Embed Size (px)
Citation preview

American Journal of Primatology 1:157- 165 (1981)
Endocrine Control of Spermatogenesis in Primates CHARLES E. GRAHAM Reproductive Biology Diuzswn, Primate Research Imtitute, New Mexico State University, Holloman AFB, New Mexico
Control of primate spermatogenesis is reviewed in terms of endogenous regula- tory mechanisms and endocrine approaches to contraception and treatment of infertility. The role of gonadotropins and steroid hormones in maintaining spermatogenesis in primates is incompletely understood because A) hormonal interactions are complex, and B) most studies have used rodents rather than primates. Feedback control, interaction of LH and testosterone, the role of androgen, androgen in secondary sex organs, regulation of receptor proteins, roles of prolactin and growth hormone, and the breakdown and modification of the endocrine control mechanism are reviewed. The treatment of infertility with GnRH, gonadotropins, and androgen is discussed. Information is included on contraception research using the following methods: immunization against GnRH, use of GnRH analogs, immunization against gonadotropins, induced suppression of FSH secretion or action, and steroid suppression of sper- matogenesis. The importance of studying testicular steroid metabolism in pri- mates is stressed. Significant advances in the understanding of endocrine con- trol of spermatogenesis have been made in recent years, but no primate species have been thoroughly studied. Variability between species in endocrine control mechanisms is an important factor in selecting primate models, and it is clear that such models can be valuable in the development of male contraceptives.
Key words: androgens, contraception, endocrine control, follicle stimulating hormone (FSH), gonadotropin releasing hormone (GnRH), infertility, lutenizing hormone (LH), primates, spermatogenesis, testosterone
INTRODUCTION Control of primate spermatogenesis is reviewed in terms of the endogenous regula- tory mechanisms and endocrine approaches to treatment of infertility and contraception. Much of our basic understanding of the subject is drawn from studies of rodents and our experience of infertility management from the human.
ENDOGENOUS REGULATION OF SPERMATOGENESIS The role of gonadotropins and steroid hormones in maintaining spermatogenesis is
incompletely understood, in part because of complex hormonal interactions, and also because there are differences between rodents, in which most studies have been done, and primates [review by Steinberger & Steinberger, 19721.
Received January 10, 1981; accepted January 23, 1981
Address reprint requests to Charles E. Graham, New Mexico State University, P.O. Box 1027, Holloman AFB, NM 88330.
0275-2565/81/0102-0157$03.00 0 1981 Alan R. Liss, Inc.

158 Graham
Feedback Control of Gonadotropin Secretion Castration of the adult primate results in a marked elevation of circulating luteinizing
hormone (LH) and follicle stimulating hormone (FSH) levels, and administration of testosterone results in depression of LH to precastration levels, indicating the presence of one or more regulatory feedback loops. Possible sites of action of testosterone or its metabolites could be the brain, presumably the hypothalamus, so regulating the release of gonadotropin releasing hormone (GnRH); or the pituitary, regulating the sensitivity of the gonadotropins to GnRH. Testosterone administration is incapable of restoring FSH to precastration levels, nor
can large doses of testosterone inhibit circulating levels of FSH in intact men [Swerdloff & Odell, 19681. Since testosterone negative feedback on the hypothalamus would be expected to affect both gonadotropins equally, it appears likely that testosterone selec- tively inhibits the sensitivity of the LH gonadotrophs to GnRH. I t also appears that another hormone, dubbed “inhibin,” may provide negative feedback inhibition of FSH in intact males.
Role of Gonadotropins in Control of Spermatogenesis Although LH or human chorionic gonadotropin (hCG) alone can maintain Leydig cell
function and basic spermatogenesis in hypophysectomized rodents, FSH appears to have a central role in the control of spermatogenesis in primates. Gonadotropin preparations rich in FSH, such as human menopausal gonadotropin (hMG), can produce full sper- matogenesis in human hypogonadotropic eunuchs [Paulsen, 19681 and hypophysec- tomized men [MacLeod et al, 1964, 19661. Specific receptors exist for FSH in the testis: labeled FSH selectively binds to the tubules [Means & Vaitukaitis, 19721 and to isolated Sertoli cells, inducing a dramatic increase in cyclic AMP [Steinberger et al, 19751. The relative roles of LH and FSH are unclear because there is a shortage of purified LH and FSH preparations; heterologous gonadotropins can induce neutralizing antibodies [Maddock, 19491.
