Embed Size (px)
Citation preview

, . 184: 424–429 (1998)
EGF-r GENE COPY NUMBER CHANGES IN RENALCELL CARCINOMA DETECTED BY FLUORESCENCE
IN SITU HYBRIDIZATION
1*, 1, . 2, 1, 1, . , 3, . 4 . 1
1Institute of Pathology, University of Basel, 4003 Basel, Switzerland2Urologic Clinics, University of Basel, 4003 Basel, Switzerland
3Department of Urology, University of California, San Francisco, San Francisco, CA 94143-0738, U.S.A.4Division of Molecular Cytometry, Department of Laboratory Medicine, University of California, San Francisco, San Francisco,
CA 94143-0808, U.S.A.
SUMMARY
Expression of epidermal growth factor receptor (EGF-r) is a frequent event in renal cell carcinoma (RCC). To investigate the role ofEGF-r gene copy number changes related to EGF-r overexpression, 50 RCC specimens were examined by fluorescence in situhybridization (FISH) and immunohistochemistry. Dual-labelling FISH with a repetitive pericentromeric probe for chromosome 7 and aprobe for the EGF-r gene (at 7p13) was performed to analyse the EGF-r copy number in relation to chromosome 7 copy number on acell-by-cell basis. Polysomy 7 was frequent in all histological types of RCC. Chromosome 7 polysomy was found in 26 of 35 clear cell(74 per cent), nine of nine papillary, and three of three chromophobe RCCs. EGF-r gene copy number was closely associated with thechromosome 7 copy number on a cell-by-cell basis. No EGF-r gene amplifications were found. EGF-r positivity was found in 37 of 50cases (74 per cent) by immunohistochemistry. EGF-r positivity was more common in clear cell (81 per cent) than in papillary tumours(40 per cent; P=0·029). Neither chromosome 7 nor EGF-r gene copy number was associated with EGF-r expression, indicating that anincreased gene dosage is not a mechanism of EGF-r overexpression in RCC. ? 1998 John Wiley & Sons, Ltd.
J. Pathol. 184: 424–429, 1998.
KEY WORDS—renal cell carcinoma; EGF-r gene; fluorescence in situ hybridization; chromosome 7
INTRODUCTION
EGF-r is a transmembranous receptor for epidermalgrowth factor (EGF), transforming growth factor alpha(TGF-á), and other ligands. Overexpression of epi-dermal growth factor receptor (EGF-r) is a frequentevent in renal cell carcinoma (RCC)1–3 and has beenproposed as a prognostic parameter in RCC.4–6 We haverecently found an association between EGF-r expressionand tumour cell proliferation rate in bladder cancer,7suggesting that the poor prognosis found for EGF-r-positive tumours6 may be due to increased tumour cellproliferation.The mechanism of EGF-r expression differs markedly
between tumour types. Whereas amplification of theEGF-r gene, located on chromosome 7p13, is a majorcause for EGF-r expression in brain tumours,8,9 thispathway is uncommon in breast, ovarian, and bladder
tumours. Also, in RCC, clear-cut EGF-r amplification isapparently rare.1 It has been postulated that chromo-some 7 copy number alterations might be associatedwith EGF-r overexpression in RCC,10 since polysomy 7has frequently been observed in RCC. However, therelationship between EGF-r expression and low-levelEGF-r gene copy number changes, for example, due topolysomy 7, duplication, or low-level amplification, hasnot been examined.In this study, dual-labelling fluorescence in situ
hybridization (FISH) was used with probes for EGF-rand the chromosome 7 centromere to explore the associ-ation of EGF-r gene copy number alteration with EGF-roverexpression in RCC. In addition, the relationshipbetween chromosome 7 and EGF-r gene copy numberand proliferation rate [Ki-67 labelling index (LI)] wasexamined.
MATERIALS AND METHODS
Patient material
Frozen RCC tissue samples from 50 patients wereanalysed. Histological classification was performed asdescribed by Weiss et al.11 The tumours were gradedaccording to the Thoenes grading system12 and stagedaccording to the UICC classification.13
*Correspondence to: Holger Moch, MD, Institut für Pathologie derUniversität Basel, Schönbeinstr. 40, CH-4003 Basel, Switzerland.
Contract grant sponsor: Deutsche Forschungsgemeinschaft;Contract grant number: Mo 625/1.
Contract grant sponsor: Schweizerische Krebsliga; Contract grantnumber: SKL 291-2-1996.
Contract grant sponsor: Swiss National Science Foundation;Contract grant number: 3200-043969.95.
