Embed Size (px)
Citation preview

ORNITOLOGIA NEOTROPICAL 15 (Suppl.): 61–72, 2004© The Neotropical Ornithological Society
EL NIÑO EFFECTS ON AVIAN ECOLOGY: LESSONS LEARNED FROM THE SOUTHEASTERN PACIFIC
Fabián M. Jaksic
Center for Advanced Studies in Ecology & Biodiversity, Pontificia Universidad Católica de Chile, Santiago, Chile. E-mail: [email protected]
Resumen. – Efectos de El Niño sobre ecología de aves: lecciones aprendidas en el Pacífico sudori-ental. – A través de una revisión de la literatura proveniente del borde suroriental de la cuenca del Pacífico,intento identificar qué aspectos de El Niño determinan la disponibilidad de recursos (alimento y hábitat)que a su vez afectan la ecología de las aves. La evidencia muestra que El Niño afecta a las aves marinasesencialmente al reducir su base alimentaria. Las aves marinas comedoras de peces y de peces + calamaresgeneralmente decrecen en abundancia poblacional, posiblemente como resultado combinado de la morta-lidad de adultos (hambruna) y de la falla reproductiva. Las aves marinas omnívoras sobreviven los períodosEl Niño sin declinaciones importantes, mientras que aquellas que incluyen desechos pesqueros en sus die-tas muestran cambios mínimos o inclusive incrementan sus poblaciones durante tales períodos. Las avesacuáticas parecen beneficiarse de la precipitación que acompaña a El Niño, la cual agranda sus hábitat(cuerpos acuáticos). El hábitat incrementado provee mayores oportunidades para instalar nidos adecuadosy para obtener alimento suficiente. Finalmente, las aves terrestres son afectadas por las precipitaciones aso-ciadas a El Niño, las que desencadenan la productividad primaria (biomasa aérea de plantas y banco desemillas), la cual a su vez permite incrementos en artrópodos y mamíferos. Las aves granívoras reaccionanrápidamente a los incrementos en semillas, mientras que las insectívoras responden de manera similar a laabundancia de artrópodos. Las aves carnívoras toman algo más de tiempo en reaccionar a los incrementosen poblaciones de mamíferos. Mis conclusiones son que: (a) La escasez de alimento puede ser un factorclave para las poblaciones de aves, pero sus efectos dependen de la amplitud de dieta, alternancia de presas,y parámetros de historia de vida; (b) Los patrones de productividad predecibles (estacionales) puedendeterminar la distribución y abundancia de aves residentes, mientras que las variaciones impuestas por ocu-rrencias impredecibles de El Niño y La Niña pueden determinar la presencia y abundancia de aves migra-torias; (c) La dicotomía El Niño versus La Niña u otros años “normales” puede ser artificial; (d) Losestudios de largo plazo son necesarios para entender la conexión entre el clima y las aves.
Abstract. – Through a literature review from the southeastern fringe of the Pacific Basin, I attempt toidentify what aspects of El Niño determine the availability of resources (food and habitat) that in turnaffect avian ecology. The evidence shows that El Niño affects seabirds essentially by reducing their foodbase. Fish- and fish + squid-eating seabirds generally decrease their population abundance, likely as a com-bined result of adult mortality (famine) and reproductive failure. Omnivorous seabirds negotiate El Niñoperiods without important declines, while those including fishery wastes in their diets show minimalchanges or even increase their populations during such periods. Waterbirds seem to benefit from precipita-tion that accompanies El Niño, which enlarges their habitats (waterbodies). Increased habitat affords moreopportunities for installing adequate nests and obtaining enough food. Finally, landbirds are affected by ElNiño-driven precipitation, which fires up primary productivity (aboveground plant biomass and seedcrop), which in turn fuels arthropod and mammal increases. Granivorous birds react promptly to peaks inseed output, while insectivores respond similarly to arthropod abundance. Carnivorous birds take some-what longer to react to increases in mammal populations. I conclude that (a) Food shortage may be an
61

JAKSIC
important key factor for bird populations, but its effects depend on diet breadth, prey switching, and lifehistory parameters; (b) Predictable (seasonal) patterns of productivity may determine distribution andabundance of resident birds, but variations imposed by unpredictable occurrences of El Niño and La Niñamay determine the presence and abundance of migratory birds; (c) The dichotomy El Niño versus LaNiña or other “normal” years may be artificial; (d) Long-term monitoring is needed to understand the con-nection between climate and birds. Accepted 9 December 2003.
Key words: El Niño, La Niña, effects, seabirds, waterbirds, landbirds, food resources.
INTRODUCTION
There is a large and disperse body of litera-ture reporting that El Niño affects differentaspects of avian ecology, ranging from indi-vidual to population and community aspects.Most of the evidence is anecdotal, qualitativeor correlational at the most, with fewattempts to relate emerging patterns to under-lying causes (see recent reviews by England2000, Jaksic 2001, Jaksic et al. ms.). A pioneerexception was the publication of the proceed-ings of the “Symposium on birds and the ElNiño Southern Oscillation” (Schreiber &Schreiber 1988), which concentrated on ElNiño 1982. That symposium was meant toencourage ornithologists to carry out long-term studies that could shed light on theinteractions between global atmosphericcycles, oceanographic phenomena, and avianpopulations. But ornithologists did not followthe advice. Most of the blame may be placedon the fact that the following El Niño’s weremuch weaker and less spectacular than that of1982. Indeed, those of 1986 and 1991 practi-cally failed to elicit interest from ornitholo-gists. Only with the advent of the strong ElNiño of 1997 there was a renewed effort ontrying to understand this phenomenon withregard to birds (Jaksic et al. ms.). Here, Iintend to provide a short synthesis on theputative effects of El Niño on birds, througha review of accumulated evidence. I attemptto disentangle what aspects of El Niño deter-mine the availability of food and habitatresources that in turn affect avian ecology in
the southeastern Pacific.
