Embed Size (px)
Citation preview

Cell Tiss. Res. 177, 331-345 (1977) Cell and Tissue Research �9 by Springer-Verlag 1977
Electron Microprobe Investigations into the Process of Hard Tissue Formation* W.A.P. Nicholson, B.A. Ashton**, H.J. H6hling, P. Quint, J. Schreiber Institut fOr Medizinische Physik, Universit/it MOnster, Federal Republic of Germany
I.K. Ashton** Anatomisches Institut, UniversitM MOnster, Federal Republic of Germany
A. Boyde Department of Anatomy, University College London, U.K.
Summary. The electron microprobe microanalyser has been used to measure the concentrations of Ca, P and S in the predentine of young rat incisors. The specimens were prepared as alcohol fixed embedded ultrathin sections, unfixed vacuum embedded dry cut ultrathin sections and as thin cryostat sections. The results show the influence of preparation on the measured compositions and indicate that Ca is tightly bound to the matrix, whereas P can be easily washed out. Measurements along the dentine-predentine border demonstrated zones of Ca enrichment, the average size of which suggests that the zones could be the prestages of calcospherites. A mineralisa- tion mechanism is discussed in which the high Ca concentration activates pyrophosphatase or ATPase before the onset of nucleation.
Key words: Mineralisation - Rat predentine - Microprobe analysis.
Introduction
Many processes involved in hard tissue formation are still not well understood. Using the electron probe microanalyser we have sought to contribute to the knowledge of how calcium and phosphate ions become enriched in the extra- cellular organic matrix of hard tissue. After some measurements in the osteoid of the tibial and femoral diaphyses of chick embryos (H6hling et al., 1970), we concentrated primarily on rat incisors. This organ has the advantages that the predentine is relatively wide (about 20 gm) so that the odontoblasts are well separated from the dentine. As well, there is a continuous process of minerali- sation, so that mineral remodeling does not have to be considered. Nevertheless,
Send offprint requests to : Dr. W.A.P. Nicholson, Dept. of Natural Philosophy, University of Glasgow, Glasgow, U.K. Prof. Dr. Hans J. H6hling, Institut for Med. Physik, HOfferstr. 68, D-4400 MOnster, Federal Republic of Germany
* The authors express their thanks to the Deutsche Forschungsgemeinschaft for financial support ** Present Address: Bone Research Unit, Nuffield Orthopaedic Centre, Oxford, England

332 W.A.P. Nicholson et al.
it seems likely that most of the stages present in dentine formation will also occur in the mineralisation of foetal bone.
It is now generally accepted that the first mineral formation in foetal bone and dentine begins in collagen free vesicles (dentine: Croissant, 1971; Ozawa and Yajima, 1972; Slavkin et al., 1972; Sisca and Provenza, 1972; Eisenmann and Glick, 1972; Bernard, 1972). In the case of dentine mineralisation, after the deposition of the first layers, the mantle dentine, the vesicles no longer play a role and mineralisation takes place directly in the matrix at the dentine-predentine border. This is also true in the later stages of development when the odonto- blastic processes have developed and can be used by the cells as transport paths for ions and proteins. Our measurements have been made in predentine mainly in these later stages of mineralisation, the results of which have been previously reported in part in the form of short notes (H6hling et al., 1967, 1968, 1972; Ashton et al., 1973; Nicholson et al., 1974).
Materials and Methods
The continuously growing ends of young incisors from rats were fixed in alcohol to minimise the loss of mineral and then directly embedded in methyl methacrylate. Ultrathin sections were then removed from the surface of the water in the microtome trough and mounted on standard electron microscopic copper grids. Most of the sections were coated on both sides with a 5 nm film of carbon and then analysed in an electron microscopical microanalyser, the prototype of the AEI EMMA IV, for Ca, P and S using ultrathin sections of methacrylate embedded apatite and CaSO 4 as standards.
In the next series of experiments the same type of tissue was quick-frozen in Freon 12 at - 155~ and then freeze-dried under vacuum at - 8 0 ~ in an Edwards-Pearse tissue dryer. The dried tissue was then vacuum-infiltrated in araldite or methacrylate, which were polymerised at 60 ~ C and 50 ~ C, respectively, after which the specimens were sectioned dry at a thickness of 100 to 200 rim. The sections were transferred from the microtome knife using either a glass fibre or an eye lash. They were then flattened (under a microscope) using a small glass ball and by placing the holders on the edge of a damp region of a filter paper. The sections were placed upper-most to prevent direct contact with the water. They were then coated with a 5 nm layer of either carbon or aluminium. The holders used for these measurements were 3 mm discs of either copper or aluminium with a I mm central hole covered with a pioloform foil which was coated with aluminium or carbon of 5 nm thickness. This type of holder was chosen in preference to standard EM grids in order to minimise the intensity of stray x-rays which we have shown to be very high within 200 p.m of any metal specimen support such as grid bars (Nicholson, 1974). We used the quantitative analytical method of Hall (1971, 1973) in which the specimen mass is monitored by the intensity of part of the continuous radiation: it is thus important that this should originate from the specimen alone and not be stray radiation.
These analyses were made in the Cameca microprobe fitted with the electron microscope attach- ment at about 70 nA. Since this EM attachment has only one useful magnification of 5000 times and the transparency of 100 to 200 nm sections is poor with 40kV electrons, it was necessary to have previously recorded micrographs of the predentine regions to be analysed. These were produced using a Siemens Elmiskop 1 a electron microscope. The measured points could then be recorded directly on montages of these micrographs (e.g. Figs. 2 and 3).
The most recent series of measurements have been carried out on cryostat sections of rat incisor predentine. These have been prepared by rapidly freezing the tissue at liquid nitrogen temperature and then by cutting 4 to 6 gm sections at - 2 5 ~ in a freezing microtome. The sections were placed on the holders and left to dry in the cryostat before bringing to room temperature in a dessicator. After warming up, the sections were coated with 20 to 40 nm of aluminium. The holders for these experiments were 10 mm diameter aluminium tubes with a specimen supporting foil of 100 to 200 nm nylon stretched over the end of the tube and then coated with 40 nm of aluminium.
