Embed Size (px)
Citation preview

JOURNAL OF BACrERIOLOGY, Jan. 1969, p. 448452 Vol. 97, No. ICopyright © 1969 American Society for Microbiology Printed In U.S.A.
Electron Microscopic Studies on Mode of Actionof Polymyxin
M. KOIKE, K. IIDA, AND T. MATSUODepartment of Bacteriology, School of Medicine, Kyushu University, Fukuoka, Japan
Received for publication 12 October 1968
The antibacterial activity of the polymyxingroup against gram-negative bacteria has beenexplained as owing to increased permeability ofthe cell envelope consisting of cell wall and cyto-plasmic membrane and the resultant leakage ofthe cell contents (7). Consequently, morpho-logical studies should contribute much to theelucidation of the mechanism of action of theseagents (2). We have made detailed observationsof the changes' in the cell envelope caused bypolymyxin B and colistin, using Escherichia coli Band Pseudomonas aeruginost P29 (furnished byY. Homma, Institute of Medical Science, TokyoUniversity).The cells of these gram-negative bacteria, grown
in Nutrient Broth (Difco), were harvested in thelogarithmic phase by centrifugation and washingwith tris(hydroxymethyl)aminomethane buffer(pH 7.2), and they were finally suspended in thesame buffer to give 5 X 108 viable cells per fnl.The cell suspension was added to one-ninthvolume of polymyxin B sulfate (PLB, Taito-phizer Co., Tokyo, Japan), colistin sulfate, orcolistin methanesulfonate (CL and CLM, KakenCo., Tokyo, Japan) solutions of various concen-trations. After incubation at 37 C for various timeintervals, each mixture was examined for theviable cell count and prepared for electronmicroscopy. A cell wall fraction from E. coli Bdisrupted in a French pressure disintegrator cellwas also treated with PLB as described above. Ac-tion of PLB on the spheroplasts of E. coli B ob-tained by Repaske's procedure (8) was measuredby the decrease of optical density of the sphero-plast suspension at 660 nm. For electron micros-copy, the cells were doubly fixed with 1% glutar-aldehyde solution and 1 % OS04 solution,dehydrated through the graded alcohol solution,and embedded in Epon 812. The specimens werecut with a Porter-Blum MT2 Ultramicrotome andexamined in a JEM7 electron microscope. Thecell wall fraction was negatively stained withphosphotungstate (pH 7.0).The viability of the cells of these bacteria was
reduced by 10-s or less within 10 min contact with25 ,ug of PLB per ml and CL and 250 ,ug of CLMper ml. Figure la shows a section of the un-
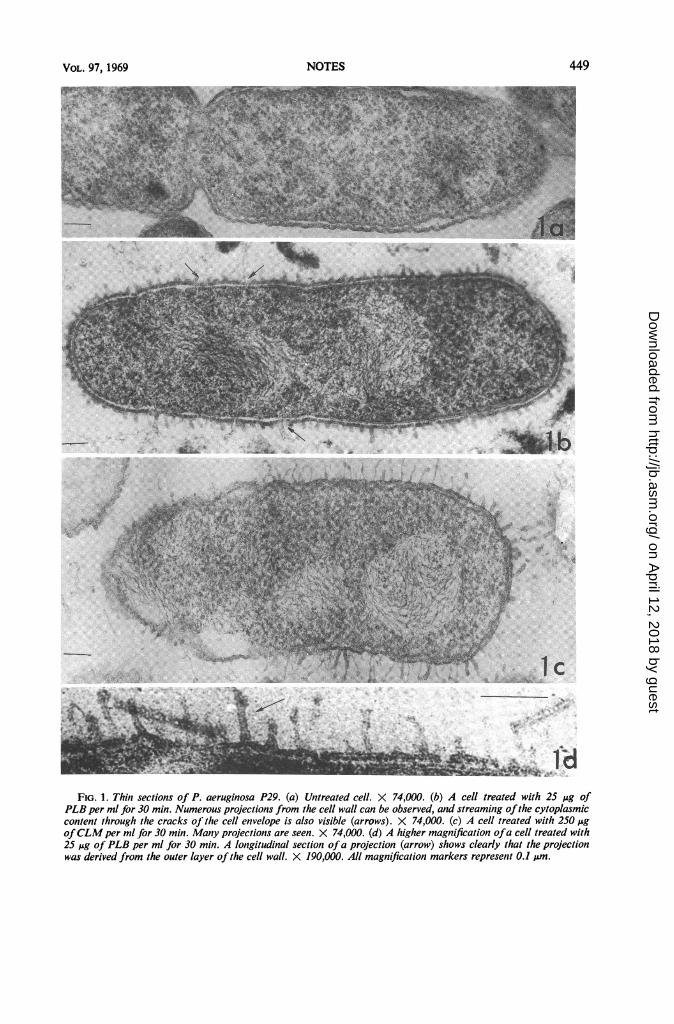
treated P. aeruginosa P29. Figure lb is a sectionof the same organism treated with 25 Mug of PLBper ml for 30 min. Figure lc is the same bacteriumtreated with 250 Mg of CLM per ml for 30 min.Numerous projections appear on the cell wall; thecytoplasmic membrane appears to be damaged,and part of the cytoplasmic material is released infibrous forms through the cracks (Fig. lb).Figure ld shows a magnified part of a treated cell;the outer layer of the cell wall bears projectionsas wide as 12 to 14 nm.
