target of BMP signaling,was upregulated in preadipocytecell lines with high adipogenicpotential compared with those(e.g., NIH 3T3) that had a low adi-pogenic potential (Gupta et al.,2010). Interestingly, Zfp423 loss-of-function severely impaired butdid not eliminate adipogenesis inmouse embryos, which suggeststhat Zfp423 is not absolutely indis-pensable (Gupta et al., 2010). Inaddition, although Zfp423 expres-sion was noted to increase in prea-dipocytes compared with precursorcells, its expression remainedunchanged during the preadipocyteto adipocyte transition. This furthersuggests that Zfp423 may beinvolved in the commitment ofMSCs to the adipocyte lineage butmight not be required for the main-tenance of the preadipocyte pheno-type (Gupta et al., 2010).Another key developmental
pathway involved in adipogenesisis the Wnt signaling pathway. Itwas shown some time ago thatpreadipocytes do not differentiateinto adipocytes in the presence ofnoncanonical Wnt signaling (Rosset al., 2000). The reciprocal rela-tionship between adipocyte andosteocyte commitment and differ-entiation is well documented andis thought to involve a shift in theflow of MSCs from osteoblastic toadipogenic lineages (Shockleyet al., 2007). The balance betweenthe osteogenic and adipogenic line-ages is thought to be mediated bythe presence of PPARc, and it wasshown that Wnt-5a expressioninhibited the expression of PPARc,thereby diverting MSCs toward theosteogenic lineage (reviewed inTakada et al., 2009). Thus, repres-sion of noncanonical Wnt-5a sig-naling together with active BMPsignaling is required for MSCs toproceed toward the adipogenic andaway from the osteogenic lineage(Bilkovski et al., 2010).
The confluence of several signal-ing pathways to allocate stem cellsbetween adipogenic and osteo-genic fates provides multiple pos-sibilities for disruption. However,there are only very few studiestesting how EDCs might influenceMSC fate. The pesticides chlorpyri-fos and carbofuran inhibited theability of MSCs to differentiate intobone (Hoogduijn et al., 2006).Although one might expect that adecrease in the allocation of MSCsto the bone lineage would result inan increase in adipogenic progeni-tors, this possibility was nottested. We found that prenataltreatment with the environmentalobesogen, tributyltin, or the phar-maceutical obesogen, rosiglita-zone, altered lineage allocation inMSCs. Pregnant dams were treatedwith a single dose of tributyltin orrosiglitazone and MSCs harvestedfrom WAT at 8 weeks of age. WAT-derived MSCs were enriched inpreadipocytes and also in cells pre-disposed toward the adipocyte line-age. MSCs differentiated into adi-pocytes about twice as frequentlyin culture and the frequency wasfurther increased by in vitro treat-ment with tributyltin or rosiglita-zone (Kirchner et al., 2010). Theability of these cells to differentiateinto bone was correspondinglyinhibited (Kirchner et al., 2010).MSCs harvested from the tributyl-tin or rosiglitazone-exposed pupswere preprogrammed to prefer theadipogenic fate. Remarkably,MSCs from pups exposed prena-tally to TBT or rosiglitazone andthen treated with a bone differen-tiating cocktail instead differenti-ated into adipocytes at high fre-quency (Kirchner et al., 2010).This observation is closely corre-
lated with clinical findings. Indeed,osteoporosis has been called ‘‘obe-sity of bone’’ to reflect the obser-vation that bone marrow fromosteoporotic women is often filledwith adipocytes (Verma et al.,2002; Rosen and Bouxsein, 2006).Moreover, MSCs cultured from post-menopausal women with osteopo-rosis or low bone density accumulatetwofold more lipid, and twofold lesstype I collagen (part of the boneextracellular matrix) compared with
ENDOCRINE DISRUPTORS AND OBESITY 39
Birth Defects Research (Part C) 93:34–50, (2011)

women with healthy bones (Rodri-guez et al., 2000). Considering theseobservations, it is perhaps not sur-prising that women taking thiazolidi-nedione anti-diabetes medications(such as rosiglitazone) that activatePPARc are at increased risk for bonefractures (Habib et al., 2010). Theseresults lead to the prediction thatobesogens that act through PPARc,such as tributyltin, will have a similareffect on osteoporosis.As noted above, much of the
most recent evidence supports themodel that adipocytes are regen-erated from an existing populationof MSCs that reside in the vascula-ture of adipose tissues (Rodehefferet al., 2008; Tang et al., 2008).As MSCs are known to be mobile,it is possible that they circulatebeyond adipose depots and/or canbe recruited to WAT from otherniches such as the bone marrow.The majority of studies on MSCmigration focus on their ability toescape from bone marrow, migratealong blood vessels toward aninjury site following chemotacticsignals then moving through theextracellular matrix to flood anarea of injury (Orlic et al., 2001;Hofstetter et al., 2002; Liu et al.,2009). It is not unreasonable tohypothesize that a similar mecha-nism might lead MSCs to migrateto adipose depots from bone mar-row on receipt of an appropriatesignal (perhaps adipokines). Arecent study tested this hypothe-sis by transplanting green fluores-cent protein labeled MSCs intoirradiated wild-type mice and ask-ing whether green fluorescing cellscould be found in adipose depots(Crossno et al., 2006). It wasfound that rosiglitazone or a highfat diet increased migration ofthese MSCs to omental or dorsalintrascapular fat depots (Crossnoet al., 2006). However, their inter-pretation was challenged by a sub-sequent study that claimed thebone marrow derived cells didnot differentiate into adipocytes,because the green fluorescing,lipid-containing cells were not uni-locular as is typically the case foradipocytes (Koh et al., 2007). Fur-ther research will be required toestablish the fate and nature of
the transplanted cells and thepotential effects of EDC exposureon this process. It is also currentlycontroversial whether MSCsderived from tissues outside theadipose vasculature can differenti-ate into adipocytes, in vivo. Someresearchers believe that adiposeprogenitors are exclusively local-ized in the bone marrow and adi-pose vasculature (Rodehefferet al., 2008; Tang et al., 2008),whereas others favor a modelwherein MSCs from many tissuescan give rise to adipocytes in situunder appropriate stimulation(Crisan et al., 2008). If the latterpossibility is true, whether or notMSCs can migrate from their pointof origin to generate adipocytes inremote parts of the body may be acompletely moot point. Overall, itremains to be demonstrated whereadipogenic MSCs are located in thebody, whether or not they migrateto points beyond their origin inresponse to dietary or other stimuliand to what extent obesogen expo-sure influences these processes.
ENDOCRINE DISRUPTION
DURING THE
DIFFERENTIATION PHASE
OF ADIPOGENESIS
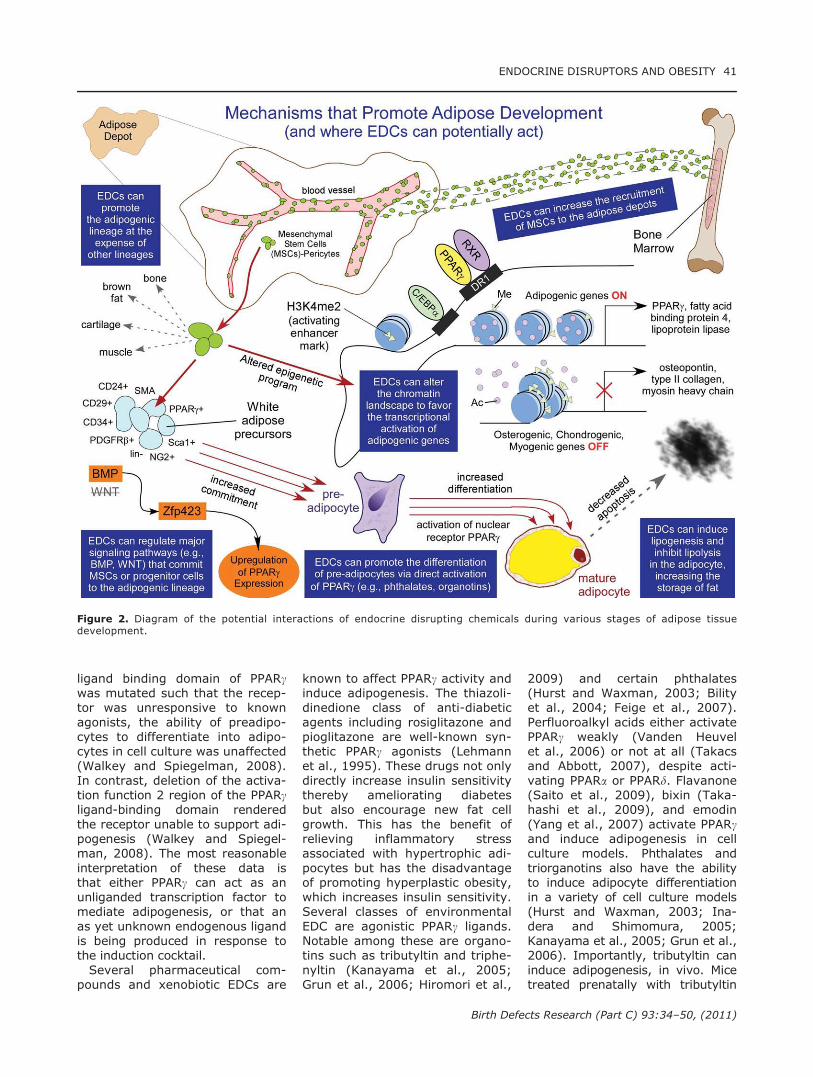
In contrast to the relative paucityof data regarding the commitmentof MSCs to become preadipocytes,there is much known about theprocess of adipocyte differentiation(reviewed in Rosen and MacDou-gald, 2006; Tontonoz and Spiegel-man, 2008). The central regulatorin this process is the peroxisomeproliferator activated receptorgamma (PPARc), which associateswith the retinoid X receptor (RXR)and binds DNA targets as a hetero-dimer to directly regulate theexpression of its target genes atthe transcriptional level (Tontonozet al., 1994). PPARc is consideredto be the master regulator of adi-pogenesis (reviewed in Evanset al., 2004) and plays key roles innearly all aspects of adipocyte biol-ogy (reviewed in Tontonoz andSpiegelman, 2008). Figure 2 sum-marizes the important events inadipocyte differentiation, focusing
