Embed Size (px)
Citation preview

ACTA NEUROBIOL. EXP. 1990, 50: 23-36
ELECTRICAL ACTIVITY AND FEEDING CORRELATES OF INTRACRANIAL HYPOTHALAMIC INJECTION OF GABA,
MUSCIMOL AND PICROTOXIN IN THE RATS
Anil Kumar RATTAN and Harjit Kaur MANGAT
Neuroscience Reseamh Laboratmy, Schold of Life Sciences Guru Nanak Dev University, Amritsar-143005, India
Key words: hypothalamus, hunger, thirst, EEG, inhibition, rats
Abstract. Assemblies of electrodes and a cannula were stereota- xically implanted in the ventromedial (VMH), lateral (LHA) and pa- raventricular (PVH) hypothalamic areas in male albino rats. Electrical activity of these regions was recorded electrographically before and following intracranial injection (ICI) of GABA, muscimol and picrotoxin. In another set of animals, food intake and water intake were also mea- sured. The activity of the ventromedial hypothalamus changed from slow to fast after ICI of GABA and picrotoxin and fast to slow after muscimol. The activity of the lateral hypothalamus changed from slow to fast with ICI of muscimol and picrotoxin and from fast to slow with GABA, while that of the paraventricular hypothalamic nucleus changed from slow to fast with ICI GABA and fast to slow with muscimol and picrotoxin. ICI of GABA into VMH and LHA and muximol in VMH, LHA and PVH caused a decrease in food intake. Water intake was also decreased after ICI of GABA in PVH and muscimol in LHA and PVH. On the opposite picrotoxin increased food intake in VMA and LHA and water intake in PVH. The possible interaction of GABAergic drugs with the areas of the brain controlling feeding and drinking is being discussed.
Send mrrespondence to H. K. Mamgat.

INTRODUCTION
The role of central nervous system in the regulation of feeding is now well recognized and sufficient experimental evidence was provided to demonstrate the existence of two opposing mechanisms in the hypo- thalamus which regulate food intake (7, 8, 16, 18).
Several suggestive data with lesion studies have been reported, con- cerning possible roles of VMH, LHA and PVH as satiety, feeding and drinking centres of the brain (1-3). Grandiston and Guidotti (10) reported that microinjection of GABA agonist muscimol into the VMH stimulated food intake in satiated rats. Leibowitz and Hor (13) found that f3-endor- phin caused this effect when injected into the PVH. Chronic injections of morphine into the third cerebral ventricle decreased food intake and water intake in rats (8). The functional states of the feeding and satiety centres in the hypothalamus have also been attributed to GABA inhi- bitory neurons or such synaptic links (12).
Although innumerable number of experiments have been reported in the literature to study the site of drug action, the knowledge of the correlation of the changes in ingestive behavior and EEG activity is scanty. The present study attempts to correlate the food intake with electrical activity of VMH, LHA and PVH after microinjection of the gamma-aminobutyric acid (GABA), to the respective hypothalamic areas.
MATERIAL AND METHODS
Male albino rats (Wistar strain) weighing between 200-250 g, fed with natural pellets and water ad lib. were individually housed in a well ventilated and temperature controlled room (22 + 1°C) and maintained on an artificial 14 : 10 h light-dark cycle. The rats were fed with natural food pellets (Hindustan Levers Ltd., Bombay) and water ad lib.
The animals were divided into a feeding group and an electrical activity group. The rats of the both groups were surgically implanted with intracranial cannula (23 gauge, 17 mm legnth) and bipolar stainless steel electrodes (20 mm length, 0.25 mm diameter, varnish insulated except for the last 0.5 mm of the tip), placed in various hypothalamic nuclei using bregma as landmark, i.e. for VMH (A 5.8, L 0.8, D 9.0), LHA (A 6.6, L 1.5, D 9.5) and PVH (A 6.6, L 0.5, D 8.5) taken from the atlas of DeGroot (6). The animals were anesthetized with Nembu tal (35 mglkg b.wt, i.p.). In each animal, only one hypothalamic nucleus was explored.
The animals of the feeding group were injected intracranially (ICI) with GABA (0.5 pg/0.2 pl) or muscimol (0.5 pg/0.2 pl or picrotoxin

(0.25 y*g/0.2 p1) at a speed of 0.1 pl/min. Immediately after injection, each rat was returned to its home cage and a reweighed food pellet and weighed water bottle were placed in the cage. All significant spillage was collected and included in the calculations of food intake. For water intake, spillage was collected in containers placed directly under water bottles. The quantities of food intake and water intake consumed were measured at 2 h after the start of the experiment. The experiment began at approximately lC.OO a.m. so that the intakes were measured during the light period.