Interaction of LH and Testosterone in Control of Spermatogenesis It is generally accepted that LH stimulation of the Leydig cells maintains a high, local,
peritubular concentration of testosterone essential for spermatogenesis. The mechanism by which LH controls androgen secretion has been explored by Arslan
et a1 [1978], who showed that testosterone production can be stimulated in vitro by hCG in adult, but not in prepubertal or juvenile rhesus monkeys. However, priming in vivo with 300 IU hCG daily for five days, sensitized the immature testis to hCG in vitro. Arslan et a1 also showed that mature and immature monkeys can be stimulated with chronic hCG in vivo to increase testosterone levels as much as sevenfold and twofold, respectively. These data suggest that chronic hCG exposure activates the response system, probably by enhancing LH/hCG receptor availability.
Role of Androgen in Support of Spermatogenesis In the hypophysectomized rat, testosterone alone can substantially maintain sper-
matogenesis; whereas in the rhesus monkey, spermatogenesis is only partly maintained [Smith, 19381. In hypophysectomized men treated with testosterone, the recovery of the seminiferous epithelium is much poorer than in the other species; only spermatogonia and Sertoli cells remain in the seminiferous tubules [Smith, 19381. In view of these interspecific differences, primate models for study of the endocrine control of sper- matogenesis should be chosen with great care. The mode of action of androgen in supporting spermatogenesis remains unclear, and
whether the active androgen acts on the Sertoli cell or directly on the germ cells is unknown.

Endocrinology of Primate Spermatogenesis 159
Androgen Action in Secondary Sex Organs Since the secondary sex structures depend upon androgen for their integrity, androgen
affects sperm maturation at least indirectly. Androgen action in epididymal and testicu- lar epithelia is mediated by a cytoplasmic receptor protein [Hansson et al, 19731. A separate binding protein is produced by Sertoli cells and is secreted into the rete testis fluid and caput epididymidis, probably accounting for the high concentration of androgen in these locations and providing additional input for androgen on sperm maturation.
Regulation of Receptor Proteins As previously mentioned, LH levels regulate the availability of LH receptor in the
Leydig cells. Prolactin, and perhaps other hormones, also does so. It should be anticipated that similar endocrine mechanisms regulate the availability of FSH and androgen receptors since estrogen can induce FSH receptors in the rat ovary ILouvet & Vaitu- kaitis, 19761.
Role of Prolactin and Growth Hormone in Control of Spermatogenesis In rodents, prolactin potentiates the action of LH in stimulating spermatogcnesis
[review by Bartke et al, 19781. Prolactin may also play a key role in seasonal testicular regression. In the Syrian hamster, prolactin, but not exogenous gonadotropins, reverses the testicular regression and depressed gonadotropin levels induced by a shortened photoperiod. Prolactin also has antigonadotropic effects, however, and it is noteworthy that prolactin levels are inversely correlated with seasonal testicular activity in the rhesusmonkey [Wickings & Nieschlag, 19801. Seasonal primates such asprosimians, the squirrel monkey (Saimiri sczureus), and the rhesus monkey [Bernstein et al, 1974; Kinzey, 19711 may be valuable models for the endocrine control of spermatogenesis. The tropic action of prolactin appears to involve increasing the number of LH receptors
in Leydig cells. Proper prolactin balance is essential; hyperprolactinemia in men can produce hypogonadism and impotence [review by Bartke et al, 19781. Pharmacological elevation of prolactin in normal men can result in testosterone elevation [Rubin et al, 19781. Growth hormone overlaps prolactin in many of its biological properties; it, too, can
partially reverse the effects of shortened photoperiod in the hamster. The role of both of these hormones in primate spermatogenesis needs to be defined.
Breakdown and Modification of the Endocrine Control Mechanism It is clear that such a complex hormonal regulatory system offers many possibilities for
failure and points of entry for contraception. Thelability of the system is indicated by the fact that high doses of FSH in the rat [Doener & Deckart, 19621 and of hCG in men [Maddock & Nelson, 19521 and low doses of androgens, estrogens, and progestins can damage the seminiferous epithelium or inhibit spermatogenesis. The effect of steroids is secondary to blockage of gonadotropin secretion 1 Ludwig, 19501.
