Embed Size (px)
Citation preview

www.elsevier.com/locate/brainres
Brain Research 1044
Research report
Electroacupuncture attenuates morphine withdrawal signs and c-Fos
expression in the central nucleus of the amygdala in freely moving rats
Sheng Liua, Wenhua Zhoub,T, Huifen Liub, Guodong Yangb,T, Weikang Zhaoa
aShanghai University of Traditional Chinese Medicine, Shanghai, 200032, P.R. ChinabNingbo Addiction Research and Treatment Center, 42, Xibeijie Street, Ningbo 315010, P.R. China
Accepted 23 February 2005
Available online 15 April 2005
Abstract
Experimental efforts for understanding the mechanisms of electroacupuncture (EA) for opiate addiction are partially hampered by restraint
stress. In unrestrained animals, it is difficult to perform EA stimulation at acupuncture points frequently selected on the four limbs. The
present study was performed to evaluate the effect of EA at the acupuncture point Shen-Shu (BL.23) on morphine withdrawal signs and c-Fos
expression of the amygdala in freely moving rats or restrained rats. We applied immunohistochemistry to detect c-Fos-positive nuclei.
Corticosterone levels and behavioral responses were measured during EA stimulation. The needles were bilaterally inserted and fixed at
BL.23, and 100-Hz electric stimulation was conducted 30 min before naloxone-precipitated withdrawal. In both freely moving rats and
restrained rats, EA significantly reduced the signs of morphine withdrawal. Notably, EA stimulation in freely moving rats attenuated c-Fos
expression in the central nucleus of the amygdala while EA in restrained animals increased this response. In addition, the restrained rats
emitted greater levels of vocalization and facial expression than freely moving rats during EA stimulation. Corticosterone levels were also
significantly higher in restrained animals after EA stimulation. The new EA paradigm demonstrated in the present study might help the
analysis of certain physiological responses induced by EA that would otherwise have been hindered by restraint stress.
D 2005 Elsevier B.V. All rights reserved.
Theme: Neural basis of behavior
Topic: Drugs of abuse: opioids and others
Keywords: Acupuncture point BL.23; Morphine withdrawal; The amygdala; c-Fos immunohistochemistry; Restraint stress
1. Introduction
Abrupt cessation of chronic opiate use results in a
characterized withdrawal syndrome that includes nausea,
dysphoria, and anxiety [23]. These consequences of
abstinence are thought to be important factors contributing
to opiate addiction. Acupuncture and electroacupuncture
(EA) have been applied with great success to attenuate
behavioral signs of morphine withdrawal in addicts
[8,30,39]. The effects of acupuncture on drug addiction
have also been verified by animal experiments. The with-
0006-8993/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.brainres.2005.02.075
T Corresponding authors. Fax: +86 574 87345976.
E-mail address: [email protected] (G. Yang).
drawal syndrome observed in morphine-dependent rats can
be effectively suppressed by 100-Hz EA [20,47]. Morphine-
induced conditioned place preference can be successfully
suppressed by 2- or 100-Hz EA [38,42]. These animal
studies have provided important information for under-
standing the underlying neurobiological mechanisms of
acupuncture and EA in the treatment of opiate addiction.
However, experimental efforts are, at least partially, ham-
pered by several limitations of laboratory animals. The
most important one is that laboratory animals must be
restrained for several minutes to complete acupuncture
treatment. This has the following inherent problems: (1)
acute restraint stress itself can interfere with acupuncture
stimulation [27]; (2) both a single restraint session and a
repeated restraint stress similarly enhance the effects of
(2005) 155–163

S. Liu et al. / Brain Research 1044 (2005) 155–163156
morphine on locomotor activity [37]. Those rats with high
locomotor activity are more vulnerable to drug addiction
[34]; (3) restraint stress induces the release of endogenous
opioids [3] and produces opioid-like effects [2,32]; (4)
restraint stress can activate several brain regions [10],
representing a major confounding variable for the assess-
ment of EA-induced c-Fos expression in brain areas.
In animal studies, the acupuncture points most frequently
selected are Zusanli (ST.36) and Sanyinjiao (SP.6) located
on the leg. It is difficult to perform the needle manipulation
at these acupuncture points, which are located on the four
limbs, in unrestrained animals. In fact, according to the
theory of traditional Chinese medicine, some acupuncture
points represent discrete locations on the body, where
manual or electrical stimulation can exhibit similar, if not
identical, therapeutic effects [4]. This raises the possibility
that some acupuncture points located on the back and head
have similar therapeutic effects as Zusanli (ST.36) and
Sanyinjiao (SP.6). An important acupuncture point, Shen-
Shu (BL.23), is located on the back and is commonly used
for analgesia and sedation in our clinic. It has been shown
previously that EA at BL.23 has similar antinociceptive
effects as EA at ST.36 [4,17]. We hypothesized that some
acupuncture points that induce analgesia could play a role in
the treatment of opioid addiction.
The goals of our present study were to evaluate the effect
of EA at BL.23 on morphine withdrawal syndrome in both
freely moving and restrained rats. We compared the
behavioral responses during EA stimulation. To determine
whether EA exhibits different activation within certain brain
regions in restrained or unrestrained rats, we initially applied
c-Fos immunomapping to investigate functional activation
of the amygdala during precipitated withdrawal from
morphine and EA stimulation. The amygdala is thought to
be important in mediating the behavioral, autonomic, and
endocrine responses to stressors [11,12,22]. Functional
neuroimaging studies demonstrate that the human amygdala
is activated during negative affective states [11]. Further-
more, the amygdala is well known to be involved in opioid
withdrawal [23,28]. A number of studies have shown that
both morphine withdrawal and restraint stress induce the
activation of c-Fos in the amygdala [5,16,18,35,41]. By
excluding or minimizing interference from restraint stress,
the present study assessed the brealQ effects of EA stimu-
lation on c-Fos expression of the amygdala in morphine
withdrawal rats.
2. Materials and methods
2.1. Subjects
Male Sprague–Dawley rats (250–300 g) from the Zhe-
jiang Center of Experimental Animals were used. They were
randomly assigned and housed collectively (four per cage)
under controlled environmental conditions (22 8C, 12-h
light/dark cycle) with free access to food and water. All
animal treatments were performed in strict accordance with
the National Institutes of Health Guide for the Care and Use
of Laboratory Animals. All experiments were conducted
during the light cycle.
2.2. Drug and withdrawal behavioral scores
Morphine treatment and precipitation of withdrawal were
performed according to published procedures [48]. Rats
were subcutaneously injected with morphine–HCI twice
daily. The dose of each injection was 10 mg/kg in the first
day and increased by 10 mg/kg each day thereafter (10, 20,
30, 40, 50 mg/kg). On day 6, morphine withdrawal
syndrome was precipitated by an intraperitoneal (ip)
injection of 4 mg/kg naloxone hydrochloride (Sigma,
USA), 4 h after one single injection of 50 mg/kg morphine.
