Embed Size (px)
Citation preview

© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim 1434-2944/07/4-508-378
TADEUSZ FLEITUCH*, 1 and MARIA LEICHTFRIED2
1Institute of Nature Conservation, Polish Academy of Sciences, al. Mickiewicza 33, 31–120 Cracow,Poland; e-mail: [email protected]
2Institute of Limnology, Austrian Academy of Sciences, Mondseestr. 9, A-5310 Mondsee, Austria; e-mail: [email protected]
Electron Transport System (ETS) Activity in Alder Leaf Litter in Two Contrasting Headwater Streams
key words: decomposition, ETS activity, nutrients, geomorphology and hydrology, thermal regime
Abstract
Decomposition rates, carbon and nitrogen concentrations and respiration electron transport (ETS)activity in alder leaf litter were examined by bag exposition method in two contrasting 2nd order streams.Oberer Seebach, Austria (alpine, limestone, karstic) and Goscibia, Poland (sub mountain, flysh) con-trasted in catchment geology, channel hydrology, thermal regime and water chemistry. Despite differ-ences in water temperature, the breakdown rates did not show statistical differences. However, the C:Nratio in alder leaf litter varied significantly between two sites. The potential ETS activity was signifi-cantly higher in the colder Goscibia and weakly related to stream thermal regimes. The effect of tem-perature on ETS of alder leaves was not the dominating factor. It was masked by variation of other fac-tors like stream chemistry and the contribution of fine sediments, which are related to stream morphol-ogy and channel hydrology.
1. Introduction
Allochthonous organic matter, as terrestrial leaf litter input from the riparian zone of astream, is an important source of energy in forested headwater streams (e.g. FISHER andLIKENS, 1973; WEBSTER and BENFIELD, 1986). Leaf litter breakdown is thus a key elementof stream ecosystem function (GESSNER et al., 1999; GESSNER and CHAUVET, 2002; ROWE
et al., 1996). Differences in catchment land use and riparian vegetation, as well as reach-specific geomorphic factors alter light availability and organic matter supply. These are thefundamental factors controlling organic matter production, respiration, and transport instreams (YOUNG and HURYN, 1999). In addition to environmental factors, differences inbreakdown rates among species have been attributed to intrinsic factors of the leaves (BOUL-TON and BOON, 1991; POZO, 1993; WEBSTER and BENFIELD, 1986). The absolute concentra-tion of nitrogen (N) and the amount of carbon (C) relative to the amount of nitrogen (C/Nratio) in degraded leaf litter have generally been considered of primary importance (GULIS
and SUBERKROPP, 2003; ROYER and MINSHALL, 2001). In general, high concentrations of dis-solved nutrients, primarily nitrogen (N) and phosphorus (P), accelerate the processing ofleaves (KAUSHIK and HYNES, 1971; MEYER and JOHNSON, 1983). Breakdown also appears tooccur faster in hard water streams than in soft water streams (SUBERKROPP and CHAUVET,1995).
Functioning of streams and organic matter respiration rates are controlled by the availabili-ty of electron acceptors (e.g. oxygen, nitrate), electron donors (organic matter) and inorganic
Internat. Rev. Hydrobiol. 92 2007 4–5 378–391
DOI: 10.1002/iroh.200610986
* Corresponding author

nutrients. The availability of these substrates is often linked to trophic processes, which, inturn, reflect local geomorphic influences and regional hydrologic and climatic patterns (SIN-SABAUGH, 1997). HALL and TANK (2003) suggested that ecosystem metabolism provides adirect measurement of the food base of river ecosystems and thus helps to determine their lifesupporting capacity. Ecosystem metabolism has also recently been linked with nutrient uptake,another important function of river ecosystems. Therefore, information on ecosystem metabo-lism gives an indication of the likely capability of rivers to remove and transform nutrientsfrom the water. Moreover, functional measures (e.g. primary production, respiration) integratediverse communities into a few attributes and allow easier comparison among different sys-tems or within one system (FLEITUCH, 1998; GESSNER and CHAUVET, 2002; HILL et al., 2000).
The lack of a substantial literature on metabolic activity of leaf litter in ecosystem assess-ments is caused largely by the lack of understanding of the aquatic community within theecosystem. This is changing in light of studies of microbial community structure and meta-bolic activity in freshwater sediments (HILL et al., 2002; SINSABAUGH and FINDLAY, 1995)and few studies on leaf liter respiration in springs and streams (BARTODZIEJ and PERRY, 1990;GRAÇA and ABELHO, 2005; GULIS and SUBERKROPP, 2003; MULHOLLAND et al., 1984). Theresults of these studies indicate that measures of microbial metabolic processes may be thebest indicator of integrity of aquatic ecosystems and changes in microbial metabolic ratesmay be construed as an impact on these systems (NEWMAN et al., 1987; PERRY et al., 1987).
Community respiration is one of the most commonly measured functional attributes ofecosystems and has been proven to be a sensitive indicator of stream ecosystem integrity(BOTT et al., 1978, 1985, 1997; HILL et al., 1997, 1998, 2000). Respiration in aquatic sys-tems is usually determined by direct measurement of oxygen consumption.
Alternatively, respiration has also been measured as electron transport system (ETS) activ-ity in leaf litter (BARTODZIEJ and PERRY, 1990; MULHOLLAND et al., 1984). The ETS assay,proposed originally by PACKARD (1971), has been used to estimate respiratory potential ofzooplankton, microplankton, benthos, sediments, and macrophytes (see methods and litera-ture overview in G.-TÓTH, 1999; SZABO 2003; SIMCIC, 2005). Production in ETS assays hasbeen found to be closely correlated to oxygen consumption (DEL GIORGIO, 1992).
The aim of this study was to test the hypothesis that different geomorphologic catchmentproperties and regional climatic conditions in organic matter-based streams would influenceecosystem functioning through effects on metabolic activity of leaf litter. For this purpose,we compared leaf litter breakdown and respiration activity in two different headwaterstreams (eco-regions: Alps and Carpathians) running through mixed forests in CentralEurope. We used a fast decomposing leaf species as a high quality food resource for aquat-ic organisms in order to assess how differences between mountain streams influence leaf lit-ter breakdown, leaf nutrition quality and its potential respiration activity.
2. Study Area
Two pristine, nature-protected stream areas were investigated simultaneously in 2003 and2004: the Goscibia stream in southern Poland and the Oberer Seebach near Lunz/See, LowerAustria.
The 2nd order stream (STRAHLER, 1957) Goscibia (GOS) is situated in the Beskid Makow-ski Mountains and has a catchment area of 16 km2, forested up to 88%. The stream lengthis 7.8 km, mean width is 4.4 m and the mean depth is 30 cm. The streambed consists main-ly of gravels, small stones and fine sediments.
The second site, Oberer Seebach (OSB), was selected in the lower reach of a 2nd orderalpine stream on the northern fringe of the Alps, near Biological Station Lunz (Lower Aus-tria). The stream length is around 7 km and its catchment area is 20 km2. The catchment isuninhabited and forested. The mean width at the sampling site is 4.4 m and the mean depth
ETS Activity in Alder Leaf Litter 379
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com

is 40 cm. The bed sediments are badly sorted; the grain size median is around 30 mm indiameter (LEICHTFRIED, 1995). Grain size smaller 1 mm in diameter is always under 10% byweight but this fraction always contains more than 80% of all nutrients in bed sediments(LEICHTFRIED, 1986, 1988).