TREATMENT OF INFERTILITY The chief problem in the development of strategies for treatment of human fertility has
been lack of adequate controlled studies, variable causes of infertility, and biases intro- duced by case selection. There is a need for replicable primate models of human endo- crine-based infertility in which the endocrine requirements for initiation and mainte- nance of spermatogenesis can be evaluated. Careful validation of such models must be undertaken before results can be extrapolated to the human with confidence.
Treatment of Infertility With GnRH Although the synthesis of GnRH and highly potent agonistic analogs promised an
effective approach to the treatment of infertility associated with an intact pituitary

160 Graham
gland, it was soon found that these agents actually caused damage to the rodent and human seminiferous epithelium. Perhaps we should have been forewarned by the discov- ery of Maddock and Nelson in 1952 that hCG administration could block sper- matogenesis in rats.
Treatment of Infertility With Gonadotropins As was mentioned earlier, crude gonadotropin preparations rich in FSH, such as hMG,
are capable of inducing or maintaining spermatogenesis in some hypogonadotropic patients. The success of such preparations may depend on their partially purified state, since the exact endocrine requirements of spermatogenesis remain undefined.
Treatment of Infertility With Androgen Treatment with testosterone seems to have met with little success, probably because it
prevents the attainment of sufficient intratesticular levels of testosterone because of negative feedback inhibition of LH secretion and, hence, lack of Leydig cell activity.
CONTRACEPTION Immunization Against GnRH Hodgen and Hearn 11977 I immunized five male marmosets (Callithrixjacchus) with
GnRH-BSA and adjuvants; three of the five males developed high antibody titer, marked testicular atrophy, and suppression of testosterone secretion. Histological study showed marked inhibition of seminiferous tubules and of spermatogenesis and a reduction in size of Leydig cells. In addition, animals with a high anti-GnRH titer showed a reduced LH response to exogenous GnRH. Chappel et a1 [1980], in a similar study with male rhesus monkeys, also noted a
reversible inhibition of peripheral serum LH and testosterone levels inversely correlated with GnRH antibody levels. A dramatic reduction in testis effluent, pituitary content of LH, venous blood concentration of testosterone, estradiol, dihydrotestosterone, and pro- gesterone also occurred. The immunological approach will become more promising if the need for adjuvant can
be overcome.
Administration of GnRH Analogs Competitive antagonistic analogs of GnRH have antifertility action in male rodents. In
female chimpanzees they can block the induction of LH surges induced by exogenous GnRH, suggesting their potential for ovulation supression [Gosselin et al, 19791 ; how- ever, the antagonists have not been explored as contraceptive agents in male nonhuman primates. Agonistic analogs of GnRH also have potent antifertility action. Such compounds in the
male rat result in involution ofthe seminiferous tubules and accessory organs. Auclair et a1 [1979] and Defau et a1 119791 have shown that potent GnRH agonists, GnRH itself, or hCG result in great reductions in the number of testicular LH-hCG and prolactin receptors, and of testosterone production. Direct effects on the gonads have also been postulated, based on the observation that GnRH inhibits FSH-induced steroidogenesis in rat granulosa cells in vitro and in hypophsectomized rats [Hsueh & Erickson, 19791. Another possible mode of action is pituitary desensitization by the high level of stimula- tion exerted by superagonists. Attempts to inhibit spermatogenesis in nonhuman pri- mates with superagonists have met with mixed results. Wickings and associates in this symposium, and Vickery and McRae [1980b, and personal communication] have noted no depression of testosterone levels or of spermatogenesis in rhesus monkeys or M . fascicularis. By contrast a marked depression of testosterone levels has been noted in

Endocrinology of Primate Spermatogenesis 161
baboons treated with superagonists of GhRH [Vickery & McRae, 198Oal. To what extent these differences reflect interspecific differences or experimental variables remains to be defined. Primates have considerable value as models for the development of contraceptive
strategies using GnRH analogs. However, careful species selection is necessary in view of interspecific differences in response to GnRH and analogs. In addition to examples already mentioned, it may be noted that acute GnRH administration does not elicit consistent responses in rhesus monkeys, whereas it does in chimpanzees and women [Hobson & Fuller, 1977; Graham et al, 1979; Yen et al, 19731. The bonnet monkey (Macaca radiatu) is sensitive to GnRH, but it is totally insensitive
to an analog highly potent in rodents [Levitan et al, 19771. Poor activity correlations have also been noted for some GnRH antagonists between rodents and male and female chimpanzees [Bowers et al, 1980; Graham & Gosselin, unpublished observation]. The significance of these differences for humans are not yet clear, but it is possible that some analogs active in rodents are not active in primates, in which case primates will become important for preclinical screening of promising contraceptive G&H analogs.
