Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Zhuo Li Æ Xujia Zhang
Electron-microscopic structure of the V-ATPase from mung bean
Received: 30 December 2003 / Accepted: 22 April 2004 / Published online: 5 June 2004� Springer-Verlag 2004
Abstract The vacuolar H+-ATPase from mung bean(Vigna radiata L. cv. Wilczek) was purified to homoge-neity. The purified complex contained all the reportedsubunits from mung bean, but also included a 40-kDasubunit, corresponding to the membrane-associatedsubunit d, which has not previously been observed. Thestructure of the V-ATPase from mung bean was studiedby electron microscopy of negatively stained samples.An analysis of over 6,000 single-particle images obtainedby electron microscopy of the purified complex revealedthat the complex, similar to other V-ATPases, is orga-nized into two major domains V1 and Vo with overalldimensions of 25 nm·13.7 nm and a stalk region con-necting the V1 and Vo domains. Several individual areasof protein density were observed in the stalk region,indicating its complexity. The projections clearly showedthat the complex contained one central stalk and at leasttwo peripheral stalks. Subcomplexes containing subun-its A, B and E, dissociated from the tonoplast mem-brane by KI, were purified. The structure of thesubcomplex was also studied by electron microscopyfollowed by single-molecule analysis of 13,000 projec-tions. Our preliminary results reveal an area of highprotein density at the bottom of the subcompleximmediately below the cavity formed by the A andB subunits, indicating the position of subunit E.
Keywords Electron microscopy Æ Single-particleanalysis Æ Vacuolar H+-ATPase Æ Vigna
Abbreviations MSA: Multivariate statisticalanalysis Æ 2D, 3D: Two-, three-dimensional Æ V-ATPase:Vacuolar H+-ATPase
Introduction
The vacuolar H+-ATPase, or V-ATPase, is a primaryactive pump located at the vacuolar membrane (tono-plast) of plant cells (Marin 1985; Sze 1985; Sze et al.1992). By hydrolyzing ATP, the V-ATPase generates anelectrochemical gradient across the membrane and pro-vides a driving force for secondary transport processescontributing to osmoregulation, ion and pH homeosta-sis, nutrient and remnant storage, and plant defense(Finbow and Harrison 1997; Ratajczak 2000).
The V-ATPase is a multisubunit complex consistingof two distinct domains: a transmembrane protonchannel (Vo) and an attached catalytic complex (V1)(Nishi and Forgac 2002). The overall structure of the V-ATPase is similar to that of the well-characterized F-ATPase (Abrahams et al. 1994; Stock et al. 1999). Thesoluble V1 domain is connected via a stalk region to themembrane-integrated Vo domain. Because of the struc-tural similarity, V-ATPases are believed to functionthrough a binding change mechanism similar to F-ATPases (Boyer 1999). ATP hydrolysis in the V1 domaindrives the rotation of the central stalk with respect to theA and B subunits of V1, which in turn drives the rota-tion of the ring of c subunits of the Vo domain againstthe static part of Vo, leading to proton translocationacross the membrane.
The exact subunit composition and stoichiometryofV-ATPases, however, is still controversial and varies be-tween plant species (Sze et al. 1999; Ratajczak 2000). TheV-ATPase from yeast has beenwell characterized (Forgac1999). Its V1 domain is composed of eight different su-bunits (subunits A–H), which are responsible for ATPhydrolysis; and the Vo domain is composed of five dif-ferent subunits (subunits a, d, c, c¢ and c¢¢), which areresponsible for proton translocation. Homologues of thec¢and c¢¢ subunits have not yet been identified in plants.Moreover, in mung bean and pea, the subunits other thanA, B, D, E and c have not been conclusively assigned(Matsuura-Endo et al. 1990; Kawamura et al. 2000).
Z. Li Æ X. Zhang (&)National Laboratory of Biomacromolecules,Institute of Biophysics, Chinese Academy of Sciences,100101 Beijing, ChinaE-mail: [email protected].: +86-10-64888517Fax: +86-10-64872026
Planta (2004) 219: 948–954DOI 10.1007/s00425-004-1298-2

Unlike F-ATPases, for which detailed knowledge ofthe structural organization of various domains is avail-able (Abrahams et al. 1994; Stock et al. 1999), little isknown about the structure of V-ATPases. The grossstructure and tentative subunit positions in the complexhave emerged through cross-linking studies (Grahamet al. 2000; Gruber et al. 2001) complemented by low-resolution structural information from electron micros-copy (Boekema et al. 1997, 1999; Wilkens et al. 1999;Ubbink-Kok et al. 2000; Domgall et al. 2002). Electronmicroscopy has confirmed that the isolated V1 domain isa hetero hexametric complex of alternating A andB subunits (Radermacher et al. 1999, 2001; Gruber et al.2000; Rizzo et al. 2003). The asymmetric protein ring, aswell as the 7-fold symmetry of the c ring, has beenvisualized by electron microscopy, demonstrating thatthe Vo domain is the rotor responsible for protontranslocation (Wilkens and Forgac 2001; Murata et al.2003). However, the remaining subunit positions havenot so far been convincingly determined within thestructures of the complexes. For instance, whether theV1 domain has two or three peripheral stalks is stillcontroversial (Boekema et al. 1997, 1999; Wilkens et al.1999; Ubbink-Kok et al. 2000; Domgall et al. 2002).Moreover, it has been proposed that the central stalkcontains the E subunit (Gruber et al. 2000; Chaban et al.2002; Rizzo et al. 2003), although recent cross-linkingexperiments have suggested that the E subunit mightcontribute to the peripheral stalk of the V-ATPase(Arata et al. 2002).
