Embed Size (px)
Citation preview

Proc. Nat. Acad. Sci. USAVol. 72, No. 12, pp. 5056-5060, December 1975Genetics
Electron microscopic observations on the meiotic karyotype ofdiploid and tetraploid Saccharomyces cerevisiae
(synaptonemal complex/recombination nodules)
BRECK BYERS AND LORETTA GOETSCHDepartment of Genetics, University of Washington, Seattle, Wash. 98195
Communicated by Herschel L. Roman, September 26,1975
ABSTRACT Certain strains of Saccharomyces cerevisiaecontain visible segments of synaptonemal complex which areapparent components of bivslents in pachytene of meioticprophase. The synaptonemal complex has the typical widthin the frontal plane but is unusually thin in the sagittal plane,thus accounting for its poor visibility. Amorphous densitiessituated adjacent to the central element occur at intervalssuggesting their coincidence with sites of crossing over. Re-construction of the synaptonemal complex from serial sec-tions has permitted karyotypic analysis. The number of seg-ments of synaptonemal complex and the distribution of theirlengths is consistent with the genetic map. Two, possiblythree, segments enter the nucleolus as if bearing sequencesencoding ribosomal RNA. Reconstruction of tetraploid nucleireveals an approximate doubling of the diploid chromosomenumber and confirms the pattern of nucleolar entry. Quadri-valent pairing is evident between the pairs of synaptonemalcomplex segments in the tetraploid nuclei.
Although the budding yeast, Saccharomyces cerevisiae, hasproven highly amenable to genetic analysis, cytogeneticanalysis has been limited by the absence of a karyotype vis-ible by light microscopy. The greater resolution of electronmicroscopy has revealed several features of spindle behaviorduring nuclear division but has failed to reveal the individu-al chromosomes. Nevertheless, genetic studies of the sporesderived from meiosis indicate that yeast chromosomes prob-ably undergo synaptic behavior similar to that of other eu-karyotes. Meiotic nuclei have occasionally been found tocontain the synaptonemal complex (SC) both in aggregatesand as individual segments (1, 2). Recent observations byZickler and Olson (3) demonstrate the typical phases of me-iosis in yeast, confirming that the individual segments of SCrepresent components of pachytene bivalents as they do inother species. We have now achieved serial section electronmicroscopy of meiotic yeast adequate for reconstructing thedistribution of these segments of the synaptonemal complex.On this basis, we report here the numbers and lengths ofmeiotic chromosomes in diploid and tetraploid strains of S.cerevisiae.
MATERIALS AND METHODSStrains. The original diploid strain used for these observa-
tions is strain 212-1, constructed by Simchen (4). It is homo-zygous for the temperature-sensitive cell division cycle allelecdc 4-3 (5). This strain sporulates well, forming over 75%four-spored asci with viable spores after transfer from ace-tate growth medium to sporulation medium under permis-sive conditions, as described by Simchen (4). The tetraploidderivative, 212-1-T, was obtained by crossing clones thathad been induced by UV irradiation to become capable ofmating (by conversion of the mating type to aa or aa); thelatter were generously supplied by Ms. Hannah Klein. The
Abbreviation: SC, synaptonemal complex.
cells of 212-1-T were significantly larger than 212-1 andsporulated equally well at the permissive temperature (200),which was used for the observations reported here.
Electron Microscopy. Cells were fixed in glutaraldehyde,subjected to wall removal, further fixed with osmium tetrox-ide, and embedded for serial sectioning and electron micros-copy as described (6).
Reconstruction. The distribution of segments of SC wasdetermined by tracing their contours in micrographs (totalmagnification: 10,000 diameters) onto clear plastic sheets,which were then bound into booklets in appropriate register.Sketches of the courses followed by each bivalent were pre-pared as guides. We then determined lengths of each com-plete SC by summing segments of constant pitch. Segmentlengths were derived from the Pythagorean Theorem (c2 =a2 + b2) with measurements parallel to the sections as a anddepth (included section number times single section thick-ness of 100 nm) as b. Comparability of values for a and bwas tested by summing all values of each for an entire nu-cleus; these sums never varied by more than 10%, demon-strating that these two perpendicular projections from therandomly oriented chromosomes were indeed similar. Esti-mated errors of measurement were 10% for larger chromo-somes and 20% for shorter ones.