on their origin from multipotentprecursors, how they commit to theadipocyte lineage instead of theother potential pathways down-stream of MSCs and how EDCsmight affect the generation, func-tion, or apoptosis of adipocytes.PPARc sits at the center of a
complex web of interacting signal-ing pathways that regulate itsexpression (see Fig. 3 in Tontonozand Spiegelman, 2008). In 3T3-L1cells, PPARc is first induced at thetranscriptional level by CCAAT/enhancer binding protein (C/EBP)b and d (Wu et al., 1996; Shaoand Lazar, 1997) and thenengages in a feed-forward loopwith C/EBPa, amplifying the adipo-genic signal (Rosen et al., 1999).To moderate the effects of thisfeed-forward loop, C/EBPa alsoinduces Sirt-1 (Jin et al., 2010),which curbs adipogenesis via inhi-bition of PPARc target genes(Picard et al., 2004). Studies ofadipogenesis in cell culture gener-ally require that MSCs, or preadi-pocytes be treated with a sensitiz-ing cocktail that stimulates the dif-ferentiation process (Studentet al., 1980), then with a PPARcligand that strongly promotes thedifferentiation process (Tontonozet al., 1994). Such cocktails oftencontain differentiation agents suchas insulin, glucocorticoids, methyl-isobutylxanthine, and indometha-cin, which act through the PI3K/AKT, glucocorticoid receptor, cAMPprotein kinase, and PPARc signal-ing pathways, respectively. Gener-ally speaking, the function of theinduction cocktail is to increasethe expression levels of C/EBPa orPPARc to levels where adipogene-sis is favored, rather than in thedirect ligand activation of PPARc.The insulin-induced transcriptionfactor, sterol regulatory elementbinding protein 1c (SREBP1c), canalso lead to the synthesis of fattyacids that can bind PPARc, and it isknown that high levels of insulinare sufficient to fully stimulate adi-pogenesis (Kim et al., 1998).It was recently proposed that
PPARc may function in adipogene-sis without requiring the ability tobe activated by ligand (Walkeyand Spiegelman, 2008). When the
40 JANESICK AND BLUMBERG
Birth Defects Research (Part C) 93:34–50, (2011)
ligand binding domain of PPARcwas mutated such that the recep-tor was unresponsive to knownagonists, the ability of preadipo-cytes to differentiate into adipo-cytes in cell culture was unaffected(Walkey and Spiegelman, 2008).In contrast, deletion of the activa-tion function 2 region of the PPARcligand-binding domain renderedthe receptor unable to support adi-pogenesis (Walkey and Spiegel-man, 2008). The most reasonableinterpretation of these data isthat either PPARc can act as anunliganded transcription factor tomediate adipogenesis, or that anas yet unknown endogenous ligandis being produced in response tothe induction cocktail.Several pharmaceutical com-
pounds and xenobiotic EDCs are
known to affect PPARc activity andinduce adipogenesis. The thiazoli-dinedione class of anti-diabeticagents including rosiglitazone andpioglitazone are well-known syn-thetic PPARc agonists (Lehmannet al., 1995). These drugs not onlydirectly increase insulin sensitivitythereby ameliorating diabetesbut also encourage new fat cellgrowth. This has the benefit ofrelieving inflammatory stressassociated with hypertrophic adi-pocytes but has the disadvantageof promoting hyperplastic obesity,which increases insulin sensitivity.Several classes of environmentalEDC are agonistic PPARc ligands.Notable among these are organo-tins such as tributyltin and triphe-nyltin (Kanayama et al., 2005;Grun et al., 2006; Hiromori et al.,
2009) and certain phthalates(Hurst and Waxman, 2003; Bilityet al., 2004; Feige et al., 2007).Perfluoroalkyl acids either activatePPARc weakly (Vanden Heuvelet al., 2006) or not at all (Takacsand Abbott, 2007), despite acti-vating PPARa or PPARd. Flavanone(Saito et al., 2009), bixin (Taka-hashi et al., 2009), and emodin(Yang et al., 2007) activate PPARcand induce adipogenesis in cellculture models. Phthalates andtriorganotins also have the abilityto induce adipocyte differentiationin a variety of cell culture models(Hurst and Waxman, 2003; Ina-dera and Shimomura, 2005;Kanayama et al., 2005; Grun et al.,2006). Importantly, tributyltin caninduce adipogenesis, in vivo. Micetreated prenatally with tributyltin
Figure 2. Diagram of the potential interactions of endocrine disrupting chemicals during various stages of adipose tissuedevelopment.
ENDOCRINE DISRUPTORS AND OBESITY 41
Birth Defects Research (Part C) 93:34–50, (2011)

are born with more fat stored thancontrols and Xenopus laevis tad-poles exposed to tributyltin exhib-ited replacement of testicular tissuewith adipocytes (Grun et al.,2006). It was inferred that tributyl-tin is likely to be promoting the de-velopment of new adipocytes at theexpense of other cell types.Other EDCs are known to pro-
mote adipogenesis but probably donot act through PPARc. Theseinclude bisphenol A (Rubin andSoto, 2009), nicotine (Somm et al.,2008), organophosphate pesticides(Slotkin, 2010), monosodium gluta-mate (Matsuyama et al., 1973;Bunyan et al., 1976), and polybro-minated diphenyl ethers (PBDEs)(Hoppe and Carey, 2007). Coplanarpolychlorinated biphenyls (PCBs;e.g., PCB-77) bind the aryl hydro-carbon receptor in adipocytes andincrease adipogenesis (Arsenescuet al., 2008). Bisphenol A andalkylphenols stimulate adipogene-sis in 3T3-L1 cells (Masuno et al.,2005), and bisphenol A diglycidylether was recently shown toinduce adipogenesis in humanand mouse bone marrow-derivedMSCs (Kirchner et al., 2011).Hence, EDCs can influence adi-
pocyte differentiation; however,the mechanisms are not alwaysclear. Because PPARc is such animportant regulator of adipogene-sis, it is common to associate thisreceptor with the obesogenic phe-nomenon observed. It should benoted, though, that numerousother nuclear receptors and theircofactors are important during adi-pogenesis (Feige et al., 2007; Fuet al., 2005; Rosen and MacDou-gald, 2006). These include positiveregulators of adipogenesis such asthe glucocorticoid receptor (GR)(Sargis et al., 2010), the liver ‘‘X’’receptor (LXR) (Seo et al., 2004),COUP-TFII (Li et al., 2009), estro-gen-related receptors alpha andgamma (ERRa, ERRc) (Kubo et al.,2009; Morganstein et al., 2010),Rev-ERB (Kumar et al., 2010;Wang and Lazar, 2008), and theNR4A family (Zhao and Bruemmer,2010). Negative regulators of adi-pogenesis include the thyroid hor-mone receptor (TR) (Lu and Cheng,2010), RORa (Ohoka et al., 2009),
Vitamin D (VDR) (Wood, 2008),DAX-1 (Kim and Surh, 2008), andthe nuclear receptor corepressorSMRT (Sutanto et al., 2010). Fur-thermore, the retinoid X receptor,RXR, is also upregulated duringadipogenesis and is activated bythe obesogens tributyltin andtriphenyltin (Grun et al., 2006; leMaire et al., 2009). RXR is an obli-gate heterodimeric partner forPPARc, as well as some of thereceptors noted above (TR, LXR,PPAR, VDR). RXR itself can be acti-vated in a subset of these hetero-dimers. In principle, any of thesereceptor-mediated pathways couldbe targeted by EDCs and this areais ripe for future studies.