For electrical activity recordings, the rats were placed in a trans- parent plastic box contained in a sound-proof shielded room. The elec- trical activity of VMH, LHA and PVH was bipolarly recorded on a poly- graph (Inco, India) at a time contant of 0.1 s and a pass filter set at 50 Hz. Following one hour record, the rats were given ICI of GABA or muscimol or picrotoxin or saline at a speed of 0.1 pllmin. Dosages were the same as in the feeding group. Injections were never repeated in any one of these rats. Evaluation and quantification of the EEG data made as previously described (17).
After the completion of experimental procedure, electrode sites were marked by passing direct current of 3 mA for 5 s to pmduce irop deposits at the tips of electrodes and cannula. The animals were sacri- ficed and their brains perfused with 10°/o formalin solution containing 1°/o potassium ferrocyanide. This produced a Prussian blue reaction with iron deposits, resulting in blue spots at the sites of electrodes and cannula in 25 pM thick sections of the brain.
The data were evaluated statistically with Students t-test (20).
RESULTS
The effect of GABA, muscimol and picrotoxin oln 2 h food intake and water intake is shown in Table I. GABA and muscimol significantly (P < 0.01) decreased food intake when injected into the VMH and LHA. The two drugs also reduced water intake when injected into the PVH and VMH respectively. On the contrary, picrotoxin increased food intake or water intake after injection on the same studies where GABA caused a decrease.
Electrical activity of the hypothalamus
Marginal potentiation of electrical activity appeared in VMH at about 15 min after ICI of GABA, which persisted throughout the recording period (Figs. 1A and 4; Tables I1 and 111). The electrical activity of
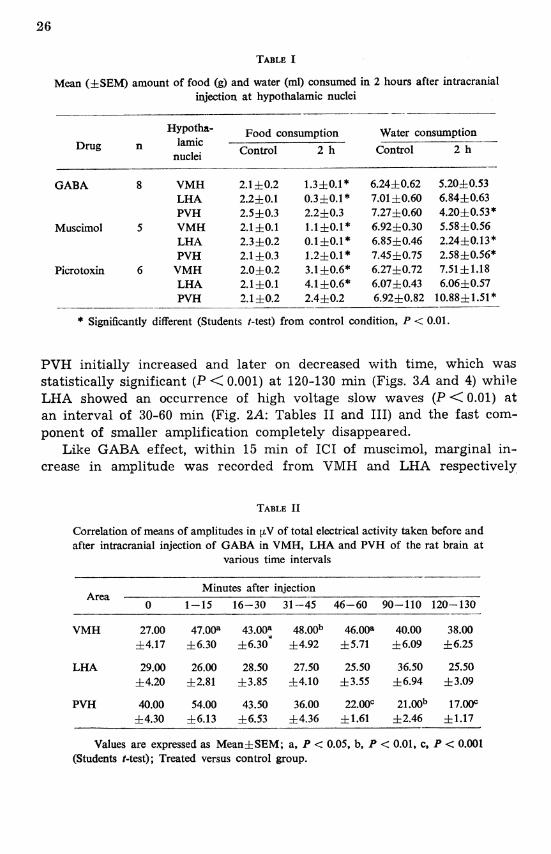
Mean (f SEM) amount of food (g) and water (ml) consumed in 2 hours after intracranial injection at hypothalamic nuclei
Hypotha- ~ o o d consumption Water consumption lamic - .- -
Drug Control 2 h Control nuclei 2 h
GABA
Muscimol
Picrotoxin
8 VMH LHA PVH
5 VMH LHA PVH
6 VMH LHA PVH
* Significantly different (Students I-test) from control condition, P < 0.01.
PVH initially increased and later on decreased with time, which was statistically significant (P < 0.001) at 120-130 min (Figs. 3A and 4) while LHA showed an occurrence of high voltage slow waves (P < 0.01) at an interval of 30-60 min (Fig. 2A: Tables I1 and 111) and the fast corn- ponent of smaller amplification completely disappeared.
Like GABA effect, within 15 min of ICI of rnuscirnol, marginal in- crease in amplitude was recorded from VMH and LHA respectively
Correlation of means of amplitudes in pV of total electrical activity taken before and after intracranial injection of GABA in VMH, LHA and PVH of the rat brain at
various time intervals - -- --
Minutes after injection Area
0 1-15 16-30 31-45 46-60 90-110 120-i%
VMH 27.00 47.00a 43.00: 48.00~ 46.00a 40.00 38.00 f4.17 f6.30 f 6.30 f 4.92 f 5.71 f 6.09 f 6.25
PVH 40.00 54.00 43.50 36.00 22.01)~ 21.00b 17.00~ 44.30 f 6.13 f 6.53 f 4.36 f1.61 52.46 f 1.17
Values are expressed as Meanf SEM; a, P < 0.05, b, P < 0.01, c, P < 0.001 (Students t-test); Treated versus control group.