The withdrawal behavioral scores were calculated during
the following 45 min (divided into three 15-min periods for
scoring) in a quiet room by one observer who did not know
what experimental treatment had been administered. The
scored signs are based on published criteria [26,36,48], with
minor modifications. The scores obtained for each 15-min
period were noted on the score sheet and were summed over
the 45-min observation period. Withdrawal scores included
teeth chattering, wet dog shake, diarrhea, irritability,
salivation, and abnormal posture including writhing, dig-
ging, and hunching. The amount of weight loss experienced
by the animal during the observation period was also
analyzed. Weight loss during the 45-min observation period
was scored as 1 for a loss of b2%, 5 for a loss of b4%, 10
for a loss of b6%, 15 for a loss of b8%, and 20 for a loss
of N8%.
2.3. EA stimulation
Electric stimulation was conducted for 30 min before
naloxone-precipitated withdrawal. Stainless steel needles
were bilaterally inserted to a depth of 5 mm into BL.23 (one
to two rib’s width lateral to the caudal border of the spinous
process of the second lumbar vertebra) or ST.36 (located
near the knee joint, between the muscle anterior tibialis and
muscle extensor digitorum longus). Constant current square-
wave electric stimulation produced by an electroacupunc-
ture apparatus (Model G-6805-2, Shanghai Medical Elec-
tronic Apparatus, China) was administered via the two
needles. The frequency of stimulation used was 100 Hz
(0.2 ms pulse width). The intensity of the stimulation was
increased stepwise from 1.5 to 2 mA, with each step lasting
for 15 min.
2.4. c-Fos immunohistochemistry
Rats were deeply anesthetized with sodium pentobarbital
(60 mg/kg, ip) and killed by transcardial perfusion of
200 ml saline followed by 200 ml 4% paraformaldehyde in

Fig. 1. Photograph of the electroacupuncture procedures and placement of
acupuncture needles. Two needles were fixed bilaterally at acupuncture
point BL.23. The needles were attached to an electrical stimulator for EA
treatment. The rat could move freely in an observation chamber during EA
stimulation.
S. Liu et al. / Brain Research 1044 (2005) 155–163 157
0.1 mol/L phosphate buffer (PB). Brains were dissected and
postfixed in the same fixative and then stored in 30%
sucrose at 4 8C for 3–5 days. Coronal sections (30 Am in
thickness; �2.56 mm from bregma according to the atlas of
Paxinos and Watson [33]) were cut on the cryostat at
�25 8C. Sections were rinsed in 0.01 M phosphate-buffered
saline (PBS) and incubated in PBS containing 5% normal
goat serum and 0.3% Triton X-100 for 30 min, and then in
Fos antibody (rabbit polyclonal antibody. Santa Cruz, USA)
diluted at 1:200 in PBS at 4 8C for 48 h. After rinsing three
times with PBS, sections were incubated in the biotinylated
goat anti-rabbit secondary antibody (Sigma, USA, diluted
1:200 with PBS) for 2 h and washed again. Then all
corresponding sections were placed in the avidin-biotin-
peroxidase complex solution for 60 min. Finally, DAB
was used for visualization of Fos immunoreactivity. The
reaction was stopped by several PBS washes. Sections were
then mounted on gelatin-coated slides, air-dried, dehydrated
through graded alcohols, cleared in xylene, and cover-
slipped with Eukitt.
2.5. Quantification of Fos-positive nuclei in the amygdala
The amygdala is divided into a number of subregions.
Our study focused on two subregions: (1) the basolateral
amygdala (BLA)—this region is comprised of principal
output neurons that are glutamatergic, pyramidal-shaped
neurons and provide excitatory output to a number of rostral
brain regions; (2) the central nucleus (CeA)—this region
contains primarily GABAergic neurons that project to
brainstem structures, including the locus caeruleus and
hypothalamus. The two subregions have been proposed to
play an important role in the expression of many withdrawal
signs [23,28]. Sections were scanned using an Olympus
BX51 microscope. Image analysis was carried out with the
aid of an image analysis system (Microimage, Olympus
Optical (Europa), Hamburg, Germany. Three consecutive
sections were taken from each animal, and the Fos-positive
nuclei were counted bilaterally, based on a randomization
procedure. A computer-generated rectangle (250 � 600 Am)
was placed in a fixed area of the BLA and CeA of each
section, and the analysis software counted stained nuclei
within the area (see Fig. 5F for specific area used for CeA
and BLA).
2.6. Experimental protocols
In experiment 1, to determine whether BL.23 has similar
therapeutic effects as ST.36 on naloxone-precipitated with-
drawal signs, rats were restrained and submitted to EA
bilaterally at BL.23 or ST36 30 min before naloxone-
precipitated withdrawal on day 6. Rats merely restrained
and not administered EA, and rats that received no treatment
before morphine withdrawal were used as controls.
In experiment 2, to observe the effect of EA at BL.23 on
morphine withdrawal and c-Fos expression in the amygdala
in restrained or freely moving rats, rats were divided into
four groups (n = 7): (1) freely moving and EA group: on day
5, rats were anesthetized with light halothane. Two stainless-
steel needles were inserted at the acupuncture point BL.23
on both sides of the spine and fixed in place with
cyanoacrylate glue and acrylic dental cement. On day 6,
30 min before naloxone-precipitated withdrawal, the nee-
dles were connected with the output terminals of the
stimulator. Freely moving rats received electric stimulation
in an observation chamber (21 � 28 � 20.5 cm) as shown in
Fig. 1. (2) Sham-EA group: the needles were fixed at BL.23
on day 5, but electric stimulation was not conducted on day
6. (3) Restrained and EA group: rats were restrained and
submitted to EA at BL.23 on day 6. (4) The morphine
withdrawal group: EA was not administered before nalox-
one-precipitated morphine withdrawal. Considering that it
was difficult to perform the needle manipulation at ST.36 in
freely moving rats, we did not run the group restrained and
ST.36 as control in experiment 2.
After calculating withdrawal scores, four rats from each
group were randomly selected for c-Fos immunohistoche-
mistry. Four additional morphine-dependent rats (that
received an isotonic saline injection but not naloxone on
day 6) were used for studying the expression of c-Fos in the
amygdala only.
In experiment 3, we observed different behavioral res-
ponses in restrained rats and in freely moving rats during
EA stimulation. In order to exclude the possible effects of
morphine dependence on the behavioral observation, we
used 12 opiate-naive rats. Six rats were restrained and
submitted to EA at BL.23, while six freely moving rats
received EA as described in experiment 2. Vocalization and
facial expression were measured during EA stimulation.