ENDOCRINE DISRUPTOR
ACTION THAT ALTERS
METABOLISM OR LIPID
HOMEOSTASIS
Although several EDCs are associ-ated with adipogenesis and obe-sity in animal models, tributyltin isthe only EDC known to cause inutero effects on adipocytes viaactivation of PPARc (Kirchneret al., 2010; reviewed in Janesickand Blumberg, in press). Prenatalexposure to tributyltin in mice ledto a substantial increase in theamount of triglycerides in newborntissues that normally have little tono fat at all (Grun et al., 2006);although, the experiments did notdistinguish whether more lipid wasstored in existing cells, more cellswere produced, or both. OtherEDCs are likely to promote adipo-genesis, in utero, although it ispossible that this is secondary tobroader metabolic imbalances. Forexample, certain PCBs and PBDEsreduce thyroid function (Hallgrenet al., 2001) as does the antibac-terial compound triclosan (Paulet al., 2010; Rodriguez and San-chez, 2010). The mechanisms ofaction are not completely certain,but possible modes include inter-ference with thyroid hormone syn-thesis, transport, metabolism, orclearance (Diamanti-Kandarakiset al., 2009). High levels of mater-nal PCBs and PBDEs are correlatedwith reduced total and free T4
levels in infant cord blood(Herbstman et al., 2008) and thy-roid hormone stimulates lipolysisin adipocytes (Van Inwegen et al.,1975; Smith et al., 1991). Thyroidhormone also inhibits lipogenesisby downregulating expression ofSREBP1c (Viguerie et al., 2002).Taken together, these results leadto the inference that exposure toEDCs such as triclosan, PCBs, andPBDEs can reduce circulating thy-roid hormone levels leading to aconsequent increase in adipocytelipid accumulation.Estrogens are another class of
EDCs that can globally affectenergy metabolism (reviewed inChen et al., 2009). Estrogens areantiobesogenic in adults; they pro-tect against adiposity by promot-ing exergonic, energy-consuming,reactions in glycolysis, fatty acidoxidation, and electron transport(reviewed in Chen et al., 2009).However, perinatal exposure tolow doses of estrogens such asdiethylstilbestrol (DES) or genis-tein leads to obesity in mice (New-bold et al., 2009). Perfluoroaklylsulfonate exposure in mice led toincreased body weight in offspring,along with higher serum insulin andleptin levels (Hines et al., 2009).There is evidence both in support of(Sakurai et al., 2004; Hugo et al.,2008; Ben-Jonathan et al., 2009;Rubin and Soto, 2009) and against(Ryan et al., 2010) bisphenol A asan obesogen. Nicotine, which pro-motes weight loss in adults,increases adipose hypertrophy inyoung rats prenatally exposed tonicotine via osmotic pump (Sommet al., 2008). This has a parallel inhumans where epidemiologicalstudies of maternal smoking showthat the adjusted odds ratio forobesity is between 1.5–2.0 foldgreater if children were exposedduring, but not prior or after, thepregnancy (Power and Jefferis,2002; Oken et al., 2005; Al Mamunet al., 2006).Taken together, the studies pre-
sented above support the conten-tion that obesogens can act onadipocyte commitment, adipocytedifferentiation, by altering meta-bolic setpoints, modulating lipidhomeostasis, and by other mecha-
42 JANESICK AND BLUMBERG
Birth Defects Research (Part C) 93:34–50, (2011)

nisms not yet characterized. Wewill next consider the possiblemechanisms through which obeso-gen action during critical windowsof development might perma-nently alter the phenotype ofexposed individuals.
EPIGENETIC
MODIFICATIONS DURING
EARLY LIFE
One seemingly perplexing aspectof the burgeoning obesity epi-demic is the very rapid increase inthe rates of obesity, particularly inthe U.S. As recently as the 1980s,�15% of the population was obesecompared with 34% in 2008 (Flegalet al., 2010). Although genetic var-iability is sufficient to explain whysome people may have the propen-sity to become obese, it is incon-ceivable that the rapid increasein the rate of obesity in the U.S.has any genetic basis. In contrast,epigenetic phenomena can occurvery rapidly and could easilybecome established within a popu-lation within a single generation(Gluckman et al., 2005). There-fore, epigenetic changes mediatedby dietary and environmental fac-tors, rather than genetic changes,are a more plausible explanationfor the ‘‘epidemic’’ of obesity inWestern countries. Moreover, epi-genetics can also explain the typesof rapid metabolic adaptationsdescribed in the DOHaD studies(Barker, 1994; Hales and Barker,2001; Gluckman and Hanson,2004). It is likely that epigeneticchanges also underlie adaptationsmade during other critical develop-mental time windows, like adoles-cence, because the epigenomechanges substantially during life,even in monozygotic twins (Fragaet al., 2005).At the chromatin level, EDC ex-
posure can alter the expression ofproteins required for DNA accessi-bility or structure. Changing theexpression levels of DNA methyl-transferases, histone acetyltrans-ferases, histone deacetylases, andhistone methyltransferases willhave significant and sustainedeffects on gene expression. For
example, prenatal exposure toDES led to a significant increase inthe mRNA expression of DNAmethyltransferase 1 (Dnmt1) andDNA methyltransferase 3b(Dnmt3b). In turn, this led to hy-permethylation of the homeoboxgene HOXA10 and region-specificalterations in its expression levelsin the uteri of exposed animals(Bromer et al., 2009). Exposure tothe commonly used fungicide vin-clozolin caused a decrease in theexpression of Dnmt1, Dnmt3a,Dnmt3L, and euchromatic histonemethyltransferase (Ehmt1) in thetestes of male rats, and thesechanges were shown to persist forat least three subsequent genera-tions without further exposure(Anway et al., 2008). Whatremains an important question ishow EDCs can lead to epigeneticalterations on particular metabolicpathways to the exclusion ofothers. There are relatively fewDNA and histone methyl transfer-ases, compounding the problemeven further. One possibility isthat DNA methyl transferases suchas Dnmt1 and Dnmt3 are escortedby specific transcription factors tothe target DNA sequences thatthey, themselves recognize. Inthis case, target gene selectivityresides in the interaction betweentranscription factor and the chro-matin remodeling factor and thespecificity of the EDC for a particu-lar pathway would be conferred bythe transcriptional programs inindividual cells (Robertson et al.,2000; Burgers et al., 2002).Epigenetic changes related to
adipogenesis and obesity originatewithin the stem cell compartment.The observation that diet andexercise primarily alter adipocytevolume rather than adipocytenumber in adults probably resultsfrom the developmentally pro-grammed bias in the stem cellpopulation that causes the adipo-cyte pool to be replenished to its‘‘set point.’’ Understanding howadipocyte number is programmedat the genomic level will be of crit-ical importance in understandingthis set point phenomenon, andhow it is modified by EDCs, dietaryfactors, or the intrauterine envi-
ronment. At least one example ofEDC-induced changes in MSC fatehas already been identified. MSCsfrom mice exposed to tributyltin inutero exhibited alterations in themethylation status of the CpGislands of adipogenic genes suchas AP2 and PPARc. This led to anincrease in the number of preadi-pocytes at birth and an increasedpropensity of MSCs to differentiateinto adipocytes on adipogenicstimulation (Kirchner et al., 2010).It is likely that other such exam-ples will be identified in the future.The observation that EDCs, such
as tributyltin, can modify DNAmethylation to alter the expres-sion of adipogenic genes in MSCs(Kirchner et al., 2010) is only thebeginning of possible EDC-derivedepigenetic modifications within thestem cell compartment of adiposetissue. Recent studies using stemcells, including MSCs haverevealed the influence of histonemethylation on lineage program-ming. MSCs, such as embryonicstem cells (Azuara et al., 2006;Bernstein et al., 2006), naı̈ve Tcells (Wei et al., 2009), and neuralprogenitors (Mikkelsen et al.,2007), exhibit bivalent chromatinmarks on histone H3 proteins asso-ciated with promoters of lineagespecific genes (Noer et al., 2009).That is, both activating markssuch as tri-methylation of H3 atlysine 4 (H3K4me3) and repres-sive marks, such as H3K27me3,are present in the same histonemolecules (Azuara et al., 2006;Bernstein et al., 2006; Roh et al.,2006). These opposing histonemodifications are thought to‘‘prime’’ genes such that they canbe quiescent but ready to bequickly activated when differentia-tion is induced simply by demethy-lating H3K27 (Bernstein et al.,2006). A similar phenomenon hasbeen identified in the PPARc pro-moter in adipose-derived MSCs.Both marks are present in theMSCs, but when differentiation isstimulated, H3K27 is demethy-lated completely, leaving only theactivating mark on the PPARc pro-moter (Noer et al., 2009). It wouldbe readily possible for obesogenexposure to alter early program-
ENDOCRINE DISRUPTORS AND OBESITY 43
Birth Defects Research (Part C) 93:34–50, (2011)

ming events when these lineage-specific histone modifications areestablished. In addition, inappro-priate temporal or spatial stimula-tion of the demethylases thatremove the H3K27me3 marks,JMJD3 and UTX (Lan et al., 2007),could lead to inappropriate activa-tion of PPARc and the adipogenicprogram.An intriguing situation occurs
with genes such as PPARc, whichare important for multiple differ-entiation processes. For example,PPARc is expressed in monoblasts(Greene et al., 2000) and pro-motes macrophage differentiation(Tontonoz et al., 1998), but PPARcis also expressed in preadipocyteswhere it is required for the adipo-genic pathway (Tontonoz et al.,1994). Adipocytes and macro-phages diverge from a commonprogenitor early in development,but both share a requirement forPPARc expression. In order forPPARc to promote a cell-specifictranscriptional program, an addi-tional layer of regulation isrequired: differential recruitmentof PPARc to enhancer elements.Similar to the estrogen receptor,PPARc has an affinity for gene regu-latory regions in introns and distalto the promoter (Nielsen et al.,2008). The PPARc/RXR hetero-dimer binds to direct repeats sepa-rated by one nucleotide (DR1) withPPARc binding 50 to RXR (Jpenberget al., 1997; Chandra et al., 2008).Enhancer-specific histone methyla-tion increases the probability that apotential DR1 element is an actual,functional PPARc response element(Heintzman et al., 2009). Methyla-tion patterns on active enhancersare distinct from the promotermarks discussed above. HistoneH3 in active enhancers displaysmonomethylated and dimethylatedhistone, at lysine 4 (H3K4me1,H3K4me2) (Heintzman and Ren,2009; Heintzman et al., 2007).Macrophages have activatingH3K4me1 marks associated withthe enhancers of cytokine and im-munity genes. These are closelylinked to binding of PPARc and theets-factor PU1 (Heinz et al., 2010;Lefterova et al., 2010). In contrast,the same enhancers are repressed
in adipocytes, whereas those ofhighly induced genes are associ-ated with adjacent PPARc and C/EBPa binding (Siersbaek et al.,2010). An important, open ques-tion is whether the presence ofthese histone marks increases thelikelihood of PPARc binding to theDR1 consensus or if PPARc itselfdirectly recruits methyltransfer-ases to the DNA. In the latterinstance, obesogens such as tribu-tyltin could directly act to alter thechromatin leading to preferentialrecruitment of PPARc to theenhancers of its target genes inadipocyte progenitors.