Immunization Against Gonadotropins Wickings and associates, in this symposium, have reported that both passive and active
immunization against FSH can inhibit spermatogenesis. The latter approach achieved its objective without affecting testicular steroid secretion, although use of adjuvant was necessary.
Suppression of FSH Secretion and Action Identification and synthesis of a specific testicular FSH inhibitor (inhibin) might
provide a means of controlling spermatogenesis without affecting androgen-dependent secondary sex organs. In male nonprimates, peptides with inhibin-like properties have been isolated from testicular extracts, testicular lymph, rete testis fluid, semen, and medium used to culture Sertoli cells [review by Setchell, 19801, and other evidence for inhibin derives from human subjects with damaged seminiferous epithelium who show selective elevation of FSH levels [Van Thiel et al, 19711.
Steroid Suppression of Spermatogenesis Various androgens (eg, Danazol, an ethynyltestosterone derivative) inhibit LH secre-
tion, and so secretion of testosterone by Leydig cells, resulting in insufficient local concentration of testosterone to support spermatogenesis [Heller et al, 19701. There is no loss of libido or potentia, but undesirable side effects on lipoprotein metabolism and blood cell formation occur [Heller et al, 1950; Hotchkiss, 1944; Skoglund & Paulson, 1973; Reddy & Rao, 19721. Estrogens and progestins can inhibit spermatogenesis, but they also cause painful
gynecomastia, as well as loss of libido and potentia [Heller et al, 19591. Estrogen or progestin appropriately balanced with androgen could avoid such undesired side effects. Reversible infertility can be induced with such preparations [Coutinho & Melo, 1973; Briggs & Briggs, 19741. Ewing et a1 [ 19781 have described a primate model for the eficient evaluation of efficacy
and safety of steroid combinations, using 20 rhesus monkeys for 50 months to test 30 testosterone-estradiol combinations. The design assumes no carryover of drug effect from one treatment to the next; end points were number of sperm and volume per ejaculate. This experimental model seems a powerful analytical tool, although the observation of seasonal variation in sperm number points to the importance ofhaving a separate control group rather than using each animal as its own control; alternatively, a less seasonal

162 Graham
species such as the stumptail or pigtail monkey (Mucaca speciosa or M. nemestrina) could be selected. Anand Kumar et a1 11980J have administered steroids by intranasal spray to rhesus
monkeys. The intranasal route results in a relatively high concentration of administered steroid in the cerebrospinal fluid and hence in the brain, without raising peripheral levels significantly. Considerable depression oftestosterone levels and azoospermia after 30 pgiday of estradiol, progesterone, or norethisterone was noted. The testicular regres- sion resembled that which follows oral or systemic administration ofestrogens or proges- tins in men, except that the intranasal route required M O O or less of the dosage calculated on a body weight basis. This seems to offer an exciting new approach to male contraception that might avoid the toxicological problems associated with higher steroid doses and systemic effects. However, as with other approaches to male contraception that involve suppression of testosterone levels, a means to maintain the integrity of libido and accessory organs will have to be worked out before application to the human can be envisaged. The competitive testosterone inhibitor cyproterone acetate (CAI can inhibit sper-
matogenesis in rhesus monkeys and man LPrasad et al, 19771; 697 mgikg CA released from Silastic capsules did not alter circulating testosterone levels in rhesus monkeys, but caused reversible oligospermia or azoospermia. CA also appeared to inhibit conversion of testosterone to dihydrostestosterone (the intracellular active androgen) selectively in the caput epidymidis, so reducing caput weight and secretory activity.