In the present study, we purified the V-ATPase frommung bean and performed single-particle analysis byelectron microscopy. Our purified complex contained a40-kDa subunit that was not observed in previous studies.Electron-microscopic images of the isolated V1 domaincontaining only A, B and E subunits support the sugges-tion that the E subunit is located at the central stalk.
Materials and methods
Materials
Fresh etiolated mung bean (Vigna radiata L. cv. Wil-czek) seedlings were purchased from a local market.Lysophosphatidylcholine (egg yolk, type I), L-a-phos-phatidylcholine (soybean, Type II-S) and ATP wereobtained from Sigma. All other chemical agents wereA.R. grade available in P.R. China.
Purification of the mung bean V-ATPase complex
The V-ATPase was purified from mung bean hypocotylsas described previously (Matsuura-Endo et al. 1990;Kawamura et al. 2000) with minor modifications.Tonoplast vesicles were prepared from tissue homo-genates by sucrose density ultracentrifugation (Matsu-ura-Endo et al. 1990). Vacuolar membranes (3 mg of
protein ml�1) were treated with 5% (w/v) Triton X-100plus 0.1 M KCl, and then V-ATPase was solubilizedfrom the membrane with 2 mg ml�1 lys-ophosphatidylcholine for 10 min at 25�C. After centri-fugation for 30 min at 150,000 g, the supernatant wasimmediately subjected to ion-exchange chromatographyusing a column of Source Q15 (Amersham Biosciences)pre-equilibrated with a running buffer containing20 mM Tris–HCl (pH 7.5), 1 mM dithiothreitol, 1 mMEGTA, 2 mM MgCl2, 20% (w/v) glycerol and 0.05%dodecyl maltoside (buffer A), and eluted with a lineargradient of 0–1 M NaCl in buffer A and fractionatedinto 0.3-ml aliquots. The major peak fractions of AT-Pase activity were pooled and further purified by size-exclusion chromatography on a Superose 6 column(Amersham Biosciences), which was equilibrated withthe elution buffer (buffer A). Fractions of 0.3 ml werecollected. All steps were carried out at 4�C.
Partial purification of mung bean Vo domain
The vacuolar membrane (1 mg protein ml�1) obtainedabove was treated with 0.4 M KI plus 5 mM ATP for1 h at 4�C to strip the peripheral V1 domain and sub-sequently centrifuged for 30 min at 150,000 g (Zhanget al. 1992). The KI/ATP-stripped membranes were thensolubilized with 2 mg ml�1 lysophosphatidylcholine for10 min at 25�C. After centrifugation for 30 min at150,000 g, the supernatant was immediately subjected tosize-exclusion chromatography as described above.Fractions of 0.3 ml were collected.
Purification of mung bean V1 subcomplex
The V1 domain was dissociated from the vacuolarmembrane (1.5 mg of protein ml�1) obtained above bytreatment with 0.3 M KI plus 5 mM ATP for 1 h at 4�C.The membrane fraction was removed by centrifugationat 150,000 g for 30 min and the supernatant was dia-lyzed against buffer B [20 mM Tris–HCl (pH 7.5),1 mM dithiothreitol, 1 mM EGTA, 2 mM MgCl2, 3%(w/v) glycerol] overnight at 4�C. The dialyzed samplewas then subjected to an anion-exchange column(Source Q15; Amersham Biosciences) that was equili-brated with buffer B and eluted with a linear gradient of0–1 M NaCl in buffer B. Fractions containing subunitsA and B as detected by SDS–PAGE were pooled andfurther purified by size-exclusion chromatography on aSuperose 6 column (Amersham Biosciences) equili-brated with the elution buffer (buffer B containing150 mM NaCl). Fractions of 0.3 ml were collected.