Both vocalization and facial expression in which the corners
of the mouth are retracted, resulting in exposure of the teeth,
were recorded on videotape. A standard scoring system was
used to analyze the presence or absence of vocalization or
the facial expression during EA stimulation. After EA
stimulation, animals were decapitated rapidly. Trunk blood
samples (500 Al in heparinized tubes) were collected and
S. Liu et al. / Brain Research 1044 (2005) 155–163158
centrifuged for 5 min at 4000 rpm. The plasma was
immediately frozen on dry ice and kept frozen at �20 8Cuntil measurement of corticosterone. A radioimmunoassay
for plasma corticosterone was conducted with the help of
Dr. Sun using a kit from ICN Biochemicals (Costa Mesa,
CA) and 125I-corticosterone as the tracer. An additional
group of six opiate-naive rats without any treatment was
used as a control for corticosterone response.
2.7. Statistical analysis
Data from experiments 1 and 2 were analyzed by one-
way or two-way analysis of variance (ANOVA). When
significance was found using ANOVA procedures, Fisher’s
LSD post hoc testing was used to compare individual
treatment groups. Values are expressed as mean F SEM. In
experiment 3, similar statistics were performed on the
corticosterone levels. Fisher’s exact test was used to analyze
the presence and absence of vocalization and facial ex-
pression during EA stimulation. P b 0.05 was considered
statistically significant.
3. Results
3.1. Effects of EA at BL.23 acupuncture point on
naloxone-precipitated withdrawal syndrome
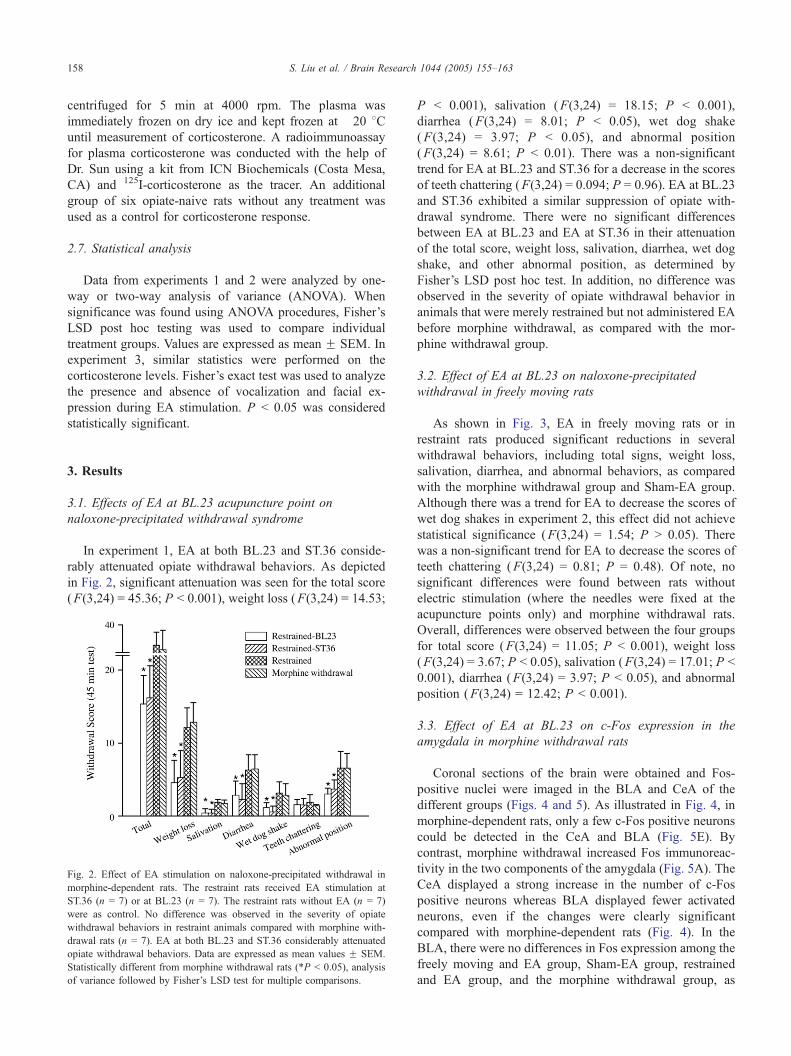
In experiment 1, EA at both BL.23 and ST.36 conside-
rably attenuated opiate withdrawal behaviors. As depicted
in Fig. 2, significant attenuation was seen for the total score
(F(3,24) = 45.36; P b 0.001), weight loss (F(3,24) = 14.53;
Fig. 2. Effect of EA stimulation on naloxone-precipitated withdrawal in
morphine-dependent rats. The restraint rats received EA stimulation at
ST.36 (n = 7) or at BL.23 (n = 7). The restraint rats without EA (n = 7)
were as control. No difference was observed in the severity of opiate
withdrawal behaviors in restraint animals compared with morphine with-
drawal rats (n = 7). EA at both BL.23 and ST.36 considerably attenuated
opiate withdrawal behaviors. Data are expressed as mean values F SEM.
Statistically different from morphine withdrawal rats (*P b 0.05), analysis
of variance followed by Fisher’s LSD test for multiple comparisons.
P b 0.001), salivation (F(3,24) = 18.15; P b 0.001),
diarrhea (F(3,24) = 8.01; P b 0.05), wet dog shake
(F(3,24) = 3.97; P b 0.05), and abnormal position
(F(3,24) = 8.61; P b 0.01). There was a non-significant
trend for EA at BL.23 and ST.36 for a decrease in the scores
of teeth chattering (F(3,24) = 0.094; P = 0.96). EA at BL.23
and ST.36 exhibited a similar suppression of opiate with-
drawal syndrome. There were no significant differences
between EA at BL.23 and EA at ST.36 in their attenuation
of the total score, weight loss, salivation, diarrhea, wet dog
shake, and other abnormal position, as determined by
Fisher’s LSD post hoc test. In addition, no difference was
observed in the severity of opiate withdrawal behavior in
animals that were merely restrained but not administered EA
before morphine withdrawal, as compared with the mor-
phine withdrawal group.
3.2. Effect of EA at BL.23 on naloxone-precipitated
withdrawal in freely moving rats
As shown in Fig. 3, EA in freely moving rats or in
restraint rats produced significant reductions in several
withdrawal behaviors, including total signs, weight loss,
salivation, diarrhea, and abnormal behaviors, as compared
with the morphine withdrawal group and Sham-EA group.
Although there was a trend for EA to decrease the scores of
wet dog shakes in experiment 2, this effect did not achieve
statistical significance (F(3,24) = 1.54; P N 0.05). There
was a non-significant trend for EA to decrease the scores of
teeth chattering (F(3,24) = 0.81; P = 0.48). Of note, no
significant differences were found between rats without
electric stimulation (where the needles were fixed at the
acupuncture points only) and morphine withdrawal rats.
Overall, differences were observed between the four groups
for total score (F(3,24) = 11.05; P b 0.001), weight loss
(F(3,24) = 3.67; P b 0.05), salivation (F(3,24) = 17.01; P b
0.001), diarrhea (F(3,24) = 3.97; P b 0.05), and abnormal
position (F(3,24) = 12.42; P b 0.001).