The Ca, P and S determinations were made in the standard Cameca microprobe model MS46, using either the specimen current, secondary electron or transmission scanning images to localise

Electron Microprobe Investigations of Hard Tissue Formation 333
the border regions to be measured. Because of the moderate resolution of the microprobe (about 0.8 gin) and the small contrast difference between the predentine and the odontoblast regions of the specimen, it was not possible to distinguish between these regions from the display images. For this reason we have restricted our measurements to a distance of about I 0 gm from the dentine-predentine border. As the predentine is about 20 gm wide, we could therefore be sure that all our measurements were made in the predentine and not in the cellular regions.
The standards used for the microprobe measurements of Ca and P were ultrathin sections of apatite powder embedded in methyl methacrylate. Similar sections of FeS 2 and K 2 S O 4 were used as standards for S and later standards of sulphur in araldite of the type described by Jessen et al. (1974) were used. These two sorts of standards led to differing results, but it has not yet been possible to resolve the problem of which set is more accurate.
Bovine predentine has been prepared from frozen calf molars. The state of the phosphorus in this predentine was elucidated from the determination of total phosphate and inorganic orthophosphate by the molybdate method (Fiske and Subba Row, 1954) following hydrolysis and oxidation, and the enzymic method described by Lowry and Passoneau (1972), respectively. The difference between total and orthophosphate represents the amount of phosphate in polyphosphatc, organophosphate esters and phosphoproteins.
Results and Discussion
Discussion of Specimen Preparation for Microanalysis
T h e c o n v e n t i o n a l p r e p a r a t i o n used fo r m o r p h o l o g i c a l e x a m i n a t i o n in t he e l e c t r o n
m i c r o s c o p e o f h a r d t issue s p e c i m e n s such as ra t p r e m o l a r s i n v o l v e s f ix ing the
t i ssue in g l u t a r a l d e h y d e a n d O s O 4 b e f o r e e m b e d d i n g . F i g u r e 1 is a t yp i ca l m i c r o -
g r a p h o f a sec t ion o f p r e m o l a r f r o m a ra t o f a few days old . By such p r e p a r a t i o n
the o d o n t o b l a s t s , the p r e d e n t i n e wi th the o d o n t o b l a s t i c p rocesses , a n d the
Fig. 1. Electron micrograph of normally stained ultrathin section with odontoblasts, predentine and dentine. Od odontoblast, Pr predentine, OP odontoblastic process, D dentine, x 4,000

334 W.A.P. Nicholson et al.
mineralised dentine can all be very easily recognised. Unfortunately, this type of preparation is unsuitable for the microprobe since foreign elements such as osmium will be introduced into the tissue, and soluble ions (e.g. PO4) will be washed out during the fixing process and sectioning, as the sections are normally taken off the surface of a water bath in the microtome.
An improved method of preparing ultrathin sections by freeze-drying and vacuum-embedding the tissue before dry sectioning eliminates some of the disadvantages of conventional preparation, but it must be accepted that the embedding medium may displace diffusible ions to a certain extent, and the flattening of the sections with water vapour may remove some (variable) fraction of diffusible substances. Further, because the method of quantitation by Hall (1971) for thin sections uses part of the continuous, "white" radiation to monitor the total mass being analysed, we can only measure weight fractions related to the mass of the specimen plus embedding materials. It is clear that by registering the white radiation it is not possible to distinguish between x-rays coming from the tissue and those coming from the embedding materials. However, it is possible to compensate for this error if an estimate is made of the fraction of embedding material in the measured zone. In spite of the above-mentioned problems with sections of embedded material, there is the important advantage that the structure is still well preserved, so that the measurements of the distributions of Ca, P and S can be well correlated to the morphology of the sections, in particular to the shape of the developing mineral front.
A further improvement in the preparation of specimens for the microprobe has led to the use of 6 ~tm cryostat sections of frozen material that have had no contact with any kind of fluid. The tissue must be brought from the temperature of about -150~ used for freezing up to the cryostat temperature of -25~ used for sectioning. During this warming-up process the danger exists that some elements may be moved from their original sites in the specimen by the growth of ice crystals which were formed during the first freezing of the tissue. It is thought, however, that such movements may not be important when compared to the 1 ~tm diameter of the electron beam, which determines the resolution of the analyses. The principal difficulty with 6 lam thick frozen sections is that until now the only structure recognisable in the microprobe is the dentine- predentine border to which all measurements must be referred. Although the specimens are protected by coating both sides with a layer of aluminium 20 to 40 nm thick, and the current of the electron beam used for the measurements is very low (20 nA), still another problem exists that a certain loss of organic matrix occurs under the electron beam (H6hling et al., 1973; Hall and Gupta, 1974). However, we have achieved close agreement between parallel chemical and microprobe analyses for Ca and P of rat tail tendon (see entries 4 and 5, Table 1).
The next development in the preparation of hard tissues for the microprobe in the future must be the use of ultrathin frozen sections, as has been used by Appleton (1974) for soft tissues. Such sections eliminate the introduction of foreign substances into the sections, avoid contact with water and are also suitable for electron microscopy. Thus, good morphological correlation of the analyses is possible, added to which a much higher spatial resolution (about 0. I ~tm) will be possible for the measurements.