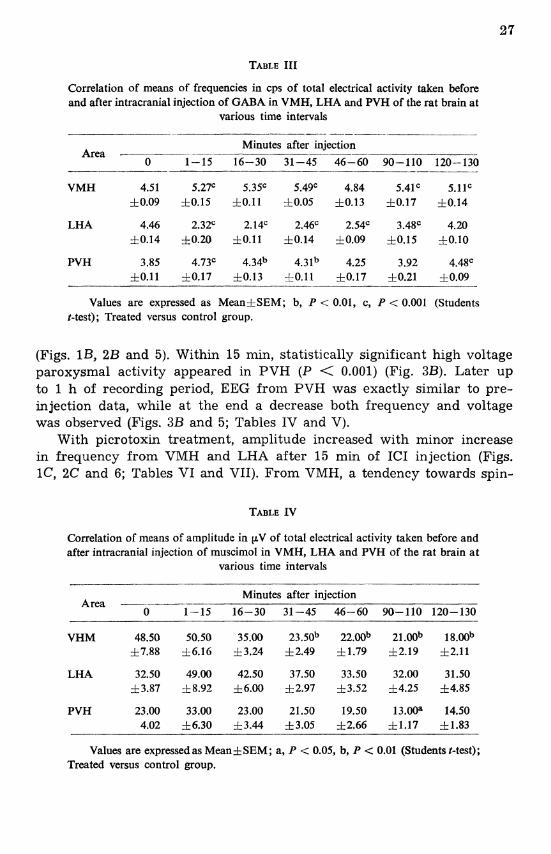
Correlation of means of frequencies in cps of total electrical activity taken before and after intracranial injection of GABA in VMH, LHA and PVH of the rat brain at
various time intervals
Minutes after injection Area -- -- - - 0 1-15 16-30 31-45 46-60 90-110 120-130
. - - - - -- - - - - - -- -- - . - .- --
VMH 4.51 5.27C 5.3F 5.49C 4.84 5.4lc 5.1lc 10.09 zk0.15 hO.11 i0.05 zk0.13 h0.17 f0.14
LHA 4.46 2.32" 2.14C 2.46C 2.54C 3.4Sc 4.20 f0.14 rf10.20 10.11 f0.14 h0.09 f0.15 +0.10
PVH 3.85 4.73' 4.34b 4.31b 4.25 3.92 4.4Sc f 0.11 10.17 f 0.13 10.11 f 0.17 10.21 10.09 - -- - - --- - -- - -
Values are expressed as Meanf SEM; b, P < 0.01, c, P < 0.001 (Students t-test); Treated versus control group.
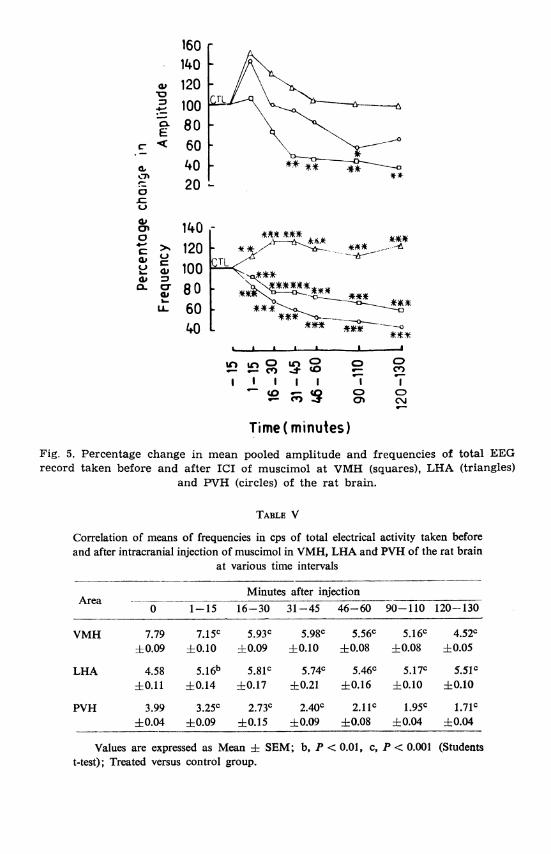
(Figs. lB, 2B and 5). Within 15 min, statistically significant high voltage paroxysmal activity appeared in PVH (P < 0.001) (Fig. 3B). Later up to 1 h of recording period, EEG from PVH was exactly similar to pre- injection data, while at the end a decrease both frequency and voltage was observed (Figs. 3B and 5; Tables IV and V).