3.3. Effect of EA at BL.23 on c-Fos expression in the
amygdala in morphine withdrawal rats
Coronal sections of the brain were obtained and Fos-
positive nuclei were imaged in the BLA and CeA of the
different groups (Figs. 4 and 5). As illustrated in Fig. 4, in
morphine-dependent rats, only a few c-Fos positive neurons
could be detected in the CeA and BLA (Fig. 5E). By
contrast, morphine withdrawal increased Fos immunoreac-
tivity in the two components of the amygdala (Fig. 5A). The
CeA displayed a strong increase in the number of c-Fos
positive neurons whereas BLA displayed fewer activated
neurons, even if the changes were clearly significant
compared with morphine-dependent rats (Fig. 4). In the
BLA, there were no differences in Fos expression among the
freely moving and EA group, Sham-EA group, restrained
and EA group, and the morphine withdrawal group, as
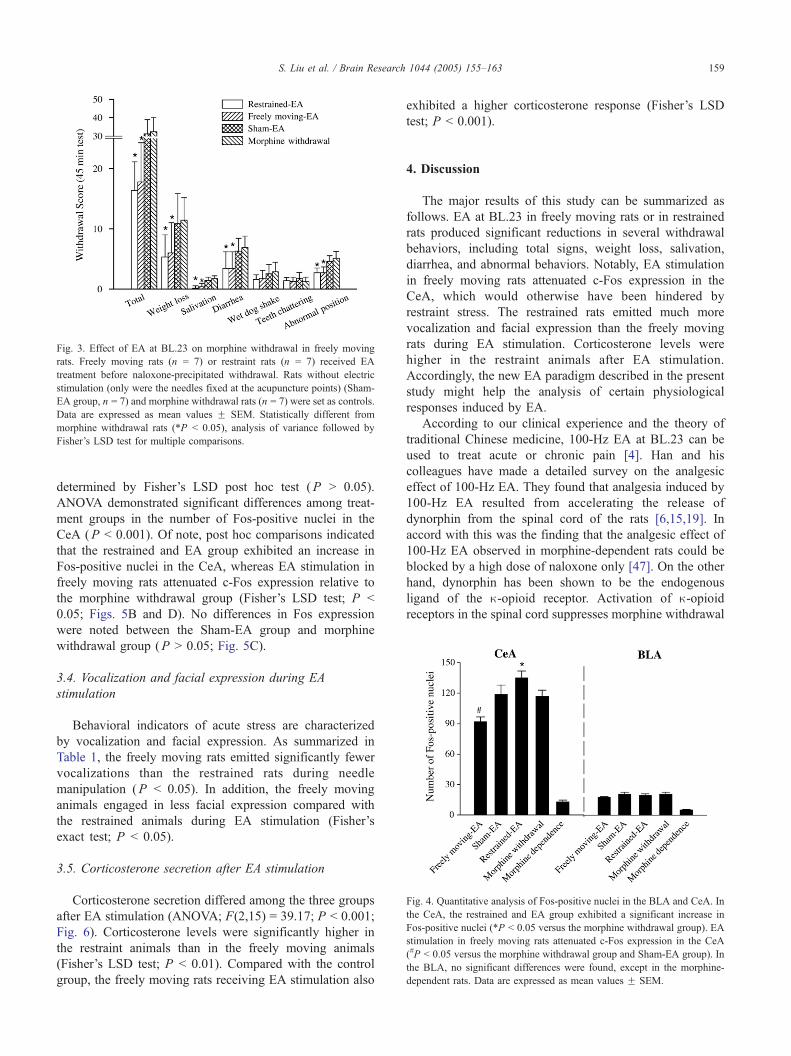
Fig. 4. Quantitative analysis of Fos-positive nuclei in the BLA and CeA. In
the CeA, the restrained and EA group exhibited a significant increase in
Fos-positive nuclei (*P b 0.05 versus the morphine withdrawal group). EA
stimulation in freely moving rats attenuated c-Fos expression in the CeA
(#P b 0.05 versus the morphine withdrawal group and Sham-EA group). In
the BLA, no significant differences were found, except in the morphine-
dependent rats. Data are expressed as mean values F SEM.
Fig. 3. Effect of EA at BL.23 on morphine withdrawal in freely moving
rats. Freely moving rats (n = 7) or restraint rats (n = 7) received EA
treatment before naloxone-precipitated withdrawal. Rats without electric
stimulation (only were the needles fixed at the acupuncture points) (Sham-
EA group, n = 7) and morphine withdrawal rats (n = 7) were set as controls.
Data are expressed as mean values F SEM. Statistically different from
morphine withdrawal rats (*P b 0.05), analysis of variance followed by
Fisher’s LSD test for multiple comparisons.
S. Liu et al. / Brain Research 1044 (2005) 155–163 159
determined by Fisher’s LSD post hoc test (P N 0.05).
ANOVA demonstrated significant differences among treat-
ment groups in the number of Fos-positive nuclei in the
CeA (P b 0.001). Of note, post hoc comparisons indicated
that the restrained and EA group exhibited an increase in
Fos-positive nuclei in the CeA, whereas EA stimulation in
freely moving rats attenuated c-Fos expression relative to
the morphine withdrawal group (Fisher’s LSD test; P b
0.05; Figs. 5B and D). No differences in Fos expression
were noted between the Sham-EA group and morphine
withdrawal group (P N 0.05; Fig. 5C).
3.4. Vocalization and facial expression during EA
stimulation
Behavioral indicators of acute stress are characterized
by vocalization and facial expression. As summarized in
Table 1, the freely moving rats emitted significantly fewer
vocalizations than the restrained rats during needle
manipulation (P b 0.05). In addition, the freely moving
animals engaged in less facial expression compared with
the restrained animals during EA stimulation (Fisher’s
exact test; P b 0.05).
3.5. Corticosterone secretion after EA stimulation
Corticosterone secretion differed among the three groups
after EA stimulation (ANOVA; F(2,15) = 39.17; P b 0.001;
Fig. 6). Corticosterone levels were significantly higher in
the restraint animals than in the freely moving animals
(Fisher’s LSD test; P b 0.01). Compared with the control
group, the freely moving rats receiving EA stimulation also
exhibited a higher corticosterone response (Fisher’s LSD
test; P b 0.001).
4. Discussion
The major results of this study can be summarized as
follows. EA at BL.23 in freely moving rats or in restrained
rats produced significant reductions in several withdrawal
behaviors, including total signs, weight loss, salivation,
diarrhea, and abnormal behaviors. Notably, EA stimulation
in freely moving rats attenuated c-Fos expression in the
CeA, which would otherwise have been hindered by
restraint stress. The restrained rats emitted much more
vocalization and facial expression than the freely moving
rats during EA stimulation. Corticosterone levels were
higher in the restraint animals after EA stimulation.