Both streams are oligotrophic and their discharge regimes are natural. The riparian canopyof the streams is mostly closed in summer and is similar between the streams. Deciduoustrees dominate the riparian vegetation with the presence of mountain maple (Acer pseudo-platanus L.), beech (Fagus silvatica L.), European ash (Fraxinus excelsior L.), willow (Salix carea L.), spruce (Picea abies (L.) KARST.) and others (MOSER, 1991, 1992). The selected sites differ in catchment geology, altitude, latitude, longitude and hydrologicalparameters (precipitation and water discharge). The data for each site are included in Table 1with the main water chemistry variables.
3. Material and Methods
3.1. Leaf-Bag Experiments
To assess leaf breakdown rate and the plant litter-associated ETS activity, we used alder leaves (Alnusglutinosa (L.) GAERTN.) representing a fast decomposing species (PETERSEN and CUMMINS, 1974). Thelitter mesh bag method was used to expose the leaves to decomposition in the streams (BOULTON andBOON, 1991). Freshly fallen leaves of alder were collected from one alder tree at abscission during lateOctober 2003 in Poland, placed into single mesh bags and distributed at both study sites.
Portions of air-dried leaves (ca. 5 g) were weighted to 0.01 g accuracy and enclosed in coarse (10 mmmesh) plastic bags and closed in a tetrahedral shape (JONSSON et al., 2001). The coarse mesh allowedmicrobial and large shredder colonization (BOULTON and BOON, 1991). On November 4th 2003, fiveexperimental sets of litter mesh bags were installed concurrently in riffles in each of the two streamsand secured by rebar anchored in the streambed. Each set consisted of four bags.
Just after weighing and prior to transporting to the field, the leaf litter was sprayed with distilledwater to prevent breakage during handling and transport. In total, 20 bags were exposed in each stream.Four additional fine mesh-size bags of alder leaves were prepared but kept in the laboratory as controlsto determine initial leaf ash-free dry mass (AFDM). Four replicates of bags were retrieved from differ-ent rebars in each stream after 7, 14, 35, 77, and 126 days and stored individually in plastic zip-lockbags. On the last sampling date, two bags were lost due to dislodgment by melting ice (GOS site) andtwo others due to high flow (OSB site).
3.2. Sample Processing
Upon retrieval, the leaves were washed individually with tap water to remove sediments, sand, exoge-nous leaves, non-alder leaves and invertebrates. One set of five 12 mm diameter discs were cut in fiveleaves from each bag, avoiding the central veins. Additionally, the 5 discs were cut from unexposedleaves in order to test their initial metabolic activity. Immediately after cutting, all discs were frozen(–30 °C) and used later to determine ETS activity. The remaining leaf material was dried at 105 °C for48 h and weighed to 0.01 g accuracy. Then, leaf sub-samples of about 300–500 mg were ashed at550 °C for 4 h and weighed to determine the ash free dry mass (AFDM). Alder mass loss (MR) wasexpressed as % of AFDM remaining (MR = (Mt/M0) ¥ 100; where: Mt – AFDM after time t, M0 – ini-tial AFDM). Moreover, leaf breakdown rates (kd) were estimated by fitting the AFDM data to the expo-nential model, Yt = Y0 e–kd t, where Yt is the AFDM remaining at time t in days, and Y0 the AFDM at thebeginning of the experiment (PETERSEN and CUMMINS, 1974). To calculate decomposition rates on adegree day basis (kdd) from the exponential model, time t´ (in degree days) was used.
Sub-samples of the dried bulk material (about 1 g) were used to estimate total organic carbon (TOC)and total nitrogen (TN) concentrations with the use of a LECO CHN 600 analyzer. Sub-samples of extracollected leaves were used for determination of initial concentration (blank sample) of total organic car-bon (TOC) and total nitrogen (TN). Concurrently with the dates of experiments with leaf breakdown,
380 T. FLEITUCH and M. LEICHTFRIED
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com

the physical and chemical parameters of the stream water were measured monthly: pH, specific con-ductance, alkalinity, dissolved inorganic nitrogen (sum of ammonia-, nitrates- and nitrites- nitrogen) andphosphorus concentrations (APHA, 1989). Based on these analyses, the C: N ratio for each sample wascalculated. During the breakdown study, water temperature was recorded every 2 h (Smart Button, ACRSystem Inc.). For each stream and sampling date, thermal regime was calculated as degree-days.
ETS activity was used as an indicator of potential alder leaf respiration. We used the laboratorymethod described by PACKARD (1971) and further modified by KENNER and AHMED (1975), OWENS andKING (1975), and G.-TÓTH (1999). All chemicals were purchased from Sigma, except for NADH2 (ICNBiomedicals Inc.). Immediately before the analysis, five discs of each sample were weighted using amicrobalance of 0.1 mg accuracy and grained with liquid nitrogen in a porcelain mortar until the mate-rial particles were reduced to a fine powder. The contents of the mortar were washed with 1 ml of icecold homogenization buffer, pH 8.25 (0.1 M Na2HPO4, 0.1 M KH2HPO4, 0.075 mM MgSO4, 0.2 vol.Triton X-100, 0.06 mM polivinyl-pyrrolidone (PVP)). Samples were centrifuged at 5000 g at 0 °C for5 minutes and kept on ice prior to analysis. When prepared by this method, none of the supernatantsshowed opalescence. The test was performed in triplicates (examined samples) with a control of unspe-cific reduction for each sample (control sample). The test tubes were placed in a warm water bath(20 °C) with gentle agitation and 750 µl of substrate buffer pH 8.25 (0.1 M Na2HPO4, 0.1 M KH2HPO4,0.075 mM MgSO4, 0.2 vol. Triton X-100, 1 mM NADH2, 0.25 mM NADPH2) was added to each ofthem. Next, 250 µl of supernatant were pipetted and after a few minutes, when the temperature stabi-lized, 250 µl of 4 mM INT (2-(4-iodophenyl)-3-(4-nitrophenyl)-5-phenyl tetrazolium chloride) solutionwere added (start of the reaction). The test tubes were capped and incubated for 60 min (this time wasdetermined experimentally; after 60 min the colour was most intensive). The reaction was stopped bythe addition of 250 µl quenching solution (1 M H3PO4 + 4% formalin 1:1v/v). The control sample(unspecific reduction) had the same content but the aliquots were added in the following sequence: sub-strate buffer, quenching solution, examined supernatant. Samples were vortexed for 5 min and cen-trifuged at 3500 rpm for 10 min. As a coloured product reaction, formazan was measured as absorbancein a LKB Ultraspec II spectrophotometer at 490 nm against a distilled water blank. The formazanabsorbance was calculated by subtracting the control sample absorbance at 490 nm from the meanabsorbance of the examined samples at the same wavelength. This value was used to calculate ETSactivity as µl O2 per hour and per g of ash-free dry mass (G.-TÓTH, 1999).