With picrotoxin treatment, amplitude increased with minor increase in frequency from VMH and LHA after 15 min of ICI injection (Figs. lC, 2C and 6; Tables VI and VII). From VMH, a tendency towards spin-
Correlation of means of amplitude in p.V of total electrical activity taken before and after intracranial injection of lnuscimol in VMH, LHA and PVH of the rat brain at
various time intervals
Minutes after injection Area - - -- - -- ----- ----- - 0 1-15 16-30 31-45 46-60 90-110 120-130
VHM 48.50 50.50 35.00 23.50~ 22.00b 21.00b 18.OC)b f7.88 f6.16 h3.24 f2.49 11.79 f2.19 f2.11
PVH 23.00 33.00 23.00 21.50 19.50 13.Wa 14.50 4.02 f 6.30 f 3.44 f3.05 12.66 f 1.17 f 1.83
Values are expressedas Mean*SEM; a, P < 0.05, b, P < 0.01 (Students t-test); Treated versus control group.
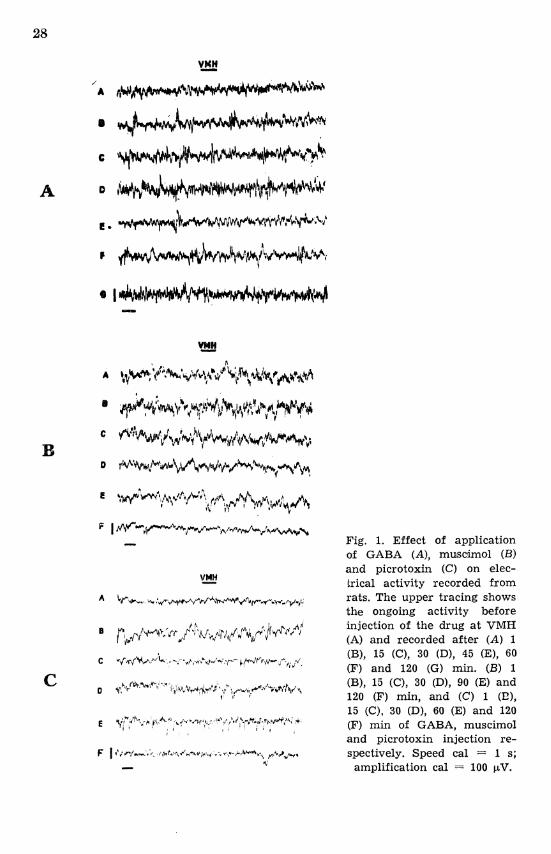
Fig. 1. Effect of application of GABA (A), muscimol (B) and picrotoxin (C) on elec- trical activity recorded from rats. The upper tracing shows the ongoing activity before injection of the drug a t VMH (A) and recorded after (A) 1 (B), 15 (C), 30 (D), 45 (E), 60 (F) and 120 (G) min. (B) 1 (B), 15 (C), 30 (D), 90 (E) and 120 (F) min, and (C) 1 (E), 15 (C), 30 (D), 60 (E) and 120 (F) min of GABA, muscimol and picrotoxin injection re- spectively. Speed cal = 1 s;
amplification cal = 100 pV.
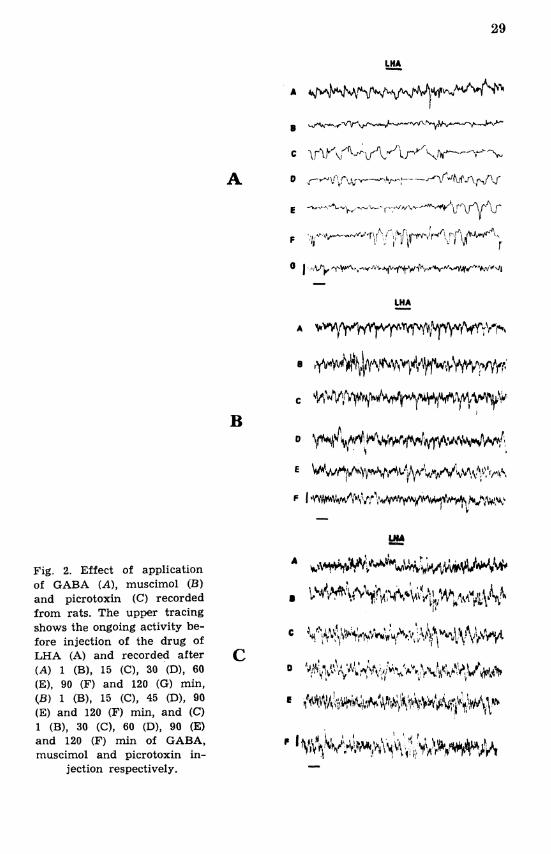
Fig. 2. Effect of application of GABA (A), muscimol (B) and picrotoxin (C) recorded from rats. The upper tracing shows the ongoing activity be- fore injection of the drug of LHA (A) and recorded after (A) 1 (B), 15 (C), 30 (Dl, 60 (E), 90 (F) and 120 (G) min, (B) 1 (B), 15 (C), 45 (Dl, 90 (E) and 120 (F) min, and (C) 1 (B), 30 (C), 60 (Dl, 90 (E) and 120 (F) min of GABA, muscimol and picrotoxin in-
jection respectively.