Electron Microprobe Investigations of Hard Tissue Formation
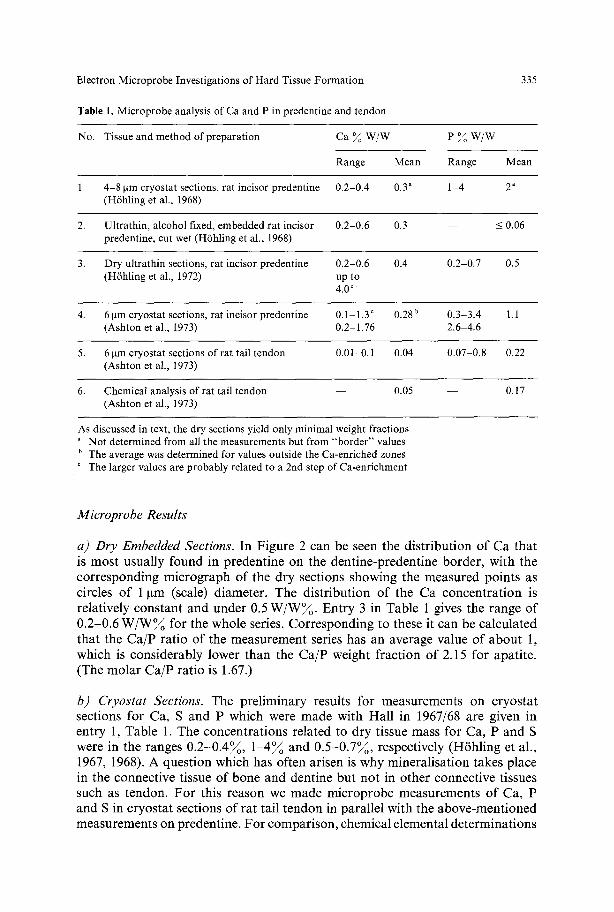
Table 1. Microprobe analysis of Ca and P in predentine and tendon
335
No. Tissue and method of preparation Ca ~ W/W e % w / w
Range Mean Range Mean
1. 4-8 ~tm cryostat sections, rat incisor predentine 0.2-0.4 0.3 a 1-4 2 a (H6hling et al., 1968)
2. Ultrathin, alcohol fixed, embedded rat incisor 0.2-0.6 0.3 - - < 0.06 predentine, cut wet (H6hling et al., 1968)
3. Dry ultrathin sections, rat incisor predentine 0.2-0.6 0.4 0.2-0.7 0.5 (Hfhling et al., 1972) up to
4.0 ~
4. 6 gm cryostat sections, rat incisor predentine 0.1-1.3 ~ 0.28 u 0.3-3.4 1.1 (Ashton et al., 1973) 0.2 1.76 2.6-4.6
5. 6 gm cryostat sections of rat tail tendon 0.01-0.1 0 . 0 4 0.07-0.8 0.22 (Ashton et al., 1973)
6. Chemical analysis of rat tail tendon - - 0.05 - - 0.17 (Ashton et al., 1973)
As discussed in text, the dry sections yield only minimal weight fractions a NOt determined from all the measurements but from "border" values b The average was determined for values outside the Ca-enriched zones
The larger values are probably related to a 2nd step of Ca-enrichment
Microprobe Results
a) Dry Embedded Sections. In F igure 2 can be seen the d i s t r ibu t ion o f Ca tha t is mos t usua l ly found in p reden t ine on the den t ine -p reden t ine borde r , wi th the c o r r e s p o n d i n g m i c r o g r a p h o f the d ry sect ions showing the m e a s u r e d po in t s as circles o f 1 ~tm (scale) d iameter . The d i s t r ibu t ion o f the Ca c onc e n t r a t i on is re la t ive ly cons t an t and under 0.5 W / W ~ . Ent ry 3 in Table 1 gives the range o f 0 .2 -0 .6 W / W % for the whole series. C o r r e s p o n d i n g to these it can be ca lcu la t ed tha t the C a / P ra t io o f the m e a s u r e m e n t series has an average value o f a b o u t 1, which is cons ide rab ly lower than the C a / P weight f rac t ion o f 2.15 for apa t i te . (The m o l a r C a / P ra t io is 1.67.)
b) Cryostat Sections. The p re l im ina ry results for measu remen t s on c ryos t a t sect ions for Ca, S and P which were m a d e wi th Hal l in 1967/68 are given in ent ry 1, Table 1. The concen t ra t ions re la ted to d ry t issue mass for Ca, P and S were in the ranges 0 . 2 - 0 . 4 ~ , 1 - 4 ~ and 0 . 5 - 0 . 7 ~ , respect ively (H6h l ing et al. , 1967, 1968). A ques t ion which has of ten ar isen is why mine ra l i s a t ion takes p lace in the connect ive t issue o f bone and dent ine but no t in o the r connect ive t issues such as tendon . F o r this reason we m a d e m i c r o p r o b e me a su re me n t s o f Ca, P and S in c ryos t a t sect ions o f ra t ta i l t endon in pa ra l l e l wi th the a b o v e - m e n t i o n e d measu remen t s on predent ine . F o r compar i son , chemica l e lementa l d e t e r m i n a t i o n s

336 W.A.P. Nicholson et al.
were also made on samples of similar pieces of tissue. The results for these sets of measurements (Ashton et al., 1973) are given in entries 4, 5 and 6 of Table 1. The microprobe average weight fractions for predentine were 0.28~o for Ca and 1.1 for P, compared with the results for tendon of 0.04~o for Ca and 0.22~ for P. By comparing entries 5 and 6, it can be seen that there is good agreement between chemical and microprobe results for Ca and P. Estimation of S in predentine and tendon by the microprobe method has been beset with problems in the choice of suitable standards. Mean values ranging between 0.1 and 0.4~o W/W for tendon and 0.25 to 1.1 ~ W/W for predentine have been obtained. For the chemical determinations in tendon, the method of Dodgson and Price (1962) has been used to determine the content of the sulphur bound in glycosaminoglycans leading to a value of 0.07~o S (Ashton et al., 1973). The method of Zimmermann (1961) for determination of the whole sulphur content gave a value of 0.37~ S. The two different chemical methods gave values of S in tendon which lie at opposite ends of the range of values determined in the microprobe, but it is not yet possible to say in which part of the range the true values lie.
