Embed Size (px)
Citation preview

Drug Development Research 14: 123-139 (1988)
Electrophysiological and Antiarrhythmic Effects of the K-Channel Opener, BRL 34915, in Cardiac Purkinje Fibers 80 Liu, Faraidoon Golyan, John R. McCullough, and Mario Vassalle
Department of Physiology, State University of New York, Health Science Center, Brooklyn, (8. L., F. G., M. V.) and Research Department, Pharmaceuticals Division, Ciba-Geigy Corp., Summit, NJ (J. R. M.)
ABSTRACT
Liu, B., F. Golyan, J.R. McCullough, and M. Vassalle: Electrophysiological and anti- arrhythmic effects of the K-channel opener, BRL 34915, in cardiac Purkinje fibers. Drug Dev Res 14:123-139, 1988.
The effects of BRL 34915 (6-cyano-3,4-dihydro-2,2-dimethyl-frans-4-[2-oxo-l-pyrrolidyl]- 2H-benzo[b]pyran-3-oI, to be referred to as BRL) on the electromechanical properties of superfused dog and sheep ventricular Purkinje fibers were studied in vitro. At 5 pM, BRL shortened the action potential and decreased contractile force; these effects were greater in dog than in sheep Purkinje fibers. BRL reduced the slope and amplitude of diastolic depolarization measured during interruptions of drive. BRL suppressed spontaneous activ- ity and antagonized the enhancement of automatic discharge induced by norepinephrine (0.5 pM), barium (50 pM), or strophanthidin (0.5 pM). BRL reduced or abolished the oscillatory potentials induced by high [Ca2+], (10.8 mM). It also antagonized the sponta- neous responses in low [K+], (1 mM) and hyperpolarized fibers depolarized in a K-free solution. It is concluded that BRL modifies the action potential and force and has antiar- rhythmic actions as it antagonizes abnormal pacemaker activity in Purkinje fibers by modifying potassium conductance and secondarily reducing intracellular calcium.
Key words: transmembrane potential, contractile force, dog, sheep, potassium, norepineph- rine, strophanthidin, calcium, barium
Received final version April 11, 1988; accepted April 11, 1988.
Address reprint requests to Dr. Mario Vassalle, Department of Physiology, Box 31, State University of New York, Health Science Center, 450 Clarkson Ave., Brooklyn, NY 11203.
Dr. McCullough’s present address is Squihb Institute for Medical Research, Pharmacology Department, P.O. Box 4000, Princeton, NJ 08543.
0 1988 Alan R. Liss, Inc.

124 Liuet al.
INTRODUCTION
The benzopyran derivative BRL 3491 5 (6-cyano-3,4-dihydro-2,2-dimethyl-trans-4-[2- oxo-l-pyrrolidyl[-2H-benzo[b]pyran-3-ol, referred to as BRL) has an antihypertensive action in intact animals [Buckingham et al., 1984, 1986; Ashwood et al., 19861 and in patients [VandenBurg et al., 19861. This antihypertensive action is related to the inhibition of spontaneous mechanical [Ruckingham et al., 19841 and electrical [Hamilton et al., 19861 activity of vascular smooth muscle. The mechanism of this action appears to be an increased potassium conductance since, in different smooth muscles, BRL hyperpolarizes the membrane potential to a value close to the calculated potassium equilibrium potential and increases the efflux of radioactive rubidium [Hamilton et al., 1986; Weir and Weston, 1986b; Allen et al., 1986).
An increase in potassium conductance by BRL makes it a suitable candidate as an antiarrhythmic agent. The aim of the present experiments was to investigate the actions of BRL on the electrical and mechanical activity of driven and spontaneous Purkinje fibers from sheep and dog hearts. BRI, was tested under several conditions that modify potassium conductance and, therefore, spontaneous activity. Also, BRL actions were studied in conditions that inhibit the activity of the Na-K pump, since some of drug actions could be explained also by the stimulation of an electrogenic Na-K pump. Finally, BRL cffects were evaluated on abnormal automaticity at different levels of rnembranc potential. A preliminary report of some of the data presented here has appeared in abstract form [Liu et al., 19871.
MATERIALS AND METHODS
Sheep (30-69 kg) and dogs (15-30 kg) of either sex were anesthetized with sodium pentobarbital (30 mg/kg, JV). The heart was quickly excised through a left intercostal incision and strands of Purkinje fibers were removed from the ventricles and perfused in a tissue bath with oxygenated (97% O2 and 3 8 C02) Tyrode’s solution at 37°C. The composition of the Tyrode’s solution in mM was NaCl 137; KCI 4.0; NaHCO, 11.9; NaH2P0, 0.45; MgC12 1.05; CaC12 2.7; dextrose 5.5. The [Ca”], was increased to 16.2 mM in some experiments, as will be specified. The extra calcium chloride was added as a powder to the oxygenated freshly prepared Tyrode’s solution.
The fibers were stimulated by means of two steel pins connected to a Tektronix stimulator (model 62) via Grass stimulation isolation unit (SIU 4678). The duration of the stimuli was 1-3 msec and the voltage was 50-80% above the threshold. The fibers were driven at 60 Hz and equilibrated for a minimum of 1 hr before the experiments began. During the different tests, the drive was temporarily interrupted until the membrane potential reached a steady value or spontaneous activity developed. BRL was tested under these various conditions.
One end of the fiber bundle was immobilized with one of the steel pins used as stimulating electrodes, whereas the other end was tied by means of a short silk thread to a rigid rod attached to a force transducer (Grass IT0 3C). The other steel pin was positioned near the preparation. The steel pin that immobilized one end of the strand pressed against the bottom of the bath under tension and prevented any movement of that end. The rod to which the other end of the strand was attached did not touch the bottom of the bath and kept the strand slightly lifted in the perfusing solution. The preparations were stretched by approximately one-third of their resting length, as sheep Purkinje fibers do not tolerate large degrees of stretch well. In the different tests, the changes in force were compared with the control value prior to the test, thus providing a useful correlate to the electrical measurements. Transmembrane potentials were recorded by means of glass microelectrodes filled with 3 M KCl (15-30 M a ) . Both electrical and mechanical events were displayed on a Tektronix Model 5 1 1 1 oscilloscope and were