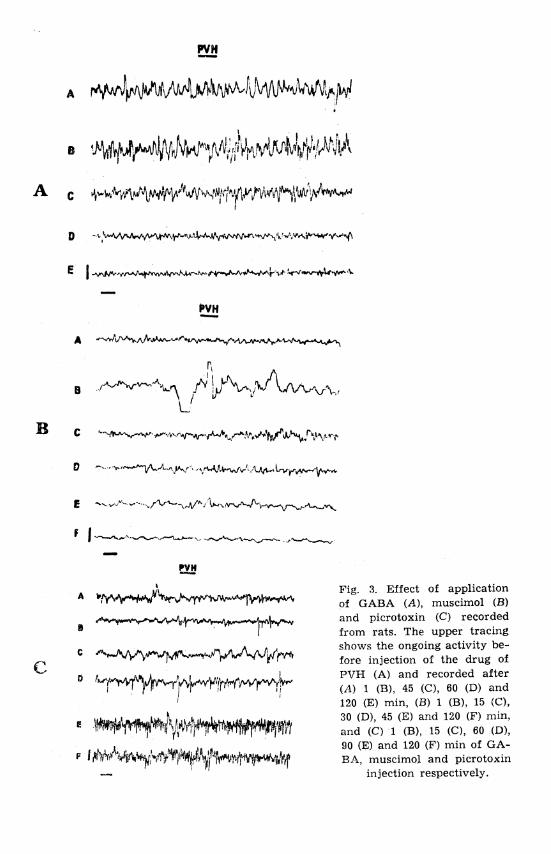
Fig. 3. Effect of application of GABA (A), muscimol (B) and picrotoxin (C) recorded from rats. The upper tracing shows the ongoing activity be- fore injection of the drug of PVH (A) and recorded after (A) 1 (B), 45 (C), 60 (D) and 120 (E) min, (B ) 1 (B), 15 (C), 30 (D), 45 (E) and 120 (F) mln, and (C) 1 (B), 15 (C), 60 (D), 90 (E) and 120 (F) rnin of GA- B,%, muscimol and picrotoxin
injection respectively.
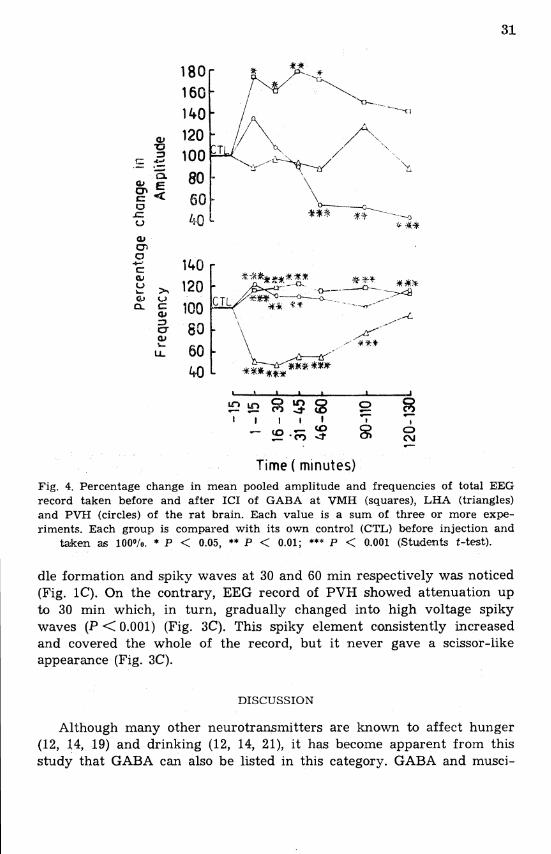
Time ( minutes) Fig. 4. Percentage change in mean pooled amplitude and frequencies of total EEG record taken before and after ICI of GABA at VMH (squares), LHA (triangles) and PVH (circles) of the rat brain. Each value is a sum of three or more expe- riments. Each group is compared with its own control (CTL) before injection and
taken as 100o/o. * P < 0.05, ** P < 0.01; *** P < 0.001 (Students t-test).
dle formation and spiky waves at 30 and 60 min respectively was noticed (Fig. 1C). On the contrary, EEG record of PVH showed attenuation up to 30 min which, in turn, gradually changed into high voltage spiky waves (P < 0.001) (Fig. 3C). This spiky element consistently increased and covered the whole of the record, but it never gave a scissor-like appearance (Fig. 3C).