ENDOCRINE DISRUPTION
ACROSS MULTIPLE
GENERATIONS
A profound and provocative possi-bility is that epigenetic changescaused by environmental expo-sures may be transmitted acrossgenerations. Although the dataare currently scant, there aresome indications that this mightbe the case. Vinclozolin exposurein the parental generation is linkedto infertility, behavior, and matepreference for at least four subse-quent generations (Anway et al.,2005). Many, but not all of theobserved alterations in chromatinremodeling were stably inheritedin subsequent generations (atleast through F4) despite that vin-clozolin exposure only occurred inthe parental (F0) animals (Anwayet al., 2008). This ‘‘memory’’ of anancestral exposure is thought tobe epigenetically maintainedwithin the male germ cells (Anwayand Skinner, 2006).Epidemiological studies using a
cohort of Swedish farmers fromthe O†verkalix region of Swedendemonstrated that food availabilityduring the prepubescent periodaffected the longevity and mortal-ity (from cardiovascular disease)of one’s grandchildren. Remark-ably, a single winter of overeatingcould lead to a 6-year decrease inlongevity of a boy’s grandsons,but not granddaughters, or a girl’sgranddaughters but not grandsons(Kaati et al., 2007). During the
prepubescent period, the testes orovaries are developing, and theprimordial germ cells incorporatesex-specific imprinting patterns,making this time exquisitely sensi-tive to epigenetic changes thatcan be transmitted down the gen-erations (Hajkova et al., 2002).Although the O†verkalix example
was a nutritional study, it serves toillustrate the possibility of environ-mental effects being epigeneticallytransmitted to one’s descendentsin humans. We propose that it is noless likely that exposure to EDCsduring sensitive developmentalwindows could lead to similartransgenerational effects. Thereare currently no data regardingpotential transgenerational effectsof obesogens; however, it is rea-sonable to propose that obesogeniccompounds that can directly altergene expression (e.g., tributyltin,BPA, or phthalates) could alsoinfluence the propensity to beobese for some generations afterthe initial exposure. Whether or notthe effects could be truly transge-nerational (i.e., persist in F3 andbeyond) compared with multige-nerational (persist to F2) wouldlikely depend on whether exposureoccurred during a critical develop-mental window (Jirtle and Skinner,2007).
CONCLUSIONS AND AREAS
OF FUTURE RESEARCH
For a long time, obesity researchhas focused on what we call the‘‘central dogma’’ of obesity—thatall humans possess sufficient ‘‘freewill’’ to manipulate their fat-ladensomatotypes relatively simply viaproper nutrition and physical activ-ity. Much has been written about thebehavioral origins of obesity and thefact that we live in a habitat thatpromotes a positive energy balance(Hill and Peters, 1998). Various gov-ernmental measures have beenundertaken to improve this environ-ment where fructose is more con-venient than carrots, and sedentarychoices champion over activitiesthat require exercise. San Franciscobanned toys from Happy Mealsunless they were adjusted to meetnutritional guidelines (Martinez,
44 JANESICK AND BLUMBERG
Birth Defects Research (Part C) 93:34–50, (2011)

2010). Vermont Attorney General,Bill Sorrell, proposed legislature totax soda pop (Kinzel, 2010). Yet,tap into any voice in the ‘‘fato-sphere’’ (Rabin, 2008) and onequickly realizes that obesity maynot solely be a matter of personalresponsibility and discipline, or eventhe dearth of healthy food optionsand exercise opportunities. Manyobese individuals passionatelybelieve that an underlying, drivingforce keeps them fat. This phenom-enon is commonly explained bygenetics (Walley et al., 2009), anobesogenic environment (Gorin andCrane, 2009), and the unremittingbiochemical and neurobiologicalforces that maintain the body in anobese state (Lustig, 2006; Fried-man, 2009; Rosenbaum and Leibel,2010).A newly recognized contributing
factor to the seemingly intractableproblem of obesity is exposure toEDCs, during the prenatal periodor early life. Although EDCs arehypothesized to interfere withbroad metabolic processes to en-courage adipogenesis, fat storage,and feeding (Grun and Blumberg,2009a; b), the strongest evidencefor obesogen action is at the levelof stem cell programming. Prenatalexposure to tributyltin alters theadipose vascular network of MSC-pericytes, pushing them down theadipocyte lineage (Kirchner et al.,2010). We showed that obesogensencourage MSC-pericytes to giverise to a higher proportion of com-mitted WAT precursors at theexpense of bone precursors, andthat tributyltin exposure gives riseto larger adipose depots than thoseof unexposed animals (Grun et al.,2006). Whether these changes inadipogenesis create an adiposemass that defies reduction in sizeremains to be determined. How-ever, there are data to suggestthat adipocytes ‘‘crave’’ to be filled(although this point remains con-troversial), and we propose thathaving a larger adipocyte progeni-tor population will result in alarger steady state adipocyte pop-ulation that may interfere with thesuccess of subsequent weight lossattempts.
An alarming recent trend is theincreasing rate of obesity in veryyoung children, even infants (Tav-eras et al., 2009; Koebnick et al.,2010; McCormick et al., 2010).Unless one wants to argue thatthe typical infant is now consum-ing far more calories than in thepast and refraining from exercisethat previous generationsembraced, the most reasonableconclusion is that the infant wasborn with more fat, and/or thatsomething about the early post-natal environment is vastly differ-ent than in the past. Remarkably,a recent study showed that ani-mals (pets, cats and dogs; labora-tory animals, rats, mice, and fourspecies of primates; and feralrats) living in proximity to humansin industrialized societies exhibitedpronounced increases in obesityover the past several decades (Kli-mentidis et al., 2010). Notably,these populations included labora-tory animals living in strictly con-trolled environments, as well asferal animals living in cities (Kli-mentidis et al., 2010). The likeli-hood of 24 animal populationsfrom eight different species allshowing a positive trend in weightover the past few decades bychance was estimated at 1.2 31027 (Klimentidis et al., 2010).These increases in weight over
time in humans, and in animalpopulations associated withhumans, argue for alternativeexplanations than simply diet andexercise; obesogens have a role toplay here. What remains unknownis the extent to which obesogensinfluence obesity in humans com-pared with other recognized fac-tors such as the timing, amountand nature of calories consumed,physical activity, other lifestylefactors such as stress, amount ofsleep, virus exposure, gutmicrobes, and genetic factors suchas single polynucleotide polymor-phisms in a variety of genes. Theobesogen hypothesis fits well withthe DOHaD model to provide mo-lecular explanations for how obe-sity might begin in the womb. Epi-genetics, a strong component ofDOHaD, is predicted to drive earlyprogramming events in the MSC-
pericyte compartment, where cellsreceive cues from their local envi-ronment that limit potential forfuture differentiation. Since criticalevents in the development of theadipocyte compartment occurearly in life, this is when obeso-genic chemicals likely act to alterepigenetic programming eventsthat predispose a stem or progeni-tor cell toward a particular lineage.Evidence to support an epigeneticbasis for obesogen action is onlynow emerging (Kirchner et al.,2010) as is evidence supportingepigenetic effects of EDC exposureon fertility, behavior, stress andother endpoints (Jirtle and Skinner,2007; Skinner and Guerrero-Bosa-gna, 2009). The field of adiposedevelopment, beginning at thestem cell stage, is still in itsinfancy, and future research shouldendeavor to understand whetherand how this process can be misre-gulated by EDCs in obesity.
ACKNOWLEDGMENTS
A.J. is a pre-doctoral trainee ofNSF IGERT DGE 0549479. We wouldalso like to thank James Janesick forartistic contributions to Figure 1.
REFERENCES
Ahima RS, Flier JS. 2000. Leptin. AnnuRev Physiol 62:413–437.
Al Mamun A, Lawlor DA, Alati R, et al.2006. Does maternal smoking duringpregnancy have a direct effect onfuture offspring obesity? Evidencefrom a prospective birth cohortstudy. Am J Epidemiol 164:317–325.
Alkhouri N, Gornicka A, Berk MP, et al.2010. Adipocyte apoptosis, a linkbetween obesity, insulin resistance,and hepatic steatosis. J Biol Chem285:3428–3438.
Anway MD, Cupp AS, Uzumcu M, Skin-ner MK. 2005. Epigenetic transge-nerational actions of endocrine dis-ruptors and male fertility. Science308:1466–1469.
Anway MD, Rekow SS, Skinner MK.2008. Transgenerational epigeneticprogramming of the embryonic testistranscriptome. Genomics 91:30–40.
Anway MD, Skinner MK. 2006. Epige-netic transgenerational actions of en-docrine disruptors. Endocrinology147(6 Suppl):S43–S49.
Arner E, Westermark PO, Spalding KL,et al. 2010. Adipocyte turnover: rele-vance to human adipose tissue mor-phology. Diabetes 59:105–109.
ENDOCRINE DISRUPTORS AND OBESITY 45
Birth Defects Research (Part C) 93:34–50, (2011)

Arsenescu V, Arsenescu RI, King V, et al.2008. Polychlorinated biphenyl-77induces adipocyte differentiation andproinflammatory adipokines and pro-motes obesity and atherosclerosis. En-viron Health Perspect 116:761–768.
Azuara V, Perry P, Sauer S, SpivakovM, Jorgensen HF, John RM, Gouti M,Casanova M, Warnes G, Merkenschl-ager M, Fisher AG. 2006. Chromatinsignatures of pluripotent cell lines.Nat Cell Biol 8:532–538.
Barker DJP. 1994. Programming thebaby. London: BMJ Publishing Group.
