Embed Size (px)
Citation preview

Department of Botany, Royal Holloway College, University of London, Egham Hill, Egham, Surrey TW20 OEX, England
Effects of Two Triethylamines on the Carotenogenesis of Turkish Lemons and Oranges
L. R. G. VALADON and ROSEMARY S. MUMMERY
Recieved 5 May 1978 . Accepted 5 June 1978
Summary
The two triethylamines, 2-( 4-chlorophenylthio )-triethylamine (CPT A) and 4-chloro-[j'l-(diethylamine)-ethyl]-benzoate (CDEB) were used to investigate carotenoid biosynthesis in Turkish lemons and oranges, C. limonia var. Messina and C. sinensis var. Valencia respectively, as a function of time. In all cases, lycopene not present in controls appeared in large amounts, as did ~ -carotene in the oranges. Since j'l-carotene does not accumulate with either chemical the inhibition may be specii-ic for the steps between lycopene and j'l-carotene in our case. When different concentr~.tions of CDEB were used the color of the flavedo was improved and, with a 0.01 M solution, was still red after 4 weeks. An attempt to study apo-carotenal synthesis with the aid of these triethylamines was unsuccessful. The differences obtained with CDEB on Turkish oranges compared with Californian Citrus fruits suggest that it is not possible to generalize on the use of these compounds although they seem to be lycopene inducers.
Key words: triethylamines, lycopene, carotenoids, bioregulators, Citrus, Rutaceae.
Introduction
Ever since it was first reported that 2-(4-chlorophenylthio)-triethylamine hydrochloride (CPTA) affected carotenoid biosynthesis (COGGINS, HENNING and YOKOYAMA, 1970), a number of workers have used this compound in a variety of tissues to show that lycopene accumulated even though it was not present in the original material (YOKOYAMA, COGGINS, HENNING and DE BENEDICT, 1972; Hsu, POLING and YOKOYAMA, 1974; SIMPSON, RAHMAN, BUCKLE and LEE, 1974; ELAHI, LEE, SIMPSON and CHICHESTER, 1973; VALADON and MUMMERY, 1977). Other di- and triethylamines have been used extensively by YOKOYAMA and co-workers (POLING, Hsu and YOKOYAMA, 1975, 1976; POLING, Hsu, KOEHRN and YOKOYAMA, 1977) on Marsh white seedless grapefruits and they have shown that some of these compounds are lycopene and others e.g. 4-chloro-[p-(diethylamine)-ethyl]-benzoate (CDEB) are p-carotene inducers as well. In the deeper-pigmented citrus fruits, such as oranges, the degree of response due to these ethylaminc compounds is much less than in the lighter-coloured fruits such as lemons. Furthermore, it is known that varietal
z. Pjlanzenphysiol. Bd. 90. s. 11-19. 1978.

12 L. R. G. VALADON and ROSEMARY S. MUMMERY
differences also play an important role in response to these compounds (YOKOYAMA, personal communication).
The present study was carried out on Turkish oranges and lemons, using CPTA and CDEB to determine whether carotenogenesis in these fruits was affected in the same way as in Californian ones. Also YOKOYAMA and co-workers (YOKOYAMA et a!., 1972; Hsu et a!., 1974; POLING et a!., 1975, 1976, 1977) have been mainly concerned with carotenes and have not identified xanthophylls, even though these accounted for approximately one third total carotenoids in their grapefruits. Since xanthophylls are on the same biosynthetic pathways as carotenes one or more xanthophylls may also be affected. We therefore identified all the carotenoids present in our fruits which, as they contained more xanthophylls than carotenes, showed varietal differences.
It is well known that as fruits mature, different carotenoids in various amounts appear which may further elucidate carotenoid biosynthesis (V ALADON and MUMMERY, 1977). Carotenoids of orange and lemon flavedos of increasing maturity were investigated in order to attempt to observe biogenesis of apocarotenals from C40
carotenoids. Further, different concentrations of CDEB were used with a view to improving the colour of orange flavedos; carotenoid accumulation was also studied as a function of time.
Materials and Methods
Fruit samples
Turkish lemons (Citrus limonia Osbeck var. Messina = C. medica var. Limon LINN.) and Turkish oranges (C. sinensis Osbeck var. Valencia = C. aurantium var. Sinensis LINN.) were harvested by Mr. A. K. CETIN at different times from trees located in Izmir, Turkey. The fruits were then air-freighted, taking about 4 days to reach the laboratory.
Treatment with CPT A
The fruits were treated as previously described (YOKOYAMA et a!., 1972) but the wetting agent was Triton X-I00.
Treatment with CDEB
As previously described (POLING et a!., 1976).