OBSERVATIONSAppearance of meiotic nucleiAlthough the synaptonemal complex (SC) is difficult to de-tect in many strains of yeast (1), we find it to be visible with-out difficulty in strain 212-1, which is homozygous for amutation (cdc 4) conferring temperature-sensitivity for bothmitosis (5) and meiosis (4). In meiotic arrest, 212-1 under-goes significant genetic recombination but fails to divide (4).Electron microscopy of arrested cells revealed aggregates ofSC [called "polycomplexes" (2)] as well as individual seg-ments of SC, which we assume to represent pachytene biva-lents. Attempts to reconstruct the distribution of these seg-ments in order to derive a yeast karyotype were frustratedby the presence of uninterpretable discontinuities. Wefound, however, that the SC was unusually visible in thisstrain not only during arrest but also between 16 and 20 hrin cells proceeding successfully through meiosis at the per-missive temperature, 200. The absence of detectable SC instrain 212, which differs from 212-1 in being heterozygousfor cdc 4 (4), suggests that it is the influence of the cdc 4 inhomozygous state at 200 that prolongs the appropriate stageof meiosis. During this stage at the permissive temperature,the segments of SC visible in 212-1 (Fig. la) do not show anydiscontinuities in serial sections. The visible profiles of mostidentifiable segments terminate at the nuclear envelope(Fig. Id). In agreement with studies of numerous other
5056
Proc. Nat. Acad. Sci. USA 72 (1975) 5057
g ., A,.b t a ^
~~~~(~~4,<fl.e
**;'wx*A~eO_ W~ v;eeewee}
All the' Ao*_
n~~~~~~1 [A .'
B.DBo.. f . ..._ Of. A. b
Be... Jt;,r+ R . ..
,*_*. . I,. ..
Wha; ;'st'k'-* I'd *s _ :Ks .... ,: ,_tab e£at('il'' 'P'sF A.. so ^ -07SAXS 4, I;*""*>-.1t>|* 5;J '
., _, ; S s ; -.
* .o t r }
*_ em amp- i-_L
s A; %'>.t X*, 4 And
W+s'a <* t '
,., *,- '
', .' _ . ..b;X, a. 'i. -.
%a ';';bS4.L.
-Va
4'~~~~~~~~~~~~~~~~.~ ~ ~ T
~ ~4AsS'
UC .2-
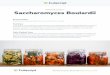
FIG. 1. Details of diploid nuclei during pachytene of meiosis at 200. (a) A nucleus bearing several segments of SC with dense nodules (N)adjacent to the central element. (b) The dense body (DB) and nucleolus (NU) together with a section of SC. (c) A double plaque intransverse section. (d) A telomere (T) in typical association with the nuclear envelope. (e) Three nodules (N) along a segment of SC. Scalebars, 0.2 Am.
species (7), we interpret this distribution of segments to rep-resent the telomeric association of pachytene bivalents withthe nuclear envelope. The few remaining ends of segmentslie at the margin of the nucleolus; the bivalent is indistinctwithin the apparent nucleolar organizing region, as it is inchromosome 2 of Neurospora (8).