By comparison between entries 4 and 5 it can be seen that the average content in predentine is higher than that in tendon by factors of 7 for Ca and 5 for P. The results clearly indicate that there is an active accumulation in predentine which does not occur in tendon.
c) Sections which had Contact with Water or Alcohol. The first set of dentine sections were treated with water during the specimen preparation procedure rather than under controlled conditions. In spite of this, these results gave pre- liminary information about the presence of soluble compounds (mainly phosphate groups) in predentine. The intensity of phosphorous radiation was so low that only an approximate value of 0.06~ for the phosphorous concentration could be estimated, this being at the limit of detectability for this element in the micro- probe. Similar results were obtained for sections of freeze-dried tissue embedded in methacrylate that were sectioned 'wet' and floated on the surface of the water bath of the microtome for several minutes.
The phosphorus concentration in these sections was also mostly just at the limit of detectability.
More recently, cryostat sections mounted on the usual microprobe holders were treated with distilled water. Water (25 gl) was placed on the sections for 15 min, after which it was removed using a pipette and replaced with a 25 gl aliquot for a further 15 min. The first results from a number of water treated sections show that even these small volumes of water remove a large amount of phosphorus (50~ and more) from the predentine region. The removal of calcium by water treatment was undetectable in some sections and was about 20~o from others. This data shows the importance of using cryostat sections when accurate concentrations of soluble ions are to be measured.
Measurements of Sulphur across the Dentine/Predentine Border
The results presented here are of measurements made of S related to tissue volume and not related to the dry mass of the section, and are thus independent of measure- ments of S standards and the related problems. It has been widely accepted that

Electron Microprobe Investigations of Hard Tissue Formation 337
a break down of S-rich proteoglycans and related substances takes place associated with the process of hard tissue formation. Pugliarello et al. (1970) performed chemical determinations on osteons at various stages of mineralisation and found a decrease in the content of non-collagenous substances with increasing minerali- sation, whereas the collagen content remained unchanged. Using the electron microprobe Baylink et al. (1972) concluded that as the Ca and P content increased in predentine and dentine, the S content and therefore the concentration of S-rich proteoglycans decreased. It should be borne in mind that the measurements made by Baylink et al. (1972) were performed on cryostat or electron opaque specimens, in which S radiation would be heavily absorbed in the dentine zones. A correction for this absorption would be extremely difficult as the absorption is dependent upon the density of mineralisation.
For our S determinations in dentine, ultrathin sections were used since it was thought that the high absorption of S X-rays in the dentine region of 6 ~tm cryostat sections would preclude accurate measurements. It was assumed that in a large number of measurements (> 100) variations in the section thickness would tend to cancel out. Point measurements were made (a) in predentine close to the border, (b) directly next to these points but in the dentine, thus 1 to 2 ~tm away, and (c) 5 to 10 lam away from the mineral front in the dentine. The results showed that in regions with a higher S content per volume in the predentine, the S content was approximately the same or slightly higher in the adjacent dentine border, but always decreased beyond the border 5 to 10 I~m into the dentine. In regions with lower S content in the predentine, the S content fell at the dentine border and was further decreased (sometimes to an undetectable amount) 5-10 ~tm into the dentine. The results indicate that no large loss of S-rich proteoglycans occurs in the border region of mineralisation, but at a region within the dentine, although only a few microns from the predentine/dentine border. This is in agreement with the findings of Sundstr6m (1971) who showed that only a fraction of the sulphate incorporated into the mineralised dentine.
The positions of measurements made 5 to 10 ~tm from the border could be only roughly estimated, so that the measurements between these and the border were not made. For this reason and because the embedding material might have moved the sulphur-containing substances, these results of H6hling et al. (1972) can be only regarded as giving a rough picture of the distribution of S across the dentine/predentine border. Further systematic measurements are required to determine a more detailed distribution of the S-loss from the predentine into the dentine zone.
Zones Showing the Second Stage of Ca Enrichment
Although the concentrations of Ca and P found in normal areas of predentine were much higher than in blood plasma and interstitial tissue fluid, they are still considerably below (about 1/30) the concentrations of Ca found in dentine, which is only 1 or 2 gm away from the measured points. Since mineralisation begins in the predentine, it was anticipated that much higher concentrations may exist in some micro areas in predentine. By systematic measurements along the

338 W.A.P. Nicholson et al.
Fig. 2a and b. Calcium measurements in predentine of a dry ultrathin section of rat incisor, deep frozen and freeze-dried embedded, a Electron micrograph of the analysed section; circles mark the areas measured, x 4,340. b Graph of a typical Ca-distribution in predentine below 1 ~ w/w; Ca calcium
Fig. 3a and b, Calcium and phosphorus measurements in predentine of a dry ultrathin section of rat incisor, deep frozen and freeze-dried embedded, a Electron micrograph of the analysed section; circles mark the areas measured. M Area in which the mineralisation front lags behind that of neigh- bouring regions. • 4,340. b Graph of calcium and phosphorus distribution in predentine. Ca calcium, P phosphorus. 1 region with a typical relatively low calcium distribution. H region with two micro- compartments of a 2rid stage of calcium enrichment. E calcium enrichraent which corresponds to the region M of Figure 3a, in which the mineralisation front lags behind that of the neighbouring regions
den t ine /predent ine border such regions showing second stages of Ca enr ichment were found (sometimes associated with cor responding P and S enrichments) .
Figure 3 a is a micrograph of a region with two ne ighbour ing compar tmen t s of Ca enr ichment , the measured 1 gm diameter points being shown as circles. F igure 3 b shows the cor responding Ca distr ibut ion. Above and below the enriched region (II) Ca has the concen t ra t ion normal ly found of less than 1 ~ (I). The upper c o m p a r t m e n t has a peak concen t ra t ion of abou t 4 ~ Ca and the lower of abou t 2~o.