Extraction, isolation and quantitative determination of pigment
The carotenoids were isolated and separated as previously described (VALADON and MUMMERY, 1975).
Carotenoids having epoxy groups were characterised by the modified cone. HCI-ether test (]UNGALWALA and CAMA, 1962).
The structural identity of individual carotenoids was established by comparison with authentic samples using various chromatographic methods as already described (VALADON and MUMMERY, 1975, 1977), and visible, UV and IR spectroscopy. The apo-compounds were further identified by reduction with LiAlH4 when the corresponding mono-ol was obtained (CURL, 1962).
The concentration of individual carotenoids was determined by measuring Emax and
comparing it with known E~~~ for pure pigments (DAVIES, 1965). For those pigments whose
E r~~ were unknown, Amax was assumed to be 2500 (VALADON and MUMMERY, 1975). All
Z. Pjlanzenphysiol. Bd. 90. S. 11-19. 1978.

Carotenogenesis in lemons and oranges 13
results were calculated on a per gram dry weight basis and each carotenoid was estimated as the percentage of total carotenoids.
'I)-Carotene was identified tentatively as it had absorption of 378, 400 and 424 nm in n-hexane and was found between phytofluene and a-carotene (YOKOYAMA and WHITE, 1967).
All the experiments were repeated at least three times. The trend was always the same but amounts varied depending on the maturity of the oranges and lemons with which we started. We had controls of fruits (Table 3) stored for the same period as the treated ones. With both lemons and oranges, these controls contained very similar carotenoids to controls at 0 days but total carotenoids increased over the first 5 weeks and thereaiter decreased. These controls never contained lycopene and only small amounts of ~-carotene.
Results and Discussion
Fruits not treated with CPTA and CDEB were yellow (oranges) or pale yellow (lemons) whilst those so treated were red, red-orange or red-yellow depending on various factors. The test compounds did not penetrate into the interior of the fruits and therefore only the flavedo changed colour. At the highest concentration of CDEB used (0.5 M), the oranges were a dull orange after 3 weeks i.e. the red colour had completely disappeared and parts had become brown, whilst at the lowest concentration (0.01 M) the oranges were still red-orange after 5 weeks and at 3 weeks were bright red.
Tables 1 and 2 give the results obtained when lemons and oranges were treated with 5000 ppm CPTA over periods of 3 and 12 weeks respectively. Lycopene was not present in the untreated fruits but this red carotene accumulated in both the treated fruits as is already well known (YOKOYAMA et aI., 1972). In lemons there were also increased syntheses of phytofluene, ~ -carotene, y-carotene but not of neurosporene which was never observed in our studies. In the case of oranges the increase in ~-carotene was as spectacular as that in lycopene, whilst the syntheses of phytofluene and y-carotene were slightly increased (Table 2). There were obvious differences between lemons and oranges even though in the controls neither possessed y-carotene and lycopene which appeared in the ePTA-treated fruits. Since we observed increases in certain carotenes, decreases would be expected in other compounds. With lemons there was a significant decrease in p-apo-8' -carotenal (Cao), and in p-zeacarotene, violaxanthin, zeaxanthin, lutein, mutatochrome and antheraxanthin (all C40 compounds). Of these p-zeacarotene, lutein and mutatochrome were identified in the controls but were absent in the CPTA-treated lemons. Carotenoids found in the latter but not in the former were: citranaxanthin (Cas), and flavochrome, aurochrome and auroxanthin (all C40 compounds). Although generally CPTA-treated oranges were similar to treated lemons, there were some differences. There was an increase in total carotenoids up to 5 weeks and then a decrease to a minimum of 392.9 fig g-l after 12 weeks which was only about half that of untreated controls (698.7 fig g-l). Once again, y-carotene, absent in the untreated oranges, appeared in the CPTA-treated fruits but only after 5 weeks. Though sintaxanthin (C3l) did not appear in the controls it increased up to the seventh week
Z. P/lanzenphysiol. Bd. 90. S. 11-19. 1978.

14 L. R. G. VALADON and ROSEMARY S. MUMMERY
and then disappeared. This is in direct contrast to observations with CPTA-treated lemons, where sintaxanthin was identified in the controls and then disappeared after two weeks. The increase in lycopene (= 25 0/0) was never as great as in lemons where it was about 80010; and whereas after three weeks total carotenoids of CPTA-treated lemons were approximately ten times those of controls, values for CPTA-treated oranges had not even doubled compared to controls.