Ben-Jonathan N, Hugo ER, Brande-bourg TD. 2009. Effects of bisphenolA on adipokine release from humanadipose tissue: implications for themetabolic syndrome. Mol Cell Endo-crinol 304:49–54.
Berken GH, Weinstein DO, Stern WC.1984. Weight gain. A side-effect oftricyclic antidepressants. J Affect Dis-ord 7:133–138.
Bernstein BE, Mikkelsen TS, Xie X,et al. 2006. A bivalent chromatinstructure marks key developmentalgenes in embryonic stem cells. Cell125:315–326.
Bility MT, Thompson JT, McKee RH,et al. 2004. Activation of mouse andhuman peroxisome proliferator-acti-vated receptors (PPARs) by phthalatemonoesters. Toxicol Sci 82:170–182.
Bilkovski R, Schulte DM, Oberhauser F,et al. 2010. Role of WNT-5a in thedetermination of human mesenchy-mal stem cells into preadipocytes. JBiol Chem 285:6170–6178.
Blumberg B. 2010. Obesogens, stemcells and the maternal programming ofobesity. J Dev Origins Health Diseasedoi:10.1017/S2040174410000589.
Bromer JG, Wu J, Zhou Y, Taylor HS.2009. Hypermethylation of homeoboxA10 by in utero diethylstilbestrol ex-posure: an epigenetic mechanism foraltered developmental programming.Endocrinology 150:3376–3382.
Bunyan J, Murrell EA, Shah PP. 1976.The induction of obesity in rodentsby means of monosodium glutamate.Br J Nutr 35:25–39.
Burgers WA, Fuks F, Kouzarides T.2002. DNA methyltransferases getconnected to chromatin. TrendsGenet 18:275–277.
Butte NF, Christiansen E, Sorensen TI.2007. Energy imbalance underlyingthe development of childhood obesity.Obesity (Silver Spring) 15:3056–3066.
Chandra V, Huang P, Hamuro Y, et al.2008. Structure of the intact PPAR-gamma-RXR-alpha nuclear receptorcomplex on DNA. Nature 456:350–356.
Chen JQ, Brown TR, Russo J. 2009.Regulation of energy metabolismpathways by estrogens and estro-genic chemicals and potential impli-cations in obesity associated withincreased exposure to endocrine dis-ruptors. Biochim Biophys Acta 1793:1128–1143.
Cornelius P, MacDougald OA, Lane MD.1994. Regulation of adipocyte devel-opment. Annu Rev Nutr 14:99–129.
Crisan M, Yap S, Casteilla L, et al. 2008.A perivascular origin for mesenchymalstem cells in multiple human organs.Cell Stem Cell 3:301–313.
Crossno JT, Jr, Majka SM, Grazia T,et al. 2006. Rosiglitazone promotesdevelopment of a novel adipocytepopulation from bone marrow-derived circulating progenitor cells. JClin Invest 116:3220–3228.
da Silva Meirelles L, Caplan AI, NardiNB. 2008. In search of the in vivoidentity of mesenchymal stem cells.Stem Cells 26:2287–2299.
da Silva Meirelles L, Chagastelles PC,Nardi NB. 2006. Mesenchymal stemcells reside in virtually all post-natalorgans and tissues. J Cell Sci 119(Pt11):2204–2213.
Dahlman I, Mejhert N, Linder K, et al.2010. Adipose tissue pathwaysinvolved in weight loss of cancercachexia. Br J Cancer 102:1541–1548.
de Ferranti S, Mozaffarian D. 2008.The perfect storm: obesity, adipocytedysfunction, and metabolic conse-quences. Clin Chem 54:945–955.
Diamanti-Kandarakis E, BourguignonJP, Giudice LC, et al. 2009. Endo-crine-disrupting chemicals: an Endo-crine Society scientific statement.Endocr Rev 30:293–342.
Evans RM, Barish GD, Wang YX. 2004.PPARs and the complex journey toobesity. Nat Med 10:355–361.
Fava M. 2000. Weight gain and antide-pressants. J Clin Psychiatry 61(Suppl11):37–41.
Feige JN, Gelman L, Rossi D, et al.2007. The endocrine disruptor mono-ethyl-hexyl-phthalate is a selectiveperoxisome proliferator-activated re-ceptor gamma modulator that pro-motes adipogenesis. J Biol Chem282:19152–19166.
Fischbach C, Spruss T, Weiser B, et al.2004. Generation of mature fat padsin vitro and in vivo utilizing 3-D long-term culture of 3T3-L1 preadipo-cytes. Exp Cell Res 300:54–64.
Flegal KM, Carroll MD, Ogden CL, Cur-tin LR. 2010. Prevalence and trendsin obesity among US adults, 1999–2008. JAMA 303:235–241.
Fraga MF, Ballestar E, Paz MF, et al.2005. Epigenetic differences ariseduring the lifetime of monozygotictwins. Proc Natl Acad Sci USA102:10604–10609.
Friedman JM. 2009. Obesity: Causesand control of excess body fat. Na-ture 459:340–342.
Fu M, Sun T, Bookout AL, et al. 2005.A nuclear receptor atlas: 3T3-L1 adi-pogenesis. Mol Endocrinol 19:2437–2450.
Gluckman PD, Hanson MA. 2004. Liv-ing with the past: evolution, develop-ment, and patterns of disease. Sci-ence 305:1733–1736.
Gluckman PD, Hanson MA, SpencerHG. 2005. Predictive adaptiveresponses and human evolution.Trends Ecol Evol 20:527–533.
Glucksman ML, Hirsch J. 1968. Theresponse of obese patients to weightreduction: a clinical evaluation ofbehavior. Psychosom Med 30:1–11.
Glucksman ML, Hirsch J. 1969. Theresponse of obese patients to weightreduction. 3. The perception of bodysize. Psychosom Med 31:1–7.
Glucksman ML, Hirsch J, McCully RS,et al. 1968. The response of obesepatients to weight reduction. II. Aquantitative evaluation of behavior.Psychosom Med 30:359–373.
Gorin AA, Crane MM. 2009. the obeso-genic environment. In: Jelalian E,Steele RG, editors. Handbook ofchildhood and adolescent obesity.US: Springer. pp 145–161.
Greene ME, Pitts J, McCarville MA, et al.2000. PPARgamma: observations inthe hematopoietic system. Prostaglan-dins Other Lipid Mediat 62:45–73.
Grun F, Blumberg B. 2006. Environ-mental obesogens: organotins andendocrine disruption via nuclear re-ceptor signaling. Endocrinology147(6 Suppl):S50–S55.
Grun F, Blumberg B. 2009a. Endocrinedisrupters as obesogens. Mol CellEndocrinol 304:19–29.
Grun F, Blumberg B. 2009b. Minire-view: the case for obesogens. MolEndocrinol 23:1127–1134.
Grun F, Watanabe H, Zamanian Z, et al.2006. Endocrine-disrupting organotincompounds are potent inducers ofadipogenesis in vertebrates. MolEndocrinol 20:2141–2155.
Gupta RK, Arany Z, Seale P, et al.2010. Transcriptional control of prea-dipocyte determination by Zfp423.Nature 464:619–623.
Gustafson B, Gogg S, Hedjazifar S,et al. 2009. Inflammation and im-paired adipogenesis in hypertrophicobesity in man. Am J Physiol EndocrinolMetab v297:E999–D1003.
Habib ZA, Havstad SL, Wells K, et al.2010. Thiazolidinedione use and thelongitudinal risk of fractures inpatients with type 2 diabetes mellitus.J Clin Endocrinol Metab 95:592–600.
Hager A, Sjostrm L, Arvidsson B, et al.1977. Body fat and adipose tissuecellularity in infants: a longitudinalstudy. Metabolism 26:607–614.
Hager A, Sjorstrom L, Arvidsson B, et al.1978. Adipose tissue cellularity in obeseschool girls before and after dietarytreatment. Am J Clin Nutr 31:68–75.
Hajkova P, Erhardt S, Lane N, et al.2002. Epigenetic reprogramming inmouse primordial germ cells. MechDev 117:15–23.
Hales CN, Barker DJ. 2001. The thriftyphenotype hypothesis. Br Med Bull60:5–20.
Hallgren S, Sinjari T, Hakansson H,Darnerud PO. 2001. Effects of poly-brominated diphenyl ethers (PBDEs)
46 JANESICK AND BLUMBERG
Birth Defects Research (Part C) 93:34–50, (2011)

and polychlorinated biphenyls (PCBs)on thyroid hormone and vitamin Alevels in rats and mice. Arch Toxicol75:200–208.
Hatch EE, Nelson JW, Qureshi MM, et al.2008. Association of urinary phthalatemetabolite concentrations with bodymass index and waist circumference:a cross-sectional study of NHANESdata, 1999–2002. Environ Health7:27. doi:10.1186/1476-069X-7-27
Heintzman ND, Hon GC, Hawkins RD,et al. 2009. Histone modifications athuman enhancers reflect global cell-type-specific gene expression. Na-ture 459:108–112.
Heintzman ND, Ren B. 2009. Findingdistal regulatory elements in thehuman genome. Curr Opin Genet Dev19:541–549.
Heintzman ND, Stuart RK, Hon G,et al. 2007. Distinct and predictivechromatin signatures of transcrip-tional promoters and enhancers inthe human genome. Nat Genet 39:311–318.
Heinz S, Benner C, Spann N, et al.2010. Simple combinations of line-age-determining transcription factorsprime cis-regulatory elementsrequired for macrophage and B cellidentities. Mol Cell 38:576–589.