METHODS
I concentrated my literature search in thesoutheastern fringe of the Pacific Basin, anarea with which I am more familiar and wheremost of the known effects of El Niño onbirds come from. I searched the primary liter-ature for quantitative, or at least categorical,data on bird abundance, mortality and repro-duction, in connection with fluctuations inprey and habitat resources. The literaturesources and specific methods used may befound in Jaksic et al. (ms.) I recognized threefunctional bird types: seabirds, waterbirds,and landbirds, regardless of systematic affini-ties. With regard to seabirds, I analyzed semi-quantitative information in 10 studies onthe effects of El Niño on abundance andreproduction of seabird populations in fourareas of the South Pacific Ocean (Jaksic et al.ms.). I characterized the diet of 41 seabirdspecies. By summing abundance and repro-ductive effects, either negative or positive orneutral, one arrives at a single total effect ofEl Niño for each of the 41 seabird species.For waterbirds, the amount of informationavailable was so limited that there was noneed to use screening or summary proce-dures. For landbirds, I further divided thiscategory among granivorous, insectivorousand carnivorous birds, and obtained as muchquantitative information as feasible. Most ofthese data are unpublished results from thisauthor.
62

EL NIÑO EFFECTS ON AVIAN ECOLOGY
RESULTS
SeabirdsThere is an extensive body of literature onthese birds. I chose not to cite specific papersin this section, because a detailed analysis ofsuch literature is being published elsewhere(Jaksic et al. ms.). Below is a summary of myfindings.
Effects on migration, abundance & distribution ofseabirds. Massive migrations of resident sea-birds away from equatorial waters occur dur-ing El Niño events. Distributions of seabirdsshift with movements of their preferred watertypes: During El Niño events, cool-water spe-cies become less abundant, and warm-waterspecies more abundant. Species normally notpresent appear in extra-tropical areas.
Effects on mortality & reproduction of seabirds.Mortality of guano birds as a result of El Niñois well known. During El Niño, large numbersof dead and dying birds are observedthroughout the eastern Pacific. The absenceof, or the decrease in, adults attempting tonest during El Niño results in a lower repro-ductive output for eastern Pacific seabirds.Species show smaller clutch size, reducedchick size and lower survival.
Effects of food web structure & rainfall regime on sea-birds. Migration and mortality of seabird pop-ulations could be associated with alterationsof the marine food web, while changes inreproduction could be related to increasedrainfall. Changes in the marine food webinvolve a replacement of primary producers –from dinoflagellates to diatoms –, and of pri-mary consumers – from copepods to chaetog-naths –. This in turn produces the migrationto deeper water of the main secondary con-sumers – anchovies and sardines –, the pre-ferred prey of seabirds. Hence follow massivemigrations and die-offs of seabirds. Concur-
rent changes such as a higher plant cover dueto increased rainfall, and flooding of seabirdreproductive grounds due to increased sealevel, are also important causes of reproduc-tive failures.
Effects of fisheries on seabirds. Due to their depen-dence on anchovies, which usually disappearduring El Niño, seabirds are highly sensible tothe occurrence of this event. Overfishing hasled to a dramatic reduction in catches ofanchovies. Because of this, the fisheryswitched to sardines in Peru and to horsemackerel in Chile. The recovery rate of sea-birds after El Niño events has been gettingincreasingly slower. A possible explanation forthis phenomenon is that the anchovy fisherytook up the superabundance of food onwhich the seabirds depended.
Trophic ecological effects on seabirds. Seabird dietsconsist mainly of three prey types: Fishes,crustaceans, and mollusks. The problem offinding enough food to survive and reproduceis solved in three ways. Coastal or inshore sea-birds, such as gulls and terns, gather to feed inareas where prey are abundant or where theyare forced to the surface. Diving seabirds,including penguins and alcids, exploit agreater range of depths to obtain food.Pelagic species, such as albatrosses and shear-waters, soar over vast expanses of the ocean insearch of widely dispersed surface prey. Dur-ing periods of reduced food availability, suchas El Niño years, seabirds may be forced toswitch their diet. Seabirds with a restricteddietary range and with no chances to effectprey switching, suffer important reproductivefailures. Seabirds with a broad dietary rangeand with abilities to perform prey switching,may display no changes in reproduction andsurvival.
With regard to my own analyses of tenstudies reporting data on 41 seabird species, asummary follows (see Table 1 and Fig. 1). The
63
JAKSIC
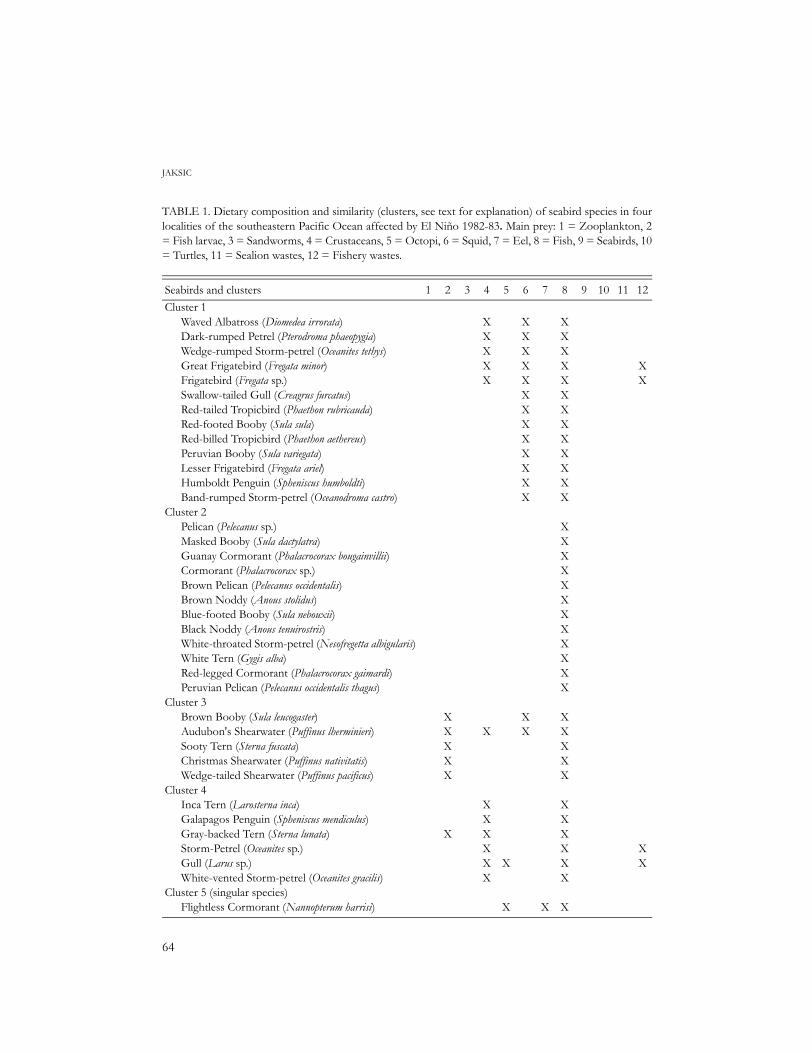
TABLE 1. Dietary composition and similarity (clusters, see text for explanation) of seabird species in fourlocalities of the southeastern Pacific Ocean affected by El Niño 1982-83. Main prey: 1 = Zooplankton, 2= Fish larvae, 3 = Sandworms, 4 = Crustaceans, 5 = Octopi, 6 = Squid, 7 = Eel, 8 = Fish, 9 = Seabirds, 10= Turtles, 11 = Sealion wastes, 12 = Fishery wastes.