Accordingly, the new EA paradigm described in the present
study might help the analysis of certain physiological
responses induced by EA.
According to our clinical experience and the theory of
traditional Chinese medicine, 100-Hz EA at BL.23 can be
used to treat acute or chronic pain [4]. Han and his
colleagues have made a detailed survey on the analgesic
effect of 100-Hz EA. They found that analgesia induced by
100-Hz EA resulted from accelerating the release of
dynorphin from the spinal cord of the rats [6,15,19]. In
accord with this was the finding that the analgesic effect of
100-Hz EA observed in morphine-dependent rats could be
blocked by a high dose of naloxone only [47]. On the other
hand, dynorphin has been shown to be the endogenous
ligand of the n-opioid receptor. Activation of n-opioidreceptors in the spinal cord suppresses morphine withdrawal
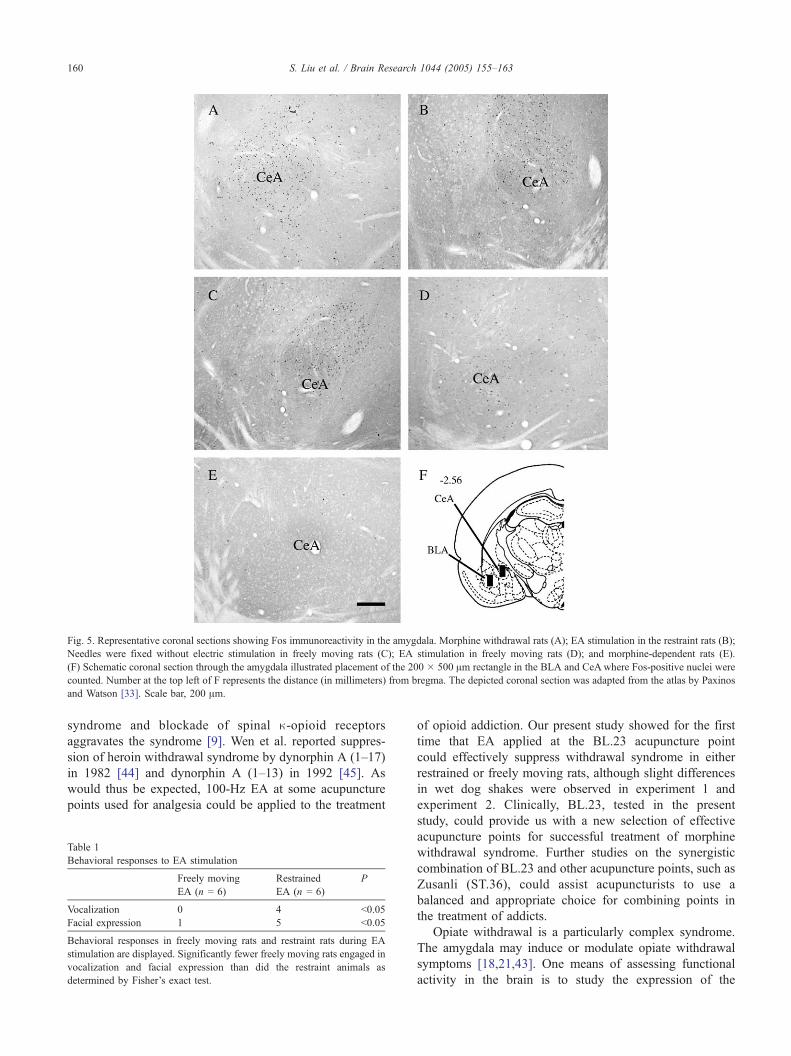
Fig. 5. Representative coronal sections showing Fos immunoreactivity in the amygdala. Morphine withdrawal rats (A); EA stimulation in the restraint rats (B);
Needles were fixed without electric stimulation in freely moving rats (C); EA stimulation in freely moving rats (D); and morphine-dependent rats (E).
(F) Schematic coronal section through the amygdala illustrated placement of the 200 � 500 Am rectangle in the BLA and CeAwhere Fos-positive nuclei were
counted. Number at the top left of F represents the distance (in millimeters) from bregma. The depicted coronal section was adapted from the atlas by Paxinos
and Watson [33]. Scale bar, 200 Am.
S. Liu et al. / Brain Research 1044 (2005) 155–163160
syndrome and blockade of spinal n-opioid receptors
aggravates the syndrome [9]. Wen et al. reported suppres-
sion of heroin withdrawal syndrome by dynorphin A (1–17)
in 1982 [44] and dynorphin A (1–13) in 1992 [45]. As
would thus be expected, 100-Hz EA at some acupuncture
points used for analgesia could be applied to the treatment
Table 1
Behavioral responses to EA stimulation
Freely moving
EA (n = 6)
Restrained
EA (n = 6)
P
Vocalization 0 4 b0.05
Facial expression 1 5 b0.05
Behavioral responses in freely moving rats and restraint rats during EA
stimulation are displayed. Significantly fewer freely moving rats engaged in
vocalization and facial expression than did the restraint animals as
determined by Fisher’s exact test.
of opioid addiction. Our present study showed for the first
time that EA applied at the BL.23 acupuncture point
could effectively suppress withdrawal syndrome in either
restrained or freely moving rats, although slight differences
in wet dog shakes were observed in experiment 1 and
experiment 2. Clinically, BL.23, tested in the present
study, could provide us with a new selection of effective
acupuncture points for successful treatment of morphine
withdrawal syndrome. Further studies on the synergistic
combination of BL.23 and other acupuncture points, such as
Zusanli (ST.36), could assist acupuncturists to use a
balanced and appropriate choice for combining points in
the treatment of addicts.
Opiate withdrawal is a particularly complex syndrome.