DISCUSSION
Although many other neurotransmitters are known to affect hunger (12, 14, 19) and drinking (12, 14, 21), it has become apparent from this study that GABA can also be listed in this category. GABA and musci-
% 140 - 0
LA 60 'to
Time (minutes)
Fig. 5. Percentage change in mean pooled amplitude and frequencies of total KEG record taken before and after ICI of muscimol a t VMH (squares), LHA (triangles)
and W H (circles) of the ra t brain.
Correlation of means of frequencies in cps of total electrical activity taken before and after intracranial injection of muscimol in VMH, LHA and PVH of the rat brain
at various time intervals - - -- -- - - - -- -- - -
Minutes after injection Area - - - - - . pp - - - - -- - - - -. --
0 1-15 16-30 31-45 46-60 90-110 120--130
VMH 7.79 7.15' 5.93C 5.98' 5.56C 5.16C 4.52C 1k0.09 10.10 10.09 10.10 f 0.08 f 0.08 1 0.05
PVH 3.99 3.25' 2.73C 2.4Oc 2.1lc 1.9Y 1.7lc f0.04 10.09 f 0.15 &0.09 f 0.08 f 0.04 f 0.04
Values are expressed as Mean f SEM; b. P < 0.01, c, P < 0.001 (Students t-test); Treated versus control group.
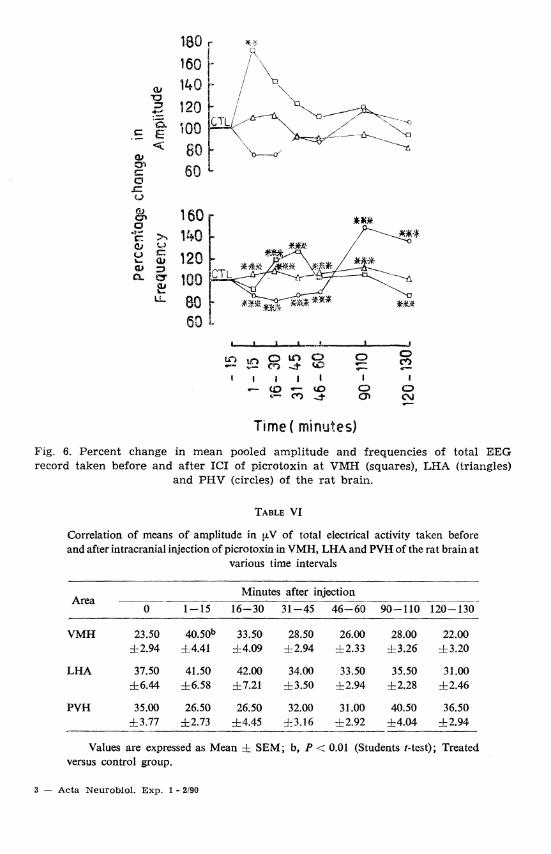
Fig. 6. Percent change in mean pooled amplitude and frequencies of total EEG record taken before and after ICI of picrotoxin a t VMH (squares), LHA (triangles)
and PHV (circles) of the rat brain.
Correlation of means of amplitude in pV of total electrical activity taken before and after intracranial injection of picrotoxin in VMH, LHA and PVH of the rat brain a t
various time intervals
Minutes after injection Area -- - - .- .--
0 1-15 16-30 31-45 46-60 90-110 120-130
VMH 23.50 40.50b 33.50 28.50 26.00 28.00 22.00 i 2 . 94 14.41 14.09 k2.94 rt2.33 f3.26 rt3.20
LHA 37.50 41.50 42.00 34.00 33.50 35.50 31.00 56.44 16.58 17.21 rt3.50 f 2.94 f 2.28 +2.46
PVH 35.00 26.50 26.50 32.00 31.00 40.50 36.50 i 3 . 7 7 12.73 f4.45 13.16 -12.92 &4.04 -t2.94
--- - - - - -- --
Values are expressed as Mean 5 SEM; b, P -< 0.01 (Students t-test); Treated versus control group.