It is well known that as fruits mature there is an increase in furanoid compounds such as mutatochrome and aurochrome at the expense of 5,6- and 5,6 : 5', 6' -epoxy compounds. Further there is also an increase in cis-compounds which are not normally found in younger fruits (VALADON and MUMMERY, 1977). This is clearly shown in the present study in which diepoxy-p-carotene and violaxanthin decrease and aurochrome and auroxanthin increase. YOKOYAMA et al. (1972) concluded that the pattern of large accumulation of lycopene and to a smaller extent y-carotene ascribes to CPTA an effect due to inhibition of cyclases or feedback inhibition. YOKOYAMA (personal communication) considers that this is rather conjectural and that other factors may be involved.
In their latest papers YOKOYAMA and co-workers (POLING et aI., 1976, 1977) have observed that certain triethylamines not only increased lycopene synthesis but also that of a- and p-carotene. One such compound is CDEB, which was made available to us and with which we investigated carotenoid synthesis in Turkish oranges. Three concentrations of CDEB were used over varying periods of time. The oranges were not as mature as in the previous experiment and contained only 139 Ilg g-l total carotenoids compared with the previous 698 Ilg g-t, so that variations would be expected (YOKOYAMA et aI., 1972). The results (Table 3) show the same general pattern as with CPTA in that lycopene, although not present in the controls, increased significantly in CDEB-treated oranges, but there was no y-carotene and ~-carotene accumulation. There was however limited a- and p-carotene accumulation at the lowest concentration (0.01 M CDEB) after 5 weeks. Concentrations of 0.5 M and 0.1 M were not very useful in producing good-coloured oranges and after 3 and 4 weeks the fruits were a dull orange containing only 69 Ilg g-l carotenoids and a fading-red containing only 160 j.lg g-l respectively. In the case of the lowest concentration used (0.01 M), the flavedo although fading was still bright red-orange and contained as much total carotenoids as controls. Neurosporene was not identified and in most cases C-carotene and violaxanthin were obtained in both the cis- and trans-forms; there were no increased syntheses of these compounds but in fact a decrease. Furthermore, p-apo-8' (Cao) and p-apo-l0'(C27)-carotenals were observed in the controls and decreased fairly rapidly in the CDEB-treated fruits. A further effect of CDEB was the production of cis-phytofluene which was not found in any of the controls nor in ePTA-treated fruits. The central double-bond of naturally-occurring phytoene - the C40 precursor of carotenoids - in higher plants is cis (DAVIS et aI., 1961), although natural carotenoids are usually in the trans-forms. RAYMUNDO and SIMPSON (1972) working with the tangerine tomato suggested an
z. PJlanzenphysiol. Ed. 90. S. 11-19. 1978.

Carotenogenesis in lemons and oranges
cis-Phytoene ---'- trans-Phytoene ....,..-
~ ~ cis-Phytofluene ---'- trans-Phytofluene ....,..-
~ ~ cis-~ -Carotene ---'- trans-~ -Carotene ....,..-
~ ~ cis-N eurosporene ---'- trans-N eurosporene ....,..-
-I- ~ cis-Violaxanthin ---'- trans-Violaxanthin ....,..-
~ ~ cis-N eoxanthin ---'- trans-N eoxanthin ....,..-
Fig. 1: Possible pathway of cis- and trans-isomers of carotenoids higher plants. -------- indicates several steps.
15
inter conversion of the twO forms as in Fig. 1. However, the presence of large amounts of cis-compounds as the fruits mature has yet to be explained.
Our results with CDEB-treated fruits were therefore very different to those of POLING et a!. (1976) who found increased a- and p-carotene syntheses. They further showed (1977) that the time factor was very important for these p-carotene inducers. These compounds caused a very rapid initial accumulation of lycopene, which was later converted to p-carotene in Marsh white seedless grapefruit. This was not the case with Turkish oranges, as there was very little if any p-carotene synthesis even after five weeks, and hence no conversion of lycopene to p-carotene. POLING et a!. (1977) found that the compound which yielded the best result as a p-carotene inducer was the hexanoate of 2-diethylamine-ethyl ester and it would be interesting to ascertain whether this compound can induce p-carotene synthesis in Turkish oranges. It is well known that different varieties respond differently to the same compounds. This has been shown quite clearly here, and it is not always possible to generalize when using one variety or one genus. Nicotine induces lycopene synthesis in many tissues but not in mature grapefruits nor in navel oranges (YOKOYAMA et a!., 1972).
The biosynthetic pathway of C30 and C33 carotenoids is relatively unknown but from chemical studies it appears that some of them are formed from p-carotene according to the pathway proposed by LIAAEN-JENSEN (1971).