The control sample was prepared to eliminate both any possible unspecific (i.e. unconnected with theETS enzymes activity) reduction of INT and absorbance due to reagents and substances present inleaves. The quenching solution applied (acidified formaldehyde) completely terminates formazan pro-duction (CHRISTENSEN and PACKARD, 1979) so it was not necessary to prepare blanks with boiled orautoclaved homogenate.
3.3. Data Analysis
To assess an overall dispersion of data between the two sites, the coefficient of variance (CV) wascalculated. Processing mass remaining was tested and regressed on time to find the best fit with the useof different regression models. ETS activity was regressed on stream water temperature to predict themost significant model for the relationship. Analysis of variance (ANOVA; time versus site) was usedto test the differences in terms of carbon, nitrogen and C/N ratio in alder litter. Analysis of covariance(ANCOVA), with time as covariate, was used in the GLM model to compare slopes between sites interms of processing rates. Data were lognormal transformed whenever necessary to achieve normal dis-tribution. For graphic presentation, non-transformed data (mean and standard error of the mean (SE))were used. Differences were considered significant when P < 0.05. Calculations were made with statis-tical packages SYSTAT and SPSS.
4. Results
4.1. Stream Characteristics
Both study streams showed similar, slightly basic pH of around 8 and relatively low spe-cific conductance (<270 µS cm–1). In terms of water chemistry, the Goscibia was richer in
ETS Activity in Alder Leaf Litter 381
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com

nutrient, with twice the inorganic phosphorus and ammonia nitrogen (Table 1) of the Obe-rer Seebach. In the limestone stream (OSB), alkalinity was about two times higher than inthe sandstone stream (GOS). The two sites differed mainly in altitude, catchment geology,bed structure (small gravel and clay prevailed in GOS), water discharge (on average 20 timeshigher in OSB) and water thermal regime (mean temperatures and degree days, Table 1).The Goscibia stream, mean temperature 3.3 °C, was significantly colder than Oberer See-bach, where the mean temperature was 6.8 °C.
4.2. Processing Rates
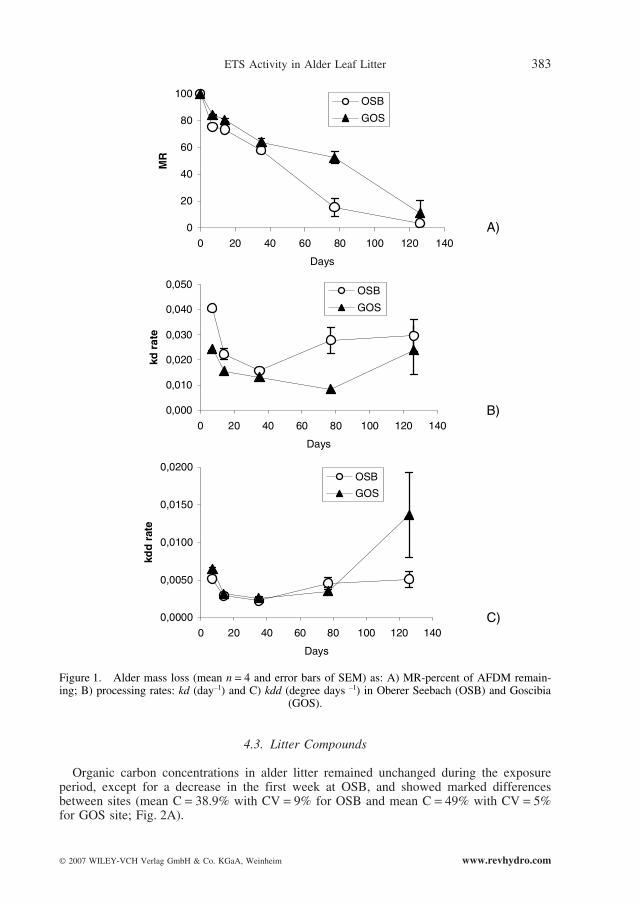
Overall processing of alder leaves in the two streams was similar and rapid, with approx-imately 40% mass lost in the first month (Fig. 1A). Between December and March, alderprocessing was distinctly slower at the GOS site. The data on mass remaining, obtained fromeach stream, were used to test two models: linear and exponential. For the OSB site, the bestfit of alder mass remaining was obtained by use of an exponential model whilst, for the GOSstream, a linear model provided the best fit (Table 2).
During the same initial period, mean kd showed a wider range at the warmer OSB site(from 0.041 to 0.016) than at the colder GOS site (from 0.021 to 0.013). Processing rates ofkd and kdd (Fig. 1 B and C) indicated that breakdown in both streams consisted of two dis-tinct phases. An initial, rapid decrease, within 35 days, was followed by an increase in thetwo processing rates until March. In the final study period (January–March), a greaterincrease in processing rate was noted for the GOS site. Observed differences in processingrates between sampling dates throughout the study period were not confirmed by ANCOVA, which showed a non-significant interaction term “stream ¥ time” (P > 0.05) andindicated that similarities in patterns between sites of the three processing rates were consistent.
382 T. FLEITUCH and M. LEICHTFRIED
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
Table 1. Selected catchment characteristics and water physico-chemical parameters of thestream water of two study sites on the Oberer Seebach (OSB) and the Goscibia stream(GOS) from November 2003 to March 2004. Parameter’ ranges are given in parentheses.
(OSB: n = 5, GOS: n = 6).
Site characteristics OSB GOS
Stream order (STRAHLER, 1957) 2 2Annual precipitation (mm) 1515 750Altitude (m) 605 350Longitude 15°04´ E 19°49´ ELatitude 47°85´ N 49°50´ NEcoregion (Water Framework Directive) Alps CarpathiansMean width (m) 15.8 4.4Mean depth (cm) 0.50 0.30Mean discharge (l s–1) 720 34Water temperature °C (mean) 6.8 3.3Degree days (126 days) 733.6 222.5pH 8.1 (7.9–8.3) 7.9 (7.6–8.0)Specific conductance (µS/cm at 20 °C) 216 (112–283) 269 (157–378)Alkalinity (mg CaCO3/l) 218 (145–307) 107 (36–157)DIN (dissolved inorganic nitrogen, µg/l) 1010 (840–2000) 1084 (870–2100)N–NH4 µg/l 126 (89–168) 284 (135–349)P–PO4 µg/l 9 (5–14) 42 (17–60)
4.3. Litter Compounds
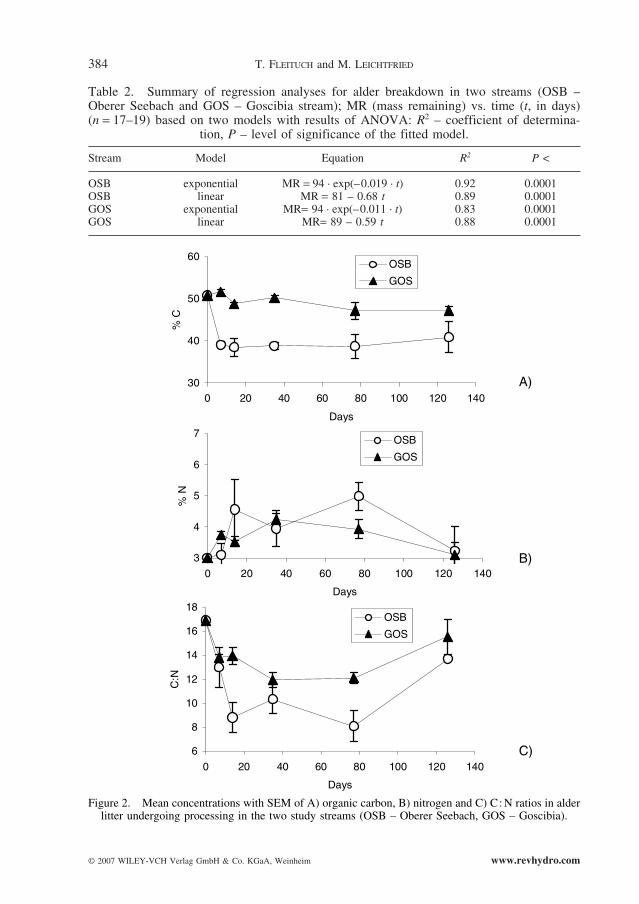
Organic carbon concentrations in alder litter remained unchanged during the exposureperiod, except for a decrease in the first week at OSB, and showed marked differencesbetween sites (mean C = 38.9% with CV = 9% for OSB and mean C = 49% with CV = 5%for GOS site; Fig. 2A).