Actions of BRL 34915 in Purkinje Fibers 125
BRL CONTR. .
Fig. 1. Effects of BRL on action potential, twitch, and diastolic depolarization in dog cardiac Purkinje fibers. In the left and right panels, the top trace shows the action potential and the bottom trace the twitch; thc middle panels show the lower part of the action potential at higher gain. In A, B and C, the first panel and the upper middle panel show the control traces in Tyrode’s solution; the lower middle panel and the last panel show the traces recorded in the presence of different concentrations of BRL (0.05 pM in A; 0.5 pM in B; and 5 p,M in C). Voltage, time, and force calibrations are next to the respective traces.
recorded on paper by means of a chart recorder (Gould Brush 2400). Some of the traces were also recorded on a FM tape recorder (Store 4, Lockheed Electronics, Inc.).
BRL 34915 (Ciba-Geigy) was dissolved in polyethylene glycol 400 (Fluka Co.). The final dilution of the solvent was 0.1% or less and did not have any apparent effect on the electrical or mechanical activity of the Purkinje fibers when tested alone. Strophanthidin and barium were obtained from Sigma Chemical Co. The duration of the action potential was measured at 50% (APD,,) and 100% (APDloo) repolarization: when not otherwise specified, the action potential duration given in text is APDloo. In most experiments, the lower part of the action potential was recorded also at a higher gain. The slope and the amplitude of diastolic depolarization were measured at the beginning and at the end, respectively, of the drive interruption. The results are expressed as mean 2 SE. A paired Student’s t test was used and a P < 0.05 was considered significant. In the test, n (number of experiments) indicates the number of animals.
RESULTS Effects of BRL as a Function of the Dose in Dog and Sheep Purkinje Fibers
The effects of BRL on the action potential, twitch, and diastolic depolarization (DD) in dog Purkinje fibers are shown in Figure 1 . In Figure 1 A, a low concentration of BRL (0.05 FM) had little effect on the recorded parameters. In Figure lB, an intermediate concentration (0.5 pM) substantially decreased the action potential duration (APDSo -60.0%, APD,w -31.8%) and the twitch amplitude (-83.3%). During interruption of the drive (middle panel), the slope and amplitude of DD were reduced compared with control (-23.9% and -28.3%, respectively). In Figure IC, a high concentration of BRL (5 pM) shortened the action potential markedly (APDSo -97.l%, APDloo -59.1%) and abolished the twitch. Also, during interruption of the drive, DD was abolished. In Figure 1B and C, the control panels show that
126 Liu et al.
0
-20
rn W u 5 -40 X 0 c z W u -fie a W P
-8C
-1oc
ERt concsntratlon. yM
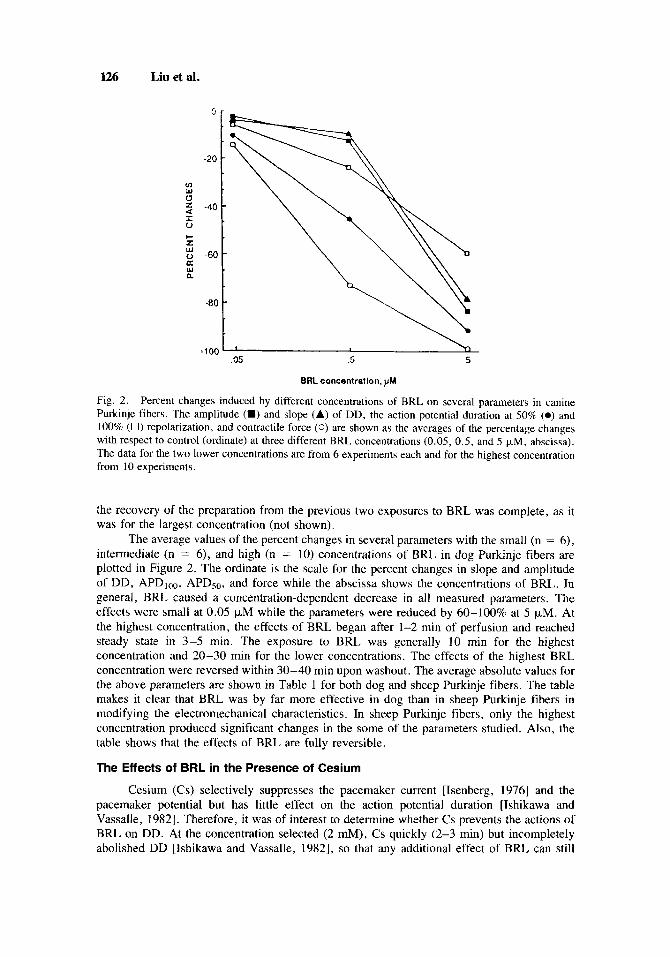
Fig. 2. Percent changes induced by different concentrations of BRL on several parameters in canine Purkinje fibers. The amplitude (M) and slope (A) of DD, the action potential duration at 50% (0) and 100% (0) repolarization, and contractile force (0) are shown as thc avcrages of the percentage changes with respect to control (ordinate) at three different BRL concentrations (0.05, 0.5, and 5 kM, abscissa). The data for the two lower concentrations are from 6 experiments each and for the highest concentration from 10 experiments.
the recovery of the preparation from the previous two exposures to BRL was complete, as it was for the largest concentration (not shown).
The average values of the percent changes in several parameters with the small (n = 6), intermediate (n = 6), and high (n = 10) concentrations of BRL in dog Purkinje fibers are plotted in Figure 2. The ordinate is the scale for the percent changes in slope and amplitude of DD, APD,,, APDSo, and force while the abscissa shows the concentrations of BRL. In general, BRL caused a concentration-dependent decrease in all measured parameters. The effects were small at 0.05 p.M while the parameters were reduced by 60-100% at 5 pM. At the highest concentration, the effects of BRL began after 1-2 min of perfusion and reached steady state in 3-5 min. The exposure to BRL was generally 10 min for the highest concentration and 20-30 min for the lower concentrations. The effects of the highest BRL concentration were reversed within 30-40 min upon washout. The average absolute values for the above parameters are shown in Table 1 for both dog and sheep Purkinje fibers. The table makes it clear that BRL was by far more effective in dog than in sheep Purkinje fibers in modifying the electromechanical characteristics. In sheep Purkinje fibers, only the highest concentration produced significant changes in the some of the parameters studied. Also, the table shows that the effects of BRL are fully reversible.
The Effects of BRL in the Presence of Cesium
Cesium (Cs) selectively suppresses the pacemaker current [Isenberg, 19763 and the pacemaker potential but has little effect on the action potential duration [Ishikawa and Vassalle, 19821. Therefore, it was of interest to determine whether Cs prevents the actions of BRL on DD. At the concentration selected (2 mM), Cs quickly (2-3 min) but incompletely abolished DD [Ishikawa and Vassalle, 19821, so that any additional effect of BRL can still
Actions of BRC 34915 in Purkinje Fibers 127
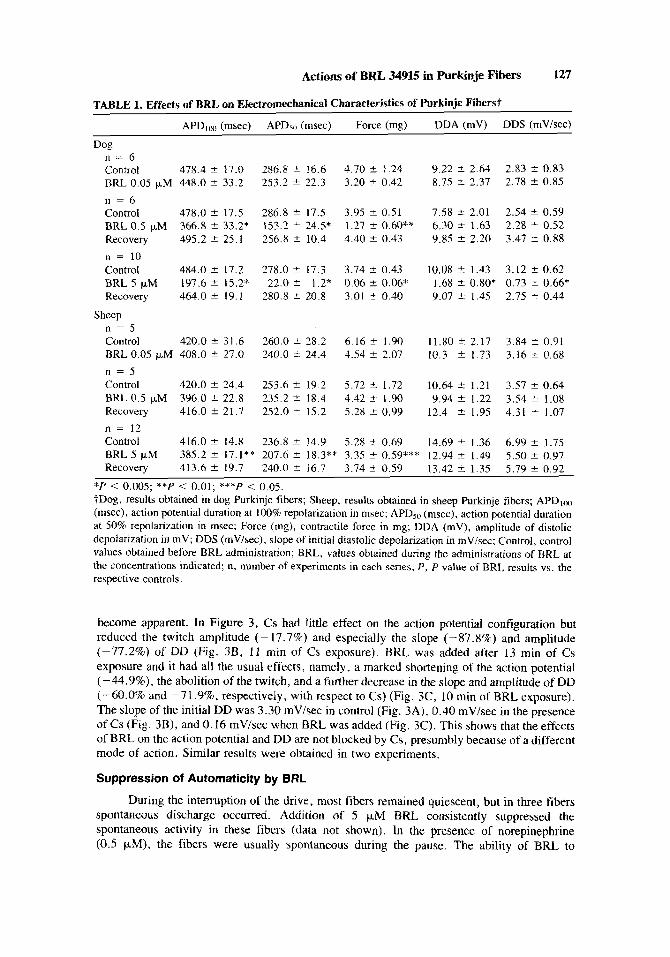
TABLE I. Effects of BRL on Electromechanical Characteristics of Purkinje Fiberst
APDlm (msec) A P h (msec) Force (mg) DDA (mV) DDS (mVisec)
Dog n = 6 Control 478.4 2 17.0 286.8 2 16.6 4.70 k 1.24 y.22 s 2.64 2.83 2 0.83 BRL 0.05 p M 448.0 5 33.2 253.2 t 22.3 3.20 ? 0.42 8.75 k 2.37 2.78 * 0.85 n = 6 Control 478.0 c 17.5 286.8 2 17.5 3.95 ? 0.51 7.58 ? 2.01 2.54 * 0.59 BRL0.5 pM 366.8 * 33.2* 153.2 S_ 24.5* 1.27 .t 0.60** 6.30 S 1.63 2.28 2 0.52 Recovery 495.2 2 25.1 256.8 * 10.4 4.40 c 0.43 9.85 2 2.20 3.47 t 0.88 n = 10 Control 484.0 2 17.2 278.0 2 17.3 3.74 & 0.43 10.08 2 1.43 3.12 2 0.62 BRL 5 pM 197.6 2 15.2* 22.0 * 1.2" 0.06 & 0.06" 1.68 2 0.80* 0.73 2 0.66* Recove3 464.0 * 19.1 280.8 2 20.8 3.01 ? 0.40 9.07 2 1.45 2.75 ? 0.44
n = 5 Control 420.0 * 31.6 260.0 2 28.2 6.16 ? 1.90 11.80 ? 2.17 3.84 * 0.91 BRL0.05 p,M 408.0 * 27.0 240.0 S 24.4 4.54 k 2.07 10.3 ? 1.73 3.16 t 0.68 n = 5 Control 420.0 * 24.4 253.6 * 19.2 5.72 5 1.72 10.64 2 1.21 3.57 * 0.64 BRL0.5 pM 396.0 2 22.8 235.2 2 18.4 4.42 i 1.90 9.94 2 1.22 3.54 ir 1.08 Recovery 416.0 * 21.7 252.0 * 15.2 5.28 f 0.99 12.4 2 1.95 4.31 k 1.07 ri = 12 Control 416.0 2 14.8 236.8 2 14.9 5.28 c 0.69 14.69 k 1.36 6.99 a 1.75 BRL 5 pM 385.2 2 17.1** 207.6 2 18.3** 3.35 ? 0.59*** 12.94 2 1.49 5.50 2 0.97 Recovery 413.6 2 19.7 240.0 * 16.7 3.74 i 0.59 13.42 2 1.35 5.79 * 0.92
*P < 0.005; **P < 0.01; ***P < 0.05. ?Dog, results obtained in dog Purkinje fibers; Sheep, results obtained in sheep Purkinje fibers; APDIoo (msec), action potential duration at 100% repolarization in msec; APDso (msec), action potential duration at 50% repolarization in msec; Force (mg), contractile force in mg; DDA (mv), amplitude of distolic depolarization in mV; DDS (mV/sec), slope of initial diastolic depolarization in mVisec; Control, control values obtained before BRL administration; BKL, values obtained during the administrations of RRL at the concentrations indicated; n, number of experiments in each series; P , P value of BRL results vs. the respective controls.
Shecp
become apparent. In Figure 3, Cs had little effect on the action potential configuration bur reduced the twitch amplitude (- 17.7%) and especially the slope (-87.8%) and amplitude (-77.2%) of DD (Fig. 3B, 1 1 min of CS exposure). BRL was added after 13 min of Cs exposure and it had all the usual effects, namely, a marked shortening of the action potential (-44.9%), the abolition of the twitch, and a further decrease in the slope and amplitude of DD (-60.0% and -71.9%, respectively, with respect to Cs) (Fig. 3C, 10 min of BRL exposure). The slope of the initial DD was 3.30 mV/sec in control (Fig. 3A), 0.40 mVisec in the presence of Cs (Fig. 3Bj, and 0.16 mV/sec when BRL was added (Fig. 3Cj. This shows that the effects of BRL on the action potential and DD are not blocked by Cs, presurnbly because of a different mode of action. Similar results were obtained in two experiments.
Suppression of Automaticity by BRL
During the interruption of the drive, most fibers remained quiescent, but in three fibers spontaneous discharge occurred. Addition of 5 FM BRL consistently suppressed the spontaneous activity in these fibers (data not shown). In the presence of norepinephrine (0.5 pM), the fibers were usually spontaneous during the pause. The ability of BRL to