These presumed pachytene nuclei invariably bear thedouble plaque (Fig. ic), or "side-by-side plaques" (2), pre-paratory to meiosis I. This stage of plaque development per-sists from about 12 to 20 hr and is accompanied by the pres-ence of a finely-fibrous dense body (DB, Fig. lb), similar tothe "polycomplex body" of Moens and Rapport (2). Aggre-gates of SC (polycomplexes) are rarely seen in cells incubat-ed at the permissive temperature.The segments of SC are occasionally indistinct in detail
but bear features typical of this organelle (Fig. 1). Frontalsections, as defined by Moses (7), display prominent lateralelements spaced 110 nm apart. Each lateral element is about25 nm thick in this view and is indistinguishable from chro-matin, which is as difficult to detect here as at any otherstage of the life cycle. The lateral element only occasionallydisplays the periodicity (20 nm) characteristic of some otherAscomycetes, like Neotiella (9). The central element is about20 nm wide, sometimes appearing as two parallel denselines; transverse filaments are faintly discernible across the50-nm clear zone between the central and lateral elements
(as in the centermost segment in Fig. la).Transverse sections (Fig. 4a, no. 26) demonstrate that the
SC is extremely narrow in the sagittal plane. The lateral ele-ment may be only 30 nm thick and the central element,where visible, is less than 20 nm thick. Sagittal sections re-veal no additional details; only the density of the lateral ele-ment makes these sections visible.
Also visible in frontal and transverse sections are amor-phous densities, 30-50 nm in diameter, situated adjacent tothe margins of the central element-that is, in the same rela-tionship to the SC as the more distinctive "recombinationnodules" of Drosophila melanogaster (10). These nodules(N, Fig. la) are undetectable in other angles of section, butwe have been able to estimate their average distributionalong the bivalents by determining their frequency in seg-ments of SC at certain angles of section (transverse and fron-tal) which permit calculation of SC segment length. In fa-vorable sets of serial sections of four nuclei, there were 1.72I 0.37 nodules per Am of bivalent. Therefore, they musttypically be spaced at intervals of about 0.6 1um when de-tectable, although the smallest interval encountered was 0.15uM (Fig. I e).Reconstruction of karyotypes from serial sectionsInterpreting the segments of SC as components of pachytenechromosomes, we undertook an analysis of the yeast
Genetics: Byers and Goetsch

5058 Genetics: Byers and Goetsch
4.
8 e N
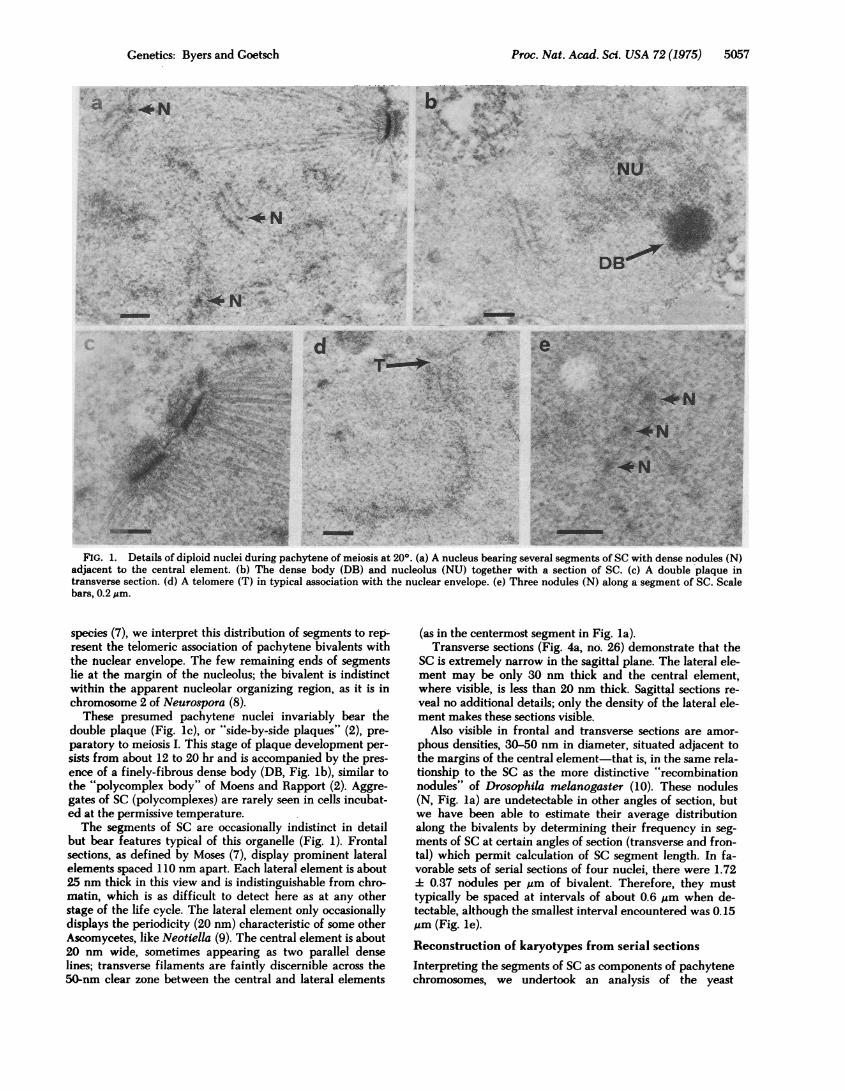
FIG. 2. Reconstruction of the segments of SC representing bi-valents in diploid nucleus D12. Wide lines, lateral elements;dashed lines, central elements; large digits, bivalent number (sameas in Fig. 3) by rank order of length; small digits, section numbers;stippling, nucleolus.
