Embed Size (px)
Citation preview

ABSTRACT: The mdx mouse is a model of Duchenne muscular dystrophy(DMD). As many DMD patients die of cardiac failure, we investigated wheth-er mdx mice exhibited clinically relevant cardiac phenotypes. We applied arecently developed method for noninvasively recording electrocardiograms(ECGs) to study male mdx mice (n = 15) and control mice (n = 15). The mdxmice had significant tachycardia and decreased heart rate variability, con-sistent with observations in DMD patients. Heart rate was nearly 15% fasterin mdx mice than control mice (P < 0.05). The rate-corrected QT intervalduration and PR interval were shorter in mdx compared to control mice (P <0.05). The muscarinic antagonist atropine significantly increased heart rateand decreased PR interval in C57 mice. In contrast, atropine significantlydecreased heart rate and increased PR interval in all mdx mice. Pharma-cological autonomic blockade and baroreflex sensitivity testing demon-strated an imbalance in autonomic nervous system modulation of heart rate,with decreased parasympathetic activity and increased sympathetic activityin mdx mice. Baseline ECGs and contrary responses to muscarinic blockadeby atropine in mice deficient in neuronal nitric oxide synthase (nNOS) sug-gest that the autonomic dysfunction in mdx mice may be independent ofdecreased myocardial nNOS. These electrocardiographic findings in dystro-phin-deficient mice may provide new bases for diagnosing, understanding,and treating DMD patients.
© 2002 Wiley Periodicals, Inc. Muscle Nerve 26: 513–519, 2002
ELECTROCARDIOGRAPHIC FINDINGS IN mdx MICE:A CARDIAC PHENOTYPE OF DUCHENNEMUSCULAR DYSTROPHY
VICTOR CHU, MSc,1 JOSE M. OTERO, MSc,2 ORLANDO LOPEZ, MSc,2 MATTHEW F. SULLIVAN, BSc,1
JAMES P. MORGAN, MD,1 IVO AMENDE, MD,1 and THOMAS G. HAMPTON, PhD1*
1 Cardiovascular Division, Beth Israel Deaconess Medical Center, Harvard Medical School, 330 Brookline Avenue,Boston, Massachusetts 02215, USA2 The CuraVita Corporation, Boston, Massachusetts, USA
Accepted 15 May 2002
Patients with Duchenne muscular dystrophy(DMD) have distinctive electrocardiograms (ECGs),many presenting with tachycardia27,29 and signifi-cant decreases in heart rate variability.52,53 The elec-trocardiographic abnormalities have been ascribedto various mechanisms, including postural syn-
dromes that exaggerate cardiac reflexes, compensa-tory mechanisms for impaired cardiac contractility,and autonomic dysfunction.18,31,33 Yet, the patho-genesis of electrical and autonomic disturbances inDMD patients remains unclear.
The dystrophin-deficient (mdx) mouse is a modelof DMD7, but its suitability for investigating thepathogenesis and treatment of cardiac defects is di-minished by a lack of cardiac phenotype. Torres andDuchen did not detect pathological changes in heartmuscle of mdx mice.44 Megeney et al. reported thatmdx mice rarely display histological evidence of car-diac abnormalities.30 Muscular dystrophy and car-diomyopathy are more severe in mice lacking utro-phin and dystrophin compared to mdx mice,19
indicating that a homologous protein effectivelycompensates for the lack of dystrophin in mdxmice.19,30 Yet, in vitro myocardial contractile prop-erties are altered in mdx mice.38 Functional changesin cardiac receptors that mediate the role of the au-
* Disclosure: Thomas G. Hampton is owner of Mouse Specifics, Inc., acompany commercializing the ECG technology described in this paper.He is also president of The CuraVita Corporation, a nonprofit organizationfor applying noninvasive technologies to characterize mouse models ofhuman diseases.Abbreviations: CV, coefficient of variance; DMD, Duchenne musculardystrophy; ECG, electrocardiogram; HR, heart rate; HRV, heart rate vari-ability; mdx, dystrophin-deficient mouse; nNOS, neuronal nitric oxide syn-thase; nNOS−/−, neuronal nitric oxide synthase–deficient; QTc, rate-corrected QT intervalsKey words: dystrophin; electrocardiogram; mdx; autonomic nervous sys-temCorrespondence to: T. G. Hampton; e-mail: [email protected]
© 2002 Wiley Periodicals, Inc.Published online 31 July 2002 in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/mus.10223
ECGs in mdx Mice MUSCLE & NERVE October 2002 513

tonomic nervous system have been described in mdxmice.25 Anesthetized mdx mice manifested abnormalECGs2 that were associated with a reduction in neu-ronal nitric oxide synthase (nNOS) activity.2,36 Wesurmised that subtle cardiac phenotypes associatedwith dystrophin deficiency might be more evident inthe conscious animal. Accordingly, we investigatedelectrocardiographic and autonomic phenotypicchanges in conscious mdx and nNOS-deficient(nNOS−/−) mice using a recently developed nonin-vasive system for screening ECGs.11
MATERIALS AND METHODS
Mice. Male control C57BL/10ScSn (C57, n = 15)and male mutant (mdx, n = 15) mice (10–12 weeksold) were obtained from The Jackson Laboratory(Bar Harbor, Maine). The animals were housed instandard conditions within the Animal Resource Fa-cility at the Beth Israel Deaconess Medical Center.Handling and care of mice were consistent with fed-eral and institutional guidelines.