Steroid Metabolism The study of testicular steroid metabolism in primates is important for the elucidation
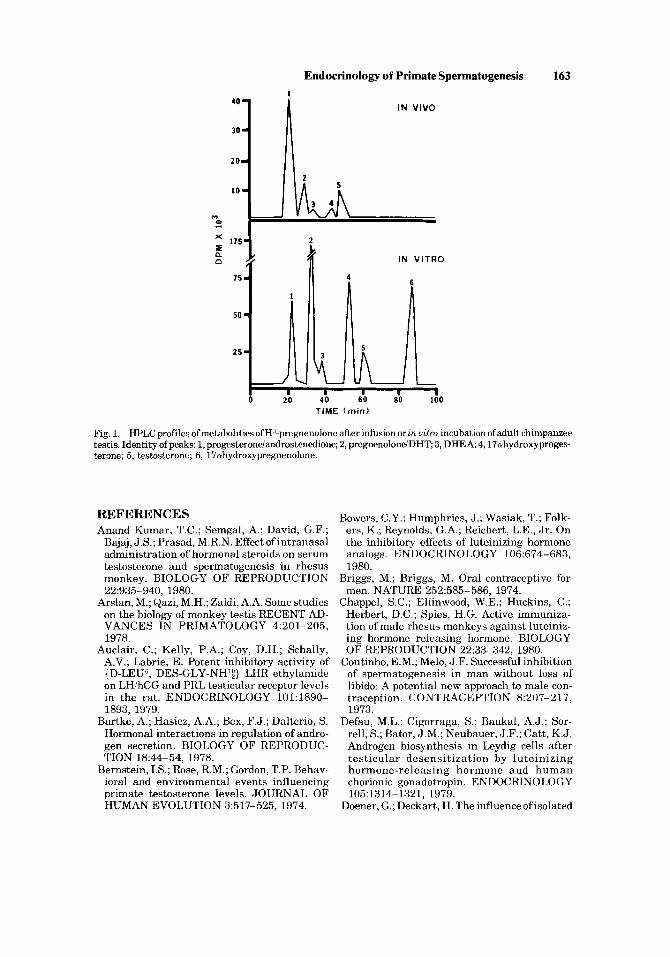
of the mechanism of action of trophic hormones, contraceptive drugs, and toxic com- pounds that affect spermatogenesis by acting on the testis. In collaboration with E. Helton and W.C. Hobson, we have commenced to develop an experimental model in the chimpanzee for this purpose because of general metabolic and endocrine similarity with man, the large testicular vessels, and the well-developed technology for semen collection [Martin e t al, 1977; McCormack, 1971; Gould et al, 19781. Pulse injection of radiolabeled substrate and collection of testiular venous effluent from
one of the relatively large collaterals of the pampiniform plexus of the chimpanzee can permit repeated in vivo study of the same subjects. The fairly long time required to clear the testis of labeled substrate, in this, case pregnenolone, plus the recovery of a variety of labeled metabolites (identified by high pressure liquid chromatography using suitable solvent systems to achieve separation) indicates that significant metabolism occurs under these conditions. Similar metabolic profiles were obtained when the contralateral unperfused testis was incubated with pregnenolone invitro (Fig. 1); these results suggest that both systems generate meaningful data and that the in vivo system can be used repeatedly to study metabolic problems relating to the control of spermatogenesis.
CONCLUSIONS 1. Significant advances have been made in the endocrine control of spermatogenesis in
recent years. 2. Our knowledge is based on relatively few primate species. 3. Variability in endocrine control mechanisms is a significant factor in the selection of
primate models for the study of the endocrinology of spermatogenesis, but the full extent of variability is unknown.
4. Primates have many applications in the development of male contraceptives.
Endocrinology of Primate Spermatogenesis 163 1
IN VlVO 40
30 9 I 20-
2 10 -
el - f X I a n
175
75
50
25
0 20 4 0 - IN VlTRO
n 0 TIME frnin)
Fig. 1. HPLC profiles ofmetabolities of H'-pregnenolone aRer infusion or in uitru incubation of adult, chimpanzee testis. Identity of peaks: 1, progesteroneiandrostenedione; 2, pregneno1ondDHT 3, DHEA; 4,17uhydroxyproges- krone; 5, testosterone; 6, 17ohydroxypregnenolone.