The amygdala may induce or modulate opiate withdrawal
symptoms [18,21,43]. One means of assessing functional
activity in the brain is to study the expression of the
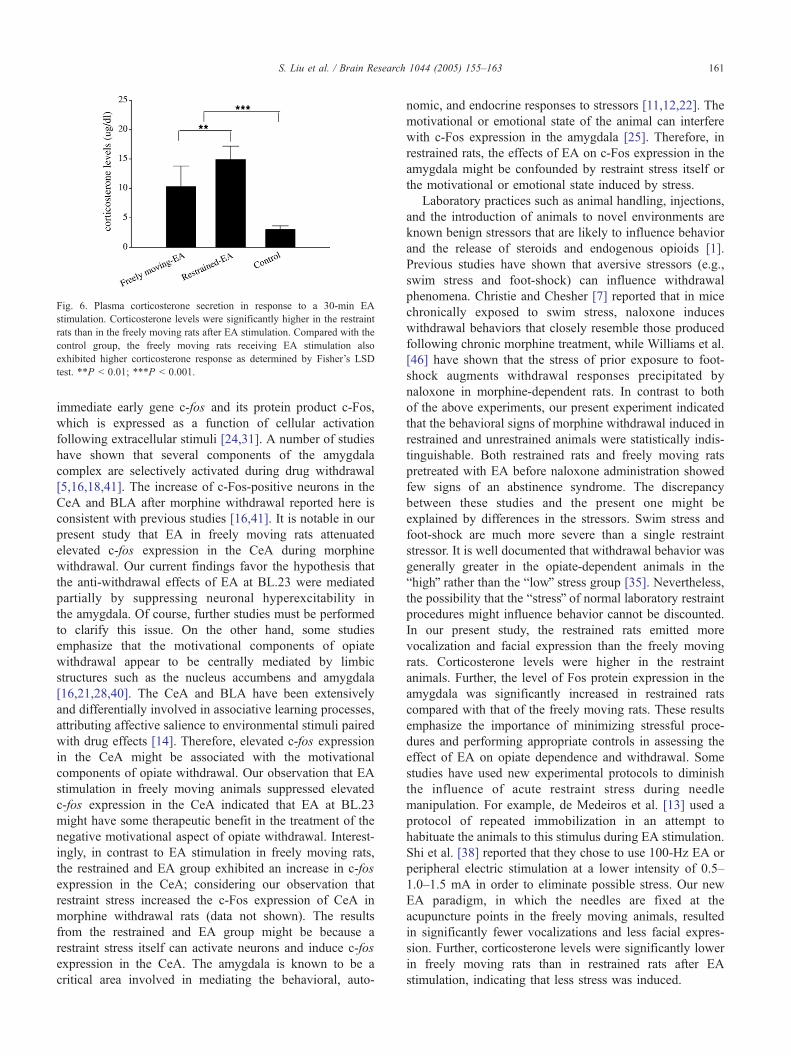
Fig. 6. Plasma corticosterone secretion in response to a 30-min EA
stimulation. Corticosterone levels were significantly higher in the restraint
rats than in the freely moving rats after EA stimulation. Compared with the
control group, the freely moving rats receiving EA stimulation also
exhibited higher corticosterone response as determined by Fisher’s LSD
test. **P b 0.01; ***P b 0.001.
S. Liu et al. / Brain Research 1044 (2005) 155–163 161
immediate early gene c-fos and its protein product c-Fos,
which is expressed as a function of cellular activation
following extracellular stimuli [24,31]. A number of studies
have shown that several components of the amygdala
complex are selectively activated during drug withdrawal
[5,16,18,41]. The increase of c-Fos-positive neurons in the
CeA and BLA after morphine withdrawal reported here is
consistent with previous studies [16,41]. It is notable in our
present study that EA in freely moving rats attenuated
elevated c-fos expression in the CeA during morphine
withdrawal. Our current findings favor the hypothesis that
the anti-withdrawal effects of EA at BL.23 were mediated
partially by suppressing neuronal hyperexcitability in
the amygdala. Of course, further studies must be performed
to clarify this issue. On the other hand, some studies
emphasize that the motivational components of opiate
withdrawal appear to be centrally mediated by limbic
structures such as the nucleus accumbens and amygdala
[16,21,28,40]. The CeA and BLA have been extensively
and differentially involved in associative learning processes,
attributing affective salience to environmental stimuli paired
with drug effects [14]. Therefore, elevated c-fos expression
in the CeA might be associated with the motivational
components of opiate withdrawal. Our observation that EA
stimulation in freely moving animals suppressed elevated
c-fos expression in the CeA indicated that EA at BL.23
might have some therapeutic benefit in the treatment of the
negative motivational aspect of opiate withdrawal. Interest-
ingly, in contrast to EA stimulation in freely moving rats,
the restrained and EA group exhibited an increase in c-fos
expression in the CeA; considering our observation that
restraint stress increased the c-Fos expression of CeA in
morphine withdrawal rats (data not shown). The results
from the restrained and EA group might be because a
restraint stress itself can activate neurons and induce c-fos
expression in the CeA. The amygdala is known to be a
critical area involved in mediating the behavioral, auto-
nomic, and endocrine responses to stressors [11,12,22]. The
motivational or emotional state of the animal can interfere
with c-Fos expression in the amygdala [25]. Therefore, in
restrained rats, the effects of EA on c-Fos expression in the
amygdala might be confounded by restraint stress itself or
the motivational or emotional state induced by stress.
Laboratory practices such as animal handling, injections,
and the introduction of animals to novel environments are
known benign stressors that are likely to influence behavior
and the release of steroids and endogenous opioids [1].
Previous studies have shown that aversive stressors (e.g.,
swim stress and foot-shock) can influence withdrawal
phenomena. Christie and Chesher [7] reported that in mice
chronically exposed to swim stress, naloxone induces
withdrawal behaviors that closely resemble those produced
following chronic morphine treatment, while Williams et al.
[46] have shown that the stress of prior exposure to foot-
shock augments withdrawal responses precipitated by
naloxone in morphine-dependent rats. In contrast to both
of the above experiments, our present experiment indicated
that the behavioral signs of morphine withdrawal induced in
restrained and unrestrained animals were statistically indis-
tinguishable. Both restrained rats and freely moving rats
pretreated with EA before naloxone administration showed
few signs of an abstinence syndrome. The discrepancy
between these studies and the present one might be
explained by differences in the stressors. Swim stress and
foot-shock are much more severe than a single restraint
stressor. It is well documented that withdrawal behavior was
generally greater in the opiate-dependent animals in the
bhighQ rather than the blowQ stress group [35]. Nevertheless,
the possibility that the bstressQ of normal laboratory restraint
procedures might influence behavior cannot be discounted.
In our present study, the restrained rats emitted more
vocalization and facial expression than the freely moving
rats. Corticosterone levels were higher in the restraint
animals. Further, the level of Fos protein expression in the
amygdala was significantly increased in restrained rats
compared with that of the freely moving rats. These results
emphasize the importance of minimizing stressful proce-
dures and performing appropriate controls in assessing the
effect of EA on opiate dependence and withdrawal. Some
studies have used new experimental protocols to diminish
the influence of acute restraint stress during needle
manipulation. For example, de Medeiros et al. [13] used a
protocol of repeated immobilization in an attempt to
habituate the animals to this stimulus during EA stimulation.
Shi et al. [38] reported that they chose to use 100-Hz EA or
peripheral electric stimulation at a lower intensity of 0.5–
1.0–1.5 mA in order to eliminate possible stress. Our new
EA paradigm, in which the needles are fixed at the
acupuncture points in the freely moving animals, resulted
in significantly fewer vocalizations and less facial expres-
sion. Further, corticosterone levels were significantly lower
in freely moving rats than in restrained rats after EA
stimulation, indicating that less stress was induced.

S. Liu et al. / Brain Research 1044 (2005) 155–163162
With regard to the EA paradigm in the present study, there
are some variable factors that need to be taken into account.