ECG Recording. ECGs were recorded in consciousmice as described previously.11 Briefly, mice weregently removed from their cages and positioned onthe ECG recording platform (AnonyMOUSE, MouseSpecifics, Inc., Boston, Massachusetts). An array ofgel-coated ECG electrodes were embedded in thefloor of the platform and spaced to provide contactbetween the electrodes and animals’ paws. The elec-trodes were connected to an amplifier by threeshielded conductive leads. As even modest handlingof mice induces significant alterations in heartrate,15 each mouse was permitted to acclimatize for10 min prior to collection of baseline data. The sig-nals were digitized with 16-bit precision at a sam-pling rate of 2,000 samples/s. When mice were sit-ting or otherwise positioned such that the paws werenot in contact with three electrodes, the output fromthe amplifier was discarded. Only data from continu-ous recordings of 15–25 ECG signals were used inthe analyses.
ECG Analyses. Analyses of the ECGs were per-formed as described previously.11 Each signal wasanalyzed using e-MOUSE, Internet-based physi-ologic waveform analyses software. The program in-corporates Fourier analyses and linear time-invariantdigital filtering of frequencies below 2 HZ and above100 HZ to minimize signal environmental distur-bances. The software uses a peak detection algo-rithm to find the peak of the R waves and to calculateheart rate (HR). In accord with Mitchell et al.,32 weincluded the inverted and/or biphasic portions of
the T wave in our calculations of the QT interval.The QT intervals were rate corrected (QTc) by analgorithm developed for use in mice.32 The softwareplots its interpretation of P,Q,R,S, and T for eachbeat so that spurious data resulting from unfilterednoise or motion artefacts may be rejected. The meanof the ECG time intervals for each set of waveformsis then calculated. Heart rate variability (HRV) wascalculated as the standard deviation of all R–R inter-vals for each set of ECG signals, and the coefficientof variance (CV,%) was calculated as the ratio ofmean HRV to mean HR.16
Autonomic Blockade. Parasympathetic and sympa-thetic nervous system control of heart rate regula-tion were investigated by pharmacological block-ade.1 6 , 2 1 , 4 7 Mice received intraperi tonealadministration of atropine (0.5 mg/kg) for parasym-pathetic blockade (n = 15 mdx, n = 15 C57), propran-olol (1 mg/kg) for sympathetic blockade (n = 10mdx, n = 7 C57), and atropine (0.5 mg/kg) pluspropranolol (1 mg/kg) for combined �-adrenergicand muscarinic cholinergic blockade (n = 10 mdx,n = 7 C57).16,21 Baseline ECGs were recorded within5 min prior to each administration and after a 5–10min equilibrium phase after drug administration toallow for HR stabilization. The pharmacological in-terventions were performed on different days to pre-vent interference between drugs. The dosages of at-ropine and propranolol were those used by othersinvestigating autonomic blockade in consciousmice.16,21
Baroreflex HR Modulation. Mice were administeredintraperitoneal phenylephrine (3 mg/kg) after �-ad-renergic blockade with propranolol (1 mg/kg) totest for a baroreflex-mediated cardio-inhibitory re-sponse (n = 10 mdx, n = 7 C57).16,21 After baselinerecordings, ECGs were recorded after a 5–10 minequilibrium phase following propranolol administra-tion to allow for HR stabilization, and then within 2min following phenylephrine administration.Changes in ECG indices following phenylephrinewere determined relative to those recorded follow-ing propranolol administration.
Role of Neuronal Nitric Oxide. To investigatewhether our observations were linked to deficiencyin neuronal nitric oxide, we performed similar pro-tocols as described above in mice deficient in nNOS(nNOS−/−, n = 7) and B6129SF2/J control mice(B6129, n = 6). These mice were obtained from TheJackson Laboratories and were housed and cared foras described above.
514 ECGs in mdx Mice MUSCLE & NERVE October 2002

Statistics. Data are presented as the means ± SE.Comparisons between mdx and C57 mice and be-tween nNOS−/− and B6129 mice were performed us-ing Student’s t-test for unpaired observations. Effectsof autonomic blockade or baroreflex sensitivity test-ing within groups were performed using Student’st-test for paired observations, and between groupcomparisons using Student’s t-test for unpaired ob-servations. Differences were considered significantwith P < 0.05.