Electron microscopy
Each purified protein sample was applied to freshlyglow-discharged carbon-coated 400-mesh copper grids
949

and stained with 1% uranyl acetate. Images of thenegatively stained V-ATPase complex were recorded ona Philips Tecnai 20 transmission electron microscopeoperating at 120 kV under ‘‘low dose’’ conditions with acalibrated magnification of 50,040·. Images were re-corded on Lekai films at an underfocus of 0.6–1.2 lm.
Image analysis
Electron micrographs were digitized on a Scitex Jazz+drum scanner with a sampling rate of 20 lm corre-sponding to a step size of 0.4 nm at the specimen level.Image analysis was carried out on a Silicon Graphicsworkstation using the EM program package (Hegerl1996). All the images were examined for astigmatism,drift, and defocus by the contrast-transfer function.Filtration can be used to reduce the influence of noise, soband-pass filtering was used to remove high [>the firstzero (Thon ring) of the contrast-transfer function or>1/1.6 nm�1 if the first zero is beyond 1/1.6 nm�1] andsuppress low (<1/16 nm�1) spatial frequencies, andsuitable images were normalized. Individual particleswere selected and extracted as 100·100 pixel images.The whole data set was subjected to rotational andtranslational alignment via cross-correlation, using arandom particle in the data set as the first reference.Subsequently, multivariate statistical analysis (MSA)and classification were applied only to pixels containingthe V1 part, leading to 32 classes. Each class was sepa-rately subjected to rotational and translational align-ment, using the class average as the reference, andsubsequently MSA/classification was carried out on thewhole complex. After 5–6 runs, using the average of theprevious run as the reference, the final two-dimensional(2D) projections were obtained.
For the V1 subcomplex, the bandpass-filtering wasdone between a low-frequency cut-off of 1/10 nm�1 anda high-frequency cut-off of 1/2.0 nm�1. A total of 13,088particles were selected from 17 micrographs as40·40 pixels and MSA/classification was run as above.
Protein determination, SDS–PAGE, and activity assay
Protein concentration was determined by the method ofLowry et al. (1951) using bovine serum albumin as astandard.
Sodium dodecyl sulfate–polyacrylamide gel electro-phoresis (SDS–PAGE) was carried out by the method ofLaemmli (1970) in the presence of 0.1% SDS.
The purified ATPases were added to a reaction mix-ture containing 3 mM ATP, 3 mM MgCl2, 50 mM KCl,1 mM sodium molybdate, 5 lg of L-a-phosphatidyl-choline (soybean, Type II-S), and 30 mM Tris–Mes(pH 7.0) in the absence or presence of 50 mM KNO3.The total reaction volume was 100 ll. After 20 min at37�C, the reaction was stopped by adding 0.2 ml of 1%SDS. The release of Pi from ATP was determined
essentially as described by Ames (1966) and the absor-bance measured at 820 nm.
Results and discussion
Subunit composition of the V-ATPase from mung bean
The V-ATPase from mung bean hypocotyls was purifiedclose to homogeneity. This enzyme consists of 12 su-bunits with apparent molecular masses determined bySDS–PAGE of 100, 68, 57, 51, 44, 40, 38, 37, 32, 16, 13and 12 kDa (Fig. 1). Our purified V-ATPase exhibitedsimilar polypeptide profiles to those reported in theprevious study by Kawamura et al. (2000), but the bandat 40 kDa was not previously observed. To rule out apossible contamination with the 40-kDa protein duringthe course of purification, the band corresponding to theactive protein complex cut from a native gel electro-phoresis of the purified V-ATPase was subjected toSDS–PAGE, and revealed the same polypeptides (datanot shown). This suggests that the band at 40 kDa isindeed a subunit of the V-ATPase from mung bean.Although some of the subunits of this enzyme have beenidentified, i.e. the 68-, 57-, 38-, 37-, 32-, and 16-kDabands correspond to A, B, E1, E2, D and c subunits,respectively (where E1 and E2 are E subunit isoforms),all other bands, i.e. 100, 51, 44, 40, 13 and 12 kDa, stillremain to be characterized (Matsuura-Endo et al. 1990;Kawamura et al. 2000). The chaotropic agent, KI, wasused to remove the V1 domain from the tonoplastmembrane of mung bean, and the Vo domain was par-tially purified. The subunit composition of the partially
Fig. 1 Subunit composition of V-ATPases in mung bean (Vignaradiata). V-ATPase (8 lg) purified from mung bean hypocotyls(lane 1) and its partially purified Vo domain (4 lg, lane 2) weresubjected to SDS–13.5% PAGE followed by silver staining. Thebands at 68, 57, 40, 32 and 16 kDa correspond to subunits A, B, d,D and c, respectively. The bands at 38 and 37 kDa representE-subunit isoforms. The molecular mass marker is shown on theright
950

purified Vo domain was compared with that of the intactV-ATPase (Fig. 1). As shown in Fig. 1, the partiallypurified Vo domain had no apparent bands of V1 su-bunits, i.e. A, B, H, and E, indicating that the V1 domainwas completely stripped from the tonoplast membraneby KI and the bands in Fig. 1 should be the membrane-associated polypeptides. In spite of the presence ofcontaminations in the partially purified Vo domain, the40-kDa polypeptide co-purified with subunits a(100 kDa) and c (16 kDa) from the gel-filtration chro-matography. This result suggests that the 40-kDa poly-peptide is a membrane-associated protein. It is knownthat the Vo domain mainly consists of three subunits, a,c and d. The excised 40-kDa band was also processed forHPLC/MS/MS peptide mapping. A peptide sequenceobtained, YPPYQAIFSK, shows identity with thed subunit of Arabidopsis thaliana. This then stronglysuggests that the band at 40 kDa represents the d sub-unit of the V-ATPase from mung bean.