Firstly, in comparison to the classical acupuncture or EA
paradigms in which EA stimulation only last tens of minutes,
our new EA stimulation paradigm in freely moving rats
seemed to show a long-term effect of acupuncture stimulation
(the needles were inserted and fixed at the acupuncture points
1 day before naloxone-precipitated morphine withdrawal).
We cannot completely exclude the possibility that an
inflammatory reaction was induced in the local insertion of
needle. However, the acupuncture sites had been treated first
with antiseptic solution to avoid contamination of the needle
manipulation, and we did not observe signs of pain in the rats
(such as reluctance to move about, eat or drink, or vocal-
ization), so we used control group Sham-EA. Interestingly,
without electric stimulation, acupuncture only (where the
needles were fixed at the acupuncture points) has no
significant effect on morphine withdrawal. These data
support the clinical and experimental observation that a
therapeutic effect of acupuncture is achieved by sufficient
stimulation intensity. Clinically, acupuncturists think highly
of the typical needling sensation. This sensation (the
characteristic needling sensation at the acupuncture points,
featured by numbness, heaviness, distention, and soreness) is
traditionally believed to be essential in achieving acupunc-
ture’s therapeutic effect. It is elicited by sufficient manual
manipulation (e.g., rotation, up-and-down motion) of the
inserted acupuncture needle. EA stimulation is widely used as
a substitute for classical needle manipulation. In agreement
with previous studies [29,38], results obtained here also
indicate that sufficient electric stimulationmight be necessary
to achieve EA’s therapeutic effectiveness. Thus, our EA
paradigm may still bear significance in mimicking the short-
term acupuncture stimulation. Secondly, it is still controver-
sial as to how to set suitable control stimulation for
acupuncture or EA. The perfect control, of course, is one in
which needles are inserted into non-acupuncture point areas
but with electric stimulation. Unfortunately, such a control is
impractical in rats. It is difficult to select suitable non-
acupuncture point areas on a rat’s back, in which so many
acupuncture points are located. In the present study, we used
Sham-EA (the needles were fixed at the same acupuncture
point in freely moving rats but without electric stimulation). It
is similar to the real EA and could be helpful for explaining
the results (only one stimulation parameter was changed). In
summary, the new EA paradigm described in the present
study might help the analysis of certain physiological res-
ponses induced by EA that would otherwise have been
hindered by restraint stress.
Acknowledgments
We thank F.Q. Zhang for helpful comments on the
manuscript and technical assistance. This work was
supported by National Basic Research program of China
(2003 CB515404) and National Nature Science Foundation
of China (30100051).
References
[1] H. Akil, J. Madden, R.L. Patrick, J.D. Barchas, Stress-induced
increase in endogenous opiate peptides: concurrent analgesia and its
partial reversal by naloxone, in: H.W. Kosterlitz (Ed.), Opiates and
Endogenous Opioid Peptides. Amsterdam; Elsvier North-Holland
Biomedical Press, 1982, pp. 63–70.
[2] S. Amir, Z. Amit, Endogenous opioid ligands may mediate stress-
induced changes in the affective properties of pain related behaviour
in rats, Life Sci. 23 (1978) 1143–1152.
[3] B.D. Appelbaum, S.G. Holtzman, Characterization of stress-induced
potentiation of opioid effects in the rat, J. Pharmacol. Exp. Ther. 231
(1984) 555–565.
[4] Beijing College of Traditional Chinese Medicine, Shanghai College
of Traditional Chinese Medicine, Nanjing College of Traditional
Chinese Medicine, Acupuncture Institute of the Academy of
Traditional Chinese Medicine, Essentials of Chinese acupuncture,
Foreign Language Press, Beijing, People’s Republic of China,
1980.
[5] B. Calvino, J. Lagowska, Y. Ben-Ari, Morphine withdrawal syn-
drome: differential participation of structures located within the
amygdaloid complex and striatum of the rat, Brain Res. 177 (1979)
19–34.
[6] X.H. Chen, J.S. Han, Analgesia induced by electroacupuncture of
different frequencies is mediated by different types of opioid
receptors: another cross-tolerance study, Behav. Brain Res. 47
(1992) 143–149.
[7] M.J. Christie, G.B. Chesher, Physical dependence on physiologically
released endogenous opiates, Life Sci. 30 (1982) 1173–1177.
[8] V. Clement-Jones, L. McLoughlin, P.J. Lowry, G.M. Besser, L.H.
Rees, H.L. Wen, Acupuncture in heroin addicts; changes in Met-
enkephalin and beta-endorphin in blood and cerebrospinal fluid,
Lancet 2 (1979) 380–383.
[9] C.L. Cui, L.Z. Wu, J.S. Han, Spinal k-opioid system plays an
important role in suppressing morphine withdrawal syndrome in the
rat, Neurosci. Lett. 295 (2000) 45–48.
[10] W.E. Cullinan, J.P. Herman, D.F. Battaglia, H. Akil, S.J. Watson,
Pattern and time course of immediate early gene expression in rats
brain following acute stress, Neuroscience 64 (1995) 477–505.
[11] R.J. Davidson, W. Irwin, The functional neuroanatomy of emotion and
affective style, Trends Cogn. Sci. 3 (1999) 11–21.
[12] M. Davis, The role of the amygdala in fear and anxiety, Annu. Rev.
Neurosci. 15 (1992) 353–375.
[13] M.A. de Medeiros, N.S. Canteras, D. Suchecki, L.E. Mello,
Analgesia and c-Fos expression in the periaqueductal gray induced
by electroacupuncture at the Zusanli point in rats, Brain Res. 973
(2003) 196–204.
[14] B.J. Everitt, J.A. Parkinson, M.C. Olmstead, M. Arroyo, P. Robledo,
T.W. Robbins, Associative processes in addiction and reward. The role
of amygdala-ventral striatal subsystems, Ann. N. Y. Acad. Sci. 877
(1999) 412–438.
[15] H. Fei, G.X. Xie, J.S. Han, Low and high frequency electro-
acupuncture stimulations release (Met5) enkephalin and dynorphin
A in rat spinal cord, Chin. Sci. Bull. 32 (1987) 1496–1509.
[16] F. Frenois, M. Cador, S. Caille, L. Stinus, C. Le Moine, Neural
correlates of the motivational and somatic components of naloxone-
precipitate morphine withdrawal, Eur. J. Neurosci. 16 (2002)
1377–1389.
[17] M.X. Gao, The effect of thumb-pressure at Shen-Shu on renal colic,
JCAM 17 (2001) 9–10.
[18] K.N. Gracy, L.A. Dankiewicz, G.F. Koob, Opiate withdrawal-induced
fos immunoreactivity in the rat extended amygdala parallels the

S. Liu et al. / Brain Research 1044 (2005) 155–163 163
development of conditioned place aversion, Neuropsychopharmacol-
ogy 24 (2001) 152–160.
[19] J.S. Han, Acupuncture: neuropeptide release produced by electrical
stimulation of different frequencies, Trends Neurosci. 6 (2003)
17–22.