CCC 0022–3417/98/040424–06 $17.50 Received 20 February 1997? 1998 John Wiley & Sons, Ltd. Accepted 15 October 1997

Fluorescence in situ hybridization
Four-micrometre sections were used for haematoxylinand eosin (H&E) staining. Fresh touch preparationswere made after the thawing of frozen tumour sampleswithout normal renal tissue. All slides were fixed inmethanol–acetic acid (3:1) and subsequently air-dried.FISH was performed as previously described.7 Cells onslides were denatured in 70 per cent formamide/2#standard saline citrate (SSC; 1#SSC is 0·15 mol/lNaCl, 0·015 mol/l Na citrate), pH 7, at 75)C for 2·5 min.After dehydration in graded ethanol, samples weretreated with 0·5 ìg/ml proteinase K (Sigma) for 7 min at37)C, followed by ethanol dehydration. The hybridiza-tion mixture was denatured for 5 min at 75)C andapplied to denatured cells on sides. Hybridizationmixture (10 ìl) consisted of 10 ng of a digoxigenatedcosmid probe for the EGF-r gene locus (7p13; gener-ously provided by Dr J. Haley, Oncogene Science), 4 ngof a biotinylated chromosome 7 centromeric probe(p7alphaTET; provided by the UCSF/LBL Resource ofMolecular Cytometry, Director Dr Joe Gray), and 10 ngof unlabelled, sonicated (200–500 base pairs) humanplacental DNA (Sigma) in 50 per cent formamide, 10 percent dextran sulphate, and 2#SSC (pH 7). Hybridiza-tion was performed overnight at 37)C. Slides werewashed three times in 50 per cent formamide/2#SSC(pH 7) and twice in 2#SSC for 10 min each at45)C. Probe visualization was exactly as described,7using fluorescein-conjugated sheep anti-digoxigenin(Boehringer Mannheim), fluorescein isothiocyanate-conjugated anti-sheep IgG (Sigma), and Texas Redavidin (Vector Laboratories, Burlingame, CA, U.S.A.).Nuclei were counterstained with 0·07 ìg/ml 4,5-diamino-2-phenyl-indole (DAPI) in antifade solution. If excessivenuclear damage was observed, hybridization wasrepeated without proteinase K.
Scoring of FISH signals
At least 100 nuclei were analysed from each tumour.Cells were selected for scoring according to morpho-logical criteria, using DAPI staining. Only those cellshaving a malignant cytological appearance (especiallylarge nuclei) were scored. Small, round lymphocyte-likecells and overlapping or damaged nuclei were dis-regarded. Slides were analysed only if two-thirds of thecells were interpretable in representative areas. To avoidmisinterpretation due to inefficient hybridization, cellswere counted only if one bright centromere signal andone cosmid signal were present. Two cosmid signalswere counted as one if they were situated very close toeach other (less than 0·5 ìm), to avoid misinterpretationdue to sister chromatids of cells in S or G2M phase.On a cellular level, EGF-r amplification was defined
as the presence of at least three times more EGF-rsignals than centromere 7 signals. The occurrence ofmore than two centromere 7 signals per cell (polysomy)in greater than 20 per cent of tumour cells was requiredto categorize a tumour as polysomic. A monosomy 7was defined as the presence of more than 20 per cent ofmonosomic cells (cells with one centromere 7).
Immunohistochemistry
Sections for immunohistochemistry were made fromfrozen tissue blocks before preparing the touch prepara-tions. Four-micrometer sections were examined forEGF-r expression using a mouse monoclonal antibodyEGFR (1:50, Triton Biosciences, Alameda, CA,U.S.A.), without proteinase digestion or post-fixation,with an avidin–biotin complex immunoperoxidase pro-cedure (ABC Elite, Vector, Burlingame, CA, U.S.A.).Sections were weakly counterstained with haematoxylin.The optimal titer for EGF-r was defined as the dilutionthat gave clearly identifiable membrane staining andnegligible background on human placental samples. Toassess the specificity of the reaction, both positive(human placenta) and negative (primary antibody omit-ted) controls were used. Tumours were consideredEGF-r positive if staining as strong as in the placentalcontrol was found either diffusely or focally in at least 20per cent of tumour cells, as described by Neal et al.14Tumours with EGF-r positivity weaker than theplacental control were considered negative.Cells with proliferative activity were identified by
detecting the Ki-67 protein. The Ki-67 antigen isexpressed in all cells in G1, S, or G2M phase.15 Themonoclonal antibody MIB1 (1:800; Dianova, Hamburg,Germany) was used for the detection of Ki-67 protein.In this study, antigen retrieval was not performed, sincefresh frozen tumour material was used. Nuclei wereconsidered Ki-67-positive if any nuclear staining wasseen. Only nuclear staining was considered. Ki-67labelling indices (percentage of positive cells, LI) weredetermined by scoring 300 tumour cells in the tumourareas with the highest density of positive cells.
Statistics
Contingency table analysis was used to examine therelationship between EGF-r immunostaining, polysomy7, grade, stage, and tumour growth pattern. A Studentt-test was employed to evaluate the associations betweenEGF-r staining, tumour grade, stage, polysomy 7, andKi-67 LI.