RESULTS
Baseline HR Dynamics. Table 1 summarizes theECG data in mdx and C57 control mice. At baseline,HR was significantly higher in the mdx mice. Themdx mice had significantly shorter QRS duration,shorter QT interval, and shorter QTc interval. PRinterval was significantly shorter in mdx mice, andHRV and CV were significantly lower in mdx thanC57 mice. Figure 1 illustrates representative ECG re-cordings from a C57 control mouse (top panel) andan mdx mouse (bottom panel). As the mice are con-scious and ambulatory, baseline artifacts and noiseare apparent in the unfiltered signals. Nevertheless,the P waves and T waves are discernible and inter-pretable by the ECG analyses software.
Autonomic Blockade. The effects of parasympa-thetic, sympathetic, and combined blockade on HRand HRV are shown in Figure 2. Atropine adminis-tration in mdx mice led to significantly greaterchanges in HR (P < 0.05) and significantly smallerchanges in HRV (P < 0.05) compared to the effectsin C57 mice. HR decreased in all 15 mdx mice butincreased in all but 2 of the C57 mice after admin-istration of atropine. Propranolol administration ex-erted a more modest reduction in HR in mdx mice(−60 ± 27 bpm, n = 10) than C57 mice (−122 ± 53bpm, n = 7) (P = NS). Propranolol administrationled to greater changes in HRV in C57 (+7.2 ± 4.0 ms)
than in mdx mice (−2.0 ± 1.5 ms) (P < 0.05). Com-bined autonomic blockade resulted in a slightlylarger decrease in HR in mdx than C57 mice. Therewas no difference in HR and HRV between the twogroups after combined atropine and propranolol ad-ministration.
Baroreflex Pressor Response. The HR response tophenylephrine was examined in C57 (n = 7) and mdxmice (n = 10). The pressor challenge was performedafter �-adrenergic blockade so that an observedchange in HR or HRV could be attributed to activa-tion of the inhibitory limb of the baroreflex and notthe result of sympathetic withdrawal.16,21 As shown
Table 1. Baseline electrocardiographic parameters and heart rate variability in C57, mdx, B6129, and nNOS−/− mice.†
Variable C57 (n = 15) mdx (n = 15) B6129 (n = 6) nNOS−/− (n = 7)
HR (bpm) 706 ± 13 809 ± 5* 765 ± 10 686 ± 22*HRV (ms) 2.7 ± 0.9 0.5 ± 0.1* 0.8 ± 0.2 2.6 ± 0.8CV (%) 2.9 ± 0.9 0.7 ± 0.1* 1.1 ± 0.2 2.7 ± 0.8PR interval (ms) 29.4 ± 0.5 27.2 ± 0.4* 29.4 ± 0.8 27.7 ± 0.4QRS duration (ms) 8.3 ± 0.2 7.5 ± 0.1* 7.8 ± 0.1 8.6 ± 0.2*QT interval (ms) 57.2 ± 2.0 47.8 ± 0.5* 49.9 ± 0.5 60.8 ± 2.9*QTc interval (ms) 61.6 ± 1.5 55.5 ± 0.5* 56.3 ± 0.4 64.8 ± 2.0*
CV, coefficient of variance; HR, heart rate; HRV, heart rate variability.†Values are mean ± SE.*P < 0.05.
FIGURE 1. Electrocardiograms from a conscious male C57 con-trol mouse (top) and a male mdx mouse (bottom). In the controlmouse, HR = 595 bpm, QRS = 9.6 ms, and QT = 75.0 ms. In themdx mouse, HR = 814 bpm, QRS = 7.5 ms, and QT = 49.6 ms.
ECGs in mdx Mice MUSCLE & NERVE October 2002 515

in Figure 3, mdx mice administered phenylephrinedemonstrated smaller reductions in HR (P < 0.05)and smaller changes in HRV (P < 0.05) compared toC57 mice. After phenylephrine administration, HRVremained smaller in mdx than C57 mice (8.5 ± 2.5 msvs. 28.6 ± 4.1 ms, P < 0.05) and CV remained signifi-cantly smaller in mdx than C57 mice (6.0 ± 1.7% vs.15.8 ± 2.4%, P < 0.05).