3 - Acta Neurobiol. Exp. 1 - 2190
mol administered into VMH and LHA decreased food intake, which infers that it might have precise inhibitory effect on LHA, whereas VMH, an inhibitory area for feeding responses, remained inhibited in these studies. The imoprtance for lateral hypothalamus (which coincides with the region 6.4 mm anterior (A 6.4)) in appetite controlling mecha- nisms has already been reported (9, 14) and in addition, the loci of the highest concentrations of GABA content of the rat hypothalamus is LHA. Thus the two factors: (i) highest concentration of GABA in LHA, and (ii) role of LHA in appetite when correlated with observations of ICI GABA in LHA and VMH, strongly suggest that the existence of this neurotransmitter has a precise function in appetitive mechanisms. Fur-
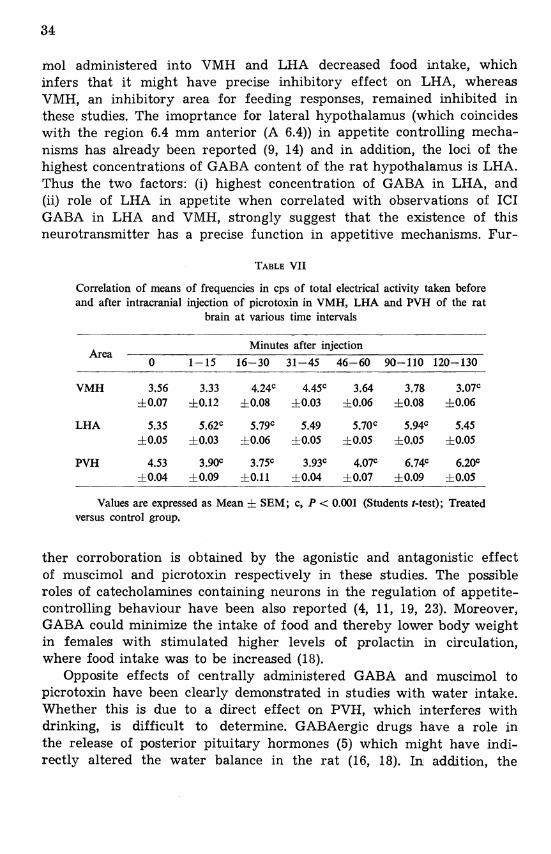
TABLE VII
Correlation of means of frequencies in cps of total electrical activity taken before and after intracranial injection of picrotoxin in VMH, LHA and PVH of the rat
brain at various time intervals -- -
Minutes after injection Area ---
0 1-15 16-30 31-45 46-60 90-110 120-130
VMH 3.56 3.33 4.24C 4.45' 3.64 3.78 3.07C f 0.07 50.12 h0.08 50.03 10.06 rf 0.08 rf0.06
LHA 5.35 5.62C 5.79' 5.49 5.70~ 5.94~ 5.45 &0.05 f0.03 f0.06 f0.05 10.05 10.05 10.05
PVH 4.53 3.90' 3.75C 3.93C 4.07C 6.74C 6.2w f0.04 6 0.09 f 0.11 60.04 f 0.07 60.09 f 0.05
Values are expressed as Mean & SEM; c, P < 0.001 (Students t-test); Treated versus control group.
ther corroboration is obtained by the agonistic and antagonistic effect of muscimol and picrotoxin respectively in these studies. The possible roles of catecholamines containing neurons in the regulation of appetite- controlling behaviour have been also reported (4, 11, 19, 23). Moreover, GABA could minimize the intake of food and thereby lower body weight in females with stimulated higher levels of prolactin in circulation, where food intake was to be in~creased (18).
Opposite effects of centrally administered GABA and muscimol to picrotoxin have been clearly demonstrated in studies with water intake. Whether this is due to a direct effect on PVH, which interferes with drinking, is difficult to determine. GABAergic drugs have a role in the release of posterior pituitary hormones (5) which might have indi- rectly altered the water balance in the rat (16, 18). In ad&tion, the

decrease in food intake could be the possible reason for the observed decrease of water intake, because assimilation and utilization of food are correlated with water intake (8).
The electrophysiological data of LHA, VMH and PVH recorded in the form of EEG could provide neither any concrete evidence nor corre- lation with respect to either appetitive regulatory mechanism or GABA- ergic drug responses of LHA, VMH and PVH. No definitive evidences have been presented concerning the identity of other transmitters media- ting the function of other neuronal inputs to LHA. Moreover, the exi- stence of reciprocal inhibitory relationships between VMH and LHA (15, 22) has also been emphasized, e.g. a possibility that VMH sends GABA inhibitory fibres to the LHA or vice-versa, cannot be ruled out. All these, along with many other unknown parameters, might be respon- sible for unsynchronized EEG changes with GABAergic drugs. Although further studies might provide definitive answers to these querries, yet these studies showed a possible involvement of GABA in hypothalamic regulatory mechanisms concerned with food intake.
The authors are thankful to the CSIR, New Delhi for providing financial assistance to AKR as JRF and SRF (Grant No. 9/254(12)/84-EMR-I).
REFERENCES
1. ANAND, B. K. 1961. Nervous regulation of food intake. Physiol. Rev. 41: 677-708.
2. ANAND, B. K. and BROBECK, J. R. 1951. Localization of a "feeding center" in the hypothalamus of the rat. Proc. Soc. Exp. Biol. Med. 77: 323-324.