Electron Microprobe Investigations of Hard Tissue Formation 339
4b 0.8
E LLJ
0.6
0 . 4 =
0 . 2 �9
O ~ o - - o ~ o ~ Co
--x--x--x-- S
- - o ~ o ~ o ~ P
e
Zo 60 Distance etong the border (#m)
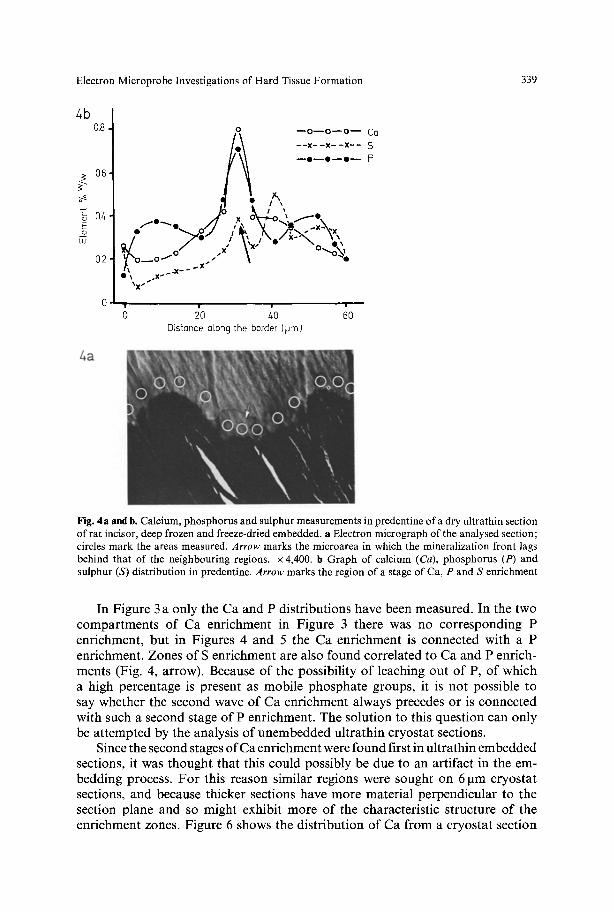
Fig. 4a and b. Calcium, phosphorus and sulphur measurements in predentine of a dry ultrathin section of rat incisor, deep frozen and freeze-dried embedded, a Electron micrograph of the analysed section; circles mark the areas measured. Arrow marks the microarea in which the mineralization front lags behind that of the neighbouring regions, x 4,400. b Graph of calcium (Ca), phosphorus (P) and sulphur (S) distribution in predentine. Arrow marks the region of a stage of Ca, P and S enrichment
In Figure 3 a only the Ca and P distributions have been measured. In the two compartments of Ca enrichment in Figure 3 there was no corresponding P enrichment, but in Figures 4 and 5 the Ca enrichment is connected with a P enrichment. Zones of S enrichment are also found correlated to Ca and P enrich- ments (Fig. 4, arrow). Because of the possibility of leaching out of P, of which a high percentage is present as mobile phosphate groups, it is not possible to say whether the second wave of Ca enrichment always precedes or is connected with such a second stage of P enrichment. The solution to this question can only be attempted by the analysis of unembedded ultrathin cryostat sections.
Since the second stages of Ca enrichment were found first in ultrathin embedded sections, it was thought that this could possibly be due to an artifact in the em- bedding process. For this reason similar regions were sought on 6 pm cryostat sections, and because thicker sections have more material perpendicular to the section plane and so might exhibit more of the characteristic structure of the enrichment zones. Figure 6 shows the distribution of Ca from a cryostat section
5b
0.8.
06. ?
~ 04.
klJ
0.2,
0,
5a
1 --o--o--o-- Ca
/ * \ p --x--x--x-- s / : / \ - ~ 1 7 6 1 7 6 p
/ I,X,\,~ 13 , , / /I',, ", ,.,,. no,, \ I ,
~, \'i,/ ",' ~ .
\3;-.. Q / \X~ tX~
�9 i i i J i
0 20 &0 60 80 100
Distance along the border (~Jm)
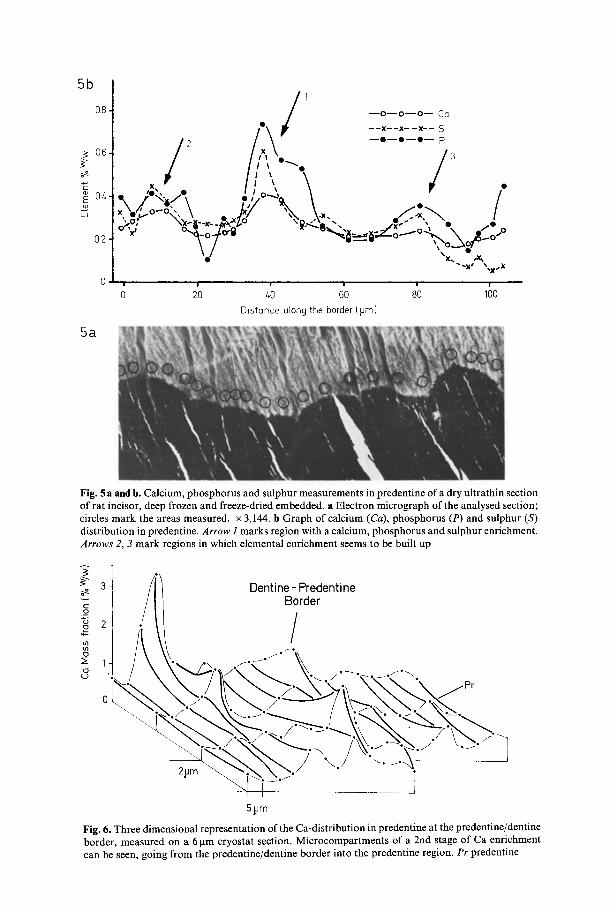
Fig. 5 a and b. Calcium, phosphorus and sulphur measurements in predentine of a dry ultrathin section of rat incisor, deep frozen and freeze-dried embedded, a Electron micrograph of the analysed section; circles mark the areas measured. • 3,144. b Graph of calcium (Ca), phosphorus (P) and sulphur (S) distribution in predentine. Arrow i marks region with a calcium, phosphorus and sulphur enrichment. Arrows 2, 3 mark regions in which elemental enrichment seems to be built up
. . . . _ . ..