128 Liuet al.
* I CONTR
- - I sec 10 sec
C S , 2 m M
+ BRL 5pM c I
Fig. 3. The effects of BRL are not prevented by cesium. In the left panels, the top trace shows the action potential and the bottom trace the twitch; in the right panels, the top trace shows the lower part of the action potential at higher gain and the bottom trace the twitch. The traces were recorded in control (A), at the 1 lth min of exposure to 2 mM cesium (B) and at the 10th min of exposure to Cs plus BRL (5 pM) (C). ‘The slope of the initial diastolic depolarization is extrapolated by the dashed lines. Force calibration is the same for all panels. The voltage Calibration for the left panels is shown next to the left A panel; that for the right panels is shown next to the right A panel.
antagonize the norepinephrine-enhanced DD is illustrated in Figure 4. In Figure 4A, the action potential and twitch (left panel) as well as DD at higher gain (right panel) are shown in control. In figure 4B, norepinephrine decreased the duration of action potential (APDlm -3.3%) and twitch (- 12.5%) and increased contractile force (+90.0%). During the pause, a steepened DD initiated spontaneous discharge. In Figure 4C (a continuation of Fig. 4B), BRL quickly (within 2.2 min) slowed the spontaneous rhythm as the threshold potential for excitation shifted gradually toward less negative values. When the potential failed to reach the threshold, during the ensuing quiescence the membrane actually became slightly more negative (2.4 mV). During the recovery from BRL, the membrane potential became less negative (by 14.5 mV) until spontaneous activity resumed.
BRL stopped the norepinephrine-induced automaticity in five out of eight experiments in dog (n = 8) or sheep (n = 8) Purkinje fibers. Even when it failed to stop the spontaneous discharge, BRL shortened the action potential and slowed the spontaneous rate by 22.8 ? 5.4%.
Actions of BRL in the Presence of Barium or Low [Kl0
If BRL acts by increasing potassium conductance, then it may also be effective when the spontaneous activity is due to a decrease in gK. In a first approach, BRL was tested in the presence of barium (Ba), which decreases g,, [see Di Francesco, 19811. In Figure 5A, the action potential and twitch (left panel) and DD at high gain (right panel) are shown in control. In Figure 5B, 50 pM Ba increased the action potential duration (+29.1%), and during the