karyotype. Serial reconstructions were prepared of eight nu-
clei from a diploid (212-1) culture and five nuclei from a
tetraploid (212-1-T) culture fixed 18 hr after initiation ofmeiosis at 200. Among the sample of diploids, between 14and 17 bivalents were detected. If some of the shorter ones
were more likely to escape detection because of unfavorableangles of sectioning, the actual number of bivalents maywell be 17 or more. Table 1 presents the length measure-
ments from three of these diploids in which the contours ofall bivalents appeared to be unambiguous. A reconstructeddiploid nucleus (D12) bearing 17 identifiable bivalents is de-picted in Fig. 2, and the lengths of these bivalents are plot-ted in Fig. 3. Fifteen bivalents clearly show both telomeresadjacent to the nuclear envelope; two others have one telom-ere there and the other end disappearing into the nucleolus,where its course is indistinct. These latter two segments ofcomplex (of length rank order 8 and 14 in nucleus D12) may
4
IMIIXIIIII e
8 ---------
12!Ixlll
;NN208
24 8111
2 PM
31 |17- DIPILOID TETRAPLOID
FIG. 3. Contour lengths of SC segments in nucleus D12 (DIP-LOID) and nucleus T14 (TETRAPLOID). Arrows represent entryinto the nucleolus (N); all other ends are telomeres. Vertical linesrepresent parallel bivalents less than 0.3 ,m apart; X indicates a
probable site of change in pairing partners.
represent either one bivalent with a medial nucleolar orga-
nizer or two bivalents with terminal organizers and unde-tected telomeres (hidden within the nucleolus). In addition,a small subtelomeric portion of a longer bivalent (rank order3 in nucleus D12) passes through the margin of the nucleolusin this and other reconstructions. Considering the three re-
constructed diploids together (Table 1), we find that the de-finitive association with the nucleolus by two segments of SCis consistent. The overall lengths of SC in these three nucleivaries considerably, averaging 28 gim. However, the relativelength of the longest bivalent in comparison with the overalllength of the karyotype is rather constant (12.8-14.3%),whereas the median length, which is more subject to error in
measurement, varies relatively more (4.8-7.2%).The interpretations derived from diploid nuclei were test-
ed by making a comparative study of tetraploid nuclei. Inthe five tetraploid nuclei studied, the local anatomy of theSC (Fig. 4) appears identical, but reconstructions of the con-
tours (Fig. 5) reveal features unique to the tetraploid. Al-though some segments of SC occur separately from others as
expected of bivalents, the reconstructions (Table 1) all con-
tain regions where segments of SC similar in length run
nearly parallel to one another along part or all of their axes
Table 1. Lengths of SC segments in serial reconstruction
Diploids Tetraploids
Nucleus D9 D10 D12 T13 T14 T17
No. of SC segments 15 15 17 30 31 28Maximum length: gm 3.56 3.01 4.74 3.36 3.66 4.61
% 12.8 13.5 14.3 7.1 7.3 8.0Median length: ;Am 1.61 1.60 1.60 1.62 1.46 1.64
% 5.8 7.2 4.8 3.4 2.9 2.9Total length: ;Am 27.7 22.4 33.1 47.1 50.3 57.4No. entering nucleolus:
definitive 2 2 2 4 4 4(partial) (0) (0) (1) (1) (2) (2)
No. of quadrivalent pairs 9 10 10Average variance in lengthof pairs: % - - 13.7 4.3 5.4
Proc. Nat. Acad. Sci. USA 72 (1975)
Proc. Nat. Acad. Sci. USA 72 (1975) 5059~~'Oab ~ 88 ~ ; 4
,A
i. k tA.