Seabirds and clusters 1 2 3 4 5 6 7 8 9 10 11 12Cluster 1
Waved Albatross (Diomedea irrorata)Dark-rumped Petrel (Pterodroma phaeopygia)Wedge-rumped Storm-petrel (Oceanites tethys)Great Frigatebird (Fregata minor)Frigatebird (Fregata sp.)Swallow-tailed Gull (Creagrus furcatus)Red-tailed Tropicbird (Phaethon rubricauda)Red-footed Booby (Sula sula)Red-billed Tropicbird (Phaethon aethereus)Peruvian Booby (Sula variegata)Lesser Frigatebird (Fregata ariel)Humboldt Penguin (Spheniscus humboldti)Band-rumped Storm-petrel (Oceanodroma castro)
Cluster 2Pelican (Pelecanus sp.)Masked Booby (Sula dactylatra)Guanay Cormorant (Phalacrocorax bougainvillii)Cormorant (Phalacrocorax sp.)Brown Pelican (Pelecanus occidentalis)Brown Noddy (Anous stolidus)Blue-footed Booby (Sula nebouxii)Black Noddy (Anous tenuirostris)White-throated Storm-petrel (Nesofregetta albigularis)White Tern (Gygis alba)Red-legged Cormorant (Phalacrocorax gaimardi)Peruvian Pelican (Pelecanus occidentalis thagus)
Cluster 3Brown Booby (Sula leucogaster)Audubon's Shearwater (Puffinus lherminieri)Sooty Tern (Sterna fuscata)Christmas Shearwater (Puffinus nativitatis)Wedge-tailed Shearwater (Puffinus pacificus)
Cluster 4Inca Tern (Larosterna inca)Galapagos Penguin (Spheniscus mendiculus)Gray-backed Tern (Sterna lunata)Storm-Petrel (Oceanites sp.)Gull (Larus sp.)White-vented Storm-petrel (Oceanites gracilis)
Cluster 5 (singular species)Flightless Cormorant (Nannopterum harrisi)
XXXXX
X
XXXXX
X
XXXXXX
X
X
XXXXXXXXXXXXX
XX
X
XXXXXXXXXXXXX
XXXXXXXXXXXX
XXXXX
XXXXXX
X
XX
XX
64
EL NIÑO EFFECTS ON AVIAN ECOLOGY
most numerous trophic or dietary group(cluster 1) was composed of seabirds feedingon fishes and squid. A second group (cluster2) was made up of seabirds feeding strictly onfishes. A third group (cluster 3) was com-posed of seabirds that fed on fishes and fish
larvae, and a fourth group (cluster 4) byseabirds feeding on crustaceans and fisherywastes. The remaining five species showeddiverse and peculiar diets sharing low similar-ity. By averaging species information foreach dietary group one finds that the five
TABLE 1. Continuation.