Actions of BRL 34915 in Purkinje Fibers 129
NE O . 5 p M ' h
u 60 sec
I 1OOsec I
Fig, 4. Suppression of norepinehrine-induced automaticity by BRL. The action potential (top trace) and the twitch (bottom trace) are shown in the left A and B panels. The lower part of the action potential at higher gain is shown in the remainder of the traces. The traces were recorded in control (A), in the presence of norepinephrine (0.5 kM, B) and of norepinephrine plus BRL (5 pM, C). The voltage calibration next to the left A panel applies to the left A and B panels; the voltage calibration next to the right A panel applies to the remainder of the traces. The last part of the second B panel was recorded at a reduced speed, as indicated by the line labelled 60 sec. The C panel is a continuation of B: between the two dournward arrows, the recording speed was further reduced as indicated by the 100 sec time calibration. In C, the interruption between the traces was 358 sec
pause (middle panel) the slope and amplitude of DD were increased (+ 118.7% and + 38.5%, respectively), initiating spontaneous discharge. At the end of the pause, the electrical drive was resumed and 5 p M BRL added to the Ba solution. In Figure 5C, the left panel shows that BRL markedly shortened the action potential (-39.6%) and abolished the twitch (as it does in the absence of barium). When the drive was interrupted again (middle panel), the DD slope (-28.6%) and amplitude (-47.5%) were reduced so that threshold was not attained and the preparation remained quiescent as long as the stimuli were withheld. In Figure 5D, the removal of BRL restored the patterns seen in the presence of Ba, namely, a longer action potential (+ 44.4%, right panel) and spontaneous activity during the pause as DD amplitude was increased by 102.4% (middle panel). In four experiments, Ba consistently induced spontane- ous activity during the pause and BRL consistently abolished it.
Since lowering [K], decreases potassium movements across the sarcolemma [Carmeliet, 19611 and increases membrane resistance [Vassalle, 19651, in another approach [K], was decreased to 1 mM. In Figure 6, in the low K solution (second panel in A), the preparation became spontaneously active, and contractile force increased ( + 87.5%). While the fiber was discharging spontaneously, the administration of BRL ( 5 p,M) gradually increased the maximum diastolic potential (7.3 mV). Within a few minutes, BRL slowed and stopped the spontaneous activity. During the ensuing quiescence, the membrane potential continued to become more negative (by 5.1 mV). During the washout of BRL, there was prompt reversal of these effects: the membrane potential rapidly became less negative, and the spontaneous activity resumed within 7 min.
In Figure 6B, further decreasing the potassium conductance by addition of barium (50 pM) to the low K solution, increased the rate of discharge (+75.0%) and contractile force

130 Liuet al.
CONTR.
Fig. 5 . BRL suppresses barium-induced automaticity. The action potential (top trace) and the twitch (bottom trace) are shown in the left and right panels; the lower part of the action potential at higher gain is shown in the middle panels. The traces are shown in control (A, first two panels, CONTR.), in the presence of Ba (B, Ba 50 @), in the presence of Ba plus 5 pM BRL (C, left bottom and third middle panel) and during recovery in Ba alone (D, Ba RECOV.). The voltage and force calibration in the left A panel apply to the left and right panels; the voltage calibration for the middle panels is shown next to the top middle panel. The time calibrations for part of the traces recorded at lower speed are indicated in the middle panels by the numbers next to the lines terminating with arrows.
(+73.1%). Administration of BRL again shifted the maximum diastolic potential to more negative values (7.0 mV) and reduced the amplitude of DD. The membrane potential failed to reach the threshold and fiber became quiescent. The effects of BRL were quickly reversible, and, in addition, the membrane potential declined transiently to less negative values than prior to the BRL administration. BRL stopped the spontaneous activity in seven experiments in 1 mM K and in two experiments in 1 mM K plus 50 pM Ba.
The Effects of BRL in the Presence of an Increased K Conductance
In high [Kf],, potassium conductance increases and, therefore, the membrane potential is fairly close to the potassium equilibrium potential [Vassalle, 1970; Baumgarten and Fozzard, 19861. Under conditions of increased potassium conductance, BRL may shorten to a smaller extent the action potential.
In Figure 7A, 5 pM BRL still shifted the plateau to more negative values ( I 8.7 mV), shortened the action potential (- 16.7%), and decreased the twitch ( - - 5 O % ) , but these actions were not as pronounced as in Tyrode’s solution. In figure 7B, adding 1 pM norepinephrine to the high K solution shifted the plateau to less negative values (25.0 mV) and increased contractile force (+ 750.0%), presumably by increasing the slow inward current. BRL ( 5 pM) had little effect on the action potential and did not decrease the twitch amplitude (Fig. 7A, middle panel). These results suggest that BRL does not decrease the norepinephrine-enhanced calcium entry and modifies the action potential less when potassium conductance is already high.
If the smaller effect of BRL in high [K’], is related to the fact that potassium conductance is already increased, then the effects of BRL might be restored even in high [K ’ 1, if potassium conductance is decreased by Ba. In Figure 7C, adding 50 pM Ba to the high K solution prolonged the action potential ( + 38.7%) while increasing the twitch amplitude (+ 70.0%) (Fig. 7C, first panel). In the presence of Ba, 5 pM BRL exerted its usual marked effects on both the action potential duration (-47.9%) and twitch amplitude (- 100%) (Fig.