[20] J.S. Han, R.L. Zhang, Suppression of morphine abstinence syndrome
by body electroacupuncture of different frequencies in rats, Drug
Alcohol Depend. 31 (1993) 169–175.
[21] S. Heinrichs, F. Menzhagi, G. Schulteis, G.F. Koob, L. Stinus,
Suppression of corticotrophin-releasing factor in the amygdala
attenuates aversive consequences of morphine withdrawal, Behav.
Pharmacol. 6 (1995) 74–80.
[22] N.H. Kalin, S.E. Shelton, R.J. Davidson, A.E. Kelley, The primate
amygdala mediates acute fear but not the behavioral and physio-
logical components of anxious temperament, J. Neurosci. 21 (2001)
2067–2074.
[23] G.F. Koob, Drug addiction: the yin and yang of hedonic homeostasis,
Neuron 16 (1996) 893–896.
[24] K.J. Kovacs, c-Fos as a transcription factor: a stressful (re) view from
a functional map, Neurochem. Int. 33 (1998) 287–297.
[25] J.E. Le Doux, Emotion circuits in the brain, Annu. Rev. Neurosci. 23
(2000) 155–184.
[26] P.H.K. Lee, R.W. Mcnutt, K.J. Chang, A nonpeptidic delta opioid
receptor agonist, BW373U86, attenuates the development and ex-
pression of morphine abstinence precipitated by naloxone in rat,
J. Pharmacol. Exp. Ther. 267 (1993) 883–887.
[27] S. Levine, H. Ursin, What is stress, in: M.R. Brown, G.F. Koob, C.
Rivier (Eds.), Stress, Neurobiology and Neuroendocrinology, Marcel
Dekker, New York, 1991, pp. 3–21.
[28] R. Maldonado, L. Stinus, L.H. Gold, G.F. Koob, Role of different
brain structures in the expression of the physical morphine withdrawal
syndrome, J. Pharmacol. Exp. Ther. 261 (1992) 669–677.
[29] A. Margolin, D.H. Kleber, S.K. Avants, J. Konefal, F. Gawwin, E.
Starkiff, E. Wells, T.R. Jackson, M. Bullock, P.D. Culliton, S. Boles,
R. Vaughan, Acupuncture for the treatment of cocaine addiction,
J. Am. Med. Assoc. 287 (2002) 55–63.
[30] K. Montazeri, M. Farahnakian, M. Saghaei, The effect of acupuncture
on the acute withdrawal symptoms from rapid opiate detoxification,
Acta Anaesthesiol. Sinics 40 (2002) 173–177.
[31] J.I. Morgan, T. Curran, Immediate-early genes: ten years on, Trends
Neurosci. 18 (1995) 66–67.
[32] G.A. Olson, R.D. Olson, A.J. Kastin, Endogenous opiates: 1989,
Peptides 11 (1990) 1277–1304.
[33] G. Paxinos, C. Watson, The Rat Brain in Stereotaxic Coordinates,
Academic, London, 1986.
[34] P.V. Piazza, S. Maccari, J.M. Deminiere, M. Moal Le, P. Mormede,
H. Simon, Corticosterone levels determine individual vulnerability to
amphetamine self administration, Proc. Natl. Acad. Sci. U. S. A. 88
(1991) 2088–2092.
[35] T.L. Pierce, C. Raper, The effects of laboratory handling
procedures on naloxone precipitated withdrawal behavior in mor-
phine-dependent rats, J. Pharmacol. Toxicol. Methods 34 (1995)
149–155.
[36] L.J. Punch, D.W. Self, E.J. Nestler, J.R. Taylor, Opposite modulation
of opiate withdrawal behaviors on microinfusion of a protein kinase a
inhibitor versus activator in the locus coeruleus of periaqueductal
gray, J. Neurosci. 17 (1997) 8520–8527.
[37] C.N. Rosario, A.M. Pacchioni, L.M. Cancela, Influence of acute or
repeated restraint stress on morphine-induced locomotion: involve-
ment of dopamine, opioid and glutamate receptors, Behav. Brain Res.
134 (2002) 229–238.
[38] X.D. Shi, W. Ren, G.B. Wang, F. Luo, J.S. Han, C.L. Cui, Brain
opioid-receptors are involved in mediating peripheral electric stim-
ulation-induced inhibition of morphine conditioned place preference
in rats, Brain Res. 981 (2003) 23–29.
[39] M. Shwartz, R. Saitz, K. Mulvey, P. Brannigan, The value of
acupuncture detoxification programs in a substance abuse treatment
system, J. Subst. Abuse Treat. 17 (1999) 305–312.
[40] L. Stinus, M. Le Moal, G.F. Koob, Nucleus accumbens and amygdala
are possible substrates for the aversive stimulus effects of opiate
withdrawal, Neuroscience 37 (1990) 767–773.
[41] P. Veinante, M.E. Stoeckel, F. Lasbennes, M.J. Freund-Mercier, C-Fos
and peptide immunoreactivities in the central extended amygdala of
morphine-dependence rats after naloxone-precipitated withdrawal,
Eur. J. Neurosci. 18 (2003) 1295–1305.
[42] B. Wang, F. Luo, Y.Q. Xia, J.S. Han, Peripheral electric stimulation
inhibits morphine-induced place preference in rats, NeuroReport 11
(2000) 1017–1020.
[43] T. Watanabe, R. Yamamoto, A. Maeda, T. Nakagawa, M. Minami,
M. Satoh, Effects of excitotoxic lesions of the central or basolateral
nucleus of the amygdala on naloxone-precipitated withdrawal-induced
conditioned place aversion in morphine-dependent rats, Brain Res.
958 (2002) 423–428.
[44] H.K. Wen, W.K.K. Ho, Suppression of withdrawal syndrome by
dynorphin in heroin addicts, Eur. J. Pharmacol. 82 (1982) 183–186.
[45] H.L. Wen, J.C.K. Kwok, N. Datta, K.Y. Sin, S.K. Teoh, W.K.K. Ho,
Detoxification of heroin abusers by dynorphin (1–13), Pharmacol.
Commun. 1 (1992) 259–265.
[46] J.L. Williams, R.C. Drugan, S.F. Maier, Exposure to uncontrollable
stress alters withdrawal from morphine, Behav. Neurosci. 98 (1984)
836–846.
[47] L.Z. Wu, C.L. Cui, J.B. Tian, D. Ji, J.S. Han, Suppression of morphine
withdrawal by electroacupuncture in rats: dynorphin and kopioid
receptor implicated, Brain Res. 851 (1999) 290–296.
[48] G.D. Yang, W.H. Zhou, F.Q. Zhang, Experimental study of
selective muscarinic receptor antagonists on attenuation of morphine
tolerance and dependence in rats, Nat. Med. J. China 77 (1997)
130–133.





