Seabirds and clusters 1 2 3 4 5 6 7 8 9 10 11 12Magnificent Frigatebird (Fregata magnificens)Lava Gull (Larus fuliginosus)Phoenix Petrel (Pterodroma alba)Northern Phalarope (Lobipes lobatus) X
XX
XX
XX
X
X XX
X X XX
XX
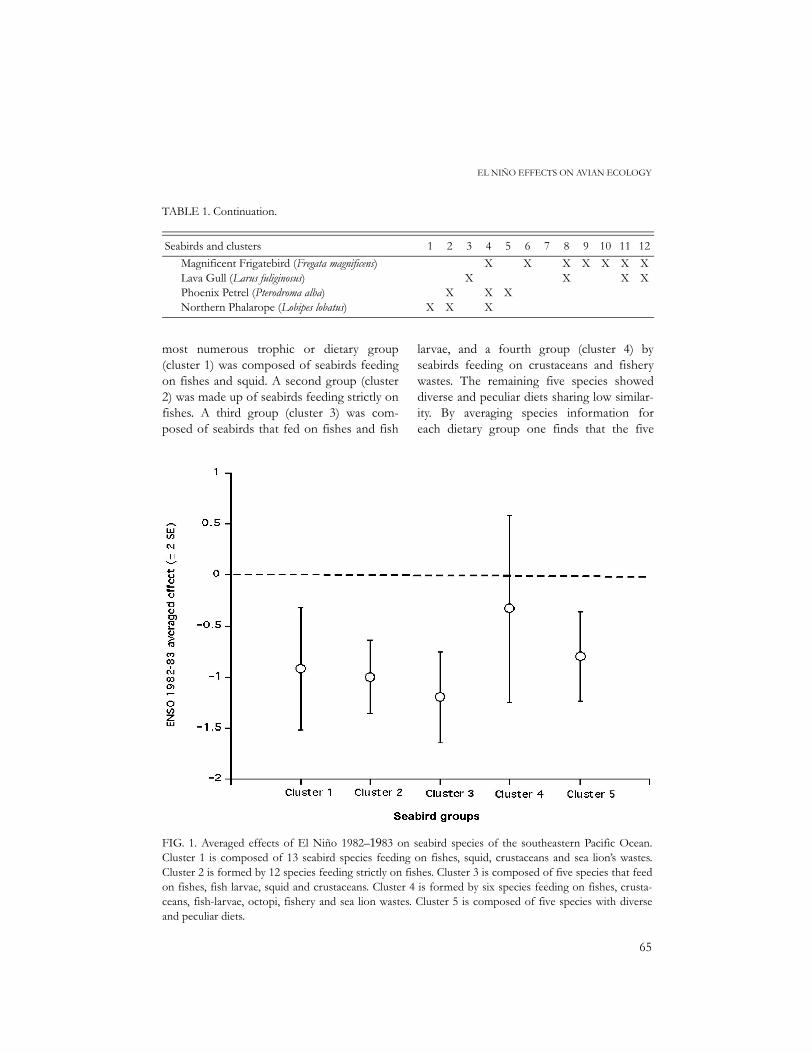
FIG. 1. Averaged effects of El Niño 1982–1983 on seabird species of the southeastern Pacific Ocean.Cluster 1 is composed of 13 seabird species feeding on fishes, squid, crustaceans and sea lion’s wastes.Cluster 2 is formed by 12 species feeding strictly on fishes. Cluster 3 is composed of five species that feedon fishes, fish larvae, squid and crustaceans. Cluster 4 is formed by six species feeding on fishes, crusta-ceans, fish-larvae, octopi, fishery and sea lion wastes. Cluster 5 is composed of five species with diverseand peculiar diets.
65
JAKSIC
groups were negatively affected by El Niño,but to different extents (Fig. 1). The mostaffected group was cluster 3 composedby seabirds with a diet based mainly on fishand fish larvae. Slightly less affected werespecies in clusters 1 and 2, feeding chieflyon fishes and squid. Less affected were spe-cies in cluster 5, which fed on a variety of dif-ferent prey. Finally, there was a group littleaffected (cluster 4) and even with positiveeffects for some species, which had in com-mon consumption of crustaceans as well asfishery wastes. In sum, my analyses show thatseabirds with a narrow diet are the mostaffected by El Niño, while species with abroader diet are less affected. In addition, itappears that the inclusion of fishery wastes inthe diet alleviates the shortage of food. Sea-birds with the ability to switch and use thisfood type are practically not affected by ElNiño.
WaterbirdsFor waterbirds, the amount of informationavailable was very limited, and thus I citedbelow all that was relevant.
Habitat & food effects on waterbirds. The distribu-tion and area of wetlands are highly variableand depend importantly on climate condi-tions, especially rainfall. Vilina & Cofre (2000)analyzed the seasonal abundance four grebespecies in a central Chilean wetland. Theyfound a positive correlation between springabundance of these species and total rainfallthe previous year. Rain fell chiefly during thewinter and spring of El Niño year; grebe pop-ulations increased during the summer, andpeaked in the spring following El Niño. Vilinaet al. (2002) analyzed the relationship betweenEl Niño, rainfall, and changes in abundance,breeding and local distribution of the Black-necked Swan (Cygnus melanocorypha) in a central
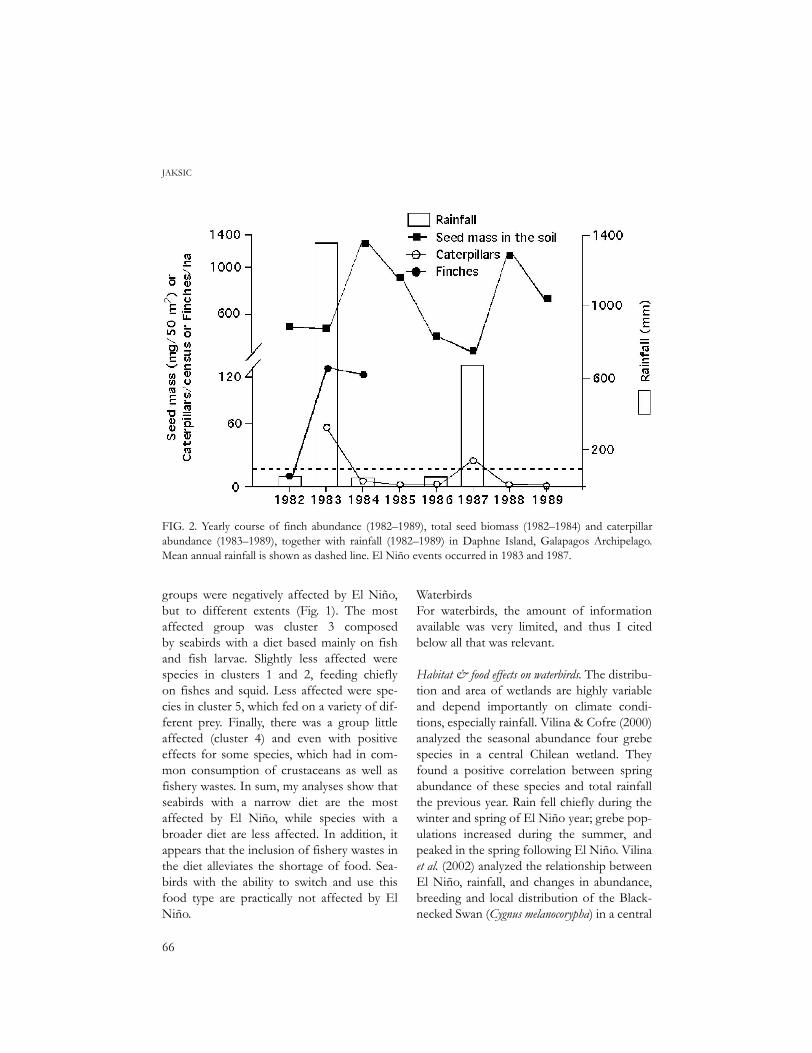
FIG. 2. Yearly course of finch abundance (1982–1989), total seed biomass (1982–1984) and caterpillarabundance (1983–1989), together with rainfall (1982–1989) in Daphne Island, Galapagos Archipelago.Mean annual rainfall is shown as dashed line. El Niño events occurred in 1983 and 1987.