Actions of BRL 34915 in Purkinje Fibers 131
Fig. 6. BRL counteracts the effects of low [K+],. The action potentials (top trace) and the twitch (bottom trace) are shown at different speeds. The traces are shown in control (first A panel) and in the presence of low [K'], ( 1 mM) (remainder of the traces, fiber spontaneously active and not driven). The periods during which BRL and Ba were administered are indicated below the respective traces. The time calibration for the left A panel is indicated underneath that panel: that for the remainder of the traces is indicated above the B panel (40 sec). During quiescence, the recording speed was reduced as indicated by the numbers next to the lines ending with the downward arrows.
7C, middle panel), followed by full recovery during the BRL washout (Fig. 7C, last panel). It should be noted that the large stimulus needed to excite the fibers depolarized in high [K++I, caused an artifact that obscured the beginning of the slow response, but the separation between stimulus artifact and slow response can be seen in Figure 7C. Similar results were obtained in four experiments.
The Effects of BRL on Calcium-Induced Oscillatory Potentials
Under conditions of calcium overload, Purkinje fibers develop an oscillatory potential (V,,), which can lead to the onset of abnormal rhythms. BRL could influence V,, in more than one way and this was tested in fibers exposed to high [Ca*+],.
In Figure 8A (control), during the interruption of the drive there were three spontaneous beats. In Figure 8B, high [Ca2+], shortened the action potential (-16.7%) and increased contractile force ( + 207.2%). During the pause, the initial DD was markedly steepened by a V,, (+ 190.0%) that was synchronous with an aftercontraction (Fig. 8B, arrows). The insert (Fig. 8B) shows V,, and aftercontraction at a higher gain and a faster time base: the dashed lines emphasize the shift in the traces induced by the V,, (4.3 niV) and aftercontraction (0.5 mg). Because of a less negative threshold in high [Ca2+], [Weidmann, 19551, the spontaneous activity was no longer present in spite of the larger DD. In Figure 8C, BRL shortened the action potential (-44.0%) and decreased force (-60.5%) as it does at normal [Ca2+],. The slope and amplitude of DD were considerably decreased (-69.8% and -61.5%, respectively) when measured after the same delay during the pause, and neither V,, nor aftercontraction were any longer present. BRL abolished V,, in three sheep and in two dog Purkinje fiber experiments, while having little effect in one sheep Purkinje fiber experiment.

132 Liu et al.
1-27 m M K-
A I I I
L -L .A- I [ + N E IpM
Fig. 7. Effects of BRL in high [K' 1,. In each panel, the top trace is the action potential and the bottom the twitch. All the traces shown in figure were recorded in high [K+], (27 mM). Norepinephrine ( 1 +M) was added in B and Ba (SO FM) in C. In A, B and C, the traces are shown in control (left panel), in the presence of 5 +M BRL (middle panel) and during recovery in high [K'], (right panel).
The Effects of BRL and the Sodium Pump
The shortening of the action potential by BRL could be brought about by different mechanisms such as an increased activity of an electrogenic Na-K pump. For this reason BRL was tested also under conditions that inhibit the Na-K pump.
Jn Figure 9, the control traces are shown in A and those recorded in the presence of strophanthidin (0.5 pM) in B. At the time strophanthidin had increased contractile force substantially ( + 281.6%), the drive was interrupted, and the preparation became spontane- ously active (second Fig. 9B panel) in contrast to control (cf. second Fig. 9A panel). When the spontaneous rhythm had stabilized, the perfusion of BRL (5 pM) increased the maximum diastolic potential (1 .O mV) and rapidly led to the cessation of spontaneous activity. During the quiescence, the membrane potential became more negative (5.1 mV). On washout of BRL, the resting potential rapidly decreased to the threshold potential and the spontaneous activity resumed.
Another way to inhibit the Na-K pump is to perfuse the fibers in zero [K+],. In Figure 10, exposure of the preparation to zero [K'l0 led to spontaneous activity and eventual quiescence at a depolarized level ( -33.8 mV, second Fig. 10A panel). In the first Figure 10B panel, addition of BRL (5 pM) to the zero K solution caused a marked repolarization (43.8 mV) to -77.6 mV, a value close to the resting potential (-87.5 mV). As usual, the effects of BRL quickly disappeared on washout of the drug. The last Figure 10B panel shows the recovery of the action potential and of twitch in Tyrode's solution.
Thus, varying degrees of Na-K pump inhibition do not prevent the usual antiarrhythmic and hyperpolarizing action of BKL. In fact, in zero [K+], BRL hyperpolarized even more than in Tyrode's solution, because in zero [K+], the potassium conductance is less and the difference between the membrane potential and the potassium equilibrium potential is greater.

Actions of BRL 34915 in Purkinje Fibers 133
A CONTR
L
10 sec
- I 5ec
I 4 sec
Fig. 8. Effects of BRL on the calcium-induced oscillatory potential. The action potential (top trace) and the twitch (bottom trace) are shown in the left panels. The lower part of the action potential recorded at higher gain (top trace) and the twitch (bottom trace) are shown in the right panels. The traces were recorded in control (A), in the presence of high [Ca2'], (16.2 mM, B) and of high [Ca2'], plus BRL (5 pM, C). In B, the arrows indicate the oscillatory potential and the aftercontraction, which are also shown at higher gain and faster time base in the inset. The voltage and time calibrations for the left panels are in the top and bottom left panels, respectively; those for the right A and B panels are in the top and middle right panels, respectively. Note that the right C panel was recorded at a faster speed than the corresponding right panels in A and B.
Similar results were obtained in three experiments with strophanthidin and in four experiments with zero [K+],.
DISCUSSION
The present results show that in cardiac Purkinje fibers, BRL 34915 1) shortens the action potential duration and reduces the slope as well as the amplitude of the pacemaker potential; 2 ) reduces contractile force, probably through the shortening of the action potential; 3) acts promptly but is more effective in the dog than sheep tissue; 4) is completely reversible in its actions; 5 ) acts on active membranes by accelerating repolarization and on resting membranes by causing hyperpolarization; 6) overcomes the actions of agents (barium) or procedures (zero [K+],) that decrease potassium conductance; 7) has the least action when potassium conductance is high (high [K'],), but even then its action can be restored by decreasing potassium conductance by means of barium; 8) acts also in the presence of inhibition of the Na-K pump (strophanthidin, zero [K+]"); and 9) inhibits normal automaticity, barium- and norepinephrine-induced automaticity, and calcium overload-induced V,, .
It is concluded that in Purkinje fibers BRL markedly modifies the action potential as well

134 Liu et al.
- U 40 5%
I sec
Fig. 9. Abolition of strophanthidin-induced automaticity by BRL. The action potential (top trace) and the twitch (bottom trace) are shown in the left panels; the lower part of the action potential at higher gain (top trace) and the twitch (bottom trace) arc shown in the tight panels. The traces were recorded in control (A) and in the presence of strophanthidin (0.5 pM) and of strophanthidin plus BRI, (B). In the second B panel, BRL was perfused during the period indicated below the traces. The voltage and time calibrations for the left panels are next to those panels; those for the right panels are in the right A panel. In the second B panel, the arrow indicates when the recording speed was reduced (see calibration at the end of the traces).
U 20sec
RECOV
40sec n i
5pM 1 Fig. 10. Reversal by BRL of depolarization caused by zero [K+],. The traces were recorded in control (left A panel), in the presence of zero [K+], (right A panel and left B panel) and during recovery (RECOV., right B panel). BRL (5 pM) was administered as indicated below the trace in B. Voltage calibration in the right A panel applies to all panels. The time calibration for the left A and right B panels is below the left A panel; that for the right A and left B panel is indicated above the left B panel. In A, the recording speed was reduced while the preparation was becoming quiescent as indicated by the line labelled 120 sec.
as DD by increasing potassium conductance, and thus may act as an effective antiarrhythmic agent.
The Effects of BRL on the Action Potential of Driven Fibers
BRL shortened the action potential of ventricular Purkinje fibers from sheep and dog hearts, but this effect was more pronounced and more consistent in the dog than in the sheep Purkinje fibers (see Table 1). This indicates that the drug can have a degree of species