Herbstman JB, Sjodin A, Apelberg BJ,et al. 2008. Birth delivery modemodifies the associations betweenprenatal polychlorinated biphenyl(PCB) and polybrominated diphenylether (PBDE) and neonatal thyroidhormone levels. Environ Health Per-spect 116:1376–1382.
Hetherington AW, Ranson SW. 1942.The spontaneous activity and foodintake of rats with hypothalamiclesions. Am J Physiol 136:609–617.
Hill JO, Peters JC. 1998. Environmentalcontributions to the obesity epi-demic. Science 280:1371–1374.
Hines EP, White SS, Stanko JP, et al.2009. Phenotypic dichotomy follow-ing developmental exposure to per-fluorooctanoic acid (PFOA) in femaleCD-1 mice: low doses induce ele-vated serum leptin and insulin, andoverweight in mid-life. Mol Cell Endo-crinol 304:97–105.
Hiromori Y, Nishikawa J, Yoshida I,et al. 2009. Structure-dependentactivation of peroxisome proliferator-activated receptor (PPAR) gamma byorganotin compounds. Chem BiolInteract 180:238–244.
Hirsch J, Knittle JL. 1970. Cellularity ofobese and nonobese human adiposetissue. Fed Proc 29:1516–1521.
Hofstetter CP, Schwarz EJ, Hess D, et al.2002. Marrow stromal cells form guid-ing strands in the injured spinal cordand promote recovery. Proc Natl AcadSci USA 99:2199–2204.
Hoogduijn MJ, Rakonczay Z, GeneverPG. 2006. The effects of anticholiner-gic insecticides on human mesenchy-mal stem cells. Toxicol Sci 94:342–350.
Hoppe AA, Carey GB. 2007. Polybromi-nated diphenyl ethers as endocrinedisruptors of adipocyte metabolism.Obesity (Silver Spring) 15:2942–2950.
Hugo ER, Brandebourg TD, Woo JG,et al. 2008. Bisphenol A at environ-mentally relevant doses inhibits adi-ponectin release from human adiposetissue explants and adipocytes. Envi-ron Health Perspect 116:1642–1647.
Hurst CH, Waxman DJ. 2003. Activa-tion of PPARalpha and PPARgammaby environmental phthalate mono-esters. Toxicol Sci 74:297–308.
Inadera H, Shimomura A. 2005. Envi-ronmental chemical tributyltin aug-ments adipocyte differentiation. Toxi-col Lett 159:226–234.
Janesick A, Blumberg B. 2011. The roleof environmental obesogens in theobesity epidemic. In: Lustig RH, edi-tor. Obesity before birth. US:Springer. pp 383–399.
Janesick A, Blumberg B. 2010. Adipo-cytes as target cells for endocrinedisruption. In: Diamanti-KandarakisE, Gore AC, editors. Endocrine dis-rupters and puberty. New York, NY:Humana Press/Springer.
Janesick A, Blumberg B. PPARg as thetarget of obesogens. J Steroid Bio-chem Mol Biol (in press).
Jin Q, Zhang F, Yan T, et al. 2010. C/EBPalpha regulates SIRT1 expressionduring adipogenesis. Cell Res 20:470–479.
Jirtle RL, Skinner MK. 2007. Environ-mental epigenomics and disease sus-ceptibility. Nat Rev Genet 8:253–262.
Johns G, Leask A. 2009 May 31.Expert: It’s your fault if you’re afatty. The New Zealand Herald.
Jpenberg I, Jeannin E, Wahli W, Des-vergne B. 1997. Polarity and specificsequence requirements of peroxisomeproliferator-activated receptor (PPAR)/retinoid X receptor heterodimer bind-ing to DNA. A functional analysis of themalic enzyme gene PPAR response ele-ment. J Biol Chem 272:20108–20117.
Kaati G, Bygren LO, Pembrey M, Sjos-trom M. 2007. Transgenerationalresponse to nutrition, early life cir-cumstances and longevity. Eur JHum Genet 15:784–790.
Kahn CR. 2008. MEDICINE: can we nipobesity in its vascular bud? Science322:542–543.
Kanayama T, Kobayashi N, Mamiya S,et al. 2005. Organotin compoundspromote adipocyte differentiation asagonists of the peroxisome prolifera-tor-activated receptor gamma/reti-noid X receptor pathway. Mol Phar-macol 67:766–774.
Karmaus W, Osuch JR, Eneli I, et al.2009. Maternal levels of dichlorodi-phenyl-dichloroethylene (DDE) mayincrease weight and body mass indexin adult female offspring. Occup En-viron Med 66:143–149.
Kershaw EE, Flier JS. 2004. Adiposetissue as an endocrine organ. J ClinEndocrinol Metab 89:2548–2556.
Kim EH, Surh YJ. 2008. The role of 15-deoxy-delta(12,14)-prostaglandinJ(2), an endogenous ligand of perox-isome proliferator-activated receptorgamma, in tumor angiogenesis. Bio-chem Pharmacol 76:1544–1553.
Kim JB, Wright HM, Wright M, Spiegel-man BM. 1998. ADD1/SREBP1 acti-vates PPARgamma through the pro-duction of endogenous ligand. ProcNatl Acad Sci USA 95:4333–4337.
Kim JY, van de Wall E, Laplante M, et al.2007. Obesity-associated improve-ments in metabolic profile throughexpansion of adipose tissue. J ClinInvest 117:2621–2637.
Kinzel B. 2010. Targeting Obesity, Sor-rell Proposes ‘Soda Tax’ Montpelier,Vt.: Vermont Public Radio.
Kirchner S, Kieu T, Chow C, et al.2010. Prenatal exposure to the envi-ronmental obesogen tributyltin pre-disposes multipotent stem cells tobecome adipocytes. Mol Endocrinol24:526–539.
Kirchner S, Ramirez M, Chow C, Blum-berg B. 2011. Bisphenol A diglycidylether (BADGE) favors adipogenic dif-ferentiation of multipotent stromalstem cells (MSCs) through a peroxi-some proliferator activated receptorgamma (PPARc)-independent mecha-nism. submitted for publication.
Klimentidis YC, Beasley TM, Lin HY,et al. 2010. Canaries in the coal mine:a cross-species analysis of the plural-ity of obesity epidemics. Proc Biol Sci.
Klont RE, Brocks L, Eikelenboom G. 1998.Muscle fibre type and meat quality.Meat Sci 49(Suppl 1):S219–S229.
Knittle JL. 1972. Obesity in childhood: aproblem in adipose tissue cellular de-velopment. J Pediatr 81:1048–1059.
Knittle JL, Timmers K, Ginsberg-FellnerF, et al. 1979. The growth of adiposetissue in children and adolescents.Cross-sectional and longitudinalstudies of adipose cell number andsize. J Clin Invest 63:239–246.
Koebnick C, Smith N, Coleman KJ,et al. 2010. Prevalence of extremeobesity in a multiethnic cohort ofchildren and adolescents. J Pediatr157:26–31 e22.
Koh YJ, Kang S, Lee HJ, et al. 2007.Bone marrow-derived circulating pro-genitor cells fail to transdifferentiateinto adipocytes in adult adipose tis-sues in mice. J Clin Invest 117:3684–3695.
Kubo M, Ijichi N, Ikeda K, et al. 2009.Modulation of adipogenesis-relatedgene expression by estrogen-relatedreceptor gamma during adipocyticdifferentiation. Biochim Biophys Acta1789:71–77.
Kumar N, Solt LA, Wang Y, et al. 2010.Regulation of adipogenesis by natu-ral and synthetic REV-ERB ligands.Endocrinology 151:3015–3025.
Kuzawa CW. 1998. Adipose tissue inhuman infancy and childhood: an ev-olutionary perspective. Am J PhysAnthropol Suppl 27:177–209.
ENDOCRINE DISRUPTORS AND OBESITY 47
Birth Defects Research (Part C) 93:34–50, (2011)

Lan F, Bayliss PE, Rinn JL, et al. 2007.A histone H3 lysine 27 demethylaseregulates animal posterior develop-ment. Nature 449:689–694.
Larsen TM, Toubro S, Astrup A. 2003.PPARgamma agonists in the treatmentof type II diabetes: is increased fat-ness commensurate with long-termefficacy? Int J Obes Relat Metab Disord27:147–161.
Lau DC, Shillabeer G, Wong KL, et al.1990. Influence of paracrine factorson preadipocyte replication and dif-ferentiation. Int J Obes 14(Suppl3):193–201.
le Maire A, Grimaldi M, Roecklin D,et al. 2009. Activation of RXR-PPARheterodimers by organotin environ-mental endocrine disruptors. EMBORep 10:367–373.
Lefterova MI, Steger DJ, Zhuo D, et al.2010. Cell-specific determinants ofperoxisome proliferator-activated re-ceptor gamma function in adipocytesand macrophages. Mol Cell Biol 30:2078–2089.
Lehmann JM, Moore LB, Smith-OliverTA, et al. 1995. An antidiabetic thia-zolidinedione is a high affinity ligandfor peroxisome proliferator-activatedreceptor gamma (PPAR gamma). JBiol Chem 270:12953–12956.
Li L, Xie X, Qin J, et al. 2009. The nu-clear orphan receptor COUP-TFIIplays an essential role in adipogene-sis, glucose homeostasis, and energymetabolism. Cell Metab 9:77–87.
Liu ZJ, Zhuge Y, Velazquez OC. 2009.Trafficking and differentiation ofmesenchymal stem cells. J Cell Bio-chem 106:984–991.
Lu C, Cheng SY. 2010. Thyroid hor-mone receptors regulate adipogene-sis and carcinogenesis via crosstalksignaling with peroxisome prolifera-tor-activated receptors. J Mol Endo-crinol 44:143–154.