REFERENCES Anand Kumar, T.C.; Semgal, A,; David, G.F.;
Bajaj, J.S.; Prasad, M.R.N. Effect of intranasal administration of hormonal steroids on serum testosterone and spermatogenesis in rhesus monkey. BIOLOGY OF REPRODUCTION
Arslan, M.; Qazi, M.H.; Zaidi, A.A. Some studies 22:935-940, 1980.
on the bioldgy of monkey testis RECENT AD- VANCES IN PRIMATOLOGY 4201-205. 1978.
Auclair, C.; Kelly, P.A.; Coy, D.H.; Schally, A.V.; Labrie, E. Potent inhibitory activity of {D-LEUb, DES-GLY-NH'!} LIlR ethylamide on LH/hCG and PRL testicular receptor levels in the rat. ENDOCRINOLOGY 101:1890- 1893, 1979.
Bartke, A,; Hasiez, A.A.; Bex, F.J.; Dalterio, S. Hormonal interactions in regulation of andro- gen secretion. BIOLOGY OF REPRODUC- TION 18:44-54, 1978.
Bernstein, IS.; Rose, R.M.; Gordon, T.P. Behav- ioral and environmental events influencing primate testosterone levels. JOURNAL OF HUMAN EVOLUTION 3:517-525, 1974.
Bowers, C.Y.; Humphries, J.; Wasiak, T.; Folk- ers, K.; Reynolds, G.A.; Reichert, L.E., Jr. On the inhibitory effects of luteinizing hormone analogs. ENDOCRINOLOGY 106:674-683, 1980.
Briggs, M.; Briggs, M. Oral contraceptive for men. NATURE 252:585-586, 1974.
Chappel, S.C.; Ellinwood, W.E.; Huckins, C.; Herbert, D.C.; Spies, H.G. Active immuniza- tion of male rhesus monkeys against luteiniz- ing hormone releasing hormone. BIOLOGY OF REPRODUCTION 2233-342, 1980.
Coutinho, E.M.; Melo, J.F. Successful inhibition of spermatogenesis in man without loss of libido: A potential new approach to male con- traception. CONTRACEPTION 8:207-217, 1973.
Defau, M.L.; Cigorraga, S.; Baukal, A.J.; Sor- rell, S.; Bator, J .M. ; Neubauer, J.F. ; Catt, K.J. Androgen biosynthesis in Leydig cells after testicular desensitization by luteinizing hormone-releasing hormone and human chorionic gonadotropin. ENDOCRINOLOGY 105:1314-1321, 1979.
Doener, G.; Deckart, H. The influence of isolated

164 Graham
and combined doses of follicle-stimulating Hsueh, A.J.W.; Erickson, G.F. Extrapituitary hormone (FSHj and chorionic gonadotropin action of gonadotropin releasing hormone: Di- (HCG) on testicular function of intact infantile rect inhibition of ovarian steroidogenesis. and juvenile rats. ACTA BTOLOGICA MED- SCIENCE 204:854-855, 1979. ICA GERMATICA 9:271, 1962. Kinzey, W.G. Male reproduction system and
Ewing, L.J.; Adams, R.J.; Cochran, R.C. The spermatogenesis. pp. 85-113 in COMPARA- effects of chemicals on spermatogenesis and TIVE REPRODUCTION OF NON-HUMAN epididymal maturation of spermatozoa: Ex- PRIMATES. E.Z. Hafez, ed. Springfield, I1- perimental principles. pp. 326-343 in ANI- linois, Charles C Thomas, 1971. MAL MODELS FOR RESEARCH ON CON- Levitan, D.; Beitins, I.Z.; Barnes, A,; McArthur, TRACEPTION AND FERTILITY. N.J. Alex- J.W. Insensitivity of Bonnet monkeys to (D- ander, ed. Hagerstown, New York, Harper and Ma6, Des-Gly'O) LHRH ethylamide, a potent Row, 1978. new LHRH analogue in rats and mice. ENDO-
Gosselin, R.E.; Fuller, G.B.; Coy, D.H.; Schally, CRINOLOGY 100:91&922, 1977. A.V.; Hobson, W.C. Inhibition of gonadotropin Louvet, J.P.; Vaitukaitis, J.C. Induction of release in chimpanzees by the LH-RH an- follicle-stimulating hormone (FSH) receptors tagonist (D-Phe', D-Trp', D-Phe"i)-LH-RH. in rat ovaries by estrogen priming. ENDO- PROCEEDINGS O F THE SOCIETY FOR CRINOLOGY 99:758-764,1976. EXPERIMENTAL BIOLOGY AND MEDI- Ludwig, D.J. The effect of androgen on sper- CINE 161:21-24, 1979. matogenesis. ENDOCRINOLOGY 46:453,