RESULTS
Patient material
Thirty-seven tumours were clear cell, ten papillary,and three chromophobe RCCs. Twenty-seven tumourswere staged as pT2, 22 tumours as pT3, and one tumouras pT4. Seven tumours were classified as grade 1, 26were grade 2, and 17 grade 3.
EGF-r and Ki-67 immunohistochemistry
EGF-r immunostaining was successfully performed inall cases. Positive EGF-r immunostaining was predomi-nantly membranous, although cytoplasmic staining wasalso present (Fig. 1). Staining of normal renal paren-chyma revealed weak cytoplasmic and membranousEGF-r expression on epithelial cells of proximal tubules,
425EGF-r COPY NUMBER CHANGES IN RENAL CELL CARCINOMA
? 1998 John Wiley & Sons, Ltd. , . 184: 424–429 (1998)
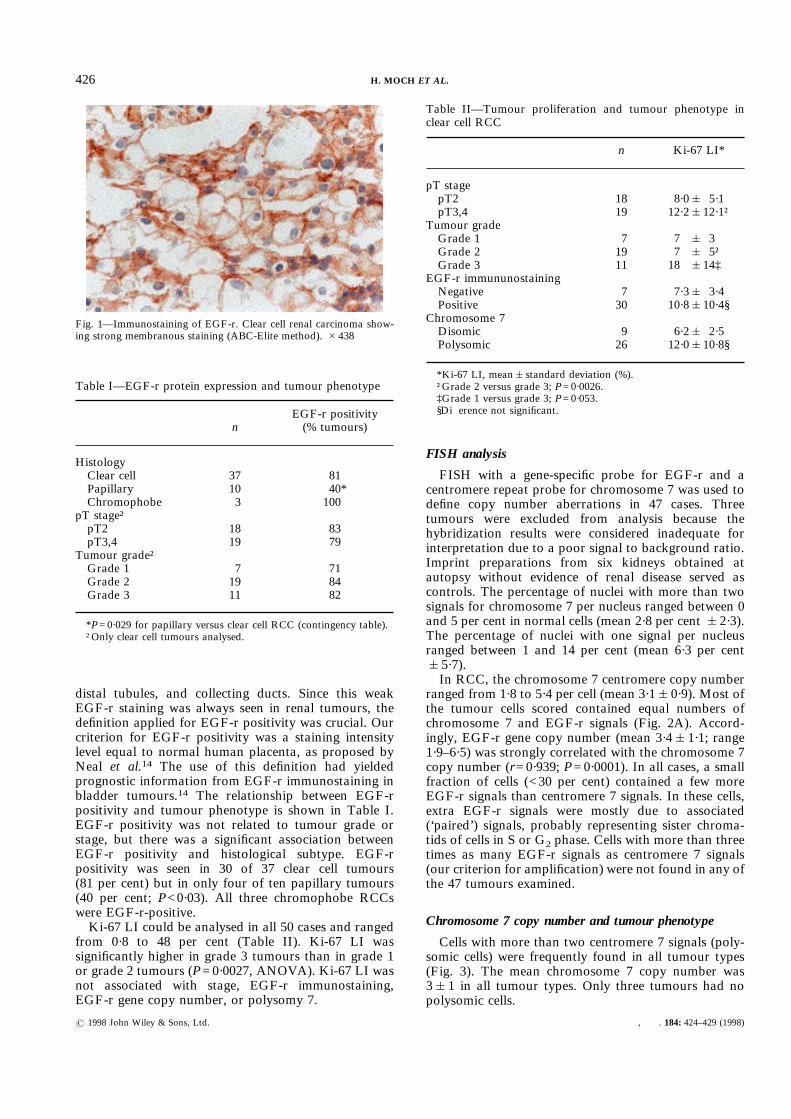
distal tubules, and collecting ducts. Since this weakEGF-r staining was always seen in renal tumours, thedefinition applied for EGF-r positivity was crucial. Ourcriterion for EGF-r positivity was a staining intensitylevel equal to normal human placenta, as proposed byNeal et al.14 The use of this definition had yieldedprognostic information from EGF-r immunostaining inbladder tumours.14 The relationship between EGF-rpositivity and tumour phenotype is shown in Table I.EGF-r positivity was not related to tumour grade orstage, but there was a significant association betweenEGF-r positivity and histological subtype. EGF-rpositivity was seen in 30 of 37 clear cell tumours(81 per cent) but in only four of ten papillary tumours(40 per cent; P<0·03). All three chromophobe RCCswere EGF-r-positive.Ki-67 LI could be analysed in all 50 cases and ranged
from 0·8 to 48 per cent (Table II). Ki-67 LI wassignificantly higher in grade 3 tumours than in grade 1or grade 2 tumours (P=0·0027, ANOVA). Ki-67 LI wasnot associated with stage, EGF-r immunostaining,EGF-r gene copy number, or polysomy 7.