ETS Activity in Alder Leaf Litter 383
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
0
20
40
60
80
100
0 20 40 60 80 100 120 140
Days
MR
OSB
GOS
0,000
0,010
0,020
0,030
0,040
0,050
0 20 40 60 80 100 120 140
Days
kdra
te
OSB
GOS
0,0000
0,0050
0,0100
0,0150
0,0200
0 20 40 60 80 100 120 140
Days
kdd
rate
OSB
GOS
Figure 1. Alder mass loss (mean n = 4 and error bars of SEM) as: A) MR-percent of AFDM remain-ing; B) processing rates: kd (day–1) and C) kdd (degree days –1) in Oberer Seebach (OSB) and Goscibia
(GOS).
A)
B)
C)
384 T. FLEITUCH and M. LEICHTFRIED
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
Table 2. Summary of regression analyses for alder breakdown in two streams (OSB –Oberer Seebach and GOS – Goscibia stream); MR (mass remaining) vs. time (t, in days)(n = 17–19) based on two models with results of ANOVA: R2 – coefficient of determina-
tion, P – level of significance of the fitted model.
Stream Model Equation R2 P <
OSB exponential MR = 94 · exp(–0.019 · t) 0.92 0.0001OSB linear MR = 81 – 0.68 t 0.89 0.0001GOS exponential MR= 94 · exp(–0.011 · t) 0.83 0.0001GOS linear MR= 89 – 0.59 t 0.88 0.0001
30
40
50
60
0 20 40 60 80 100 120 140
Days
%C
OSB
GOS
3
4
5
6
7
0 20 40 60 80 100 120 140
Days
%N
OSB
GOS
6
8
10
12
14
16
18
0 20 40 60 80 100 120 140
Days
C:N
OSB
GOS
Figure 2. Mean concentrations with SEM of A) organic carbon, B) nitrogen and C) C: N ratios in alderlitter undergoing processing in the two study streams (OSB – Oberer Seebach, GOS – Goscibia).
A)
B)
C)
A parallel increase in nitrogen concentrations at the both sites was observed during thetwo weeks of initial incubation (Fig. 2B). A maximal N concentration of 5.6% was found inthe litter on day 77 at the OSB. In separate comparisons of the C and N concentrations andthe “site ¥ time” interactions, ANCOVA did not detect significant differences (P > 0.05) foreither element or for the OSB and GOS sites. The C: N atomic ratio changed distinctly withincubation time (Fig. 2C) and on average it was higher at the GOS site (13.4) than at theOSB site (9.9). The lowest C:N proportion and the highest N concentration were found atthe OSB site in January, on day 77. Significant differences in C:N ratio (interaction“stream ¥ time”; ANCOVA, F = 4.34, P < 0.05) were detected between sites.
4.4. ETS (Electron Transport System) Activity
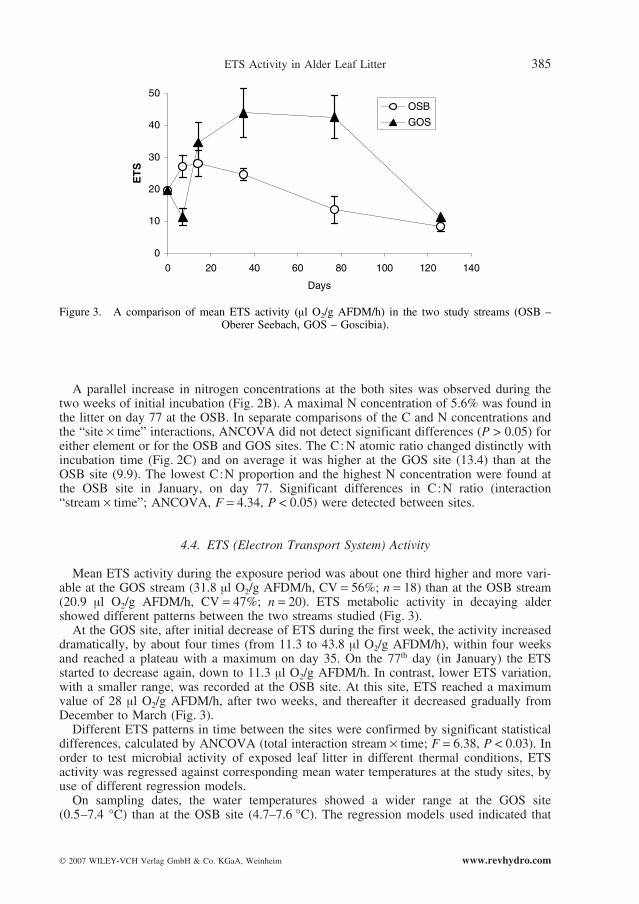
Mean ETS activity during the exposure period was about one third higher and more vari-able at the GOS stream (31.8 µl O2/g AFDM/h, CV = 56%; n = 18) than at the OSB stream(20.9 µl O2/g AFDM/h, CV = 47%; n = 20). ETS metabolic activity in decaying aldershowed different patterns between the two streams studied (Fig. 3).
At the GOS site, after initial decrease of ETS during the first week, the activity increaseddramatically, by about four times (from 11.3 to 43.8 µl O2/g AFDM/h), within four weeksand reached a plateau with a maximum on day 35. On the 77th day (in January) the ETSstarted to decrease again, down to 11.3 µl O2/g AFDM/h. In contrast, lower ETS variation,with a smaller range, was recorded at the OSB site. At this site, ETS reached a maximumvalue of 28 µl O2/g AFDM/h, after two weeks, and thereafter it decreased gradually fromDecember to March (Fig. 3).
Different ETS patterns in time between the sites were confirmed by significant statisticaldifferences, calculated by ANCOVA (total interaction stream ¥ time; F = 6.38, P < 0.03). Inorder to test microbial activity of exposed leaf litter in different thermal conditions, ETSactivity was regressed against corresponding mean water temperatures at the study sites, byuse of different regression models.