Gould, K.G.; Warner, H.; Martin, D.E. Rectal 1950. probe ejaculation of primates. JOURNAL OF MacLeod, J.; Pazianos, A,; Ray, B.S. Restoration MEDICAL PRIMATOLOGY 7:21%222,1978. of human spermatogenesis by menopausal
Graham, C.E.; Gould, K.G.; Collins, D.C.; gonadotropins. LANCET 1:1196, 1964. Preedy, J.R.K. Regulation of gonadotropin re- MacLeod, J.; Pazianos, A.; Ray, B.S. The lease by luteinizing hormone releasing hor- restoration of human spermatogenesis and of mone and estrogen in chimpanzees. ENDO- the reproductive tract with urinary gonado- CRINOLOGY 105:269-275, 1979. tropins following hypophysectomy. FERTIL-
Hansson, V.; Djoseland, D.; Reusch, E.; At- ITY AND STERILITY 17:7, 1966. tramadal, A.; Torgerson, 0. An androgen bind- Maddock, W.O. Antihormone formation com- ing protein in the testis cytosol fraction of adult plicating pituitary gonadotropin therapy in in- rats. Comparison with the androgen binding fertile men 1 . Properties of the antihormone. protein in the epididymis. STEROIDS JOURNAL OF CLINICAL ENDOCRINOL- 21:457-474, 1973. OGY 9213, 1949.
Heller, C.G.; Moore, D.J.; Paulsen, C.A.; Nelson, Maddock, W.O.; Nelson, W.O. The effects of W.; Laudlow, W.M. Effects ofprogesterone and chorionic gonadotropin therapy in infertile synthetic progestins upon the reproductive men: Increased estrogen and 17-ketosteroid physiology of normal men. FEDERATION excretion, gynecomastia, Leydig cell stimula- PROCEEDINGS 18:1057-1065, 1959. tion and seminiferous tubule damage. JOUR-
Heller, C.G.; Morse, H.C.; Su, M.; Rowley, M.J. NAL OF CLINICAL ENDOCRINOLOGY The role of FSH, ICSH and endogenous testos- 12:985, 1952. terone during testicular suppression by Martin, D.E.; Swenson, K.B.; Collins, D.C. Cor- exogenous testosterone in normal men, AD. relation of serum testosterone levels with age VANCES IN EXPERIMENTAL MEDICINE in male chimpanzees. STEROIDS 29:471- 481, AND BIOLOGY 10:249-259, 1970. 1977.
Heller, C.G.; Nelsorl, W.O.; Hill, I.B.; Hender- McCormack, S.A. Plasma testosterone concen- son, E.;Maddock, W.O.; Jungck, E.C.; Paulson, tration and binding in the chimpanzee: Effect C.A.; Mortimore, C.E. Improvement in sper- of age. ENDOCRINOLOGY 89:1171-1177, matogenesis following depression of t h e 1971. human testis with testosterone, FERTILITY Means, A.R.; Vaitukaitis, J.C. Peptide hormone AND STERILITY 1:415, 1950. receptor, specific binding of "-FSH to testis.
Hobson, W.; Fuller, G.B. LH-RH-induced ENDOCRINOLOGY 90:39-46, 1972. gonadotropin release in chimpanzees. BIOL- Paulsen, C.A. Effect of human chorionic OGY OFREPRODUCTION 17294-297,1977. gonadotropin and human menopausal go-
Hodgen, J.K.; H ~ ~ ~ , J.P. Effects of imunha- nadotropin therapy on testicular function. tion against luteinking hormone releasing PP. 491 in GONADOTROPINS. E. Rosenberg, hormone on reproduction of the rhesus man- ed. h S Altos, California, GerOn-X, 1968. key eallithrix jacchus. NATURE 265, N ~ . Prasad, M.R.N.; Arora-Dinakar, R.; Dinakar, N. 5596~746-748,1977. Use of the rhesus monkey in studies on repro-
Hotchkiss, R.S. Effect of massive doses oftestos- terone propionate upon spermatogenesis. JOURNAL OF CLINICAL ENDOCRINOL.
ductive biolo!ZY of the male: Hormonal envi- ronment and e ~ i d i d ~ m a n function. PP. 121- 141 in USE OF NON-HUMAN PRIMATES IN BIOMEDICAL RESEARCH. M.R.N. Prasad; OGY 4:117, 1944.