Effects of nNOS Deficiency. Seven male nNOS−/−
mice and six male B6129 control mice were sub-jected to the same protocols described above. Asshown in Table 1, HR was significantly lower in thenNOS−/− mice and HRV tended to be greater innNOS−/− than B6129 mice. After atropine adminis-tration to nNOS−/− mice, HR increased from 686 ±22 bpm to 721 ± 12 bpm (P = 0.07), HRV decreasedfrom 2.6 ± 0.8 ms to 0.7 ± 0.1 ms (P < 0.05), and CVdecreased from 2.7 ± 0.8% to 0.8 ± 0.1% (P < 0.05).In nNOS−/− mice, �-adrenergic blockade reducedHR by 17 ± 6 bpm (P < 0.05) and combined auto-nomic blockade reduced HR by 112 ± 3 bpm (P <0.05). As shown in Figure 4, phenylephrine admin-
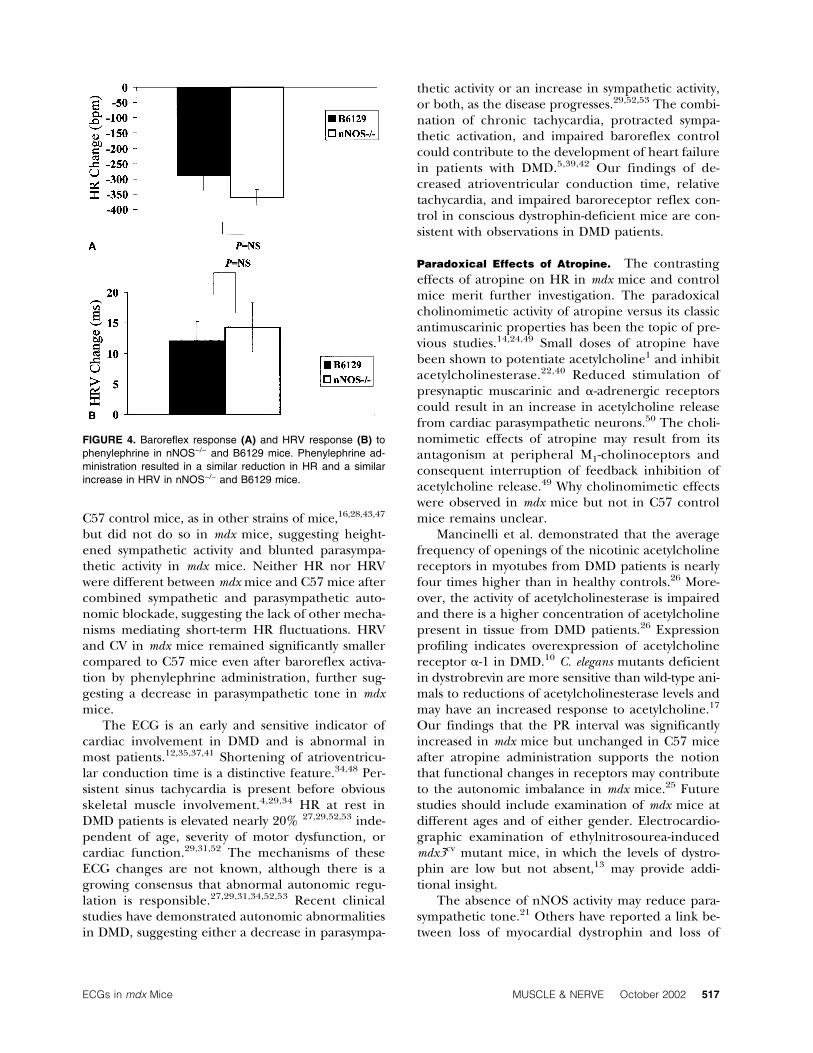
istration resulted in similar reductions in HR (−360 ±6 bpm vs. −288 ± 39 bpm, P = NS) and similar in-creases in HRV (+14.3 ± 3.9 ms vs. +12.1 ± 3.2 ms, P= NS) in nNOS−/− and B6129 mice. After phenyleph-rine administration, HRV and CV were similar innNOS−/− and B6129 mice.
DISCUSSION
Mice deficient in dystrophin have increased heartrate, decreased heart rate variability, and altered au-tonomic heart rate modulation in comparison towild-type control mice, findings consistent with clini-cal observations of autonomic dysfunction in DMDpatients.27,52,53 The elevated HR of mdx mice couldreflect increased sympathetic tone, reduced para-sympathetic tone, or both. In control mice, atropineincreased HR and decreased HRV, consistent with itswell-established antimuscarinic properties.15,49 Bycontrast, atropine significantly decreased HR in allmdx mice, suggesting a simple test that may be pre-dictive of dystrophin-deficiency.
�-Adrenergic blockade with propranolol pro-duced a more modest decrease in HR in mdx thanC57 mice, suggesting an elevated sympathetic tonein mdx mice. Propranolol increased HRV indices in
FIGURE 2. Response of HR (A) and HRV (B) to atropine, pro-pranolol, and the combination of both drugs, administered on 3separate days. After atropine administration, HR decreased 90 ±14 bpm in mdx mice, whereas HR increased 21 ± 12 bpm in C57mice (P < 0.05). HRV changes were smaller in mdx mice (+0.1 ±0.1 ms) than in C57 mice (−2.1 ± 1.1 ms) (P < 0.05).