On sampling dates, the water temperatures showed a wider range at the GOS site (0.5–7.4 °C) than at the OSB site (4.7–7.6 °C). The regression models used indicated that
ETS Activity in Alder Leaf Litter 385
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
0
10
20
30
40
50
0 20 40 60 80 100 120 140
Days
ET
S
OSB
GOS
Figure 3. A comparison of mean ETS activity (µl O2/g AFDM/h) in the two study streams (OSB –Oberer Seebach, GOS – Goscibia).
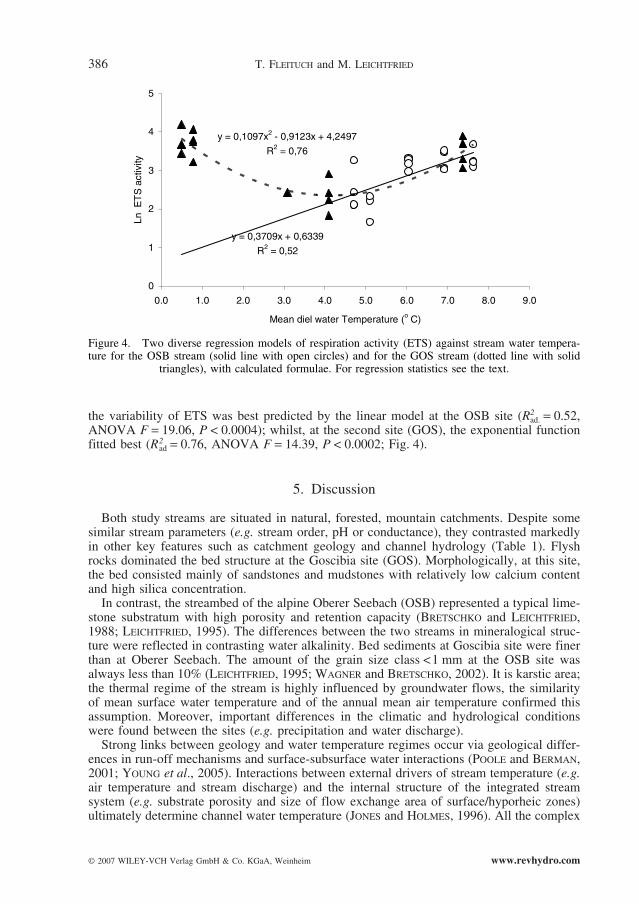
the variability of ETS was best predicted by the linear model at the OSB site (R2ad. = 0.52,
ANOVA F = 19.06, P < 0.0004); whilst, at the second site (GOS), the exponential functionfitted best (R2
ad = 0.76, ANOVA F = 14.39, P < 0.0002; Fig. 4).
5. Discussion
Both study streams are situated in natural, forested, mountain catchments. Despite somesimilar stream parameters (e.g. stream order, pH or conductance), they contrasted markedlyin other key features such as catchment geology and channel hydrology (Table 1). Flyshrocks dominated the bed structure at the Goscibia site (GOS). Morphologically, at this site,the bed consisted mainly of sandstones and mudstones with relatively low calcium contentand high silica concentration.
In contrast, the streambed of the alpine Oberer Seebach (OSB) represented a typical lime-stone substratum with high porosity and retention capacity (BRETSCHKO and LEICHTFRIED,1988; LEICHTFRIED, 1995). The differences between the two streams in mineralogical struc-ture were reflected in contrasting water alkalinity. Bed sediments at Goscibia site were finerthan at Oberer Seebach. The amount of the grain size class <1 mm at the OSB site wasalways less than 10% (LEICHTFRIED, 1995; WAGNER and BRETSCHKO, 2002). It is karstic area;the thermal regime of the stream is highly influenced by groundwater flows, the similarityof mean surface water temperature and of the annual mean air temperature confirmed thisassumption. Moreover, important differences in the climatic and hydrological conditionswere found between the sites (e.g. precipitation and water discharge).
Strong links between geology and water temperature regimes occur via geological differ-ences in run-off mechanisms and surface-subsurface water interactions (POOLE and BERMAN,2001; YOUNG et al., 2005). Interactions between external drivers of stream temperature (e.g.air temperature and stream discharge) and the internal structure of the integrated stream system (e.g. substrate porosity and size of flow exchange area of surface/hyporheic zones)ultimately determine channel water temperature (JONES and HOLMES, 1996). All the complex
386 T. FLEITUCH and M. LEICHTFRIED
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
y = 0,3709x + 0,6339
R2 = 0,52
y = 0,1097x2 - 0,9123x + 4,2497
R2 = 0,76
0
1
2
3
4
5
0.0 1.0 2.0 3.0 4.0 5.0 6.0 7.0 8.0 9.0
Mean diel water Temperature (o C)
LnE
TS
activ
ity
Figure 4. Two diverse regression models of respiration activity (ETS) against stream water tempera-ture for the OSB stream (solid line with open circles) and for the GOS stream (dotted line with solid
triangles), with calculated formulae. For regression statistics see the text.

factors above may influence the differences in thermal regimes between the streams studied.The Oberer Seebach (OSB) channel, with higher mean water temperatures, exhibited a
complex streambed topography (BRETSCHKO, 1998; WAGNER and BRETSCHKO, 2002) and hada high rate of hyporheic upwelling flow in riffle habitats. The Goscibia stream (GOS), withrelatively low streambed complexity (i.e. pool/riffle sequences), a high amount of fine sed-iments, and freezing water surface during winter (January–February), was more influencedby atmospheric conditions than by groundwater inflow. Additionally, the reduction ofgroundwater inflow at this site may have been caused by clogging effect of fine sediments,which dominated the stream reach. In this respect, several authors have demonstrated thatclogging of the streambed surface will often isolate deeper regions of the bed from thestream flow, so that even relatively low amounts of suspended sediments can substantiallychange streambed habitat (e.g. PACKMAN and MACKAY, 2003). Many investigations havefound, that hyporheic water velocity is controlled by the surface flow regime (ANGRADI andHOOD, 1998; BRUNKE and GONSER, 1997). Although WAGNER and BRETSCHKO (2002)demonstrated that, in Oberer Seebach, bed sediments formed a complex three-dimensionalnetwork of sediment areas with high hydraulic conductivities, they did not find a relation-ship between interstitial flow and stream discharge.
Levels of alder breakdown rates found in the present study confirmed the assumption thatit was a relatively fast decomposing species (PETERSEN and CUMMINS, 1974). Similar k-ratesfor alder have been obtained with the use of comparable mesh bag size, stream temperaturerange and altitude in the French Pyrenees (GESSNER, 1991). Many studies involving a widevariety of leaf species and specific thermal conditions have shown faster leaf breakdown inwarmer than in colder streams and also when warm periods have been compared with coldperiods in seasonal studies (WEBSTER and BENFIELD, 1986). However, our results indicatedthat contrasted thermal regimes between streams did not alter alder breakdown rates signi-ficantly. Moreover, after degree-day adjustment, the breakdown rates (except the last sam-pling date) showed very similar patterns between the sites. Also, the noticeable differencesbetween stream water chemistry (alkalinity, ammonia and dissolved inorganic phosphorus)did not affect alder processing.