3. ANAND, B. K. and BROBECK, J . R. 1951. Hypothalamic control of food intake in rats and cats. Yale J. Biol. Med. 24: 123-140.
4. BERGER, B. D., WISE, C. D. and STEIN, L. 1973. Nerve growth factor: Enhanced recovery of feeding after hypothalamic damage. Science (New York) 180: 506-508.
5. BRENNAN, J. J., MORIS, M. and HAYWOOD, J. R. 1984. GABA agonists inhibit the vasopress in-dependent pressor effects of central angiotensin. 11. Neuroendocrinology 39: 429-436.
6. DEGROOT, J. 1972. The rat forebrain in stereotaxic coordinates. Trans. Roy. Neth. Acad. Sci. 52: 1-40.
7. DHATT, R. K. 1987. Effect of morphine on feeding responses and electroen- cephalography of male rats. M. Phil. Thesis, Guru Nanak Dev University, Amritsar.
8. DHATT, R. K., RATTAN, A. K. and MANGAT, H. K. 1988. Effect of chronic intracerebroventricular morphine to feeding responses in male rats. Phy- siol. Behav. 43: 553-557.
9. GOLD, R. M. 1967. Aphagia and adipsia following unilateral and bilaterally asymmetrical lesions in rats. Physiol. Behav. 2: 211-220.

GRANDISON, S. and GUIDOTTI, A. 1977. Stimulation of food intake by muscimol and beta endorphin. Neuropharmacology 16: 533-536.
HILLARP, N. A,, FUXE, M. and DAHLSTROM, A. 1966. Demonstration and mapping of central neurons containing dopamine, noradrenaline and 5-hy- droxy tryptamine and their reactions to psychopharmaca. Pharmacol. Rev. 18: 727-741.
KIMURA, H. and RURIYAMA, K. 1975. Distribution of gamma-aminobutyric acid (GABA) in the rat hypothalamus: Functional correlates of GABA with activities of appetite controlling mechanisms. J. Neurochem. 24: 903- 907.
LEIBOWITZ, S. F. and HOR, L. 1982. Endiorphinergic and a-nwradrenegxic systems in the paraventricular nucleus: Effects on eating behavior. Peptides 3: 421-428.
MORGANE, P. J. 1961. Alterations in feeding and drinking behavior of rats with lesions in globi pallidi. Am. J. Physiol. 201: 420-428.
OOMURA, Y., OOYAMA, H., YAMAMOTO, T., NAKA, F., KOBAYASHI, N. and ONO, T. 1967. In W. R. Adey and T. Tokizane (ed.), Progress in brain research. Vol. 27. Elsevier, Amsterdam, p. 1-33.
RATTAN, A. K. 1985. Effect of gamma-aminobutyric acid (GABA) on the feeding and satiety areas of hypothalamus. M. Phil. Thesis, Guru Nanak Dev University, Amritsar.
RATTAN, A. K. 1987. Possible modulation of hypothalarno-hypophyseal regu- latory mechanisms by gamma-aminobutyric acid. Ph. D. Thesis, Guru Nanak Dev University, Amritsar.
RATTAN, A. K., ICAKARIA, V. K. and MANGAT, H. K. 1988. Effect of third ventricle injection of GAGA of feeding responses and blood sugar in intact and ovariectomized female rats. Ind. J. Physiol. Pharmacol. 3: 132-138.
SLANGEN, J. L. and MILLER, N. E. 1969. Pharmacological tests for the func- tion of hypothalamic norepinephrine in eating behavior. Physiol. Behav. 4: 543-552.
SOKAL, IZ. R. and ROHLF, F. J. 1981. Biometry: The principles and practices of statistics in biological research, (2nd ed.). W. ff. Freeman and Co., San Francisco, p. 145-147.
UNGERSTEDT, U. 1971. Adipsia and aphagia after 6-hydroxy-doparnine in- duced degeneration of the nigrostriatal dopamine system. Acta Physiol. Scand. (Suppl.) 367: 95-122.
VAN ATTA, L. and SUTIN, J. 1971. The response of single lateral hypotha- lamic neurons to ventromedial nucleus and limbic stimulation. Phyjiol. Behav. 6: 523-536.
ZIGMOND, M. J. and STRICKER, E. M. 1973. Recovery of feeding and drin- king by rats after intraventricular 6-hydroxydopamine or lateral hypotha- lamic lesions. Science 182: 717-720.
Accepted 5 October 1989
A. K. RATTAN, Department of Pharmacology, The Ohio State University, Colutnbus, Ohio 43210-1239, USA.





