t 51Jm
Fig. 6. Three dimensional representation of the Ca-distribution in predentine at the predentine/dentine border, measured on a 6 gm cryostat section. Microcompartments of a 2nd stage of Ca enrichment can be seen, going from the predentine/dentine border into the predentine region. Pr predentine

Electron Microprobe Investigations of Hard Tissue Formation 341
showing the second stage of Ca enrichment along and across the dentine/pre- dentine border.
Morphological Considerations of the Zones of Ca Enrichment
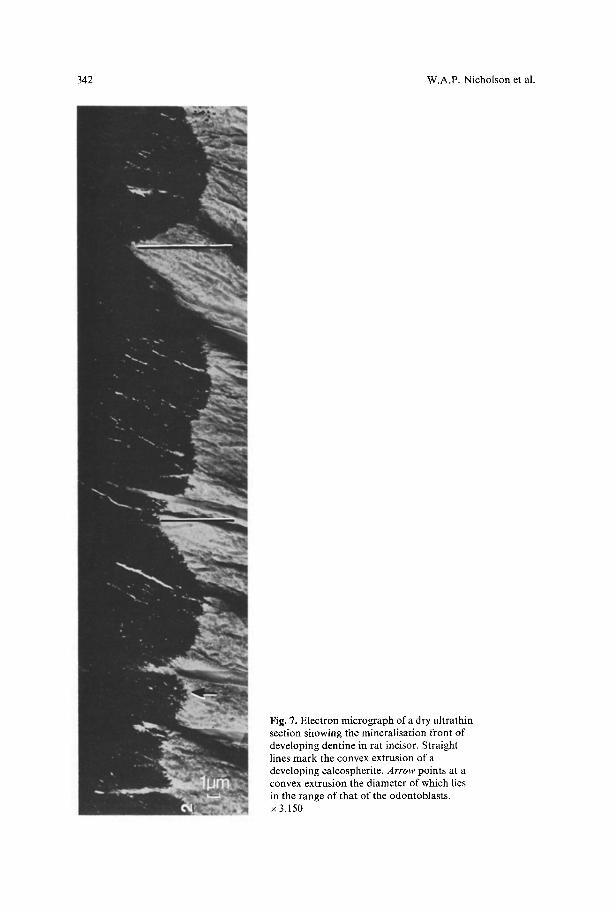
By examination of Figure 3 it can be seen that the two zones of Ca enrichment extend 12 I~m and 15 gm along the border, respectively. It is striking that the upper zone (point E) coincides with the region in which the development of the dentine front lags behind the neighbouring regions (point M). The lower zone is developing the typical convex form. This is also evident in Figures 4 and 5 where the compartments of the second stage of Ca enrichment are adjacent to a region in which the whole mineralisation front lags behind the neighbouring regions. From these observations it is interesting to compare the lateral extension of the zones of Ca enrichment with the general size of the characteristic convex regions of the dentine front (Fig. 7), which are in fact sections through the calcospherites of the mineralising surface. For this reason measurements were made of the width of about 100 regions which clearly had the convex shape (Fig. 7, lines). The width distribution lies between 6 gm and 30 ~tm, with a most frequent value of about 15 gm. It must be borne in mind that only a small part of the calco- spherites can be seen in ultrathin sections. Calcospherites are spherical to hemi- spherical in three dimensions and in the sections the calcospherites may be cut peripherally or centrally so that the observed lengths will show a large variation. It can be seen that the length of the Ca enriched zones in predentine in Figures 3 and 5 correspond with these higher values. It should be pointed out that whilst examining the mineralising dentine front, convex regions were observed such as can be seen in Figure 7 (arrow) which only had a width of about 4 -6 gm. Further measurements in ultrathin embedded sections demonstrated that in predentine Ca enriched zones of a similar size also exist.
Since the ultrathin sections can show only a narrow band through the zones of Ca enrichment, it was thought that the thicker 6 gm cryostat sections might better demonstrate the width of these zones. Figure 6 shows a three dimensional representation of the Ca distribution along the border and across the predentine of a Ca enriched zone in a cryostat section. The width of the enrichment zones along the border is about 15 gm which corresponds to the width of the zones in the ultrathin sections (Figs. 3-5) and the mean width of the calcospherites. It thus seems reasonable to suppose that the zones of a secondary Ca enrichment are at a stage which precedes mineralisation of the catcospherites, i.e. precedes their appearance as elements recognisable by morphological means.
Figure 6 also clearly indicates that the Ca-enrichment zones extend perpendi- cularly from the dentine border in the direction of the dentinal canals and a good length into the predentine, and that the Ca concentration decreases with increasing distance from the dentine border. In Figure 6 some areas of high Ca concentration can be found a short distance from the border, but these need not correspond to the maximum of the calcium enrichment. It may be that this location arises due to the angle of sectioning and that the maximum for the Ca concentration lies either above or below the section directly on the dentine border.
342 W.A.P. Nicholson et al.
Fig. 7. Electron micrograph of a dry ultrathin section showing the mineratisation front of developing dentine in rat incisor. Straight lines mark the convex extrusion of a developing calcospherite. Arrow points at a convex extrusion the diameter of which lies in the range of that of the odontoblasts. x 3,150

Electron Microprobe Investigations of Hard Tissue Formation 343
Biochemical Considerations
The calcium concentration found in predentine was approximately the same for treated and untreated cryostat sections, and for dry embedded and alcohol fixed-ultrathin sections. This indicates that the greater part of the calcium may be bound to macromolecules in the predentinal matrix.