Lack of differences in breakdown rates between streams with varied thermal conditionsmaybe a masking effect of other different, influential parameters. In this study, an examplemight be shredder abundances and their feeding activity on exposed leaf litter in coarse meshbags. According to some authors (SHORT et al., 1980), invertebrates may play a dominantrole in decomposition and breakdown rates can be high, even at very cold temperatures.Such conditions were recorded at the Goscibia site during the winter. Furthermore, indecomposition processes, the role of the complex of meiobenthic fauna and biofilms has notyet been studied.
In this study, the initial nitrogen concentration in alder leaves represented 3% of AFDM andincreased during the first 35 days of leaf decomposition at both sites. Overall, the immobi-lization of nitrogen in leaves has been partially attributed to the accumulation of microbial bio-mass (WEBSTER and BENFIELD, 1986). Similarly, in Portuguese streams, PASCOAL et al. (2005)found that the initial increase in nitrogen concentration in alder leaves was associated with highbiomass and production of fungi, the first colonizers of biofilms. The subsequent decrease innitrogen concentration was possibly caused by the release of large numbers of conidia.
In the Oberer Seebach, the first nitrogen peak was observed on day 14 and the secondlater, on day 77. These two peaks may suggest, that the early high N concentration was relat-ed to the activity of microfungi (aquatic hyphomycetes, SUBERKROPP, 1991) and the secondone to an increase of bacterial biomass in biofilms (BALDY et al., 1995). However, nitrogenimmobilization may also result from the formation of complexes between nitrogen and othercompounds in leaves (ODUM et al., 1978; RICE, 1982). The similarity in leaf decompositionrates between Oberer Seebach and Goscibia sites is in accordance with the lack of statisti-cal differences in nitrogen concentration in alder litter.
ETS Activity in Alder Leaf Litter 387
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com

In general, stream temperature influences the metabolic rates, physiology, and life-histo-ry traits of aquatic species, helping to determine important community processes such as pro-ductivity or decomposition (ALLEN, 1995). There are abundant data showing significantlyhigher respiration rates in summer compared to winter (e.g. WEBSTER et al., 1995). How-ever, several studies have suggested modest effects of temperature on ecosystem respiration(HILL et al., 1998, 2000, 2002; SINSABAUGH, 1997). At an ecosystem scale, the effect ofwater temperature seems to be weak (MULHOLLAND et al., 2001).
Little is known about the magnitude of microbial respiration on decomposing leaves infreshwater streams (GULIS and SUBERKROPP, 2003; PASCOAL et al., 2005), especially the useof the ETS assay. The only comparable exception is a study of ETS microbial activity inrelation to the decomposition of watercress and box elder in different spring categories in akarstic region of Minnesota (BARTODZIEJ and PERRY, 1990). The authors showed that break-down rates and those processes attributable to ETS activity didn’t differ greatly between thespring classes investigated. In contrast, our study demonstrated strong significant differencesin ETS microbial activity between the two sites. ETS activity is not strongly dependant onwater temperature. The different models created for this relationship indicated that factorsother than temperature might be responsible for alder processing and its respiration activityunder the two contrasting sets of environmental conditions.
Decomposition processes and their ETS activity are influenced by different biotic and ab-iotic parameters. The effects of temperature on these processes did not dominate in our studybut these could have been masked by variation in other factors.
Several studies report a positive relationship between respiration rates and the amount offine sediments on the streambed (HEDIN, 1990; HILL et al., 1998). Fine sediments have expo-nentially higher surface area than coarse sediments (LEICHTFRIED, 1985, 1988) and offer ahigh amount of colonisable surface area for the development of biofilms (defined as micro-bicoenosis, e.g. fungi, bacteria, protozoa and algae with their exocellular excretions attachedto surfaces, MARSHALL, 1984). Biofilms and their associated meiobenthic fauna play animportant role in decomposition processes (LEICHTFRIED, 1995, 1999), even if though fewinvestigations of this complex have yet been undertaken. Such mechanisms, together withthe elevated contribution of fine sediments, can partially explain the increase in ETS activ-ity at the Goscibia site during winter.
Another factor masking the effects of relatively high ETS activity at the colder Goscibiastream could be higher nutrient concentrations in the surface water. These findings can beconfirmed by studies conducted in warmer streams with different nutrient availability. Forexample, MENENDEZ et al. (2003) found a temperature-related, faster rate of leaf breakdownduring summer in an eutrophic Spanish river but not in a neighboring oligotrophic stream.
Basically, the assessment of ETS activity in leaf litter may be a promising and integrat-ing method for determining the functional status of headwater streams (GESSNER and CHAU-VET, 2002). However, the functional response (e.g. respiration) of aquatic communities maydepend on several multiple environmental factors and this makes it difficult to predict thefinal functional status of a stream ecosystem.
6. Acknowledgements
Many thanks are given to Prof. Dr. ANDRZEJ SKOCZOWSKI for his assistance in ETS analyses and toDr. PAWEL OLEJNICZAK for his statistical advice. Special thanks to Mrs TERESA FRYDRYCH and Dipl. Ing.WALBURGA FAHRNER for providing laboratory analyses. The authors also greatly appreciate the samplingin the field undertaken by SZYMON CIAPALA, LUKASZ SROKA and ENGELBERT SOLLBOECK. The authorsthank Dr. MIKE FURSE for improving English version of the manuscript and two anonymous reviewersfor their constructive comments. The project was partially supported by the Austrian Academic ExchangeService in Vienna.
388 T. FLEITUCH and M. LEICHTFRIED
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com

7. References
ALLEN, J. D., 1995: Stream Ecology: Structure and Function of Running Waters. Chapman & Hall, NewYork, 388 pp.
ANGRADI, T. and R. HOOD, 1998: An application of the plaster dissolution method for quantifying watervelocity in the shallow hyporheic zone of an Appalachian stream system. – Freshw. Biol. 39:301–315.
APHA (American Public Health Association), 1989: Standard methods for examination of water andwastewater, 17th edn. American Public Health Association, Washington D.C.
BALDY, V., M. O. GESSNER and E. CHAUVET, 1995: Bacteria, fungi and the breakdown of leaf litter ina large river. – Oikos 74: 93–102.
BARTODZIEJ, W. and J. A. PERRY, 1990: Litter processing in diffuse and conduit springs. – Hydrobiol.206: 87–97.
BOTT, T. L., J. T. BROCK, C. E. CUSHING, S. V. GREGORY, D. KING, R. and C. PETERSEN, 1978: A com-parison of methods for measuring primary productivity and community respiration in streams. –Hydrobiol. 60: 3–12.
BOTT, T. L., J. T. BROCK, C. S. DUNN, R. J. NAIMAN, R. W. OVINK and R. C. PETERSEN, 1985: Benthiccommunity metabolism in four temperate stream systems: An interbiome comparison and evaluationof the river continuum concept. – Hydrobiol. 123: 3–45.