66
EL NIÑO EFFECTS ON AVIAN ECOLOGY
Chilean wetland. During and after El Niñoevent, swans increased in abundance, breed-ing success, and time of permanence. Schlat-ter et al. (2002) showed that Black-neckedSwan populations in a wetland of southernChile more than doubled during La Niñaperiod. They hypothesized that due to migra-tions from northern wetlands, populations inthe south peaked during dry years associatedwith La Niña. In sum, waterbirds benefit fromprecipitation that accompanies El Niño,which enlarges the water surface of marshes,coastal lagoons, and other water bodies.Apparently, increased habitat affords moreopportunities for installing adequate nests andobtaining sufficient food.
LandbirdsAlthough there is not a lot of literature onthese birds, I preferred not to cite specificpapers in this section, because of my detailedanalysis being published elsewhere (Jaksic etal. ms.). Below is a summary of my findings.
Contrasting effects on landbirds. Landbirds areaffected by El Niño in two contrasting ways:Negatively, because when El Niño arrives,excessive precipitation may result in nestingfailure and increased chick mortality due tonest flooding, thus lowering reproductive suc-cess. On the other hand, El Niño-driven pre-cipitation fires up primary productivity (plantbiomass and seed crop), which in turn fuelsinsect and mammal increases, thus positivelyaffecting bird populations.
Ambiguous effects on granivorous birds. In DaphneIsland, Ecuador (Fig. 2), seed abundancepeaked with El Niño of 1983, and decreasedslightly thereafter. Finches peaked one yearafter the rainfall and seed peak, subsequentlydeclined, and again peaked with 1-year lagafter the high-rainfall year brought by El Niño1987. In Auco, semiarid Chile (Fig. 3), theincreased rainfall brought by El Niño 1997resulted in an immediate increase in annualgrass seeds, in a lack of response in perennial
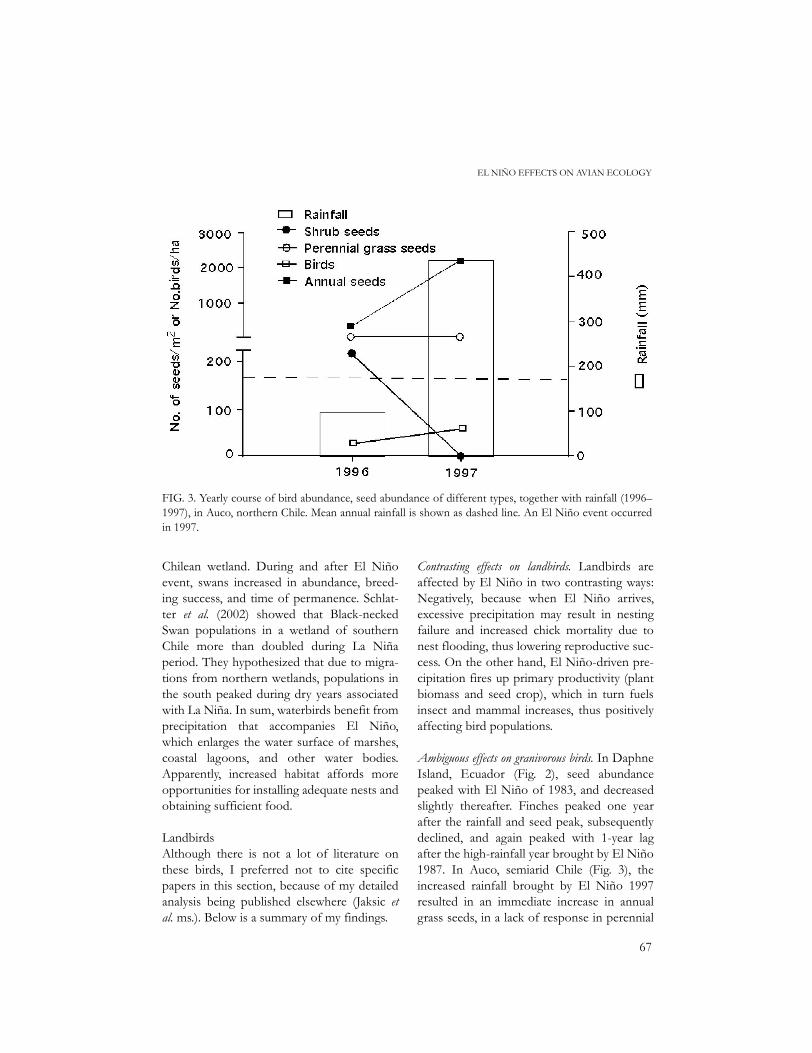
FIG. 3. Yearly course of bird abundance, seed abundance of different types, together with rainfall (1996–1997), in Auco, northern Chile. Mean annual rainfall is shown as dashed line. An El Niño event occurredin 1997.