Actions of BRL 34915 in Purkinje Fibers 135
dependence. However, in both species the action was prompt (it developed in a few minutes) and was completely reversible on washout. BRL also shortens the action potential in other tissues such as guinea pig papillary muscle [Cain and Metzler, 19851, but this action is less pronounced than in Purkinje fibers. In guinea pig papillary muscle partially depolarized in 22 mM K, BRL shortened the action potential but did not affect the action potential amplitude or the resting potential [Scholtysik, 19871. Sotalol prolongs the action potential [Strauss et a]., 1970) by decreasing the potassium current [Carmeliet, 1985). BRL shortened the action potential even in the presence of Sotalol [Scholtysik, 19871. We also found that BRL shortens the action potential of ventricular myocardial cells of the dog (unpublished experiments).
Mechanism of Action of BRL
It has been proposed that BRL acts in different tissues by increasing potassium conductance. Thus, BRL hyperpolarizes the smooth muscle membrane by shifting, in a concentration-dependent manner, the membrane potential toward the calculated potassium equilibrium potential [Hamilton et al., 1986; Weir and Weston, 1986bl. This conclusion is supported by the fact that BRL increases “Rb efflux (used as a marker of K movements) in the rat portal vein [Hamilton et al., 1986; Weir and Weston, 1986b], the guinea pig taenia caeci [Weir and Weston, 1986a1, and the guinea pig trachealis [Allen et al., 1986; Weir and Weston, 1986bl. Both the suppression of the spontaneous activity and the increase in Rb efflux were also found in the guinea pig portal vein [Quast, 19871. However, it should be added that the concentration of BRL necessary to inhibit the spontaneous activity was found to be 5 to -100 times lower (depending on the experimental conditions) than those required for the increase in Rb efflux, implying that the two effects may not be related [Quast, 1987; see also Weir and Weston, 1986b]. An alternative possibility is that the inhibition of spontaneous activity occurs because low concentrations of BRL more easily affect the pacemaker cells of these tissues [Quast, 19871. Recently, it has been suggested that the relaxant action of BRL on rat uterine smooth muscle results from an action on pacemaker cells since, although spike production was inhibited, the hyperpolarization was only slight and Rb efflux was not increased [Hollings- worth et al., 19871.
In healthy human volunteers [Fox et al., 19871 and in hypertensive patients [Vanden- Burg et al., 19861, BRL increased the heart rate. seemingly in contrast with the notion that the potassium conductance is increased. However, this is not unexpected since the blood pressure decreased, thereby activating reflex adjustments. In fact, BRL induces tachycardia also in intact rats, cats, and dogs, but the rate increase is reflex in nature since it is eliminated by propranolol (while the hypotensive action persisted) [Buckingham et al., 19861.
The Potassium Conductance involved in the Action of BRL
There are several potassium conductances in cardiac tissues. Thus, there is a voltage- dependent background potassium conductance ( g K I ) , a time- and voltage-dependent conduc- tance activated on depolarization (gJ, a conductance activated by calcium ions [lsenberg, 1977; Siegelbaum and Tsien, 19801, an apamin-sensitive conductance [see Weir and Weston, 1986a1, an adenosine triphosphate-dependent conductance [Noma, 19831, an acetylcholine- dependent conductance [Sakmann et al., 19831, an Na-dependent conductance [Kameyana et al., 19841, and that associated with the pacemaker current [Vassalle, 1966; Noble and Tsien, 19681. It should be added that, on the basis of experiments with cesium and barium, it has been proposed that the pacemaker potential in Purkinje fibers is caused by an inward current carried mostly by sodium ions [If, see Di Francesco, 19811, rather than by a decline in an outward current carried by potassium ions. It is true that cesium usually blocks potassium (and not sodium) currents and that barium also induces a pacemaker potential in myocardial fibers [see Imoto et al., 19871. However, in the present context, the knowledge of the ionic mechanism

136 Liuet al.
of the pacemaker current is not essential in view of the fact that BRL is shown not to act by modifying the pacemaker current (see below).
Some of these conductances can be readily ruled out. Since the effect of BRL on 86Rb efflux is not influenced by slow-channel blockers, apparently the effect is not mediated through a K conductance determined by an increased cellular calcium [Quast, 19871. In the present experiments, the modification of the action potential and of force are compatible with a decrease (not an increase) in calcium influx and, therefore, it is unlikely that BRL effects are duc to an increase in a calcium-dependent potassium conductance. Furthermore, an increase in [Ca’], certainly increases cellular calcium but does diminish the slope or amplitude DD. RRI, caused relaxation and an increase in “Rb efflux in the guinea pig taenia caeci. These two effectb were not blocked by apamin (in contrast to those of norepinephrine), suggesting that the effects of BRL are not mediated by the opening of apamin-sensitive potassium channels activated at membrane potential more negative than the resting potential [Weir and Weston, 1986al. Apamin also failed to modify the relaxant action of BRL in the isolated trachealis. Acetylcholine (ACh) has little effect on the action potential of‘ Purkinje fibers [see Watanabe, 19831, and there is no reason to expect that BRL depletes adenosine hiphosphate (ATP) or increases intracellular sodium (see below). Also, BRL does not act by modifying the pacemaker current since this current contributes very little to the action potential duration, presumably because of its inward-going rectification [Noble and Tsien, 19681. Thus, Cs modifies the pacemaker current but does not shorten the action potential, whereas BRL shortened the action potential in the absence and presence of Cs. In addition, BRL also shortens the action potential of ventricular muscle fibers that do not have a pacemaker current. Therefore, it is unlikely that BRL acts by increasing any of these conductances.
In isolated guinea pig ventricular myocytes, BRL hyperpolarizes the resting potential but partially inhibits gK1 [Conder and McCullough, 1987; McCullough et al., 19871. Relatively little effect of BKL on resting potential has been reported in multicellular ventricular preparations of both the guinea pig [Cain and Metzler, 1985; Scholtysik, 19871 and the dog (unpublished observations). The effects of BRL on isolated ventricular myocytes from species other than guinea pig remain to be determined. Thus, a contribution of an increase of gKI to the effects of BRL is a possibility that requires further experimentation. It is also possible that BRL increases a potassium conductance that has not been described as yet.
The K-channel blockers tetraethylammonium (TEA) and 3,4 diaminopyridine inhibited the BRL-dependent Y6Rb efflux [Quast, 19871. Procaine and 5-aminopyridine inhibited the relaxant effect of BRL of the guinea pig isolated trachealis, whereas TEA had a lesser effect [Allen et al., 19861. This may indicate that BRL may enhance gXl , the potassium Conductance activated on depolarization. This would explain why the plateau is affected by BRL more than is the resting potential. However, it should be noted that the existence of I,, has been challenged and changes attributed to this current have been explained by changes in the slow inward current [Jaeger and Gibbons, 19851. In several reports, BKL induced quiescence and/or relaxation at concentrations lower than those required for the increment in “Rb or 42K movements [Weir and Weston, 1986a,b; Quast, 1987; Shetty and Weiss, 1987; McCullough et al., 19871. This might indicate that more than one mechanism is operational, or that the onset of quiescence (and relaxation) masks a small increase in rubidium movements [Wier and Weston, 1986b] or perhaps affects the pacemaker region before it modifies rubidium efflux from the bulk of the cells of the preparation [Quast, 1987; Hollingsworth et al., 19871. Thus, in the rat portal vein, hyperpolarization also requires concentrations of BRL higher than those required for relaxation [Weir and Weston, 1986bl. Therefore, BRL does increase potassium conductance, but it is still uncertain whether this effect is causally related to its mechanism of relaxation at low concentrations.
One possible alternative or additional action of BRL could be a stimulation of the activity of an electrogenic Na-K pump that also would lead to a shortening of the action potential. However, this possibility seems less likely since the hyperpolarization was marked