BOTT, T. L., J. T. BROCK, A. BAATTRUP-PEDERSEN, P. A. CHAMBERS, W. K. DODDS, K. T. HIMBEAULT,J. R. LAWRENCE, D. PLANAS, E. SNYDER and G. M. WOLFAARDT, 1997: An evaluation of techniquesfor measuring periphyton metabolism in chambers. – Can. J. Fish. Aq. Sc. 54: 715–725.
BOULTON, A. J. and P. I. BOON, 1991: A review of methodology used to measure leaf litter decomposi-tion in lotic environments: time to turn over an old leaf? – Austral. J. Mar. Freshw. Res. 42: 1–43.
BRETSCHKO, G. and M. LEICHTFRIED, 1988: Distribution of organic matter and fauna in a second orderalpine gravel stream (RITRODAT-LUNZ). – Verh. Internat.Verein. Limnol. 23: 1333–1339.
BRETSCHKO, G., 1998: The river bottom in gravel streams. – In: BRETSCHKO, G. and J. HELESIC (eds.):Advances in River Bottom Ecology. – Backhuys Publishers, Leiden: 1–11.
BRUNKE, M. and T. GONSER, 1997: The ecological significance of exchange processes between riversand groundwater. – Freshw. Biol. 37: 1–33.
CHARACKLIS, W. G. and P. A. WILDERER (eds), 1989: Structure and Function of Biofilms. – DahlemWorkshop Rep., Life Sciences Res. Rep. 46, John Wiley & Sons Ltd.: 387 pp.
CHRISTENSEN, J. P. and T. T. PACKARD, 1979: Respiratory electron transport activities in phytoplanktonand bacteria: Comparison of methods. – Limnol. Oceanogr. 24: 576–583.
DEL GIORGIO, P. A., 1992: The relationship between ETS (electron transport system) activity and oxy-gen consumption in lake plankton: a cross-system calibration. – J. Plankt. Res. 14: 1723–1741.
FISHER, S. G. and G. E. LIKENS, 1973: Energy flow in Bear Brook, New Hampshire: an integrativeapproach to stream ecosystem metabolism. – Ecol. Monogr. 43: 421–439.
FLEITUCH, T., 1998: Responses of benthic community metabolism to abiotic factors in a mountain riverin southern Poland. – Hydrobiol. 380: 27–41.
G.-TÓTH, L., 1999: Aktivität des Elektronentransportsystems (ETS). In: TÜMPLING, W. V. and G.FRIEDRICH (eds.): Methoden der biologischen Wasseruntersuchung 2., Gustav Fischer Verlag, Jena,Stuttgart, Lübeck, Ulm: pp. 465–473.
GESSNER, M. O., 1991: Differences in processing dynamics of fresh and dried leaf litter in a streamecosystem. – Freshw. Biol. 26: 387–398.
GESSNER, M. O., E. CHAUVET and M. DOBSON, 1999: A perspective on leaf litter breakdown in streams.– Oikos 85: 377–384.
GESSNER, M. O. and E. CHAUVET, 2002: A case for using litter breakdown to assess functional streamintegrity. – Ecol. Appl. 12: 498–510.
GRAÇA and M. ABELHO, 2005: Respirometry in Methods to Study Litter Decomposition. – In: GRAÇA,M. A. S., BÄRLOCHER, F. and GESSNER, M. O. (eds.). A Practical Guide: Springer: pp. 231–236.
GULIS, V. and K. SUBERKROPP, 2003: Leaf litter decomposition and microbial activity in nutrient-enriched and unaltered reaches of a headwater stream. – Freshw. Biol. 48: 123–134.
HALL, R. O. and J. L. TANK, 2003: Ecosystem metabolism controls nitrogen uptake in streams in GrandTeton National Park, Wyoming. – Limnol. Oceanogr. 48: 1120–1128.
HEDIN, L. O., 1990: Factors controlling sediment community respiration in woodland stream ecosys-tems. – Oikos 57: 94–105.
ETS Activity in Alder Leaf Litter 389
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com

HILL, B. H., J. M. LAZORCHAK, F. H. MCCORMICK and W. T. WILLINGHAM, 1997: The effects of elevat-ed metals on benthic community metabolism in a rocky mountain stream. – Environm. Poll. 95:183–190.
HILL, B. H., A. T. HERLIHY, P. R. KAUFMANN and R. L. SINSABAUGH, 1998: Sediment microbial respi-ration in a synoptic survey of Mid-Atlantic region streams. – Freshw. Biol. 39: 493–501.
HILL, B. H., R. K. HALL, P. HUSBY, A. T. HERLIHY and M. DUNNE, 2000: Interregional comparisons ofsediment microbial respiration in streams. – Freshw. Biol. 44: 213–222.
HILL, B. H., A. T. HERLIHY and P. R. KAUFMANN, 2002: Benthic microbial respiration in AppalachianMountain, Piedmont, and Coastal Plain streams of the eastern U.S.A. – Freshw. Biol. 47: 185–194.
JONES, J. B. Jr. and R. M. HOLMES, 1996: Surface-subsurface interactions in stream ecosystems. – TrendsEcol. & Evol. 1: 239–242.
JONSSON, M., B. MALMQVIST and P. O. HOFFSTEN, 2001: Leaf litter breakdown rates in boreal streams:does shredder species richness matter? – Freshw. Biol. 46: 161–171.
KENNER, R. A., and S. I. AHMED, 1975: Measurements of electron transport activities in marine envi-ronments. – Mar. Biol. 33: 119–127.
KAUSHIK, N. K., and H. B. N. HYNES, 1971: The fate of the dead leaves that fall into streams. – Arch.Hydrobiol. 68: 465–515.
LEICHTFRIED, M., 1985: Organic matter in gravel streams (Project RITRODAT-Lunz). – Verh. Internat.Verein. Limnol. 22: 2058–2062.
LEICHTFRIED, M., 1986: Räumliche und zeitliche Verteilung von particulärer organischer Substanz (POM– Particulate Organic Matter) in einem Gebirgsbach als Energiebasis der Biozönose. – PhD thesis,Univ. Vienna, 360 pp.
LEICHTFRIED, M., 1988: Bacterial substrates in gravel beds of a second order alpine stream (ProjectRitrodat – Lunz, Austria). – Verh. Internat. Verein. Limnol. 23: 1325–1332.
LEICHTFRIED, M., 1995: Organic matter in bedsediments – an energy source for lotic ecosystems (a com-pilation of a long therm study). – Folia Fac. Nat. Univ. Masaryk. Brun., Biologia 1: 77–93.
LEICHTFRIED, M., 1999: Biofilme in Oberflächengewässern. – Schriftenreihe d. österr. Wasser- undAbfallwirtschaftsverband 127: 101–112.
LYNCH, J. M. and J. E. HOBBIE (eds.), 1988: Microorganisms in Action: Concepts and Applications inMicrobial Ecology. – Blackwell Scientific Publications: 363 pp
MARSHALL, K. C. (ed.), 1984: Microbial Adhesion and Aggregation. – Life Sciences Res. Reports 31,Dahlem Workshops Rep., Springer Verlag: 423 pp.