Electron-microscopy analysis
The NO3�-sensitive ATPase activity of the protein
sample used for electron-microscopy analysis was1.4 lmol Pi (mg protein)�1 min�1. Figure 2a shows atypical electron micrograph of the negatively stained V-ATPase from mung bean hypocotyls. Protein complexesare well dispersed, a prerequisite for single-particle im-age analysis. Most of the particles were oriented on thecarbon film in the ‘‘side view’’ position, which has aprojection perpendicular to the long axis of the complex.For image processing, only particle images of thedumbbell-like shape were chosen to ensure that all theselected complexes consisted of V1 and Vo.
A total of 6,056 single-particle images were extractedfrom 56 electron micrographs, and subsequently sub-jected to rotational and translational alignment using arandom particle in the data set as the first reference.Misaligned and low-contrast particle images were dis-carded; hence the data set was reduced to include 3,412images. The average dimensions of the molecule calcu-lated from the aligned data set are shown in Fig. 2b. Thewhole molecule is composed of three major domains asfollows: the V1 domain (11.6 nm·13.7 nm, top portion),the stalk domain (6 nm·4.8 nm, middle portion), andthe Vo domain (7.4 nm·11.9 nm, bottom portion). Theoverall dimensions of the complex are 25 nm·13.7 nm.The V1 domain located at the upper part of the complexis divided into two halves by a vertical cleft or cavityrunning from the ‘‘top’’ to the point where the centralstalk is attached. Several smaller protein densities can beseen attached at the top of the V1 domain.
As shown in Fig. 2b, an area of low protein density ofheight 2.4 nm has appeared; the high-density area,including two symmetrical knobs in the V1 domainshould be mainly composed of the A3B3 hetero hexamer(Chaban et al. 2002). The Vo domain is bean-shapedwith its hole plugged by the central stalk on the cytosolic
side. Besides the central stalk connecting the V1 and Vo
domains, a smaller area of high protein density pro-truding just above the Vo domain but attached to thecentral stalk is clearly visible, suggesting the existence ofperipheral stalks.
Since the average in Fig. 2b calculated from thealigned data set contained particles with different ori-entations along the long axis of the complex, we sub-jected these 3,412 single particles to objective alignmentprocedures followed by multivariate statistical analysisand classification to extract all the different projectionsof the V-ATPase.
The V1 subcomplex Giving consideration to the moreinformative V1 domain, we applied the first MSA/clas-sification procedure to images including only V1. Fig-ure 3a–c shows three predominant views of the V1
domain. The preferred orientations of the complex resultin the so-called ‘‘bi-lobed’’ and ‘‘tri-lobed’’ views. In thebi-lobed view (Fig. 3a,b), each of the three large su-bunits overlap in projection leading to the appearance ofa central cleft. In the ‘‘bi-lobed’’ view, the knob-likeextension appears either on the left or the right side of