FISH analysis
FISH with a gene-specific probe for EGF-r and acentromere repeat probe for chromosome 7 was used todefine copy number aberrations in 47 cases. Threetumours were excluded from analysis because thehybridization results were considered inadequate forinterpretation due to a poor signal to background ratio.Imprint preparations from six kidneys obtained atautopsy without evidence of renal disease served ascontrols. The percentage of nuclei with more than twosignals for chromosome 7 per nucleus ranged between 0and 5 per cent in normal cells (mean 2·8 per cent &2·3).The percentage of nuclei with one signal per nucleusranged between 1 and 14 per cent (mean 6·3 per cent&5·7).In RCC, the chromosome 7 centromere copy number
ranged from 1·8 to 5·4 per cell (mean 3·1&0·9). Most ofthe tumour cells scored contained equal numbers ofchromosome 7 and EGF-r signals (Fig. 2A). Accord-ingly, EGF-r gene copy number (mean 3·4&1·1; range1·9–6·5) was strongly correlated with the chromosome 7copy number (r=0·939; P=0·0001). In all cases, a smallfraction of cells (<30 per cent) contained a few moreEGF-r signals than centromere 7 signals. In these cells,extra EGF-r signals were mostly due to associated(‘paired’) signals, probably representing sister chroma-tids of cells in S or G2 phase. Cells with more than threetimes as many EGF-r signals as centromere 7 signals(our criterion for amplification) were not found in any ofthe 47 tumours examined.
Chromosome 7 copy number and tumour phenotype
Cells with more than two centromere 7 signals (poly-somic cells) were frequently found in all tumour types(Fig. 3). The mean chromosome 7 copy number was3&1 in all tumour types. Only three tumours had nopolysomic cells.