FIGURE 3. Baroreflex response (A) and HRV response (B) tophenylephrine in mdx and C57 mice. The reduction in HR afterphenylephrine administration was significantly attenuated in mdxmice (−132 ± 27 bpm) compared to C57 mice (−312 ± 27 bpm) (P< 0.05). The increase in HRV following phenylephrine adminis-tration was blunted in mdx mice (+6.2 ± 2.7 ms) compared to C57mice (+21.3 ± 5.2 ms) (P < 0.05).
516 ECGs in mdx Mice MUSCLE & NERVE October 2002
C57 control mice, as in other strains of mice,16,28,43,47
but did not do so in mdx mice, suggesting height-ened sympathetic activity and blunted parasympa-thetic activity in mdx mice. Neither HR nor HRVwere different between mdx mice and C57 mice aftercombined sympathetic and parasympathetic auto-nomic blockade, suggesting the lack of other mecha-nisms mediating short-term HR fluctuations. HRVand CV in mdx mice remained significantly smallercompared to C57 mice even after baroreflex activa-tion by phenylephrine administration, further sug-gesting a decrease in parasympathetic tone in mdxmice.
The ECG is an early and sensitive indicator ofcardiac involvement in DMD and is abnormal inmost patients.12,35,37,41 Shortening of atrioventricu-lar conduction time is a distinctive feature.34,48 Per-sistent sinus tachycardia is present before obviousskeletal muscle involvement.4,29,34 HR at rest inDMD patients is elevated nearly 20% 27,29,52,53 inde-pendent of age, severity of motor dysfunction, orcardiac function.29,31,52 The mechanisms of theseECG changes are not known, although there is agrowing consensus that abnormal autonomic regu-lation is responsible.27,29,31,34,52,53 Recent clinicalstudies have demonstrated autonomic abnormalitiesin DMD, suggesting either a decrease in parasympa-
thetic activity or an increase in sympathetic activity,or both, as the disease progresses.29,52,53 The combi-nation of chronic tachycardia, protracted sympa-thetic activation, and impaired baroreflex controlcould contribute to the development of heart failurein patients with DMD.5,39,42 Our findings of de-creased atrioventricular conduction time, relativetachycardia, and impaired baroreceptor reflex con-trol in conscious dystrophin-deficient mice are con-sistent with observations in DMD patients.
Paradoxical Effects of Atropine. The contrastingeffects of atropine on HR in mdx mice and controlmice merit further investigation. The paradoxicalcholinomimetic activity of atropine versus its classicantimuscarinic properties has been the topic of pre-vious studies.14,24,49 Small doses of atropine havebeen shown to potentiate acetylcholine1 and inhibitacetylcholinesterase.22,40 Reduced stimulation ofpresynaptic muscarinic and �-adrenergic receptorscould result in an increase in acetylcholine releasefrom cardiac parasympathetic neurons.50 The choli-nomimetic effects of atropine may result from itsantagonism at peripheral M1-cholinoceptors andconsequent interruption of feedback inhibition ofacetylcholine release.49 Why cholinomimetic effectswere observed in mdx mice but not in C57 controlmice remains unclear.
Mancinelli et al. demonstrated that the averagefrequency of openings of the nicotinic acetylcholinereceptors in myotubes from DMD patients is nearlyfour times higher than in healthy controls.26 More-over, the activity of acetylcholinesterase is impairedand there is a higher concentration of acetylcholinepresent in tissue from DMD patients.26 Expressionprofiling indicates overexpression of acetylcholinereceptor �-1 in DMD.10 C. elegans mutants deficientin dystrobrevin are more sensitive than wild-type ani-mals to reductions of acetylcholinesterase levels andmay have an increased response to acetylcholine.17
Our findings that the PR interval was significantlyincreased in mdx mice but unchanged in C57 miceafter atropine administration supports the notionthat functional changes in receptors may contributeto the autonomic imbalance in mdx mice.25 Futurestudies should include examination of mdx mice atdifferent ages and of either gender. Electrocardio-graphic examination of ethylnitrosourea-inducedmdx3cv mutant mice, in which the levels of dystro-phin are low but not absent,13 may provide addi-tional insight.
The absence of nNOS activity may reduce para-sympathetic tone.21 Others have reported a link be-tween loss of myocardial dystrophin and loss of
FIGURE 4. Baroreflex response (A) and HRV response (B) tophenylephrine in nNOS−/− and B6129 mice. Phenylephrine ad-ministration resulted in a similar reduction in HR and a similarincrease in HRV in nNOS−/− and B6129 mice.