A comparison of the high concentration of phosphorus (1 to 4 ~ W/W) found in untreated cryostat sections with the 1 ~ concentration found in dry ultrathin sections and the values of less than 0.1 ~ found in alcohol or water treated sections demonstrates that the greater fraction of phosphate groups can be easily washed out of the tissue. This indicates that the phosphate groups in predentine are present as ions and/or groups of low molecular weight which are not strongly bound to the matrix.
Recently it has been possible to study isolated bovine predentine directly to ascertain the relative proportions of orthophosphate, soluble pyro- and poly- phosphate and matrix bound phosphate (Ashton, Ashton, Boyde and HShling, in preparation; Carmichael et al., 1975). As can be seen from Table 2 ortho- phosphate accounts for 2/3 of the total phosphate present, the remainder being fairly equally divided between phosphoprotein phosphate and soluble phosphate esters.
The matrix associated phosphate is largely serine phosphate residues present in phosphoproteins which are released from the odontoblasts into the predentine adjacent to the mineralising border (Weinstock and Leblond, 1973) and which persist to some extent into the mineralised dentine, possibly in association with the insoluble collagenous fibres. The soluble phosphate ester pool is likely to be much more complicated, with contributions being made by pyrophosphate, nucleotide and hexose phosphates, polyphosphates and possibly anionic phospho- lipids.
Accepting that predentine has a moisture content of 66~ (Linde, 1973) and assuming that all the Ca (0.4~ W/W) and orthophosphate (5.6~) found in predentine are in solution, then the concentrations of Ca 2+ and HPO~ of 35 mMoles/litre and 122 mMoles/litre, respectively, would lead to an ion product far in excess of the 2 mMoles2/litre required for the spontaneous nucleation of hydroxyapatite in collagenous systems (Fleisch and Neuman, 1961).
We believe that the precipitation of apatite is temporarily prevented, firstly by the Ca-ions being bound to the organic matrix (e.g. to the SO 4 groups of proteoglycans or carboxyl groups of the acidic non-collagenous glycoproteins),
Table 2
~oo (W/W) of ~oo (W/W) of tissue total phosphate
1. Total phosphorus 2.9 2. Total P expressed as HPOg 8.2 3. EDTA-extractable orthophosphate 5.6 4. Phosphoprotein HPO, r (Carmichael et al., 1975) 1.1 5. Soluble polyphosphates [by subtraction 2.- (3 + 4)] 1.5
m
lOO 66 14 20

344 W.A.P. Nicholson et al.
and secondly because the phosphate groups that are present are bound to molecules such as phosphoserine, phosphatesters, A T P or pyrophosphate . The onset o f mineral nucleat ion cannot take place until the o r thophospha te has been split f rom the molecules.
It is interesting to note that the localisation o f alkaline phosphatase by electron mic roscopy has shown the enzyme to be present not only on the outer membrane o f odontoblas t s and their processes, but also directly on the dentine/predentine border ( F r o m m e et al., 1971, 1972; Larsson and Helander, 1974). Larsson and Helander (1974) reported further that the alkaline phosphatase found on the border can assume the funct ion o f a pyrophosphatase , in agreement with the chemical determinat ions o f W61tgens et al. (1971).
The normal Ca concentra t ion o f about 0 . 3 ~ is much higher than the value required for apatitic nucleation, so that the second stage o f Ca enr ichment in which even higher Ca concentra t ions are found is not necessary to provide the a m o u n t o f Ca required to produce mineralisation. Leonard et al. (1972) conclude that Ca A T P and ATPase m a y be necessary for the induct ion o f hard tissue mineralisation, Ca A T P hydrolysis being a source for the ions which fo rm apatite. Fur thermore , Magnusson (1974) has found by histochemical analysis that the ATPase in hard tissue forming cells needs the presence o f Ca to hydrolyse ATP. It is thus interesting to speculate whether the high concentrat ions o f Ca in the enriched zones in predentine will increase the hydrolysis o f phosphate esters by the induct ion o f the ATPase and be the stimulus for the onset ofapat i t ic nucleation. As the Ca is used up for the mineralisation o f that compar tment , the hydrolyt ic activity and hence the mineralisation m a y then be s topped by this internal regulation.