Fig. 1—Immunostaining of EGF-r. Clear cell renal carcinoma show-ing strong membranous staining (ABC-Elite method). #438
Table I—EGF-r protein expression and tumour phenotype
nEGF-r positivity(% tumours)
HistologyClear cell 37 81Papillary 10 40*Chromophobe 3 100
pT stage†pT2 18 83pT3,4 19 79
Tumour grade†Grade 1 7 71Grade 2 19 84Grade 3 11 82
*P=0·029 for papillary versus clear cell RCC (contingency table).†Only clear cell tumours analysed.
Table II—Tumour proliferation and tumour phenotype inclear cell RCC
n Ki-67 LI*
pT stagepT2 18 8·0& 5·1pT3,4 19 12·2&12·1†
Tumour gradeGrade 1 7 7 & 3Grade 2 19 7 & 5†Grade 3 11 18 &14‡
EGF-r immununostainingNegative 7 7·3& 3·4Positive 30 10·8&10·4§
Chromosome 7Disomic 9 6·2& 2·5Polysomic 26 12·0&10·8§
*Ki-67 LI, mean&standard deviation (%).†Grade 2 versus grade 3; P=0·0026.‡Grade 1 versus grade 3; P=0·053.§Difference not significant.
426 H. MOCH ET AL.
? 1998 John Wiley & Sons, Ltd. , . 184: 424–429 (1998)
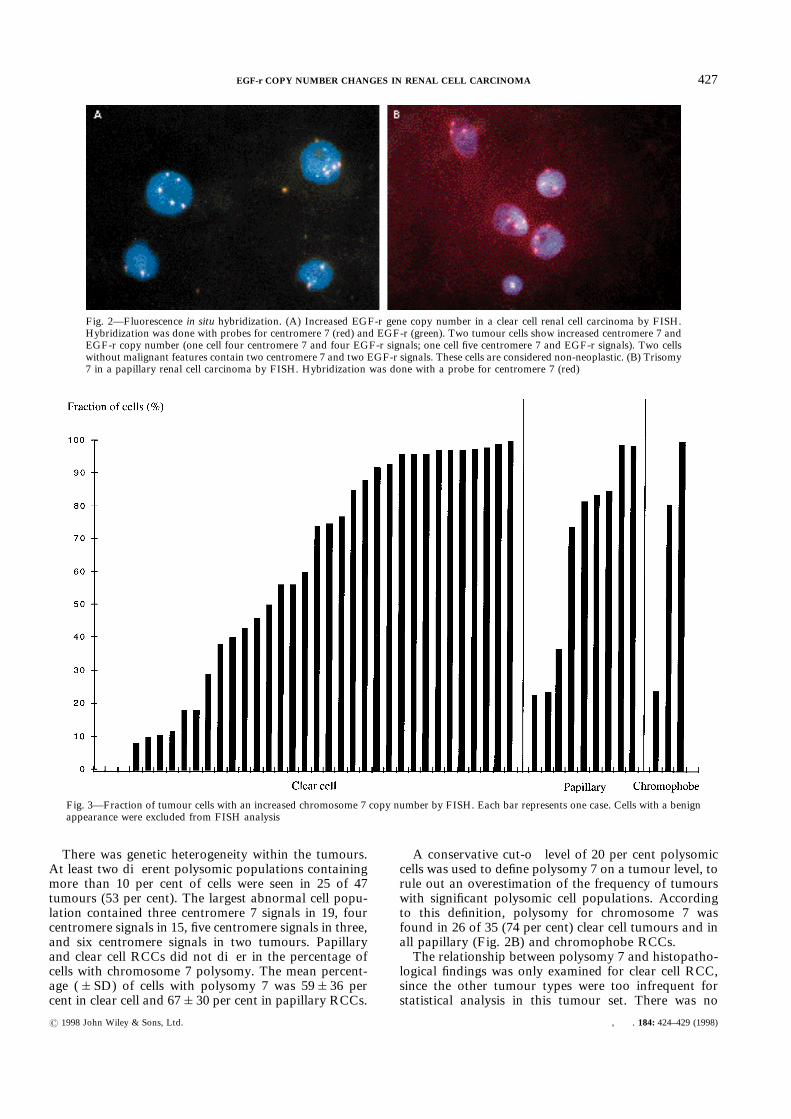
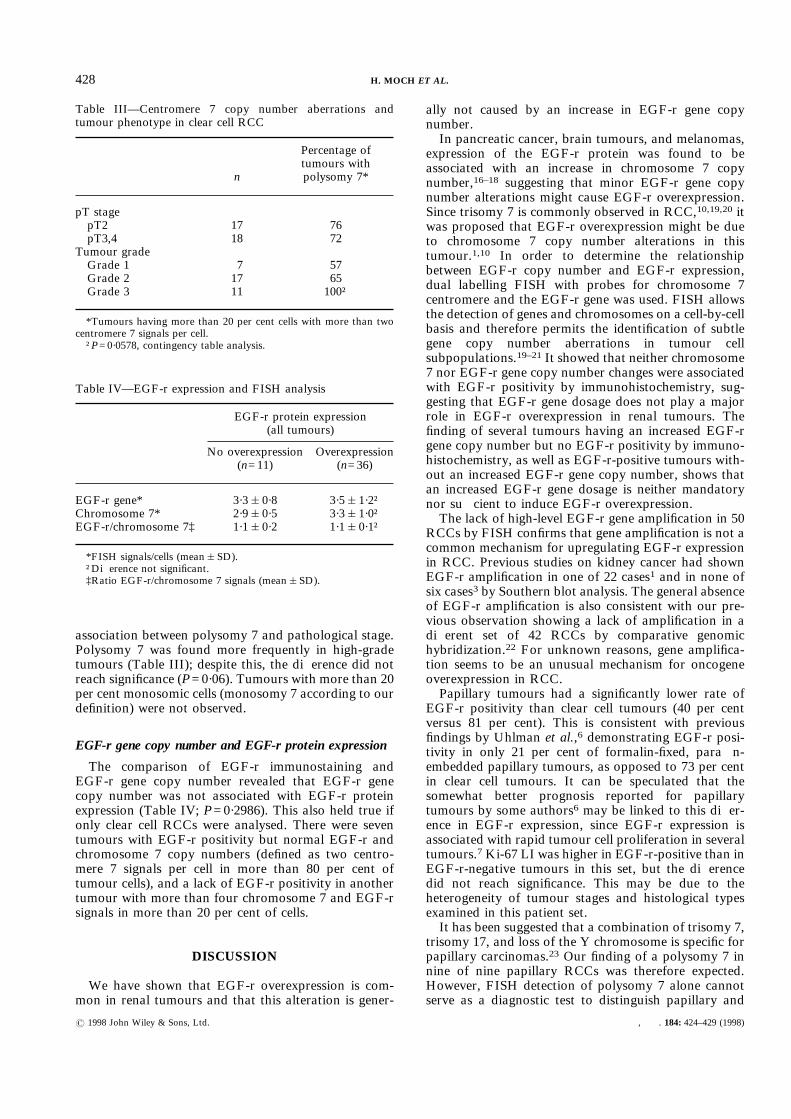
There was genetic heterogeneity within the tumours.At least two different polysomic populations containingmore than 10 per cent of cells were seen in 25 of 47tumours (53 per cent). The largest abnormal cell popu-lation contained three centromere 7 signals in 19, fourcentromere signals in 15, five centromere signals in three,and six centromere signals in two tumours. Papillaryand clear cell RCCs did not differ in the percentage ofcells with chromosome 7 polysomy. The mean percent-age (&SD) of cells with polysomy 7 was 59&36 percent in clear cell and 67&30 per cent in papillary RCCs.