ECGs in mdx Mice MUSCLE & NERVE October 2002 517

nNOS activity 6,8,9 and have associated ECG abnor-malities in the mdx heart with reduction of nNOS.2,36
We recorded ECGs in nNOS-deficient mice to estab-lish whether the absence of nNOS activity mirroredthe ECG phenotypes observed in the absence of dys-trophin. Our nNOS−/− mice exhibited slower HRand increased HRV than B6129 controls, as opposedto mdx mice that exhibited faster HR and reducedHRV than C57 controls. In nNOS−/− mice, HR in-creased by 35 ± 15 bpm after atropine administra-tion, comparable to findings in mice with intactparasympathetic function.21,47 Atropine significantlyreduced HRV and CV in nNOS−/− mice but not inmdx mice. In the control mice for mdx mice andnNOS−/− mice (C57 and B6129, respectively), phen-ylephrine administration resulted in equivalent re-ductions in HR. Moreover, phenylephrine resultedin similar reductions in HR in nNOS−/− mice andB6129 control mice. In mdx mice, however, thebaroreflex response was nearly absent. These obser-vations suggest that decreased myocardial nNOS it-self does not completely account for the perturbedparasympathetic tone elicited by the baroreflex inmdx mice.
Study Limitations. Mice are extremely sensitive toeven modest handling3,23 and transport.3,46 Ourmeasurements were made in daytime hours, disrupt-ing the less active phases of the mouse circadiancycle. The differences in HR and ECG parametersbetween mdx and C57 mice and between nNOS−/−
and B6129 may reflect variations in behavioral re-sponses to handling and transport. We allowed a 10-min acclimation period prior to recording data toattenuate these effects. We assumed complete effectsof pharmacologic blockade with propranolol and at-ropine using equivalent doses administered to allmice. The doses we used are similar to those de-scribed by others to achieve complete autonomicblockade,16,20,21,23,47 but it is possible that the differ-ent strains of mice have differing sensitivities to theseagents. Our HR and ECG data are comparable tothose obtained using radiotelemetry,16,21,47 but ourrecordings are free of the effects of anesthetic, sur-gery, or implanted devices, all of which could masksubtle cardiovascular phenotypes.51 These measure-ments are based on ECG records of relatively shortduration (∼2 s) providing 15 to 25 ECG signals. Theduration of the ECG recording should be consid-ered in the interpretation of heart rate variability.45
Mr. Kenneth Gladstone provided excellent administrative sup-port. This work was supported in part by the NHLBI Program forGenomic Applications HL066582 (Genomics of CardiovascularDevelopment, Adaptation, and Remodeling) to V. Chu and J.
Morgan. I. Amende was generously supported by Forderkreis zurVerbesserung des Gesundheitswesens e.V.
REFERENCES
1. Ashford A, Penn GB, Ross JW. Cholinergic activity of atro-pine. Nature 1962;193:1082–1083.
2. Bia BL, Cassidy PJ, Young ME, Rafael JA, Leighton B, DaviesKE, Radda GK, Clarke K. Decreased myocardial nNOS, in-creased iNOS and abnormal ECGs in mouse models of Du-chenne muscular dystrophy. J Mol Cell Cardiol 1999;31:1857–1862.
3. Blizard DA, Welty R. Cardiac activity in the mouse: straindifferences. J Comp Physiol Psychol 1971;77:337–344.
4. Boland BJ, Silbert PL, Groover RV, Wollan PC, SilversteinMD. Skeletal, cardiac, and smooth muscle failure in Du-chenne muscular dystrophy. Pediatr Neurol 1996;14:7–12.
5. Brandle M, Wang W, Zucker IH. Hemodynamic correlates ofbaroreflex impairment of heart rate in experimental canineheart failure. Basic Res Cardiol 1996;91:147–154.
6. Brenman JE, Chao DS, Xia H, Aldape K, Bredt DS. Nitricoxide synthase complexed with dystrophin and absent fromskeletal muscle sarcolemma in Duchenne muscular dystro-phy. Cell 1995;82:743–752.
7. Bulfield G, Siller WG, Wight PAL, Moore KJ. X chromosome-linked muscular dystrophy (mdx) in the mouse. Proc NatlAcad Sci USA 1984;81:1189–1192.
8. Chang W, Iannaccone T, Lau K, Masters BSS, McCabe TJ,McMillan K, Padre RC, Spencer MJ, Tidball JG, Stull JT. Neu-ronal nitric oxide synthase and dystrophin-deficient musculardystrophy. Proc Natl Acad Sci USA 1996;93:9142–9147.
9. Chao DS, Silvagno F, Bredt DS. Muscular dystrophy in mdxmice despite lack of neuronal nitric oxide synthase. J Neuro-chem 1998;71:784–789.
10. Chen YW, Zhao P, Borup R, Hoffman EP. Expression profil-ing in the muscular dystrophies: identification of novel as-pects of molecular pathophysiology. J Cell Biol 2000;151:1321–1336.