C
an- <124w
b
17 -A'i
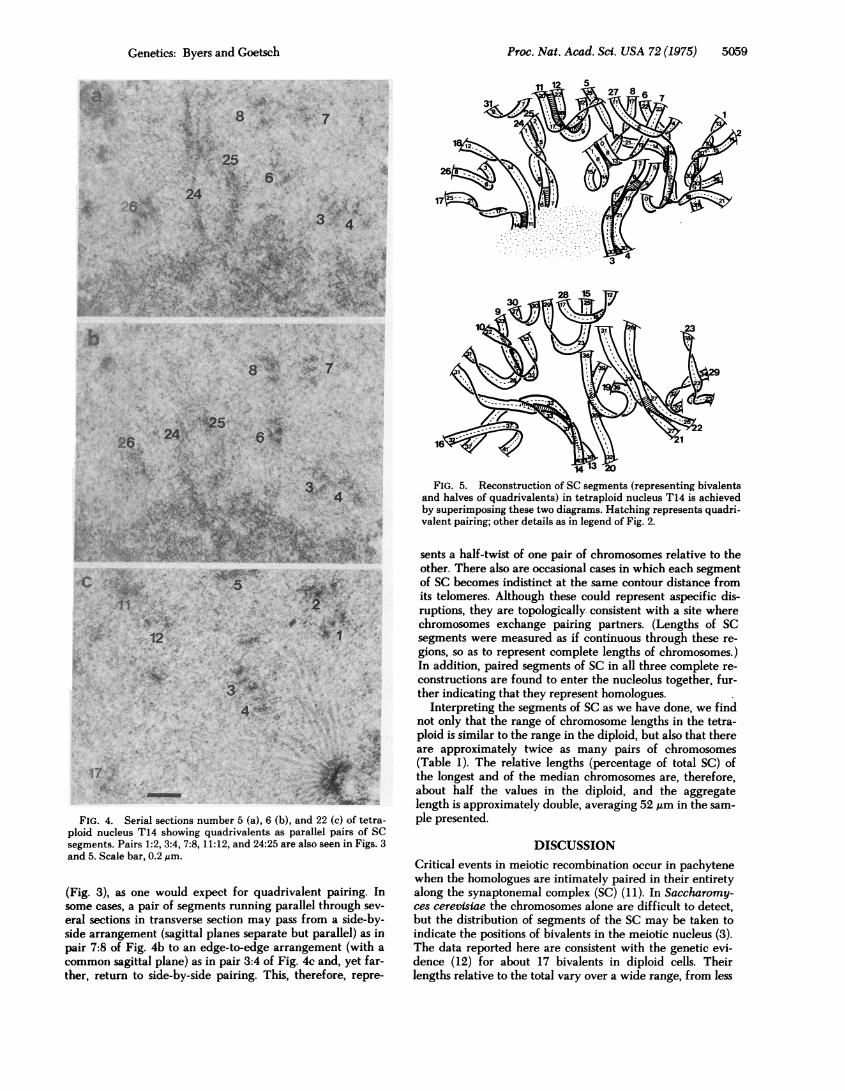
FIG. 4. Serial sections number 5 (a), 6 (b), and 22 (c) of tetra-ploid nucleus T14 showing quadrivalents as parallel pairs of SCsegments. Pairs 1:2, 3:4, 7:8, 11:12, and 24:25 are also seen in Figs. 3and 5. Scale bar, 0.2 ,um.