A conservative cut-off level of 20 per cent polysomiccells was used to define polysomy 7 on a tumour level, torule out an overestimation of the frequency of tumourswith significant polysomic cell populations. Accordingto this definition, polysomy for chromosome 7 wasfound in 26 of 35 (74 per cent) clear cell tumours and inall papillary (Fig. 2B) and chromophobe RCCs.The relationship between polysomy 7 and histopatho-
logical findings was only examined for clear cell RCC,since the other tumour types were too infrequent forstatistical analysis in this tumour set. There was no
Fig. 2—Fluorescence in situ hybridization. (A) Increased EGF-r gene copy number in a clear cell renal cell carcinoma by FISH.Hybridization was done with probes for centromere 7 (red) and EGF-r (green). Two tumour cells show increased centromere 7 andEGF-r copy number (one cell four centromere 7 and four EGF-r signals; one cell five centromere 7 and EGF-r signals). Two cellswithout malignant features contain two centromere 7 and two EGF-r signals. These cells are considered non-neoplastic. (B) Trisomy7 in a papillary renal cell carcinoma by FISH. Hybridization was done with a probe for centromere 7 (red)
Fig. 3—Fraction of tumour cells with an increased chromosome 7 copy number by FISH. Each bar represents one case. Cells with a benignappearance were excluded from FISH analysis
427EGF-r COPY NUMBER CHANGES IN RENAL CELL CARCINOMA
? 1998 John Wiley & Sons, Ltd. , . 184: 424–429 (1998)
association between polysomy 7 and pathological stage.Polysomy 7 was found more frequently in high-gradetumours (Table III); despite this, the difference did notreach significance (P=0·06). Tumours with more than 20per cent monosomic cells (monosomy 7 according to ourdefinition) were not observed.
EGF-r gene copy number and EGF-r protein expression
The comparison of EGF-r immunostaining andEGF-r gene copy number revealed that EGF-r genecopy number was not associated with EGF-r proteinexpression (Table IV; P=0·2986). This also held true ifonly clear cell RCCs were analysed. There were seventumours with EGF-r positivity but normal EGF-r andchromosome 7 copy numbers (defined as two centro-mere 7 signals per cell in more than 80 per cent oftumour cells), and a lack of EGF-r positivity in anothertumour with more than four chromosome 7 and EGF-rsignals in more than 20 per cent of cells.
DISCUSSION
We have shown that EGF-r overexpression is com-mon in renal tumours and that this alteration is gener-
ally not caused by an increase in EGF-r gene copynumber.In pancreatic cancer, brain tumours, and melanomas,
expression of the EGF-r protein was found to beassociated with an increase in chromosome 7 copynumber,16–18 suggesting that minor EGF-r gene copynumber alterations might cause EGF-r overexpression.Since trisomy 7 is commonly observed in RCC,10,19,20 itwas proposed that EGF-r overexpression might be dueto chromosome 7 copy number alterations in thistumour.1,10 In order to determine the relationshipbetween EGF-r copy number and EGF-r expression,dual labelling FISH with probes for chromosome 7centromere and the EGF-r gene was used. FISH allowsthe detection of genes and chromosomes on a cell-by-cellbasis and therefore permits the identification of subtlegene copy number aberrations in tumour cellsubpopulations.19–21 It showed that neither chromosome7 nor EGF-r gene copy number changes were associatedwith EGF-r positivity by immunohistochemistry, sug-gesting that EGF-r gene dosage does not play a majorrole in EGF-r overexpression in renal tumours. Thefinding of several tumours having an increased EGF-rgene copy number but no EGF-r positivity by immuno-histochemistry, as well as EGF-r-positive tumours with-out an increased EGF-r gene copy number, shows thatan increased EGF-r gene dosage is neither mandatorynor sufficient to induce EGF-r overexpression.The lack of high-level EGF-r gene amplification in 50
RCCs by FISH confirms that gene amplification is not acommon mechanism for upregulating EGF-r expressionin RCC. Previous studies on kidney cancer had shownEGF-r amplification in one of 22 cases1 and in none ofsix cases3 by Southern blot analysis. The general absenceof EGF-r amplification is also consistent with our pre-vious observation showing a lack of amplification in adifferent set of 42 RCCs by comparative genomichybridization.22 For unknown reasons, gene amplifica-tion seems to be an unusual mechanism for oncogeneoverexpression in RCC.Papillary tumours had a significantly lower rate of
EGF-r positivity than clear cell tumours (40 per centversus 81 per cent). This is consistent with previousfindings by Uhlman et al.,6 demonstrating EGF-r posi-tivity in only 21 per cent of formalin-fixed, paraffin-embedded papillary tumours, as opposed to 73 per centin clear cell tumours. It can be speculated that thesomewhat better prognosis reported for papillarytumours by some authors6 may be linked to this differ-ence in EGF-r expression, since EGF-r expression isassociated with rapid tumour cell proliferation in severaltumours.7 Ki-67 LI was higher in EGF-r-positive than inEGF-r-negative tumours in this set, but the differencedid not reach significance. This may be due to theheterogeneity of tumour stages and histological typesexamined in this patient set.It has been suggested that a combination of trisomy 7,
trisomy 17, and loss of the Y chromosome is specific forpapillary carcinomas.23 Our finding of a polysomy 7 innine of nine papillary RCCs was therefore expected.However, FISH detection of polysomy 7 alone cannotserve as a diagnostic test to distinguish papillary and
Table III—Centromere 7 copy number aberrations andtumour phenotype in clear cell RCC
n
Percentage oftumours withpolysomy 7*
pT stagepT2 17 76pT3,4 18 72
Tumour gradeGrade 1 7 57Grade 2 17 65Grade 3 11 100†
*Tumours having more than 20 per cent cells with more than twocentromere 7 signals per cell.†P=0·0578, contingency table analysis.