11. Chu V, Otero JM, Lopez O, Morgan JP, Amende I, HamptonTG. Method for non-invasively recording electrocardiogramsin conscious mice. BMC Physiol 2001;1:6.
12. Cox GF, Kunkel LM. Dystrophies and heart disease. CurrOpin Cardiol 1997;12:329–343.
13. Danko I, Chapman V, Wolff JA. The frequency of revertants inmdx mouse genetic models for Duchenne muscular dystro-phy. Pediatr Res 1992;32:128–131.
14. Das G, Talmers FN, Weissler AM. New observations on theeffects of atropine on the sinoatrial and atrioventricularnodes in man. Am J Cardiol 1975;36:281–285.
15. Desai KH, Sato R, Schauble E, Barish GS, Kobilka BK, Ber-stein D. Cardiovascular indexes in the mouse at rest and withexercise: new tools to study models of cardiac disease. Am JPhysiol Heart Circ Physiol 1997;272:H1053–H1061.
16. Gehrmann J, Hammer PE, Maguire CT, Wakimoto H, Tried-man JK, Berul CI. Phenotypic screening for the heart ratevariability in the mouse. Am J Physiol Heart Circ Physiol 2000;279:H733–H740.
17. Gieseler K, Mariol MC, Bessou C, Migaud M, Franks CJ,Holden-Dye L, Segalat L. Molecular, genetic and physiologi-cal characterization of dystrobrevin-like (dyb-1) mutants ofCaenorhabditis elegans. J Mol Biol 2001;307:107–117.
18. Gilroy J, Cahalan J, Berman R, Newman M. Cardiac and pul-monary complications in Duchenne’s progressive musculardystrophy. Circulation1963;1:484–493.
19. Grady MR, Teng H, Nichol MC, Cunningham JC, WilkinsonRS, Sanes JR. Skeletal and cardiac myopathies in mice lackingutrophin and dystrophin: a model for Duchenne musculardystrophy. Cell 1997;90:729–738.
20. Johansson C, Vennstrom B, Thoren P. Evidence that de-creased heart rate in thyroid hormone receptor-�-deficient
518 ECGs in mdx Mice MUSCLE & NERVE October 2002

mice is an intrinsic effect. Am J Physiol Regul Integr CompPhysiol 1998;275:R640–R646.
21. Jumrussirikul P, Dinerman J, Dawson TM, Dawson VL, Eke-lund U, Georgakopoulos D, Schramm LP, Calkins H, SnyderSH, Hare JM, Berger MD. Interaction between neuronal ni-tric oxide synthase and inhibitory G protein activity in heartrate regulation in conscious mice. J Clin Invest 1998;102:1279–1285.
22. Kato G. NMR studies on drug receptor interactions. Int J ClinPharmacol Res 1971;5:12–19.
23. Kramer K, van Acker SA, Voss H-P, Grimbergen JA, van derVijgh WJF, Bast A. Use of telemetry to record electrocardio-gram and heart rate in freely moving mice. J Pharmacol Toxi-col Methods 1993;30:209–215.
24. Lonnerholm G, Widerlov E. Effect of intravenous atropineand methylatropine on heart rate and secretion of saliva inman. Eur J Clin Pharmacol 1975;8:233–240.
25. Lu S, Hoey A. Changes in function of cardiac receptors me-diating the effects of the autonomic nervous system in mus-cular dystrophy (mdx) mouse. J Mol Cell Cardiol 2000;32:143–152.
26. Mancinelli E, Sardini A, D’Aumiller A, Meola G, Martucci G,Cossu G, Wanke E. Properties of acetylcholine-receptor acti-vation in human Duchenne muscular dystrophy myotubes.Proc R Soc Lond [Biol] 1989;237:247–257.
27. Marcello N, Baratti M, Ortaggio F, Vescovini E, Zanoni P,Tugnoli V, De Grandis D. Autonomic function and the sinustachycardia of Duchennne muscular dystrophy. ElectromyogrClin Neurophysiol 1995;35:387–395.
28. Marselos M, Eriksson K, Hanninen O. Effects of �-adrenocep-tor blocking drugs, physostigmine, and atropine on the tox-icity of doxepin in mice. Med Biol 1975;53:231–237.
29. Maruyama T, Fujino T, Fukuoka Y, Tsukamoto K, Mawatarti S.Notched T wave as evidence of autonomic nervous lability inDuchenne progressive muscular dystrophy. Jpn Heart J 1995;36:741–750.
30. Megeney LA, Kablar B, Perry RLS, Ying C, May L, RudnickiMA. Severe cardiomyopathy in mice lacking dystrophin andMyoD. Proc Natl Acad Sci USA 1999;96:220–225.