References
Appleton, T.C.: A cryostat approach to ultrathin 'dry' frozen sections for electron microscopy: a morphological and X-ray analytical study. J. Micr. 100, 49-74 (1974)
Ashton, B.A., H6hling, H.J., Nicholson, W.A.P., Zessack, U., Kriz, W., Boyde, A.: Quantitative analysis of Ca, P and S in mineralizing and non-mineralizing tissues. Naturwissenschaften 60, 392-393 (1973)
Baylink, D., Wergedal, J., Thompson, E.: Loss of protein polysaccharides at sites where bone mine- ralisation is initiated. J. Histochem. Cytochem. 20, 4, 279 292 (1972)
Bernard, G.W.: Ultrastructural observations of initial calcification in dentine and enamel. J. Ultra- struct. Res. 41, 1-7 (1972)
Carmichael, D.J., Chovelon, A., Pearson, C.H.: The composition of the insoluble collagenous matrix of bovine predentine. Calcif. Tiss. Res. 17, 263-271 (1975)
Croissant, R.D.: Isolation of an intercellular matrix "RNA-protein complex" during odontogenesis. J. dent. Res. 50, 1065 1071 (1971)
Dodgson, K.S., Price, R.G.: A note on the determination of the ester sulphate content of sulphated polysaccharides. Biochem. J. 84, 106 112 (1962)
Eisenmann, D.R., Glick, P.L.: Ultrastructure of initial crystal formation in dentine. J. Ultrastruct. Res. 41, 18-28 (1972)
Fiske, G.H., Subbarow, Y.: The colorimetric determination of phosphorus. J. biol. Chem. 66, 375-400 (1973)
Fleisch, H., Neumann, W.F.: Mechanism of calcification. Amer. J. Physiol. 200, 1296-1300 (1961) Fromme, H.G., H6hling, H.J., Riedel, H.: Elektronenmikroskopische Studien fiber die Dentin-
bildung. 1. Mitteilung: Lokalisation von Calcium und alkalischer Phosphatase. Dtsch. zahn~rztl. Z. 26, 359-364 (1971)

Electron Microprobe Investigations of Hard Tissue Formation 345
Fromme, H.G., H6hling, H.J., Riedel, H.: Elektronenmikroskopische Studien fiber die Dentin- bildung. 2. Mitteilung: Autoradiographische Studien zur Funktion der Odontoblasten. Dtsch. zahn~irztl. Z. 27, 6-13 (1972)
Hall, T.A.: Microprobe assay of chemical elements. In: Physical techniques in biological research, IA, 2nd ed., pp. 157-275. New York: Academic Press 1971
Hall, T.A., Anderson, C.H., Appleton, T.C.: The use of thin specimens for X-ray microanalysis in biology. J. Micr. 99, Pt2, 177-182 (1973)
Hall, T.A., Gupta, B.L.: Beam-induced loss of organic mass under electron microprobe conditions. J. Micr. 100, Pt 2, 177 188 (1974)
H6hling, H.J., Hall, T.A., Boyde, A.: Electron probe X-ray microanalysis of rat incisor peripheral dentine. Naturwissenschaften 54, 617-618 (1967)
H6hling, H.J., Hall, T.A., Boyde, A., von Rosenstiel, A.P.: Combined electron probe and electron diffraction analysis of prestages and early stages of dentine formation in rat incisor. Calcif. Tiss. Res. 2, Suppl. 5 (1968)
H6hling, H.J., Hall, T.A., Kriz, W., von Rosenstiel, A.P., Schnermann, J., Zessack, U.: Loss of mass in biological specimens during electron probe X-ray microanalysis. In: Modern techniques in physiological sciences (J.F. Gross, R. Kaufmann, E. Wetterer, eds.), pp. 335-343. New York: Academic Press 1973
H6hling, H.J., Nicholson, W.A.P., Schreiber, J., Zessack, U., Boyde, A.: The distribution of some elements in predentine and dentine of rat incisors. Naturwissenschaften 59, 423-424 (1972)
H6hling, H.J., Sch6pfer, H., H6hling, R.A., Hall, T.A., Gieseking, R.: The organic matrix of devel- oping tibia and femur, and macromolecular deliberations. Naturwissenschaften 57, 357 (1970)
Jessen, H., Peters, P.D., Hall, T.A. : Sulphur in different types of keratohyalin granules: a quantitative assay by X-ray microanalysis. J. Cell Sci. 15, 359-377 (1974)
Larsson, A., Helander, H.F.: Studies on dentinogenesis in the rat. Light, electronmicroscopic and histochemical studies on the interaction between lead pyrophosphate solutions and dentin- producing tissues. Calcif. Tiss. Res. 14, 87-104 (1974)
Leonard, F., Boke, J.W., Ruderman, R.J., Hegyeli, A.F.: Initiation and inhibition of subcutaneous calcification. Calcif. Tiss. Res. 10, 269-279 (1972)
Linde, A. : Glycosaminoglycans of the odontoblast-predentine layer in dentinogenically active porcine teeth. Calcif. Tiss. Res. 12, 281-294 (1973)
Lowry, O.H., Passoneav, J.V.: In: A flexible system of enzymatic analysis. New York-London: Academic Press 1972
Magnusson, B.C.: The effects of magnesium and calcium ions on phosphate activities at alkaline pH in the molar region of newborn mice. Histochemistry 42, 211-219 (1974)
Nicholson, W.A.P. : Experience of diffractive and non-diffractive analysis in the Cameca microprobe. In: Microprobe analysis of cells and tissues (T.A. Hall, P. Echlin, R. Kaufmann, eds.). New York- London: Academic Press 1974
Ozawa, H., Yajima, T. : Ultrastructure and cytochemistry of matrix vesicles in the developing cartilage and tooth germ. In: Proceedings of the 4th International Congress of Histochemistry and Cyto- chemistry, 1972, Kyoto, Japan (T. Takeuchi, K. Ogawa and S. Fujita, eds.) 1972
Pugliarello, M.C., Vittur, F., de Bernard, B., Bonucci, E., Ascenzi, A.: Chemical modification in osteones during calcification. Calcif. Tiss. Res. 5, 108-114 (1970)
Sisca, R.F., Provenza, D.V.: Initial dentine formation in human deciduous teeth. Calcif. Tiss. Res. 9, 1 16 (1972)
Slavkin, H.C., Bringas, P., Croissant, R.D., Bavetta, L.A.: Epithelial-mesenchymal interactions during odontogenesis. II. Intercellular matrix vesicles. Mech. Age. Dev. 1, 139 161 (1972)
Sundstr6m, B., Takuma, S.: A further contribution on the ultrastructure of calcifying cartilage. J. Ultrastruct. Res. 36, 419-424 (1971)
Weinstock, M., Leblond, C.P.: Radioautographic visualisation of a phosphorprotein at the min- eralisation front in the dentine of the rat incisor. J. Cell Biol. 56, 838-845 (1973)
W61tgens, J.H.M., Bonting, S.L., Bijvoet, O.L.M.: Relationship of inorganic phosphatase and alkaline phosphatase activities in hamster molars. Calcif. Tiss. Res. 5, 333-343 (1970)
Zimmermann, M. : Photometrische Metall- und Wasseranalysen mit Filterphotometern und Spektral- photometern, B -a5. Suttgart: Wissenschaftliche Verlagsgesellschaft m.b.H. 1961
Accepted October 5, 1976





