Actions of BRL 34915 in Purkinje Fibers 137
in zero K when the Na-K pump was certainly inhibited. Also, the experiments with strophanthidin clearly show that BRL can suppress spontaneous activity due to the inhibition of the Na-K pump. These results would not be conclusive since the Na-K pump might have been incompletely inhibited due to the presence of K in restricted extracellular spaces or to the nonoptimal concentration of strophanthidin. More decisive appears the demonstration that BRL does not decrease intracellular sodium activity in sheep Purkinje fibers (G. lacono, personal communication), as one would expected from a stimulation of the active transport of Na. Another possibility is that BRL enhances the Na-Ca exchange, but an electrogenic Na-Ca exchange [Mullins, 19791 would imply a depolarizing action at the resting potential. An inhibition of the Na-Ca exchange would imply a prolongation of the plateau and an increase in contractile force, but this did not occur either. Furthermore. actions on the Na-Ca exchange would leave unexplained the increase in rubidium efflux.
The Antiarrhythmic Action of BRL Because BRL has a hyperpolarizing action, it suppresses spontaneous activity by
depressing the slope and magnitude of DD, as the present experiments show. This action is potentially antiarrhythmic and it can suppress spontaneous rhythms independently of the underlying mechanisms. However, by shortening the action potential, BKL can be antiarrhyth- mic in other ways. The shift of the plateau to more negative values and its shortening should decrease the slow inward current. Not only would the activation of this current be smaller, but the residual slow inward current flowing during the plateau [see Reuter, 19791 would be curtailed as well. That this happens is suggested by the fact that the contractile force decreases. Therefore, BRL should not only increase potassium conductance but it may decrease intracellular calcium. Since certain arrhythmias (e .g . , digitalis toxicity) are caused by calcium overload and related oscillatory potentials [Kass et al., 19781, BRL should not only decrease the slope of DD but also eliminate Vos. The present experiments show that indeed V,, induced by high [Ca2+], is abolished by BRL.
The questions should be asked as to whether the antiarrhythmic and antihypertensive effects of BRL may be separated and whether the usefulness of BRL in patients with a compromised cardiac function may be limited by the negative inotropic effect. The answers must await further experimentation, but it should be noted that often diseased tissues from which the arrhythmias originate are more sensitive to drugs than the rest of the myocardium. This implies that BRL could be antiarrhythmic at doses that have little hypotensive action. In addition, it should be noted that slow channel blockers also can decrease (as BRL does) the blood pressure and myocardial contractility but, nevertheless, can be effective antiarrhythmic agents. Also, calcium overload not only can induce arrhythmias but also decrease contractile force of cardiac tissues [Vassalle and Lin, 19791, suggesting that a reduction of calcium overload in some patients with compromised cardiac function may actually increase the force of contraction. Finally, BRL might be particularly suited to treat arrhythmias in hypertensive patients.
Antian-hythmic activity has been reported for other purported K channel openers: Nicorandil in an in vitro canine Purkinje fiber model [Imanishi et al., 19841 and Pinacidil in an in vivo canine coronary artery ligation model [Kerr et al., 19851. The results of the present study suggest that it would be worthwhile to test BRL in other in vitro and in vivo models for antiarrhythmic activity.
In conclusion, BRL appears to be not only an antihypertensive drug but one that has potent antiarrhythmic actions in vitro, may increase coronary flow, have a sparing action of cardiac metabolism by decreasing the load placed upon the heart, and diminish the negative effects of calcium overload.
ACKNOWLEDGMENTS
synthesis of BRL 34915. This work was supported by the N.I.H. grant HL 17451. The authors thank Drs. K. Eickenberger and P. Wenk (Ciba-Geigy, AG, Basle) for the