(Fig. 3), as one would expect for quadrivalent pairing. Insome cases, a pair of segments running parallel through sev-
eral sections in transverse section may pass from a side-by-side arrangement (sagittal planes separate but parallel) as inpair 7:8 of Fig. 4b to an edge-to-edge arrangement (with a
common sagittal plane) as in pair 3:4 of Fig. 4c and, yet far-ther, return to side-by-side pairing. This, therefore, repre-
FIG. 5. Reconstruction of SC segments (representing bivalentsand halves of quadrivalents) in tetraploid nucleus T14 is achievedby superimposing these two diagrams. Hatching represents quadri-valent pairing; other details as in legend of Fig. 2.
sents a half-twist of one pair of chromosomes relative to theother. There also are occasional cases in which each segmentof SC becomes indistinct at the same contour distance fromits telomeres. Although these could represent aspecific dis-ruptions, they are topologically consistent with a site wherechromosomes exchange pairing partners. (Lengths of SCsegments were measured as if continuous through these re-gions, so as to represent complete lengths of chromosomes.)In addition, paired segments of SC in all three complete re-constructions are found to enter the nucleolus together, fur-ther indicating that they represent homologues.
Interpreting the segments of SC as we have done, we findnot only that the range of chromosome lengths in the tetra-ploid is similar to the range in the diploid, but also that thereare approximately twice as many pairs of chromosomes(Table 1). The relative lengths (percentage of total SC) ofthe longest and of the median chromosomes are, therefore,about half the values in the diploid, and the aggregatelength is approximately double, averaging 52 ,gm in the sam-ple presented.
DISCUSSIONCritical events in meiotic recombination occur in pachytenewhen the homologues are intimately paired in their entiretyalong the synaptonemal complex (SC) (11). In Saccharomy-ces cerevisiae the chromosomes alone are difficult to detect,but the distribution of segments of the SC may be taken toindicate the positions of bivalents in the meiotic nucleus (3).The data reported here are consistent with the genetic evi-dence (12) for about 17 bivalents in diploid cells. Theirlengths relative to the total vary over a wide range, from less
Genetics: Byers and Goetsch
0.IA I
.4 ..
I1.
I't
. 119
r'i-
.I f".
A. ",
.4 V., II I ,,

5060 Genetics: Byers and Goetsch
than 2% to nearly 14%, with an average length of about1.6-1.8 ,gm; a similar range of relative magnitudes is foundin the total map distances of the genetic linkage groups (12).Furthermore, the entry into the nucleolus by segments of SCmay be taken to represent the karyotypic location of nucleo-lar organizers. Hybridization of ribosomal RNA to fraction-ated chromosomal DNA from aneuploid strains suggests theoccurrence of more than one chromosome with rDNA se-quences (13), the majority of these sequences being on chro-mosome I (14). Although our inability to follow the SC with-in the nucleolus prevents determination of overall length(and possible continuities) of bivalents, the unambiguousentry by two segments may well represent the presence ofrDNA sequences at the ends of two bivalents. The additionalassociation of a short segment of a larger bivalent within thenucleolus may represent a few rDNA sequences on yet athird chromosome.The observations on the diploid karyotype are confirmed
by study of the tetraploid derivative, which shows a similardistribution of chromosomal lengths as well as an approxi-mate doubling of their number. Moreover, these reconstruc-tions of tetraploid nuclei also demonstrate the occurrence ofquadrivalent pairing, as predicted from the genetics of tetra-ploids (15). Quadrivalents are seen as paired segments of SCof similar length running parallel through the nucleus withoccasional interruptions at equivalent lineal positions. Theseinterruptions may well represent sites of exchange in pairingpartners, although the axial elements are not sufficiently dis-tinct to establish the courses of the individual chromatids, asis possible in Lilium (16). Perhaps the most definitive evi-dence for quadrivalents is that the pattern of their entry intothe nucleolus is the same as that shown by individual biva-lents in the diploid.The relative distribution of individual SC lengths in these
karyotypes approximates that seen in sucrose gradient sedi-mentation of yeast chromosomal DNA (17). Excluding theundetected SC within the nucleolus, the composite length ofSC in the diploids reported here averages 28 ,um, and that inthe tetraploids is 52 ,m. Based on a haploid nuclear genomeof about 5.6 X 109 daltons of DNA (18), the total length ofDNA helix for each haploid equivalent must be 2.8 X 103,um, yielding a packing ratio of 1:100, Aim of SC:,um of DNA.By comparison, Drosophila melanogaster has a haploidDNA content of 5.5 X 104 ,um of DNA (19) and 50,m ofeuchromatic SC at mid-pachytene (10), for an approximatepacking ratio of 1:1000. Yeast therefore must have aboutone-tenth as much DNA along its SC; this may account forour observation that the SC of yeast is several-fold thinnerthan that of Drosophila.