MENENDEZ, M., O. HERNANDEZ and F. A. COMIN, 2003: Seasonal comparisons of leaf processing ratesin two Mediterranean rivers with different nutrient availability. – Hydrobiol. 495: 159–169.
MEYER, J. L. and C. JOHNSON, 1983: The influence of elevated nitrate concentration on rate of leafdecomposition in a stream. – Freshw. Biol. 13: 177–183.
MOSER, H., 1991: Input of organic matter (OM) in a low order stream (Ritrodat-Lunz study area, Aus-tria). – Verh. Internat. Verein. Limnol. 24: 1913–1916.
MOSER, H., 1992: Oberflächeneintrag und Verfrachtung organischer Substanz bei einem Gebirgsbach.– PhD thesis, Univ. Vienna, 137 pp.
MULHOLLAND, P. J., J. W. ELWOOD, J. D. NEWBOLD, J. R. WEBSTER and L. A. FERREN, 1984: Phospho-rus uptake by decomposing leaf detritus: effect of microbial biomass and activity. – Verh. Internat.Verein. Limnol. 22: 1899–1905.
MULHOLLAND, P. J., C. S. FELLOWS, J. L. TANK, N. B. GRIMM, J. R. WEBSTER, S. K. HAMILTON, E.MARTI, L. R. ASHKENAS, W. B. BOWDEN, W. K. DODDS, W. H. MCDOWELL, M. J. PAUL and B. J.PETERSON, 2001: Inter-biome comparison of factors controlling stream metabolism. – Freshw. Biol.46: 1503–1517.
NEWMAN R. L., J. A. PERRY, E. L. TAM and R. L. CRAWFORD, 1987: Effects of chronic chlorine expo-sure on litter processing in outdoor experimental streams. – Freshw. Biol. 18: 415–428.
ODUM, W. E., P. W. KIRK and J. C. ZIEMAN, 1978: Nonprotein nitrogen compounds associated with par-ticles of vascular plant detritus. – Oikos 32: 363–367.
OWENS, T. G. and F. D. KING, 1975: The measurement of respiratory electron transport system activityin marine zooplankton. – Mar. Biol. 30: 27–36.
PACKARD, T. T., 1971: The measurement of respiratory electron transport activity in marine phyto-plankton. – J. Mar. Res. 29: 235–244.
PACKMAN, A. I. and J. S. MACKAY, 2003: Interplay of stream-subsurface exchange, clay particle depo-sition, and streambed evolution. – Water Resour. Res. 39 (4) Art. No. 1097.
390 T. FLEITUCH and M. LEICHTFRIED
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com

PASCOAL, C., F. CASSIO and L. MARVANOVA, 2005: Anthropogenic stress may affect aquatichyphomycete diversity more than leaf decomposition in a low-order stream. – Arch. Hydrobiol. 162:481–496.
PERRY, J. A., N. H. TROELSTRUP, M. NEWSOME and B. SHELLEY, 1987: Whole ecosystem manipulationexperiments: the search for generality. – Water Science and Technology 19: 55–71.
PETERSEN, R. C. and K. W. CUMMINS, 1974: Leaf processing in a woodland stream. – Freshwat. Biol.4: 343–368.
POOLE, G. C. and C. H. BERMAN, 2001: An ecological perspective on in-stream temperature: natural heatdynamics and mechanisms of human-caused thermal degradation environmental management. – En-viron. Manag. 27: 787–802.
POZO, J., 1993: Leaf litter processing of alder and eucalyptus in the Aguera stream system (North Spain):I. chemical changes. – Arch. Hydrobiol. 127: 299–317.
RICE, D. L., 1982: The detritus nitrogen problem: new observations and perspectives from organic geo-chemistry. – Mar. Ecol. Prog. Ser. 9: 153–162.
ROWE, J. M., S. K. MEEGAN, E. S. ENGSTROM, S. A. PERRY and W. B. PERRY, 1996: Comparison of leafprocessing rates under different temperature regimes in three headwater streams. – Freshw. Biol. 36:277–288.
ROYER, T. V. and G. W. MINSHALL, 2001: Effects of nutrient enrichment and leaf quality on the break-down of leaves in a hardwater stream. – Freshw. Biol. 46: 603–610.
SHORT, R. A., S. P. CANTON and J. V. WARD, 1980: Detrital processing and associated macroinverte-brates in a Colorado mountain stream. – Ecology 61: 727–732.
SIMCIC, T., 2005: The role of plankton, zoobenthos, and sediment in organic matter degradation in olig-otrophic and eutrophic mountain lakes. – Hydrobiol. 532: 69–79.
SINSABAUGH, R. L. and S. FINDLAY, 1995: Microbial production, enzyme activity, and carbon turnoverin surface sediments of the Hudson River estuary. – Microb. Ecol. 30: 127–141.
SINSABAUGH, R. L., 1997: Large-scale trends for stream benthic respiration. – J. N. Am. Benthol. Soc.16: 119–122.
STRAHLER, A. N., 1957: Quantitative analysis of watershed geomorphology. – Am. Geophys. UnionTrans. 38: 913–920.
SUBERKROPP, K. and M. J. KLUG, 1976: Fungi and bacteria associated with leaves during processing ina woodland stream. – Ecology 57: 707–719.
SUBERKROPP, K., 1991: Relationships between growth and sporulation of aquatic hyphomycetes ondecomposing leaf litter. – Mycol. Res. 95: 843–850.
SUBERKROPP, K. and E. CHAUVET, 1995: Regulation of leaf breakdown by fungi in streams: Influencesof water chemistry. – Ecology 76: 1433–1445.
SZABO, E., 2003: The use of the tetrazolium reduction test for the detection of the terminal electrontransport system (ETS) activity in decomposing reed (Phragmites australis/CAV./TRIN. ex STEUD.) rhi-zome. – Ann. Limnol. 39: 63–70.
WAGNER, F. H. and G. BRETSCHKO, 2002: Interstitial flow through preferential flow paths in thehyporheic zone of the Oberer Seebach, Austria. – Aquat. Sci. 64: 307–316.
WEBSTER, J. R. and E. F. BENFIELD, 1986: Vascular plant breakdown in freshwater ecosystems. – Ann.Rev. Ecol. Syst. 17: 567–594.
WEBSTER, J. R., J. B. WALLACE, and E. F. BENFIELD, 1995: Organic processes in streams of the easternUnited States. – In: CUSHING, C. E., K. W. CUMMINS and G. W. MINSHALL (eds.), River and StreamEcosystems. – Elsevier Science, Amsterdam: pp. 117–187.
YOUNG, R. G. and A. D. HURYN, 1999: Effects of land use on stream metabolism and organic matterturnover. – Ecol. Appl. 9: 1359–1376.
YOUNG, R. G., A. J. QUARTERMAN, R. F. EYLES, R. A. SMITH and W. B. BOWDEN, 2005: Water qualityand thermal regime of the Motueka River: influences of land cover, geology and position in thecatchment. – New Zeal. J. Mar. Freshw. Res. 39: 803–825.
Manuscript received April 3rd, 2006; revised February 5th, 2007; accepted April 18th, 2007
ETS Activity in Alder Leaf Litter 391
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com





