67
JAKSIC
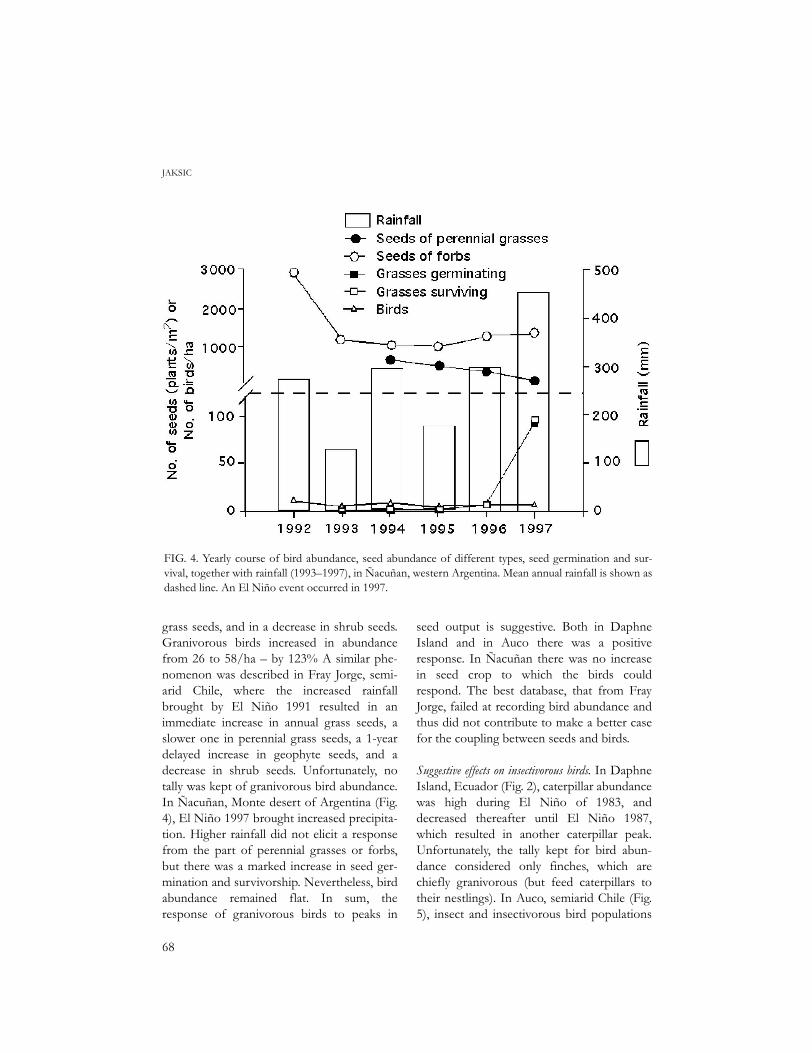
grass seeds, and in a decrease in shrub seeds.Granivorous birds increased in abundancefrom 26 to 58/ha – by 123% A similar phe-nomenon was described in Fray Jorge, semi-arid Chile, where the increased rainfallbrought by El Niño 1991 resulted in animmediate increase in annual grass seeds, aslower one in perennial grass seeds, a 1-yeardelayed increase in geophyte seeds, and adecrease in shrub seeds. Unfortunately, notally was kept of granivorous bird abundance.In Ñacuñan, Monte desert of Argentina (Fig.4), El Niño 1997 brought increased precipita-tion. Higher rainfall did not elicit a responsefrom the part of perennial grasses or forbs,but there was a marked increase in seed ger-mination and survivorship. Nevertheless, birdabundance remained flat. In sum, theresponse of granivorous birds to peaks in
seed output is suggestive. Both in DaphneIsland and in Auco there was a positiveresponse. In Ñacuñan there was no increasein seed crop to which the birds couldrespond. The best database, that from FrayJorge, failed at recording bird abundance andthus did not contribute to make a better casefor the coupling between seeds and birds.
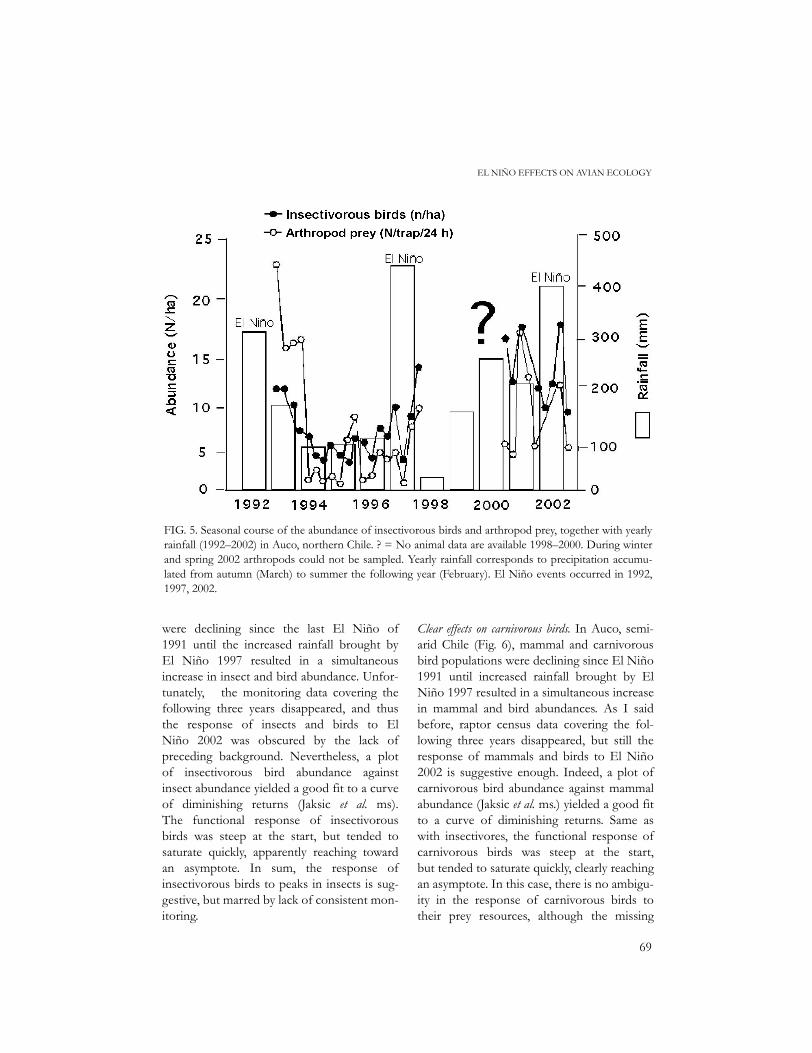
Suggestive effects on insectivorous birds. In DaphneIsland, Ecuador (Fig. 2), caterpillar abundancewas high during El Niño of 1983, anddecreased thereafter until El Niño 1987,which resulted in another caterpillar peak.Unfortunately, the tally kept for bird abun-dance considered only finches, which arechiefly granivorous (but feed caterpillars totheir nestlings). In Auco, semiarid Chile (Fig.5), insect and insectivorous bird populations
FIG. 4. Yearly course of bird abundance, seed abundance of different types, seed germination and sur-vival, together with rainfall (1993–1997), in Ñacuñan, western Argentina. Mean annual rainfall is shown asdashed line. An El Niño event occurred in 1997.