138 Liu et al.
REFERENCES
Allen, S.L., Boyle, J.P., Cortijo, J . , Foster, R.W., Morgan, G.P., and Small. R.C.: Electrical and mechanical effects of BRL 34915 in guinea-pig isolated trachealis. Br. J. Pharmacol. 89:395-40.5, 1986.
Ashwood, V.A., Buckingham, R.E., Cassidy, F., Evans, J.M., and Famk, E.A.: Synthesis and antihypertensive activity of 4-(cyclic amino)-2H-l-benzopyrans. J. Med. Chem. 29:2194-2201, 1986.
Baumgarten, C.M. and Fozzard, H.A.: The resting and pacemaker potentials. In Fozzard, H.A., Haber E., Jennings R.B., Katz A.M. and Morgan H.E. (eds): “The Heart and Cardiovascular System,” Vol. 1. New York: Raven Press, pp 601-626, 1986.
Buckingham, R.E., Clapham, J .C. , Coldwell, M.C., Hamilton, T.C. and Howlett, D.R.: Differences in vascular effects of BRL 34915 and nifedipine in vitro. In “Proceedings of IXth IUPHAR Congress of Pharmacology.” London: Macmillan Press, p. 1702P, 1984.
Buckingham, R.E., Clapham, J.C., Hamilton, T.C., Longman, S.D., Norton, J., and Poyser, R.H.: BRL 34915, a novel antihypertensive agent: Comparison of effects on blood pressure and other haemodynamic parametcrs with those or Nifedipine in animal models. J . Cardiovasc. Phmaco l .
Cain, C.R. and Metzler, V.: Electrophysiological effects of the antihypertensive agent BRL 34915 in guinea-pig papillary muscle. Naunyn-Schmiedebergs Arch. Pharmacol. 329:R.53, 198.5 (abstract).
Carmeliet, E.E.: Chloride and Potassium Permeability in Cardiac Purkinje Fibers. Bruxelles: Arscia, S.A. and Presses AcadCmiques EuropCennes, 1961.
Carmeliet, E.: Electrophysiologic and voltage clamp analysis of the effects of sotalol on isolated cardiac muscle and Purkinje fibers. J. Pharmacol. Exp. Ther. 232:817-82.5, 198.5.
Conder, M.L. and McCullough, J.R.: Purported K channel opener, BRL 34915, blocks inwardly rectifying K current in isolated guinea-pig ventricular myocytes. Biophys. J. 51:258a, 1987.
Di Francesco, D.: A study of the ionic nature of the pace-maker current in calf Purkinje fibres. J. Physiol. (Lond.) 314:377-393, 1981.
Fox, J.S . , Whitehead, E . , and Shanks, R.G.: Cardiovascular effects of the novel antihypertensive agent BRL 34915 in normal volunteers. Br. J . Chem. Pharmacol. 23:601p, 1987.
Hamilton, T.C., Weir, S.W., and Weston, A.H.: Comparisonof the effects of BRL 34915 and verapamil on electrical and mechanical activity in rat portal vein. Br. J . Pharmacol. 88:103-111, 1986.
Hollingsworth, M., AmCdte, T., Edwards, D., Mironneau, J . , Savineau, J.P., Small, R.C., and Weston, A.H.: The relaxant action of BRL 34915 in rat uterus. Br. J . Pharmacol. 91503-813, 1987.
Imanishi, S . , Arita, M., Aomine, M., and Kiyosue, T.: Antiarrhythmic effects of Nicorandil on canine cardiac Purkinje fibers. J. Cardiovasc. Pharmacol. 6:772-779, 1984.
Imoto, Y., Ehara, T., and Matsura, H.: Voltage- and time-dependent of iKI underlying Bat *-induced ventricular automaticity. Am. J. Physiol. 252:H325-H333, 1987.
Isenberg, G.: Cardiac Purkinje fibers: Cesium as a tool to block inward rectifying potassium currents. Pfliigers Arch. 36599-106, 1976.
Isenberg, G.: Cardiac Purkinje fibers. [Ca” 1, controls the potassium permeability via the conductance components gkl and gkz. Pfliigers Arch. 371:77-85, 1977.
Ishikawa, S. and Vassalle, M.: Different forms of spontancous discharge induced by strophanthidin in cardiac Purkinje fibers. Am. J. Physiol. 243:H767-H778, 1982.
Jaeger, J.M. and Gibbons W.R.: Slow inward current may produce many results attributed to IX1 in cardiac Purkinje fibers. Am. J. Physiol. 249:H122-H132, 1985.
Kameyama, M., Kakei, M., Sato, R., Shibasaki, T., Matsuda, H., and Irisawa, H.: Intracellular Na’ activates a K’ channel in mammalian cardiac cells. Nature 309:354-356, 1984.
Kass, R.S. , Lederer, N.J., Tsien, R.W., and Weingart, R.: Role of calcium ions in transient inward currents and aftereontractions induced by stropanthidin in cardiac Purkinje fibres. J. Physiol. (Lond.) 281:187-208, 1978.
Kerr, M.J., Wilson, R., and Shanks R.G.: Suprression of ventricular arrhythmias after coronary artery ligation by Pinacidil, a vasodilator drug. J . Cardiovasc. Pharmacol. 7:875-883, 1985.
Liu, B., Golyan, D., McCullough, J.R., and Vassalle, M.: Electrophysiological and antiarrhythmic effects of the K channel opener BRL 34915 in Purkinje fibers. Physiologist 30:178, 1987.
McCullough, J.R., Shetty, S . S . , Conder, M.L., and Weiss, G.B.: Multiple actions of BRL 34915, a
8:798-804, 1986.

Actions of BRL 34915 in Purkinje Fibers 139
purported K channel opener. Proceedings 9th Annual Conference of the IEEE Engineering in Medicine and Biology Society, pp. 134-136; 1987.
Mullins, T.J.: The generation of the electrical currents in cardiac fibers by NaiCa exchange. Am. J . Physiol. 236:C103-110, 1979.
Noble, D. and Tsien, R.W.: The kinetics and rectifier properties of the slow potassium current in cardiac Purkinje fibres. J . Physiol. (Lond.) 195:185-214, 1968.
Noma, A,: ATP-regulatcd K + channels in cardiac muscle. Nature 305:147-148, 1983. Quast, U.: Effect of the K + efflux stimulating vasodilator BRL 34915 on X6Rb+ efflux and spontanous
activity in guinea-pig portal vein. Br. J . Pharmacol. 91569-578, 1987. Reuter, H.: The properties of two inward membrane currents in the heart. Annu. Rev. Physiol.
Sakmann, B., Noma, A., and Trautwein, W.: Acetylcholine activation of single muscarinic K + channels in isolated pacemaker cells of the mammalian heart. Nature 303250-253, 1983.
Scholtysik, G.: Evidence for inhibition by TCS 205-930 and stimulation by BRL 34915 of K+ conductance in cardiac muscle. Naunyn-Schmiedebergs Arch. Pharmacol. 335692-696, 1987.
Shetty, S.S. and Weiss, G.B.: Dissociation of actions of BRL 34915 in the rat portal vein. Eur. J. Phannacol. 141:485-488, 1987.
Siegelbaum, S. A. and Tsien, R. W.: Calcium-activated transient outward current in calf cardiac Purkinje fibres. J. Physiol. (Lond.) 299:485-506, 1980.
Straws, H.C., Bigger, J.T., and Hoffman, B.F.: Electrophysiological and beta-blocker effects of MJ 1999 on dog and rabbit cardiac tissue. Circ. Res. 26:661-678, 1970.
VandenBurg, M.J., Woodward, S.R., Hossain, M., Stewart-Long, P., and Tasker, T.C.G.: Potassium channel activators lower blood pressure: An initial study of BRL 34915 in hypertensive patients. J . Hypertens. 4:S166-S167, 1986.
Vassalle, M.: Cardiac pacemaker potentials at different extra- and intracellular K + concentrations. Am. J . Physiol. 208:770--775, 1965.
Vassalle, M.: Analysis of cardiac potential using a “voltage clamp” technique. Am. J. Physiol.
Vassalle, M.: Electrogenic suppression of automaticity in sheep and dog Purkinje fibers. Circ. Res. 27:361-377, 1970.
Vassalle, M. and Lin, C.4: Effect of calcium on strophanthidin-induced electrical and mechanical toxicity in cardiac Purkinje fibers. Am. J . Physiol. 236:H689-H697, 1979.
Watanabe, A.M.: Cholinergic agonists and antagonists. In Rosen, M.R. and Hoffman, B.F. ( 4 s ) : “Cardiac Therapy.” Boston: Martinus Nijhoff pp. 95-144, 1983.
Weidmann, S.: Effects of calcium ions and local anaesthetics on elcctrical properties of Purkinje fibres. J. Physiol. (Lond.) 129568-582, 1955.
Weir, S.W. and Weston, A.H.: Effect of apamin on responses to BRL 34915, Nicorandil and other relaxants in the guinea-pig taenia caeci. Br. J . Pharmacol. 88:113-120, 1986a.
Weir, S.W. and Weston, A.H.: The effects of BRL 34915 and Nicorandil on electrical and mechanical activity and on R6Rbf efflux in rat blood vessels. Br. J . Pharmacol. 88:121-128, 1986b.
4 1 ~ 4 13 -424, 1979.
230: 1335-1341, 1966.