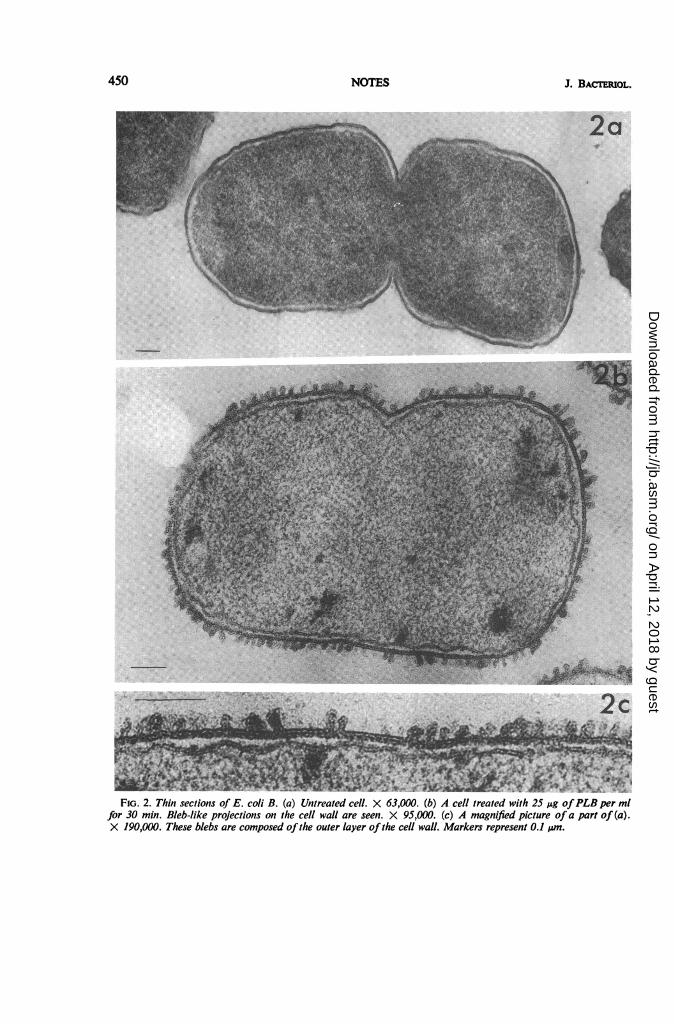
Figure 2 illustrates the appearance of E. colicells treated in the same way. In the case of E. coliB, in contrast to P. aeruginosa, the projectionsappear to be blebs derived from outer layers of thecell wall (Fig. 2c). Although such blebs areoccasionally seen even in normal bacteria, theiroccurrence in such large numbers is apparentlydue to the action of PLB. The number of theprojections decreased with reduction in concen-tration of the drugs, and this reduction was paral-leled by the higher residual viable cell count. Theantagonistic action of Mg++ to PLB (5) wascorrelated with the electron microscopically ob-servable reduction in the number of the projec-tions.These projections were also clearly visible on
the purified cell wall fraction treated with PLB.Normal cell wall has an outer layer with a smoothsurface (Fig. 3a, 3b), whereas those subjected tothe action of PLB have numerous projections ontheir surface (Fig. 3c, 3d).