This relative deficiency of DNA along the bivalent inyeast is not accompanied by a lower frequency of recombi-nation. On the contrary, yeast undergoes a relatively highlevel of recombination per length of SC. Whereas the aver-age distribution of crossovers in Drosophila is about one per9 ,um of euchromatic SC (10), their distribution in Saccharo-myces cerevlsiae must be at intervals of only 0.4 ,um alongthe bivalent [28 Am of bivalents undergo about 70 crossovers(12)]. Moreover, although the amorphous densities adjacentto the central element in these yeast nuclei are less distinctthan the "recombination nodules" of Drosophila (10) or the"nodes" of Neurospora (8), the distribution of these densitiesin yeast again suggests that they represent sites of meiotic re-combination.
We are grateful to Dr. Giora Simchen and Ms. Hannah Klein forproviding strains. This research was supported by National Insti-tutes of Health Grant GM-18541 and N.I.H. Research Career De-velopmerit Award GM-20790 to B.B.
1. Engels, F. M. & Croes, A. F. (1968) Chromosoma 25, 104-106.
2. Moens, P. B. & Rapport, E. (1971) J. Cell Sci. 9, 665-677.3. Zickler, D. & Olson, L. W. (1975) Chromosoma 50, 1-23.4. Simchen, G. (1974) Genetics 76,745-753.5. Hartwell, L. H., Mortimer, R. K., Culotti, J. & Culotti, M.
(1973) Genetics 74,267-286.6. Byers, B. & Goetsch, L. (1975) J. Bacteriol., 124,511-523.7. Moses, M. J. (1968) Annu. Rev. Genet. 2,363-412.8. Gillies, C. B. (1972) Chromosoma 36, 119-130.9. von Wettstein, D. (1971) Proc. Nat. Acad. Sci. USA 68, 851-
855.10. Carpenter, A. T. C. (1975) Proc. Nat. Acad. Sci. USA 72,
3186-3189.11. Stern, H. & Hotta, Y. (1973) Annu. Rev. Genet. 7,37-66.12. Mortimer, R. K. & Hawthorne, D. C. (1973) Genetics 74, 33-
54.13. de Kloet, S. R. (1973) J. Bacteriol. 114, 1034-1039.14. Finkelstein, D. B., Blamire, J. & Marmur, J. (1972) Nature
New Biol. 240,279-281.15. Roman, H., Phillips, M. M. & Sands, S. M. (1955) Genetics 40,
546-561.16. Moens, P. B. (1970) J. Cell Sci. 7,55-63.17. Petes, T. D., Byers, B. & Fangman, W. L. (1973) Proc. Nat.
Acad. Sci. USA 70,3072-3076.18. Whitney, P. & Hall, B. D. (1974) Fed. Proc. Fed. Am. Soc.
Exp. Biol. 33, 1282a.19. Rasch, E. M., Barr, H. J. & Rasch, R. W. (1971) Chromosoma
33, 1-18.
Proc. Nat. Acad. Sci. USA 72 (1975)