Luff AR, Goldspink G. 1967. Large andsmall muscles. Life Sci 6:1821–1826.
Lustig RH. 2006. Childhood obesity:behavioral aberration or biochemicaldrive? Reinterpreting the First Law ofThermodynamics. Nat Clin PractEndocrinol Metab 2:447–458.
Marques BG, Hausman DB, Martin RJ.1998. Association of fat cell size andparacrine growth factors in develop-ment of hyperplastic obesity. Am JPhysiol 275(6 Pt 2):R1898–R1908.
Martinez M. 2010. San Francisco over-rides mayoral veto, bans HappyMeals with toys.
Masuno H, Iwanami J, Kidani T, et al.2005. Bisphenol a accelerates termi-nal differentiation of 3T3-L1 cells intoadipocytes through the phosphatidyl-inositol 3-kinase pathway. Toxicol Sci84:319–327.
Matsuyama S, Oki Y, Yokoki Y. 1973.Obesity induced by monosodium glu-tamate in mice. Natl Inst AnimHealth Q (Tokyo) 13:91–101.
McCormick DP, Sarpong K, Jordan L,et al. 2010. Infant obesity: are we
ready to make this diagnosis? JPediatrics 157:15–19.
Mikkelsen TS, Ku M, Jaffe DB, et al. 2007.Genome-wide maps of chromatin statein pluripotent and lineage-committedcells. Nature 448:553–560.
Mohamed-Ali V, Pinkney JH, CoppackSW. 1998. Adipose tissue as an endo-crine and paracrine organ. Int J ObesRelat Metab Disord 22:1145–1158.
Morganstein DL, Wu P, Mane MR, et al.2010. Human fetal mesenchymalstem cells differentiate into brownand white adipocytes: a role forERRalpha in human UCP1 expres-sion. Cell Res 20:434–444.
Muhlhausler B, Smith SR. 2009. Early-life origins of metabolic dysfunction:role of the adipocyte. Trends Endo-crinol Metab 20:51–57.
Nemeroff CB. 1997. Dosing the anti-psychotic medication olanzapine. JClin Psychiatry 58(Suppl)10:45–49.
Newbold RR, Padilla-Banks E, JeffersonWN. 2009. Environmental estrogensand obesity. Mol Cell Endocrinol 304:84–89.
Newburgh LH, Johnston MW. 1930.The nature of obesity. J Clin Invest 8:197–213.
Nielsen R, Pedersen TA, Hagenbeek D,et al. 2008. Genome-wide profiling ofPPARgamma:RXR and RNA polymer-ase II occupancy reveals temporalactivation of distinct metabolic path-ways and changes in RXR dimercomposition during adipogenesis.Genes Dev 22:2953–2967.
Noer A, Lindeman LC, Collas P. 2009.Histone H3 modifications associatedwith differentiation and long-term cul-ture of mesenchymal adipose stemcells. Stem Cells Dev 18:725–736.
Ohoka N, Kato S, Takahashi Y, et al.2009. The orphan nuclear receptorRORalpha restrains adipocyte differen-tiation through a reduction of C/EBP-beta activity and perilipin gene expres-sion. Mol Endocrinol 23:759–771.
Oken E, Huh SY, Taveras EM, et al.2005. Associations of maternal pre-natal smoking with child adiposityand blood pressure. Obes Res 13:2021–2028.
Orlic D, Kajstura J, Chimenti S, et al.2001. Bone marrow cells regenerateinfarcted myocardium. Nature410:701–705.
Park KW, Halperin DS, Tontonoz P.2008. Before they were fat: adipocyteprogenitors. Cell Metab 8:454–457.
Park KW, Waki H, Choi SP, et al. 2010.The small molecule phenamil is amodulator of adipocyte differentia-tion and PPARgamma expression.J Lipid Res 51:2775–2784.
Paul KB, Hedge JM, Devito MJ, CroftonKM. 2010. Developmental triclosanexposure decreases maternal andneonatal thyroxine in rats. EnvironToxicol Chem 29:2840–2844.
Pearson AM. 1990. Muscle growth andexercise. Crit Rev Food Sci Nutr29:167–196.
Picard F, Kurtev M, Chung N, et al.2004. Sirt1 promotes fat mobilizationin white adipocytes by repressingPPAR-gamma. Nature 429:771–776.
Power C, Jefferis BJ. 2002. Fetal envi-ronment and subsequent obesity: astudy of maternal smoking. Int J Epi-demiol 31:413–419.
Rabin RC. 2008 January 22. In theFatosphere, Big Is In, or at LeastAccepted. The New York Times.
Rehfeldt C, Fiedler I, Wegner J. 1987.Changes in the microstructure ofmuscle tissue in laboratory mice, cat-tle and swine during growth. Z Mik-rosk Anat Forsch 101:669–680.
Robertson KD, Ait-Si-Ali S, Yokochi T,et al. 2000. DNMT1 forms a complexwith Rb, E2F1 and HDAC1 andrepresses transcription from E2F-responsive promoters. Nat Genet 25:338–342.
Rodeheffer MS, Birsoy K, Friedman JM.2008. Identification of white adipo-cyte progenitor cells in vivo. Cell 135:240–249.
Rodriguez JP, Montecinos L, Rios S,et al. 2000. Mesenchymal stem cellsfrom osteoporotic patients produce atype I collagen-deficient extracellularmatrix favoring adipogenic differen-tiation. J Cell Biochem 79:557–565.
Rodriguez PE, Sanchez MS. 2010.Maternal exposure to triclosanimpairs thyroid homeostasis andfemale pubertal development in wis-tar rat offspring. J Toxicol EnvironHealth A 73:1678–1688.
Roh TY, Cuddapah S, Cui K, Zhao K.2006. The genomic landscape of his-tone modifications in human T cells.Proc Natl Acad Sci USA 103:15782–15787.
Rosen CJ, Bouxsein ML. 2006. Mecha-nisms of disease: is osteoporosis theobesity of bone? Nat Clin Pract Rheu-matol 2:35–43.
Rosen ED, MacDougald OA. 2006.Adipocyte differentiation from theinside out. Nat Rev Mol Cell Biol 7:885–896.
Rosen ED, Sarraf P, Troy AE, et al.1999. PPAR gamma is required forthe differentiation of adipose tissue invivo and in vitro. Mol Cell 4:611–617.
Rosen ED, Walkey CJ, Puigserver P,Spiegelman BM. 2000. Transcrip-tional regulation of adipogenesis.Genes Dev 14:1293–1307.
Rosenbaum AJ, Grande DA, Dines JS.2008. The use of mesenchymal stemcells in tissue engineering: a globalassessment. Organogenesis 4:23–27.
Rosenbaum M, Leibel RL. 2010. Adapt-ive thermogenesis in humans. Int JObes (Lond) 34(Suppl 1):S47–S55.
Ross SE, Hemati N, Longo KA, et al.2000. Inhibition of adipogenesis byWnt signaling. Science 289:950–953.
Rubenstrunk A, Hanf R, Hum DW,et al. 2007. Safety issues and pros-pects for future generations of PPARmodulators. Biochim Biophys Acta1771:1065–1081.
48 JANESICK AND BLUMBERG
Birth Defects Research (Part C) 93:34–50, (2011)

Rubin BS, Murray MK, Damassa DA,et al. 2001. Perinatal exposure to lowdoses of bisphenol A affects bodyweight, patterns of estrous cyclicity,and plasma LH levels. Environ HealthPerspect 109:675–680.
Rubin BS, Soto AM. 2009. Bisphenol A:Perinatal exposure and body weight.Mol Cell Endocrinol 304:55–62.
Rupnick MA, Panigrahy D, Zhang CY,et al. 2002. Adipose tissue mass canbe regulated through the vascula-ture. Proc Natl Acad Sci USA 99:10730–10735.
Ryan KK, Haller AM, Sorrell JE, et al.2010. Perinatal exposure to bisphe-nol-a and the development of meta-bolic syndrome in CD-1 mice. Endo-crinology 151:2603–2612.
Saito T, Abe D, Sekiya K. 2009. Flava-none exhibits PPARgamma ligand ac-tivity and enhances differentiation of3T3-L1 adipocytes. Biochem BiophysRes Commun 380:281–285.
Sakurai K, Kawazuma M, Adachi T, et al.2004. Bisphenol A affects glucosetransport in mouse 3T3-F442A adipo-cytes. Br J Pharmacol 141:209–214.
Salans LB, Cushman SW, WeismannRE. 1973. Studies of human adiposetissue. Adipose cell size and numberin nonobese and obese patients.J Clin Invest 52:929–941.
Sargis RM, Johnson DN, Choudhury RA,Brady MJ. 2010. Environmental endo-crine disruptors promote adipogene-sis in the 3T3-L1 cell line throughglucocorticoid receptor activation. Obe-sity (Silver Spring) 18:1283–1288.
Seale P, Bjork B, Yang W, et al. 2008.PRDM16 controls a brown fat/skeletalmuscle switch. Nature 454:961–967.
Seo JB, Moon HM, Kim WS, et al.2004. Activated liver X receptorsstimulate adipocyte differentiationthrough induction of peroxisome pro-liferator-activated receptor gammaexpression. Mol Cell Biol 24:3430–3444.
Shao D, Lazar MA. 1997. Peroxisomeproliferator activated receptorgamma, CCAAT/enhancer-bindingprotein alpha, and cell cycle statusregulate the commitment to adipo-cyte differentiation. J Biol Chem272:21473–21478.
Sheldon W. 1940. The varieties ofhuman physique: an introduction toconstitutional psychology. New York:Harper & Bros.