Fig. 2a,b Electron microscopy and average dimensions of thepurified V-ATPase from mung bean. a Electron micrograph of thepurified V-ATPase from mung bean. The specimen was negativelystained with uranyl acetate (1% w/v). Bar = 100 nm. b Averagedimensions calculated from a total of 3,412 molecules of thepurified V-ATPase from mung bean
951
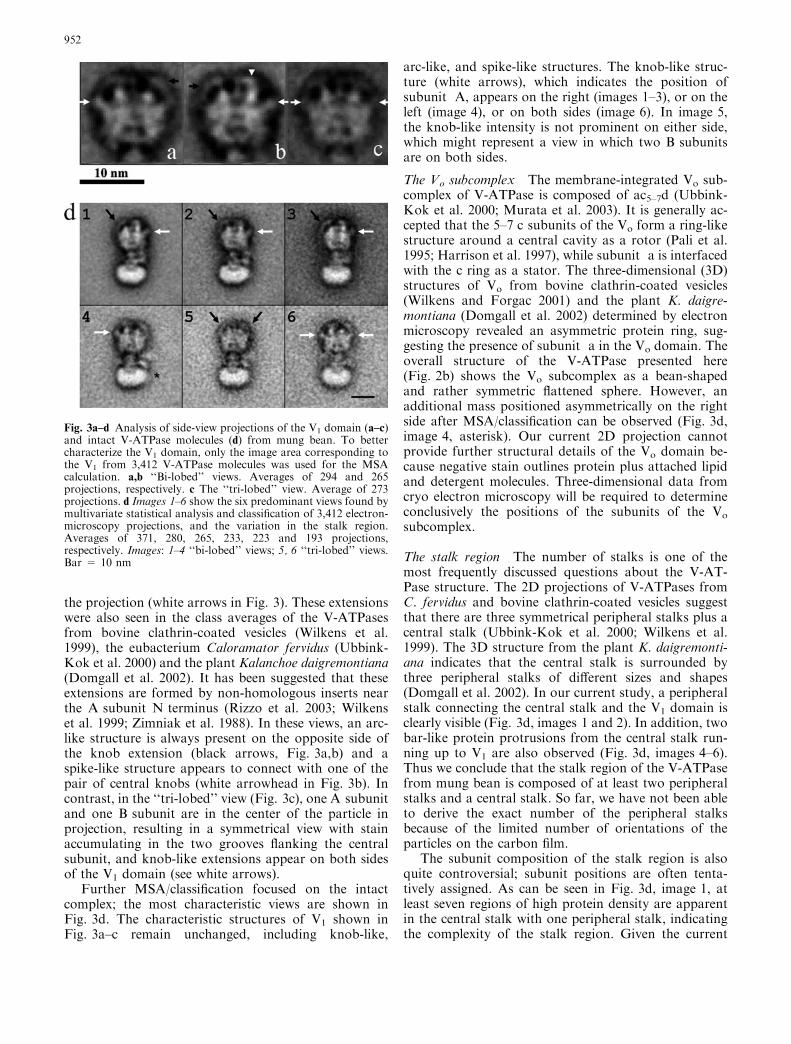
the projection (white arrows in Fig. 3). These extensionswere also seen in the class averages of the V-ATPasesfrom bovine clathrin-coated vesicles (Wilkens et al.1999), the eubacterium Caloramator fervidus (Ubbink-Kok et al. 2000) and the plant Kalanchoe daigremontiana(Domgall et al. 2002). It has been suggested that theseextensions are formed by non-homologous inserts nearthe A subunit N terminus (Rizzo et al. 2003; Wilkenset al. 1999; Zimniak et al. 1988). In these views, an arc-like structure is always present on the opposite side ofthe knob extension (black arrows, Fig. 3a,b) and aspike-like structure appears to connect with one of thepair of central knobs (white arrowhead in Fig. 3b). Incontrast, in the ‘‘tri-lobed’’ view (Fig. 3c), one A subunitand one B subunit are in the center of the particle inprojection, resulting in a symmetrical view with stainaccumulating in the two grooves flanking the centralsubunit, and knob-like extensions appear on both sidesof the V1 domain (see white arrows).
Further MSA/classification focused on the intactcomplex; the most characteristic views are shown inFig. 3d. The characteristic structures of V1 shown inFig. 3a–c remain unchanged, including knob-like,
arc-like, and spike-like structures. The knob-like struc-ture (white arrows), which indicates the position ofsubunit A, appears on the right (images 1–3), or on theleft (image 4), or on both sides (image 6). In image 5,the knob-like intensity is not prominent on either side,which might represent a view in which two B subunitsare on both sides.
The Vo subcomplex The membrane-integrated Vo sub-complex of V-ATPase is composed of ac5–7d (Ubbink-Kok et al. 2000; Murata et al. 2003). It is generally ac-cepted that the 5–7 c subunits of the Vo form a ring-likestructure around a central cavity as a rotor (Pali et al.1995; Harrison et al. 1997), while subunit a is interfacedwith the c ring as a stator. The three-dimensional (3D)structures of Vo from bovine clathrin-coated vesicles(Wilkens and Forgac 2001) and the plant K. daigre-montiana (Domgall et al. 2002) determined by electronmicroscopy revealed an asymmetric protein ring, sug-gesting the presence of subunit a in the Vo domain. Theoverall structure of the V-ATPase presented here(Fig. 2b) shows the Vo subcomplex as a bean-shapedand rather symmetric flattened sphere. However, anadditional mass positioned asymmetrically on the rightside after MSA/classification can be observed (Fig. 3d,image 4, asterisk). Our current 2D projection cannotprovide further structural details of the Vo domain be-cause negative stain outlines protein plus attached lipidand detergent molecules. Three-dimensional data fromcryo electron microscopy will be required to determineconclusively the positions of the subunits of the Vo
subcomplex.