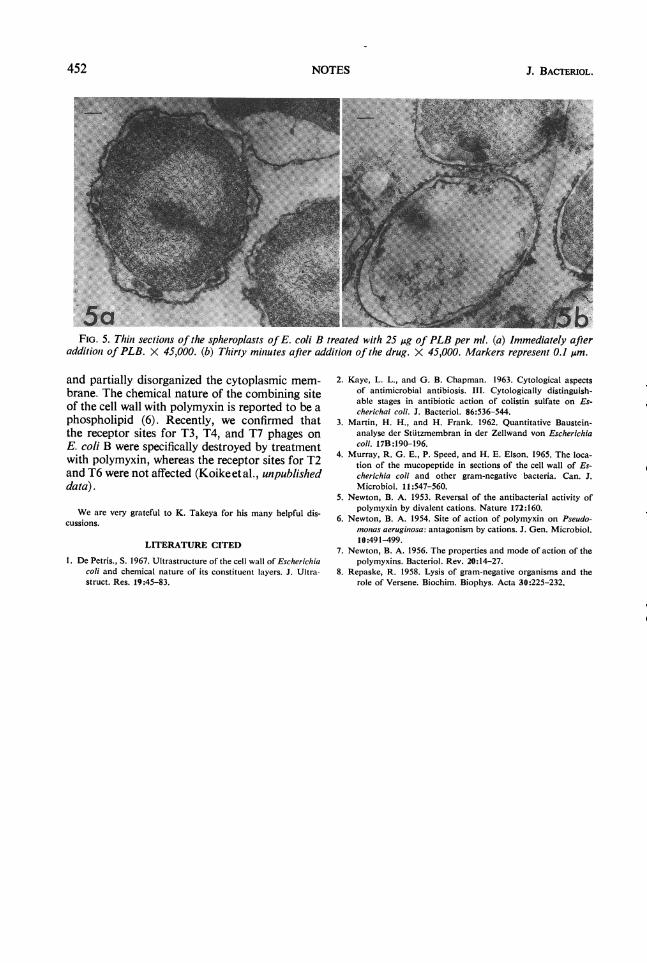
It has been shown that polymyxin acts not onlyon the cell wall but also on the cytoplasmic mem-brane (Fig. lb). Observations of the effect onspheroplasts revealed that lysis occurred rapidlyafter addition of PLB (Fig. 4). Fig. 5a shows apicture immediately after addition of PLB; blebshave been formed apparently on the outer layerbut no changes are observed in the cytoplasm.After exposure for 30 min, the protoplast lost itscontents and the cytoplasmic membrane was dis-organized (Fig. 5b).The cell wall of gram-negative bacteria is com-
posed of three layers (1, 3, 4). These experimentsrevealed that polymyxin-caused projections origi-nated from the outermost layer of the three layers
448
on April 12, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
VOL. 97, 1969
'O
- . .7.!r* &K.. .i 7
5B~~~~~~~~~~~~~~~~~~~~~ ;.i' :::s.*j'% "% si.t "-'~~~~~~~~ 2~..wn
I
J" .:*l-FIG. 1. Thin sections of P. aeruginosa P29. (a) Untreated cell. X 74,000. (b) A cell treated with 25 ,jg of
PLB per ml for 30 min. Numerous projections from the cell wall can be observed, and streaming ofthe cytoplasmiccontent through the cracks of the cell envelope is also visible (arrows). X 74,000. (c) A cell treated with 250 AgofCLM per ml for 30 min. Many projections are seen. X 74,000. (d) A higher magnification ofa cell treated with25 jsg oJ PLB per ml for 30 min. A longitudinal section ofa projection (arrow) shows clearly that the projectionwas derived from the outer layer of the cell wall. X 190,000. All magnification markers represent 0.1 m.
449NOTES
1- 4i.zl4:
I
11
lil I v.,1,
- "' .". `11..11:., -4..
t, 11 14. f t " i i :.
vel.
A'144-
O", '.,ft. "if I .:
..401
t
,,if''Y
.1 AL.
on April 12, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
NOTES J. BACTERIOL.
.
A..
. .'.
FIG. 2. Thin sections of E. coli B. (a) Untreated cell. X 63,000. (b) A cell treated with 25 uAg ofPLB per mlfor 30 min. Bleb-like projections on the cell wall are seen. X 95,000. (c) A magnified picture of a part of (a).X 190,000. These blebs are composed ofthe outer layer ofthe cell wall. Markers represent 0.1 um.