Table IV—EGF-r expression and FISH analysis
EGF-r protein expression(all tumours)
No overexpression(n=11)
Overexpression(n=36)
EGF-r gene* 3·3&0·8 3·5&1·2†Chromosome 7* 2·9&0·5 3·3&1·0†EGF-r/chromosome 7‡ 1·1&0·2 1·1&0·1†
*FISH signals/cells (mean&SD).†Difference not significant.‡Ratio EGF-r/chromosome 7 signals (mean&SD).
428 H. MOCH ET AL.
? 1998 John Wiley & Sons, Ltd. , . 184: 424–429 (1998)

clear cell tumours, since polysomy 7 was also present inthe majority of clear cell RCCs. The frequency ofpolysomy 7 in clear cell tumours (78 per cent) wascomparable to the results of previous FISH studies (68per cent),24,25 but was significantly higher than incytogenetic studies (27–67 per cent),10,19,26 despite aconservative cut-off level (20 per cent) for the definitionof a polysomic tumour in our study. This discrepancymight be explained by the considerable intratumourheterogeneity of the chromosome 7 count, almostalways including disomic cells with only two signals. It ispossible that conditions of culture before harvest permitselection for disomic tumour cell subgroups.Numerical chromosome 7 aberrations have been
associated with a more aggressive tumour phenotype inbladder cancer.26 Our results showed no correlationbetween chromosome 7 copy number and tumour stage,arguing against a role of chromosome 7 polysomy forthe development of a particularly aggressive tumourbehaviour in clear cell RCC. However, there was a trendtowards a higher rate of polysomy 7 in high-gradetumours. Recently Beck et al.25 found a strong associ-ation between chromosome 7 gains and DNA aneu-ploidy in RCC. It appears likely, therefore, thatpolysomy 7 is due to a general DNA aneuploidy in mostcases, involving many chromosomes. A frequent associ-ation of polysomy 7 with gains of other chromosomeswould also be consistent with our observation of arelatively low frequency of chromosome 7 gains (15 percent) by comparative genomic hybridization,22 wherethe copy number of chromosome 7 is measured relativeto the average of all other chromosomes.In summary, our data confirm that polysomy 7 is a
frequent event in renal cancer, occurring in all subtypesof RCC. Neither polysomy 7 nor EGF-r copy numberincrease, however, was associated with EGF-r over-expression, indicating that low-level gene dosagechanges play little part in EGF-r overexpression inRCC.
ACKNOWLEDGEMENTS
We thank Ursula Dürmüller, Rita Epper, CaroleFlesch, Hedvika Novotny, and the staff of the Institutefor Pathology, University of Basel for their technicalsupport. This work was supported by DeutscheForschungsgemeinschaft (Mo 625/1), SchweizerischeKrebsliga (SKL 291-2-1996), and the Swiss NationalScience Foundation (3200-043969.95).
REFERENCES1. Ishikawa J, Maeda S, Umezu K, Sugiyama T, Kamidono S. Amplification
and overexpression of the epidermal growth factor receptor gene in humanrenal-cell carcinoma. Int J Cancer 1990; 45: 1018–1021.
2. Sargent ER, Gomella LG, Belldegrun A, Linehan WM, Kasid A. Epidermalgrowth factor receptor gene expression in normal human kidney and renalcell carcinoma. J Urol 1989; 142: 1364–1368.
3. Weidner U, Peter S, Strohmeyer T, Hussnatter R, Ackermann R, Sies H.Inverse relationship of epidermal growth factor receptor and HER2/neugene expression in human renal cell carcinoma. Cancer Res 1990; 50:4504–4509.
4. Lager DJ, Slagel DD, Palechek PL. The expression of epidermal growthfactor receptor and transforming growth factor alpha in renal cell carci-noma. Mod Pathol 1994; 7: 544–548.
5. Yoshida K, Tosaka A, Takeuchi S, Kobayashi N. Epidermal growthfactor receptor content in human renal cell carcinomas. Cancer 1994; 73:1913–1918.