Sheldon WH, Dupertuis CW, McDer-mott E. 1954. Atlas of men: a guidefor somatotyping the adult male atall ages. J Am Med Assoc 156:1294-c-1295.
Shillabeer G, Forden JM, Lau DC. 1989.Induction of preadipocyte differentia-tion by mature fat cells in the rat.J Clin Invest 84:381–387.
Shim WS, Do MY, Kim SK, et al. 2006.The long-term effects of rosiglitazoneon serum lipid concentrations andbody weight. Clin Endocrinol (Oxf)65:453–459.
Shockley KR, Rosen CJ, Churchill GA,Lecka-Czernik B. 2007. PPARgamma2regulates a molecular signature ofmarrow mesenchymal stem cells.PPAR Res 2007:81219.
Siersbaek R, Nielsen R, Mandrup S.2010. PPARgamma in adipocyte dif-ferentiation and metabolism—novelinsights from genome-wide studies.FEBS Lett.
Sims EA, Horton ES. 1968. Endocrineand metabolic adaptation to obesityand starvation. Am J Clin Nutr 21:1455–1470.
Skinner MK, Guerrero-Bosagna C.2009. Environmental signals andtransgenerational epigenetics. Epige-nomics 1:111–117.
Slotkin TA. 2010. Does early-life expo-sure to organophosphate insecticideslead to prediabetes and obesity?Reprod Toxicol.
Smink A, Ribas-Fito N, Garcia R, et al.2008. Exposure to hexachloroben-zene during pregnancy increases therisk of overweight in children aged 6years. Acta Paediatr 97:1465–1469.
Smith CJ, Vasta V, Degerman E, et al.1991. Hormone-sensitive cyclic GMP-inhibited cyclic AMP phosphodiester-ase in rat adipocytes. Regulation ofinsulin- and cAMP-dependent activa-tion by phosphorylation. J Biol Chem266:13385–13390.
Somm E, Schwitzgebel VM, Vauthay DM,et al. 2008. Prenatal nicotine exposurealters early pancreatic islet and adi-pose tissue development with conse-quences on the control of body weightand glucose metabolism later in life.Endocrinology 149:6289–6299.
Sorisky A, Magun R, Gagnon AM.2000. Adipose cell apoptosis: deathin the energy depot. Int J Obes RelatMetab Disord 24(Suppl 4):S3–S7.
Spalding KL, Arner E, Westermark PO,et al. 2008. Dynamics of fat cell turn-over in humans. Nature 453:783–787.
Spiegelman BM, Flier JS. 2001. Obesityand the regulation of energy balance.Cell 104:531–543.
Stahlhut RW, van Wijgaarden E, DyeTD, et al. 2007. Concentrations ofurinary phthalate metabolites areassociated with increased waist cir-cumferece and insulin resistance inadult U.S. males. Environ Health Per-spect 115:876–882.
Stern JS, Batchelor BR, Hollander N,et al. 1972. Adipose-cell size and im-munoreactive insulin levels in obeseand normal-weight adults. Lancet2:948–951.
Student AK, Hsu RY, Lane MD. 1980.Induction of fatty acid synthetase syn-thesis in differentiating 3T3-L1 preadi-pocytes. J Biol Chem 255:4745–4750.
Styner M, Sen B, Xie Z, et al. 2010. In-domethacin promotes adipogenesis ofmesenchymal stem cells through acyclooxygenase independent mecha-nism. J Cell Biochem 111:1042–1050.
Sutanto MM, Ferguson KK, Sakuma H,et al. 2010. The silencing mediator of
retinoid and thyroid hormone recep-tors (SMRT) regulates adipose tissueaccumulation and adipocyte insulinsensitivity in vivo. J Biol Chem 285:18485–18495.
Takacs ML, Abbott BD. 2007. Activa-tion of mouse and human peroxi-some proliferator-activated receptors(a,b/d,c) by perfluorooctanoic acidand perfluorooctane sulfonate. Toxi-col Sci 95:108–117.
Takada I, Kouzmenko AP, Kato S.2009. Wnt and PPARgamma signal-ing in osteoblastogenesis and adipo-genesis. Nat Rev Rheumatol 5:442–447.
Takahashi N, Goto T, Taimatsu A, et al.2009. Bixin regulates mRNA expres-sion involved in adipogenesis andenhances insulin sensitivity in 3T3-L1adipocytes through PPARgammaactivation. Biochem Biophys ResCommun 390:1372–1376.
Tang QQ, Otto TC, Lane MD. 2004.Commitment of C3H10T1/2 pluripo-tent stem cells to the adipocyte line-age. Proc Natl Acad Sci USA 101:9607–9611.
Tang W, Zeve D, Suh JM, et al. 2008.White fat progenitor cells reside inthe adipose vasculature. Science322:583–586.
Taveras EM, Rifas-Shiman SL, Belfort MB,et al. 2009. Weight status in the first 6months of life and obesity at 3 years ofage. Pediatrics 123:1177–1183.
Timmons JA, Wennmalm K, Larsson O,et al. 2007. Myogenic gene expres-sion signature establishes that brownand white adipocytes originate fromdistinct cell lineages. Proc Natl AcadSci USA 104:4401–4406.
Tontonoz P, Hu E, Spiegelman BM.1994. Stimulation of adipogenesis infibroblasts by PPAR gamma 2, alipid-activated transcription factor.Cell 79:1147–1156.
Tontonoz P, Nagy L, Alvarez JG, et al.1998. PPARgamma promotes mono-cyte/macrophage differentiation anduptake of oxidized LDL. Cell 93:241–252.
Tontonoz P, Spiegelman BM. 2008. Fatand beyond: the diverse biology ofPPARgamma. Annu Rev Biochem 77:289–312.
Van Harmelen V, Reynisdottir S, Eriks-son P, et al. 1998. Leptin secretionfrom subcutaneous and visceral adi-pose tissue in women. Diabetes47:913–917.
Van Inwegen RG, Robison GA, Thomp-son WJ. 1975. Cyclic nucleotidephosphodiesterases and thyroid hor-mones. J Biol Chem 250:2452–2456.
Vanden Heuvel JP, Thompson JT,Frame SR, Gillies PJ. 2006. Differen-tial activation of nuclear receptors byperfluorinated fatty acid analogs andnatural fatty acids: a comparison ofhuman, mouse, and rat peroxisomeproliferator-activated receptor-alpha,-beta, and -gamma, liver X receptor-
ENDOCRINE DISRUPTORS AND OBESITY 49
Birth Defects Research (Part C) 93:34–50, (2011)

beta, and retinoid X receptor-alpha.Toxicol Sci 92:476–489.
Vazquez-Vela ME, Torres N, Tovar AR.2008. White adipose tissue as endo-crine organ and its role in obesity.Arch Med Res 39:715–728.
Verhulst SL, Nelen V, Hond ED, et al.2009. Intrauterine exposure to envi-ronmental pollutants and body massindex during the first 3 years of life. En-viron Health Perspect 117:122–126.
Verma S, Rajaratnam JH, Denton J,et al. 2002. Adipocytic proportion ofbone marrow is inversely related tobone formation in osteoporosis. JClin Pathol 55:693–698.
Viguerie N, Millet L, Avizou S, et al.2002. Regulation of human adipocytegene expression by thyroid hormone.J Clin Endocrinol Metab 87:630–634.
Walkey CJ, Spiegelman BM. 2008. Afunctional peroxisome proliferator-activated receptor-gamma ligand-binding domain is not required foradipogenesis. J Biol Chem 283:24290–24294.
Walley AJ, Asher JE, Froguel P. 2009.The genetic contribution to non-syn-dromic human obesity. Nat RevGenet 10:431–442.
Wang J, Lazar MA. 2008. Bifunctionalrole of Rev-erbalpha in adipocyte dif-ferentiation. Mol Cell Biol 28:2213–2220.
Wegner J, Albrecht E, Fiedler I, et al.2000. Growth- and breed-relatedchanges of muscle fiber characteristicsin cattle. J Anim Sci 78:1485–1496.
Wei G, Wei L, Zhu J, et al. 2009. Globalmapping of H3K4me3 and H3K27me3reveals specificity and plasticity inlineage fate determination of dif-ferentiating CD41 T cells. Immunity30:155–167.
Wertheimer E, Shapiro B. 1948. Thephysiology of adipose tissue. PhysiolRev 28:451–464.
Wood RJ. 2008. Vitamin D and adipo-genesis: new molecular insights.Nutr Rev 66:40–46.
Wu Z, Bucher NL, Farmer SR. 1996.Induction of peroxisome proliferator-
activated receptor gamma duringthe conversion of 3T3 fibroblastsinto adipocytes is mediated by C/EBPbeta, C/EBPdelta, and glucocorti-coids. Mol Cell Biol 16:4128–4136.
Yang Y, Shang W, Zhou L, et al.2007. Emodin with PPARgammaligand-binding activity promotes adi-pocyte differentiation and increasesglucose uptake in 3T3-Ll cells. Bio-chem Biophys Res Commun353:225–230.
Zhang X, Ji J, Yan G, et al. 2010. Silde-nafil promotes adipogenesis througha PKG pathway. Biochem BiophysRes Commun 396:1054–1059.
Zhang Y, Proenca R, Maffei M, et al.1994. Positional cloning of the mouseobese gene and its human homo-logue. Nature 372:425–432.
Zhao Y, Bruemmer D. 2010. NR4Aorphan nuclear receptors: transcrip-tional regulators of gene expressionin metabolism and vascular biology.Arterioscler Thromb Vasc Biol 30:1535–1541.
50 JANESICK AND BLUMBERG
Birth Defects Research (Part C) 93:34–50, (2011)





