450
on April 12, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
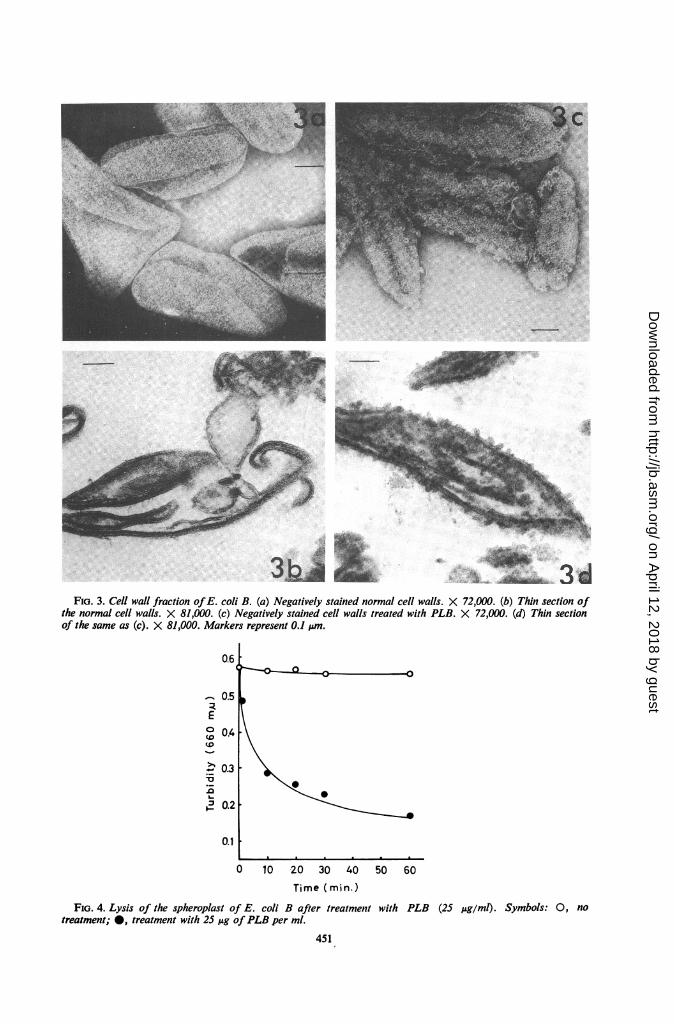
FIG. 3. Cell wall fraction ofE. coli B. (a) Negatively stained normal cell walls. X 72,000. (b) Thin section ofthe normal cell walls. X 81,000. (c) Negatively stained cell walls treated with PLB. X 72,000. (d) Thin sectionof the same as (c). X 81,000. Markers represent 0.1 ,um.
0.6
0.5
°0.4 \
0.3 -
i2 0.2
0.1
0 10 20 30 40 50 60
Time (min.)
FIG. 4. Lysis of the spheroplast of E. coli B after treatment with PLB (25 Ag/ml). Symbols: 0, notreatment; 0, treatment with 25 jug ofPLB per ml.
451
on April 12, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
452 NOTES J. BACTERIOL.
FIG. 5. Thin sections of the spheroplasts of E. coli B treated with 25 Ag of PLB per ml. (a) Immediately afteradditionz of PLB. X 45,000. (b) Thirty miniutes after addition of the drug. X 45,000. Markers represent 0.1 ,um.
and partially disorganized the cytoplasmic mem-brane. The chemical nature of the combining siteof the cell wall with polymyxin is reported to be aphospholipid (6). Recently, we confirmed thatthe receptor sites for T3, T4, and T7 phages onE. coli B were specifically destroyed by treatmentwith polymyxin, whereas the receptor sites for T2and T6 were not affected (Koikeetal., unpublisheddata).
We are very grateful to K. Takeya for his many helpful dis-cussions.
LITERATURE CITED
1. De Petris., S. 1967. Ultrastructure of the cell wall of Escherichiacoli and chemical nature of its constituent layers. J. Ultra-struct. Res. 19:45-83.
2. Kaye, L. L., and G. B. Chapman. 1963. Cytological aspectsof antimicrobial antibiosis. III. Cytologically distinguish-able stages in antibiotic action of colistin sulfate on Es-cherichai coli. J. Bacteriol. 86:536-544.
3. Martin, H. H., and H. Frank. 1962. Quantitative Baustein-analyse der Stuitzmembran in der Zellwand von Escherichiacoli. 17B:190-196.
4. Murray, R. G. E., P. Speed, and H. E. Elson. 1965. The loca-tion of the mucopeptide in sections of the cell wall of Es-cherichia coli and other gram-negative bacteria. Can. J.Microbiol. 11:547-560.
5. Newton, B. A. 1953. Reversal of the antibacterial activity ofpolymyxin by divalent cations. Nature 172:160.
6. Newton, B. A. 1954. Site of action of polymyxin on Pseudo-monias aeruginosa: antagonism by cations. J. Gen. Microbiol.10:491-499.
7. Newton, B. A. 1956. The properties and mode of action of thepolymyxins. Bacteriol. Rev. 20:14-27.
8. Repaske, R. 1958. Lysis of gram-negative organisms and therole of Versene. Biochim. Biophys. Acta 30:225-232.
on April 12, 2018 by guest
http://jb.asm.org/
Dow
nloaded from





