68
EL NIÑO EFFECTS ON AVIAN ECOLOGY
were declining since the last El Niño of1991 until the increased rainfall brought byEl Niño 1997 resulted in a simultaneousincrease in insect and bird abundance. Unfor-tunately, the monitoring data covering thefollowing three years disappeared, and thusthe response of insects and birds to ElNiño 2002 was obscured by the lack ofpreceding background. Nevertheless, a plotof insectivorous bird abundance againstinsect abundance yielded a good fit to a curveof diminishing returns (Jaksic et al. ms).The functional response of insectivorousbirds was steep at the start, but tended tosaturate quickly, apparently reaching towardan asymptote. In sum, the response ofinsectivorous birds to peaks in insects is sug-gestive, but marred by lack of consistent mon-itoring.
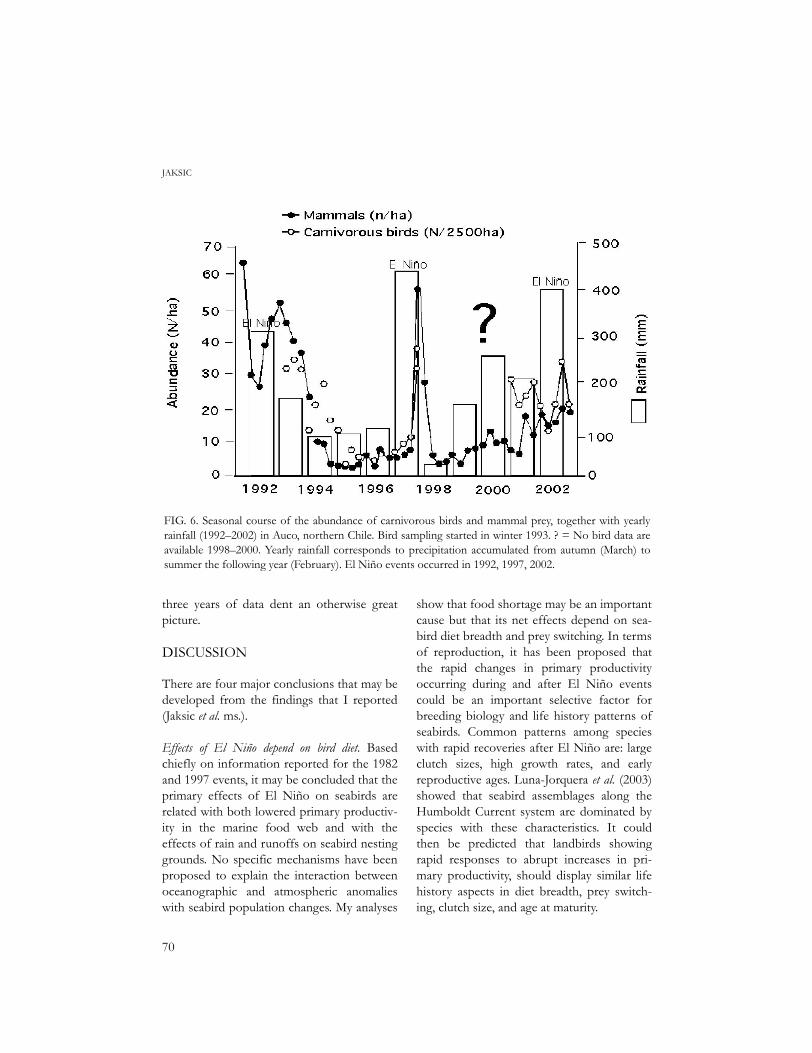
Clear effects on carnivorous birds. In Auco, semi-arid Chile (Fig. 6), mammal and carnivorousbird populations were declining since El Niño1991 until increased rainfall brought by ElNiño 1997 resulted in a simultaneous increasein mammal and bird abundances. As I saidbefore, raptor census data covering the fol-lowing three years disappeared, but still theresponse of mammals and birds to El Niño2002 is suggestive enough. Indeed, a plot ofcarnivorous bird abundance against mammalabundance (Jaksic et al. ms.) yielded a good fitto a curve of diminishing returns. Same aswith insectivores, the functional response ofcarnivorous birds was steep at the start,but tended to saturate quickly, clearly reachingan asymptote. In this case, there is no ambigu-ity in the response of carnivorous birds totheir prey resources, although the missing
FIG. 5. Seasonal course of the abundance of insectivorous birds and arthropod prey, together with yearlyrainfall (1992–2002) in Auco, northern Chile. ? = No animal data are available 1998–2000. During winterand spring 2002 arthropods could not be sampled. Yearly rainfall corresponds to precipitation accumu-lated from autumn (March) to summer the following year (February). El Niño events occurred in 1992,1997, 2002.
69
JAKSIC
three years of data dent an otherwise greatpicture.
DISCUSSION
There are four major conclusions that may bedeveloped from the findings that I reported(Jaksic et al. ms.).