The stalk region The number of stalks is one of themost frequently discussed questions about the V-AT-Pase structure. The 2D projections of V-ATPases fromC. fervidus and bovine clathrin-coated vesicles suggestthat there are three symmetrical peripheral stalks plus acentral stalk (Ubbink-Kok et al. 2000; Wilkens et al.1999). The 3D structure from the plant K. daigremonti-ana indicates that the central stalk is surrounded bythree peripheral stalks of different sizes and shapes(Domgall et al. 2002). In our current study, a peripheralstalk connecting the central stalk and the V1 domain isclearly visible (Fig. 3d, images 1 and 2). In addition, twobar-like protein protrusions from the central stalk run-ning up to V1 are also observed (Fig. 3d, images 4–6).Thus we conclude that the stalk region of the V-ATPasefrom mung bean is composed of at least two peripheralstalks and a central stalk. So far, we have not been ableto derive the exact number of the peripheral stalksbecause of the limited number of orientations of theparticles on the carbon film.
The subunit composition of the stalk region is alsoquite controversial; subunit positions are often tenta-tively assigned. As can be seen in Fig. 3d, image 1, atleast seven regions of high protein density are apparentin the central stalk with one peripheral stalk, indicatingthe complexity of the stalk region. Given the current
Fig. 3a–d Analysis of side-view projections of the V1 domain (a–c)and intact V-ATPase molecules (d) from mung bean. To bettercharacterize the V1 domain, only the image area corresponding tothe V1 from 3,412 V-ATPase molecules was used for the MSAcalculation. a,b ‘‘Bi-lobed’’ views. Averages of 294 and 265projections, respectively. c The ‘‘tri-lobed’’ view. Average of 273projections. d Images 1–6 show the six predominant views found bymultivariate statistical analysis and classification of 3,412 electron-microscopy projections, and the variation in the stalk region.Averages of 371, 280, 265, 233, 223 and 193 projections,respectively. Images: 1–4 ‘‘bi-lobed’’ views; 5, 6 ‘‘tri-lobed’’ views.Bar = 10 nm
952

level of resolution in the 2D projections, we are not ableto place the subunits into the stalk region. This could betackled by imaging V-ATPase complexes tagged withmonoclonal antibodies against individual subunits,work which is ongoing in our lab.
Recently, V1 subcomplexes that differed from eachother in the composition of subunits C and E wereisolated (Chaban et al. 2002). Difference mapping of thesubcomplex projections analyzed by electron micros-copy reveals that subunits E and C are located at thecentral stalk and subunit E is connected to the Vo do-main via subunit C (Chaban et al. 2002). Similarly, wehave purified a subcomplex by KI-driven dissociationfrom the tonoplast membrane eluting at about 15 ml ingel-filtration chromatography. SDS–PAGE revealedthat it mainly contains A and B subunits and the 38-kDaE subunit (Fig. 4a). Densitometric analysis indicatesthat these three subunits account for 90% of the inten-sity in the whole lane. Comparison with the intactcomplex indicates that the contaminants are apparentlynot from the V-ATPase (lane 1). Electron micrographsof the negatively stained subcomplex showed a ratherhomogeneous particle distribution. A total of 13,000projections were extracted and classified after MSA.Figure 4b shows the most characteristic averaged ima-ges. A pair of central knobs is visible in the subcomplexas in the intact enzyme, indicating the ‘‘side view’’ ori-entation (see arrowheads). The knob at the periphery ofV1 corresponding to the position of the A subunit is onthe left (image 2), on the right (image 3), or on both
sides (images 1, 4; see arrows). Moreover, an area ofhigh protein density can be found in all the images ofFig. 4b at the bottom of the subcomplex below thecavity formed by the A and B subunits (see asterisks).This high-density area has a diameter of 2.4 nm, and isnot obvious in the intact complex as it is enclosed in theinterior of the holoenzyme. Since the purified subcom-plex principally consists of three subunits, A, B and E,this area of dense protein most likely represents subunitE. This result is consistent with the observation thatsubunit E is located at the upper part of the centralstalk in the V-ATPase from C. fervidus (Chaban et al.2002).