Endocrinology of Primate Spermatogenesis 165
T.C. Anand Kumar, eds. New Delhi, India, In- Swerdloff, R.S.; Odell, W.D. Some aspects of the dian National Science Academy, 1977. control of secretion in LH and FSH in humans.
Reddy, P.R.K.; Rao, J.M. Reversible antifertil- In GONADOTROPINS. E. Rosenberg, ed. Los ity action of testosterone propionate in human Altos, California, Geron-X, 1Y68. males. CONTRACEPTION 5:295-301, 1972. Van Thiel, F.H.; Sherins, R.J.; Myers, G.H. Jr.;
Rubin, R.T.; Poland, R.E.; Sowers, J.R.; Hersh- DeVita, V.T. Evidence for a specific seminifer- man, J.M. Influence of methyl-TRH-induced ous tubular factor affecting follicle-stimulat- prolactin increase on serum testosterone levels ing hormone secretion in man. JOURNAL OF in normal adult men. JOURNAL OF CLINI- CLINICAL INVESTIGATION 51:1009-1019, CAL ENDOCRINOLOGY AND METABO- 1971. LISM 46:830-833, 1978. Vickery, B.H.; McRae, (2.1. Effects of continuous
Setchell, B.P. Inhibin. RESEARCH IN RE- treatment of male baboons with superagonists PRODUCTION 12:3, 1980. of LHRH. INTERNATIONAL JOURNAL OF
Skoglund, R.D.; Paulson, C.A. Danazol-testos- FERTILITY 25:179-186, 1980a. terone combination: A potentially effective Vickery, B.H.; McRae, G.I. Responses of the means for reversible male contraception, a pre- males of different laboratory species to contin- liminary report, CONTRACEPTION 7:357- uous administration of an LHRH antagonist. 365, 1973. JOURNAL OF ANDROLOGY 1:62, 1980b.
Smith, P.E. Comparative effects of hypophysec- Wickings, E.J.; Nieschlag, E. Seasonality in tomy and therapy on the testis ofmonkeys and endocrine and exocrine testicular function of rats. In LES HORMONES SEXUALIS, I,. the adult rhesus monkey (Macca mulattu) Brouha, ed. Paris, France, 1938. maintained in a controlled laboratory envi-
Steinberger, A.; Heindel, J.J.; Lindsey, N.J.; ronment. INTERNATIONAL JOURNAL OF Elkington, S.H.; Sanborn, B.M.; Steinberger, ANDROLOGY 3:87-104, 1980. E. Isolation and culture ofFSH responsive Ser- Yen, S.C.C.; Rebar, R.; Vandenberg, G.; Nafto- toli cells. ENDOCRINE: RESEARCH COM- lin, F.; Judd, M.; Ehara, Y.; Ryan K.J.; Rivier, MUNICATIONS 2:261-272, 1975. J.; Amoss, M.; Guillemin, P. Clinical studies
Steinberger, E.; Steinberger, A. Testis: Basic with synthetic LRF. pp. 217-229 in CON- and clinical aspects. pp. 144-267 in REPRO- FERENCE ON HYPOTHALAMIC HYPO- DUCTIVE BIOLOGY. H. Balin, S. Glasser, PHYSIOTROPIC HORMONES. C. Gual, E. eds. Amsterdam, Excerpta Medica, 1972. Rosenberg. Excerpta Medica Int. Congr. Ser-
ies 263. 1973.


















![Pict. of Spermatogenesis - 2015 [1]](https://img.pdfslide.us/doc/110x75/563dbb78550346aa9aad780e/pict-of-spermatogenesis-2015-1.jpg)