31. Miller G, D’Orsogna L, O’Shea JP. Cardiomyopathy of Du-chenne muscular dystrophy. Brain Dev 1989;11:247–250.
32. Mitchell GF, Jeron A, Gideon K. Measurement of heart rateand Q-T interval in the conscious mouse. Am J Physiol HeartCirc Physiol 1998;274:H747–H751.
33. Nigro G, Comi LI, Politano L, Bain RJI. The incidence andevolution of cardiomyopathy in Duchenne muscular dystro-phy. Int J Cardiol 1990;26:271–277.
34. Perloff JK. Cardiac rhythm and conduction in Duchenne’smuscular dystrophy: a prospective study in 20 patients. J AmColl Cardiol 1984;3:1263–1268.
35. Perloff JK. Neurological disorders and heart disease. In:Braunwald E, editor. Heart disease. Philadelphia: WB Saun-ders; 1998. p 1782–1783.
36. Radda GK. Of mice and men: from early NMR studies of theheart to physiological genomics. Biochem Biophys Res Com-mun 1999;266:723–728.
37. Sanyal SK, Johnson WW, Thapar MK, Pitner SE. An ultrastruc-
tural basis for electrocardiographic alterations associated withDuchenne’s progressive muscular dystrophy. Am J Cardiol1978;6:1122–1129.
38. Sapp JL, Bobet J, Howlett SE. Contractile properties of myo-cardium are altered in dystrophin-deficient mdx mice. J Neu-rol Sci 1996;142:17–24.
39. Schwarz PJ, Stone HL. The role of the autonomic nervoussystem in sudden coronary death. Ann NY Acad Sci 1982;382:162–180.
40. Sekiya A. Action of atropine on rabbit blood cholinesterase.Jpn J Pharmacol 1954;4:22–23.
41. Slucka C. The electrocardiogram in Duchenne progressivemuscular dystrophy. Circulation 1968;38:933–940.
42. Stewart JM. Autonomic nervous system dysfunction in adoles-cents with postural orthostatic tachycardia syndrome andchronic fatigue syndrome is characterized by attenuated vagalbaroreflex and potential sympathetic vasomotion. Pediatr Res2000;48:218–226.
43. Swynghedauw B, Jasson S, Chevalier B, Clairambault J, Har-douin S, Heymes C, Mangin L, Mansier P, Medigue C, MoalicJ, Thibault N, Carre F. Heart rate and heart rate variability, apharmacological target. Cardiovasc Drugs Ther 1997;10:677–685.
44. Torres LFB, Duchen LW. Morphological studies of nerves,muscles and end-plates. Brain 1987;110:269–299.
45. Tsuji H, Venditti FJ, Manders ES, Evans JC, Larson MG, Feld-man CL, Levy D. Determinants of heart rate variability. J AmColl Cardiol 1996;28:1539–1546.
46. Tuli JS, Smith A, Morton DB. Stress measurements in miceafter transportation. Lab Anim 1995;29:132–138.
47. Uechi M, Asai K, Osaka M, Smith A, Sao N, Wagner TE,Ishikawa Y, Hayakawa H, Vatner DE, Shannon RP, Homcy CJ,Vatner SF. Depressed heart rate variability and arterial barore-flex in conscious transgenic mice with over-expression of car-diac G�. Circ Res 1998;82:416–423.
48. Wahi PL. Cardiac changes in myopathy. Am Heart J 1963;66:749–751.
49. Wellstein A, Pitschner HF. Complex dose-response curves ofatropine in man explained by different functions of M1- andM2-cholinoceptors. Naunyn Schmiedebergs Arch Pharmacol1988;338:19–27.
50. Wetzel GT, Brown JH. Presynaptic modulation of acetylcho-line release from cardiac parasympathetic neurons. Am JPhysiol 1985;248:H33–H95. Mitchell GF, Jeron A, Gideon K.Measurement of heart rate and Q-T interval in the consciousmouse. Am J Physiol Heart Circ Physiol 1998;274:H747–H751.
51. Wickman K, Nemec J, Gendler S, Clapham D. Abnormalheart rate regulation in GIRK4 knockout mice. Neuron 1998;20:103–114.
52. Yotsukura M, Fujii K, Katayama A, Tomono Y, Ando H, SakataK, Ishihara T, Ishikawa K. Nine-year follow-up study of heartrate variability in patients with Duchenne-type progressivemuscular dystrophy. Am Heart J 1998;136:289–296.
53. Yotsukura M, Sasaki K, Kachi E, Sasaki A, Ishihara T, IshikawaK. Circadian rhythm and variability of heart rate in Du-chenne-type progressive muscular dystrophy. Am J Cardiol1995;76:947–951.
ECGs in mdx Mice MUSCLE & NERVE October 2002 519





