In summary, Fig. 5 represents a hypothetical modelof mung bean V-ATPase based on available biochemicaldata and the images presented here, as well as previousstructural information. The features reported here forthe V-ATPase from mung bean are remarkably similarto those deduced from projection structures of bovineclathrin-coated vesicles (Wilkens et al. 1999) and theeubacterium C. fervidus (Ubbink-Kok et al. 2000), i.e.the V1 headpiece, including knob-like structures and thecentral cleft; and the asymmetric bean-shaped Vo do-main and stalk region containing at least two peripheralstalks plus a central stalk. Our preliminary results sug-gest that the E subunit is located at the central stalk. Wealso detected a 40-kDa membrane-associated polypep-
Fig. 4a,b Subunit composition of the subcomplex as determined byKI-driven dissociation (a) and averaging of single-particle projec-tions (b). a The subcomplex resulting from KI-driven dissociation(8 lg) was subjected to SDS–15% PAGE followed by Coomassieblue staining (lane 2) and compared with the purified holoenzyme(lane 1). b Images 1–5 show the five predominant views found bymultivariate statistical analysis and classification of 13,000 elec-tron-microscopy projections, and are the averages of 415, 379, 378,442 and 299 projections, respectively. Bar = 5 nm
Fig. 5 Model of the V-ATPase from mung bean based on theknown structural data and the present study. Overlaid subunitorganizations, imposed on the side-view projection of the V-ATPase from Fig. 3d, image 1, mark the position of the A3B3
headpiece in V1, the E subunit and the membrane-bound Vo partincluding subunits a, c, d. The remaining subunits cannot bepositioned using the data currently available. Bar = 10 nm
953

tide that is found in the Vo domain together withsubunits a and c. Peptide analysis indicates that the 40-kDa polypeptide represents the d subunit. In order tounambiguously define the subunit architecture of the V-ATPase, cryo electron microscopy, which is ongoing inour laboratory, will be required in order to avoid pos-sible artifacts caused by negatively staining.
Acknowledgments We are very grateful to Prof. Fuyu Yang forcontinuous support and crucial advice on experiments. We thankG. Pfeifer and R. Hegerl for help with the EM program. This workwas supported by grants from the Chinese Academy of Sciences(KSCX2-2-05) and the Third World Academy of Sciences (00-238RG/Bio/AS) to Xujia Zhang. We thank Sarah Perrett and JudySmith for their help in improving the manuscript.
References
Abrahams JP, Leslie AG, Lutter R, Walker JE (1994) Structure at2.8 A resolution of F1-ATPase from bovine heart mitochon-dria. Nature 370:621–628
Ames BN (1966) Assay of inorganic phosphate. Total phosphateand phosphatases. Methods Enzymol 8:115–118
Arata Y, Baleja JD, Forgac M (2002) Localization of subunits D,E, and G in the yeast V-ATPase complex using cysteine-medi-ated cross-linking to subunit B. Biochemistry 41:11301–11307
Boekema EJ, Ubbink-Kok T, Lolkema JS, Brisson A, KoningsWN (1997) Visualization of a peripheral stalk in V-type AT-Pase: evidence for the stator structure essential to rotationalcatalysis. Proc Natl Acad Sci USA 94:14291–14293
Boekema EJ, van Breemen JF, Brisson A, Ubbink-Kok T, KoningsWN, Lolkema JS (1999) Connecting stalks in V-type ATPase.Nature 401:37–38
Boyer PD (1999) Molecular motors: what makes ATP synthasespin? Nature 402:247–249
Chaban Y, Ubbink-Kok T, Keegstra W, Lolkema JS, Boekema EJ(2002) Composition of the central stalk of the Na+-pumping V-ATPase from Caloramator fervidus. EMBO Rep 3:982–987
Domgall I, Venzke D, Luttge U, Ratajczak R, Bottcher B (2002)Three-dimensional map of a plant V-ATPase based on electronmicroscopy. J Biol Chem 277:13115–13121
Forgac M (1999) Structure and properties of the vacuolar (H+)-ATPases. J Biol Chem 274:12951–12954
Finbow MF, Harrison MA (1997) The vacuolar H+-ATPase: auniversal proton pump of eukaryotes. Biochem J 324:697–712
Graham LA, Powell B, Stevens TH (2000) Composition andassembly of the yeast vacuolar H(+)-ATPase complex. J ExpBiol 203:61–70
Gruber G, Radermacher M, Ruiz T, Godovac-Zimmermann J,Canas B, Kleine-Kohlbrecher D, Huss M, Harvey WR, Wie-czorek H (2000) Three-dimensional structure and subunittopology of the V1 ATPase from Manduca sexta midgut. Bio-chemistry 39:8609–8616