Effects of El Niño depend on bird diet. Basedchiefly on information reported for the 1982and 1997 events, it may be concluded that theprimary effects of El Niño on seabirds arerelated with both lowered primary productiv-ity in the marine food web and with theeffects of rain and runoffs on seabird nestinggrounds. No specific mechanisms have beenproposed to explain the interaction betweenoceanographic and atmospheric anomalieswith seabird population changes. My analyses
show that food shortage may be an importantcause but that its net effects depend on sea-bird diet breadth and prey switching. In termsof reproduction, it has been proposed thatthe rapid changes in primary productivityoccurring during and after El Niño eventscould be an important selective factor forbreeding biology and life history patterns ofseabirds. Common patterns among specieswith rapid recoveries after El Niño are: largeclutch sizes, high growth rates, and earlyreproductive ages. Luna-Jorquera et al. (2003)showed that seabird assemblages along theHumboldt Current system are dominated byspecies with these characteristics. It couldthen be predicted that landbirds showingrapid responses to abrupt increases in pri-mary productivity, should display similar lifehistory aspects in diet breadth, prey switch-ing, clutch size, and age at maturity.
FIG. 6. Seasonal course of the abundance of carnivorous birds and mammal prey, together with yearlyrainfall (1992–2002) in Auco, northern Chile. Bird sampling started in winter 1993. ? = No bird data areavailable 1998–2000. Yearly rainfall corresponds to precipitation accumulated from autumn (March) tosummer the following year (February). El Niño events occurred in 1992, 1997, 2002.
70

EL NIÑO EFFECTS ON AVIAN ECOLOGY
El Niño imposes unpredictability to bird assemblage.The structure of bird assemblages may bedetermined by basal levels of primary produc-tivity, but also by the predictable (seasonal)and unpredictable (ENSO) fluctuations ofthat variable. Hurlbert & Haskell (2003)showed that basal levels of primary productiv-ity determine resident species richness, whilethe range of predictable variation (or season-ality) of this variable determines migrant spe-cies richness. The approach of Hurlbert &Haskell could be used in the search of generalmechanisms for ENSO effects in both terres-trial and marine bird assemblages. It could bethat superimposed on seasonally predictablechanges of productivity, the range of variationimposed by unpredictable but rather frequentoccurrences of El Niño and La Niña deter-mine geographic distribution and populationabundance of the birds living in areas affectedby these anomalies.
El Niño and La Niña are extremes of a wide rangeof variability. The occurrence and magnitude ofEl Niño are declared after some pre-definedthreshold is reached, according to atmo-spheric or oceanographic indices such as seasurface temperature and southern oscillationindex. Nevertheless, physical fluctuations(anomalies) are continuously occurring at dif-ferent time scales. These anomalies are usuallytreated as departures from normality, wherenormal conditions represent either climaticmeans or non El Niño/La Niña conditions.Thus, normality is not treated as a real phe-nomenon but as the absence of extreme con-ditions. However, El Niño/La Niñathresholds encompass a wide range of realphysical fluctuations that may have effects onbird assemblage attributes. In addition, thereappears to be a weak correlation between themagnitude of El Niño/La Niña events asmeasured by physical indices, and the magni-tude of recorded biological effects. Therefore,physical and biological dynamics do not have
a direct coupling, and thus environmental sig-nals may be filtered by terrestrial and marinecommunities in such a way that they mayresist some El Niño/La Niña events regard-less of their magnitude.
The need for long-term time series. Finally, it seemsobvious that without good baseline data weare doomed to be surprised by the arrival ofEl Niño every few years. Even when ornithol-ogists are at hand to take advantage of anincoming El Niño, lack of preceding data, andof monitoring afterwards, makes it difficult tounderstand responses of birds to the succes-sive El Niño, La Niña, and normal years.Indeed, during the last century there were 12El Niño years and 12 La Niña years, thus leav-ing about 76 normal years in between (Jaksic2001). Thus, by heavily concentrating atten-tion on only 12% of the time span (El Niño),and of neglecting another 12% (La Niña), weare essentially ignoring what happens during76% of the time. This situation may be reme-died only as long as data are logged on a con-tinuous basis, that is, as long-term time series.
ACKNOWLEDGMENTS
This research was funded by grant FONDAP-FONDECYT 1501-0001 to the Center forAdvanced Studies in Ecology & Biodiversity. Iappreciate the invitation of the OrganizingCommittee of the 7th Neotropical Ornitho-logical Congress to deliver a Plenary Lectureat Termas de Puyehue, Chile. The disappeareddata from my long-term time series in Aucoare courtesy of Mr. Ivan Lazo.
REFERENCES
England, M. C. 2000. A review of bird responses toEl Niño-Southern Oscillation conditions in theNeotropics. Cotinga 13: 83–88.
Hurlbert, A. H., & J. P. Haskell. 2003. The effect ofenergy and seasonality on avian species rich-
71

JAKSIC
ness and community composition. Am. Nat.161: 83–97.
Jaksic, F. M. 2001. Ecological effects of El Niño interrestrial ecosystems of western South Amer-ica. Ecography 24: 241–250.
Jaksic, F. M., J. A. Figueroa, S. I. Silva, & J. M.Fariña. 2003. El Niño and the birds: a resource-based interpretation of climatic forcing. Manu-script submitted for publication.
Luna-Jorquera, G., A. Simenone, & R. Aguilar.2003. Ecofisiología de animales endotermos enun desierto cálido y un mar frío: el caso de lasaves marinas de la corriente de Humboldt. Pp.341–368 in Bozinovic, F. (ed.). Fisiologíaecológica y evolutiva. Ediciones Univ. Católicade Chile, Santiago, Chile.
Schlatter, R. P., R. A. Navarro, & P. Corti. 2002.Effects of El Niño Southern Oscillation onnumbers of Black-necked Swans at Rio CrucesSanctuary, Chile. Waterbirds 25: 114–122.
Schreiber, R. W., & E. A. Schreiber. 1988. Intro-duction. Proc. Int. Ornithol. Congr. 19: 1739.
Vilina, Y. A., & H. Cofre. 2000. “El Niño” effectson the abundance and habitat association pat-terns of four grebe species in Chilean wetlands.Waterbirds 32: 95–101.
Vilina, Y. A., H. L. Cofre, C. Silva-Garcia, M. D.Garcia, & C. Perez-Friedenthal. 2002. Effectsof El Niño on abundance and breeding ofBlack-necked Swans on El Yali Wetland inChile. Waterbirds 25: 123–127.
72





