6. Uhlman D, Nguyen P, Manivel J, et al. Epidermal growth factor receptorand transforming growth factor alpha expression in papillary and non-papillary renal cell carcinoma: correlation with metastatic behavior andprognosis. Clin Cancer Res 1995; 1: 913–920.
7. Sauter G, Haley J, Chew K, et al. Epidermal-growth-factor-receptorexpression is associated with rapid tumor proliferation in bladder cancer. IntJ Cancer 1994; 57: 508–514.
8. Ekstrand A, James C, Cavanee W, Seliger B, Petterson R, Collins V. Genesfor epidermal-growth-factor receptor, transforming growth factor alpha,and epidermal growth factor and their expression in human gliomas in vivo.Cancer Res 1991; 51: 2164–2172.
9. Sauter G, Maeda T, Waldman FM, Davis RL, Feuerstein BG. Patterns ofepidermal growth factor receptor amplification in malignant gliomas. Am JPathol 1996; 148: 1047–1053.
10. van den Berg E, van der Hout AH, Oosterhuis JW, et al. Cytogeneticanalysis of epithelial renal-cell tumors: relationship with a new histopatho-logical classification. Int J Cancer 1993; 55: 223–227.
11. Weiss L, Gelb A, Medeiros L. Adult renal epithelial neoplasms. Am J ClinPathol 1995; 103: 624–635.
12. Thoenes W, Stoerkel S, Rumpelt H. Histopathology and classification ofrenal cell tumors (adenomas, oncocytomas, and carcinomas): the basiccytological and histopathological elements and their use for diagnostics.Pathol Res Pract 1986; 181: 125–143.
13. UICC. TNM Classification of Malignant Tumours. 4th edn. 2nd revision.Berlin: Springer-Verlag, 1992.
14. Neal D, Sharples L, Smith K, Fenelly J, Hall R, Harris A. The epidermal-growth-factor receptor and the prognosis of bladder cancer. Cancer 1990;65: 1619–1625.
15. Gerdes J, Lemke H, Baisch H. Cell cycle analysis of a cell proliferation-associated human nuclear antigen defined by the monoclonal antibodyKi-67. J Immunol 1984; 133: 1710–1715.
16. Koprowski H, Herlyn M, Balaban G, Parmiter A, Ross A, Nowell P.Expression of the receptor for epidermal growth factor correlates withincreased dosage of chromosome 7 in malignant melanoma. Somat Cell MolGenet 1985; 11: 297–302.
17. Korc M, Meltzer P, Trent J. Enhanced expression of epidermal growthfactor receptor correlates with alterations of chromosome 7 in humanpancreatic cancer. Proc Natl Acad Sci (Wash) 1986; 85: 1571–1577.
18. Henn W, Blin N, Zang K. Polysomy of chromosome 7 is correlated withoverexpression of the erbB oncogene in human glioblastoma cell lines. HumGenet 1986; 74: 104–106.
19. Kallioniemi O, Kalioniemi A, Kurisu W, et al. C-erbB-2 oncogene amplifi-cation in breast cancer analyzed by fluorescence in situ hybridization. ProcNatl Acad Sci USA 1992; 89: 5321–5325.
20. Matsumura K, Kallioniemi O, Kallioniemi A, et al. Deletion of chromo-some 17p loci in breast cancer cells detected by fluorescence in situhybridization. Cancer Res 1992; 52: 3474–3477.
21. Sauter G, Carroll P, Moch H, et al. c-myc copy number gains in bladdercancer detected by fluorescence in situ hybridization. Am J Pathol 1995; 146:1131–1139.
22. Moch H, Presti JC Jr, Sauter G, et al. Genetic aberrations detected bycomparative genomic hybridization are associated with clinical outcome inrenal cell carcinoma. Cancer Res 1996; 56: 27–30.
23. Kovacs G, Fuzesi L, Emanual A, Kung HF. Cytogenetics of papillary renalcell tumors. Genes Chromosom Cancer 1991; 3: 249–255.
24. El-Naggar A, Hurr K, Tu Z, Tucker S, Swanson D, Hsu P. Interphasecytogenetics of renal cortical neoplasms. Am J Clin Pathol 1995; 104:141–149.
25. Beck JL, Hopman AH, Feitz WF, et al. Numerical aberrations of chromo-somes 1 and 7 in renal cell carcinomas as detected by interphase cyto-genetics. J Pathol 1995; 176: 123–135.
26. Waldman F, Carroll P, Kerschmann R, Cohen M, Field F. Centromericcopy number of chromosome 7 is strongly correlated with tumor grade andlabelling index in human bladder cancer. Cancer Res 1991; 51: 3807–3813.
429EGF-r COPY NUMBER CHANGES IN RENAL CELL CARCINOMA
? 1998 John Wiley & Sons, Ltd. , . 184: 424–429 (1998)





