Gruber G, Wieczorek H, Harvey WR, Muller V (2001) Structure–function relationships of A-, F- and V-ATPases. J Exp Biol204:2597–2605
Harrison MA, Finbow ME, Findlay JBC (1997) Postulate for themechanism of the vacuolar H+-ATPase (hypothesis). MolMembr Biol 14:1–3
Hegerl R (1996) The EM program package: a platform for imageprocessing in biological electron microscopy. J Struct Biol116:30–34
Kawamura Y, Arakawa K, Maeshima M, Yoshida S (2000)Tissue specificity of E subunit isoforms of plant vacuolar
H+-ATPase and existence of isotype enzymes. J Biol Chem275:6515–6522
Laemmli UK (1970) Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227:680–685
Lowry OH, Rosebrough NJ, Lewis Farr A, Randall RJ (1951)Protein measurement with folin phenol reagent. J Biol Chem193:265–275
Marin BP (1985) Biochemistry and function of vacuolar adenosinetriphosphatase in fungi and plants. Springer, Berlin HeidelbergNew York
Matsuura-Endo C, Maeshima M, Yoshida S (1990) Subunit com-position of vacuolar membrane H+-ATPase from mung bean.Eur J Biochem 187:745–751
Murata T, Arechaga I, Fearnley IM, Kakinuma Y, Yamato I,Walker JE (2003) The membrane domain of the Na+-motive V-ATPase from Enterococcus hirae contains a heptameric rotor. JBiol Chem 278:21162–21167
Nishi T, Forgac M (2002) The vacuolar (H+)-ATPases—nature’smost versatile proton pumps. Nat Rev Mol Cell Biol 3:94–103
Pali T, Finbow ME, Holzenburg A, Findlay JB, Marsh D (1995)Lipid–protein interactions and assembly of the 16-kDa channelpolypeptide from Nephrops norvegicus. Studies with spin-labelelectron spin resonance spectroscopy and electron microscopy.Biochemistry 34:9211–9218
Radermacher M, Ruiz T, Harvey WR, Wieczorek H, Gruber G(1999) Molecular architecture of Manduca sexta midgut V1
ATPase visualized by electron microscopy. FEBS Lett 453:383–386
Radermacher M, Ruiz T, Wieczorek H, Gruber G (2001) Thestructure of the V1-ATPase determined by three-dimensionalelectron microscopy of single particles. J Struct Biol 135:26–37
Ratajczak R (2000) Structure, function and regulation of the plantvacuolar H+-translocating ATPase. Biochim Biophys Acta1465:17–36
Rizzo VF, Coskun U, Radermacher M, Ruiz T, Armbruster A,Gruber G (2003) Resolution of the V1 ATPase from Manducasexta into subcomplexes and visualization of an ATPase-activeA3B3EG complex by electron microscopy. J Biol Chem278:270–275
Stock D, Leslie AG, Walker JE (1999) Molecular architecture ofthe rotary motor in ATP synthase. Science 286:1700–1705
Sze H (1985) H+-translocationg ATPase: advances using mem-brane vesicles. Annu Rev Plant Physiol 36:175–208
Sze H, Ward MW, Lai S (1992) Vacuolar H+-translocatingATPases from plants: structure, function, and isoforms. JBioenerg Biomembr 24:371–382
Sze H, Li X, Palmgren MG (1999) Energization of plant cellmembranes by H+-Pumping ATPases: regulation and biosyn-thesis. Plant Cell 11:677–689
Ubbink-Kok T, Boekema EJ, van Breemen JFL, Brisson A,Konings WN, Lolkema JS (2000) Stator structure and subunitcomposition of the V1/V0 Na+-ATPase of the thermophilicbacterium Caloramator fervidus. J Mol Biol 296:311–321
Wilkens S, Forgac M (2001) Three-dimensional structure of thevacuolar ATPase proton channel by electron microscopy. J BiolChem 276:44064–44068
Wilkens S, Vasilyeva E, Forgac M (1999) Structure of the vacuolarATPase by electron microscopy. J Biol Chem 274:31804–31810
Zhang J, Myers M, Forgac M (1992) Characterization of the Vo
domain of the coated vesicle (H+)-ATPase. J Biol Chem267:9773–9778
Zimniak L, Dittrich P, Gogarten JP, Kibak H, Taiz L (1988) ThecDNA sequence of the 69-kDa subunit of the carrot vacuolarH+-ATPase. Homology to the beta-chain of F0F1-ATPases. JBiol Chem 263:9102–9112
954

![V-ATPase · From Wiki: Vacuolar-type H+ -ATPase (V-ATPase) is a highly conserved evolutionarily ancient enzyme with remarkably diverse functions in eukaryotic organisms.[1] membranes](https://img.pdfslide.us/doc/110x75/5fa3fb056ad5ca477269e2ce/v-atpase-from-wiki-vacuolar-type-h-atpase-v-atpase-is-a-highly-conserved-evolutionarily.jpg)















![Mung Production [Compatibility Mode]](https://img.pdfslide.us/doc/110x75/549df729b4795974208b45dc/mung-production-compatibility-mode.jpg)