Table 1 shows a number of carotenoids were decreasing whilst lycopene was increasing, the largest decreases being in p-apo-8' -carotene, violaxanthin and izeaxanthin. In Table 2 the largest decreases were in cryptoxanthin and violaxanthin, whilst in Table 3 there was a small general decrease in a number of carotenoids and not in any particular one. If the biosynthetic pathway is the same as the chemical
Z. P/lanzenphysiol. Ed. 90. S. 11-19. 1978.
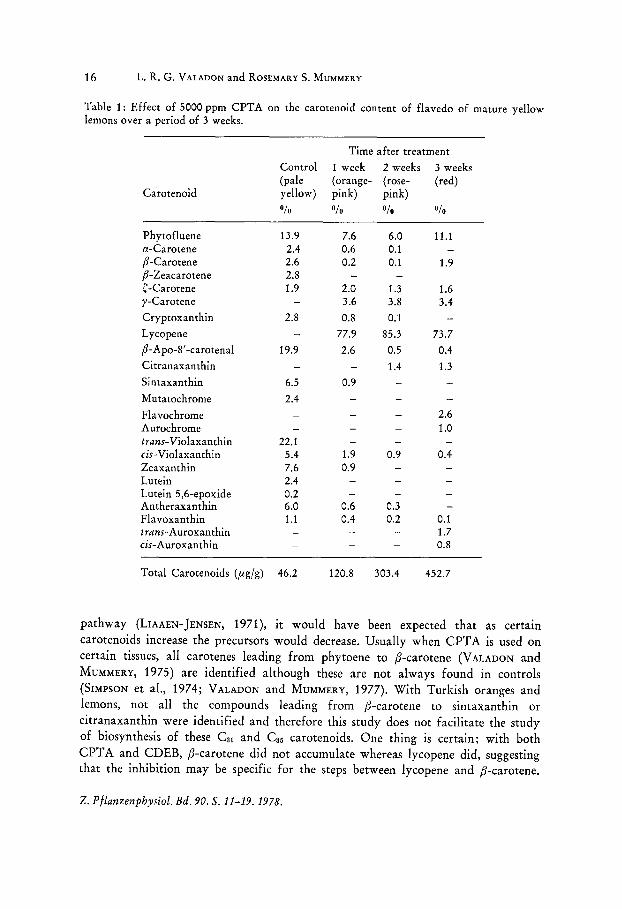
16 L. R. G. VALADON and ROSEMARY S. MUMMERY
Table 1: Effect of 5000 ppm CPTA on the carotenoid content of flavedo of mature yellow lemons over a period of 3 weeks.
Time after treatment
Control 1 week 2 weeks 3 weeks (pale (orange- (rose- (red)
Carotenoid yellow) pink) pink) 0/0 Ofo Ofo 0/0
Phytofluene 13.9 7.6 6.0 11.1 a-Carotene 2.4 0.6 0.1 p-Carotene 2.6 0.2 0.1 1.9 p-Zeacarotene 2.S C-Carotene 1.9 2.0 1.3 1.6 )I-Carotene 3.6 3.S 3.4
Cryptoxanthin 2.S O.S 0.1
Lycopene 77.9 85.3 73.7
p-Apo-S' -carotenal 19.9 2.6 0.5 0.4
Citranaxanthin 1.4 1.3
Sintaxanthin 6.5 0.9
Mutatochrome 2.4
Fla vochrome 2.6 Aurochrome 1.0 t,-ans-Violaxanthin 22.1 cis-Violaxanthin 5.4 1.9 0.9 0.4 Zeaxanthin 7.6 0.9 Lutein 2.4 Lutein 5,6-epoxide 0.2 An theraxan thin 6.0 0.6 0.3 Flavoxanthin 1.1 0.4 0.2 0.1 t rans-Auroxan thin 1.7 cis-Auroxanthin 0.8
Total Carotenoids (fig/g) 46.2 120.S 303.4 452.7
pathway (LIAAEN-JENSEN, 1971), it would have been expected that as certain carotenoids increase the precursors would decrease. Usually when CPTA is used on certain tissues, all carotenes leading from phytoene to p-carotene (V ALADON and MUMMERY, 1975) are identified although these are not always found in controls (SIMPSON et aI., 1974; VALADON and MUMMERY, 1977). With Turkish oranges and lemons, not all the compounds leading from p-carotene to sintaxanthin or citranaxanthin were identified and therefore this study does not facilitate the study of biosynthesis of these Cal and Ca5 carotenoids. One thing is certain; with both CPTA and CDEB, p-carotene did not accumulate whereas lycopene did, suggesting that the inhibition may be specific for the steps between lycopene and p-carotene.
Z. PJlanzenphysiol. Bd. 90. S. 11-19. 1978.
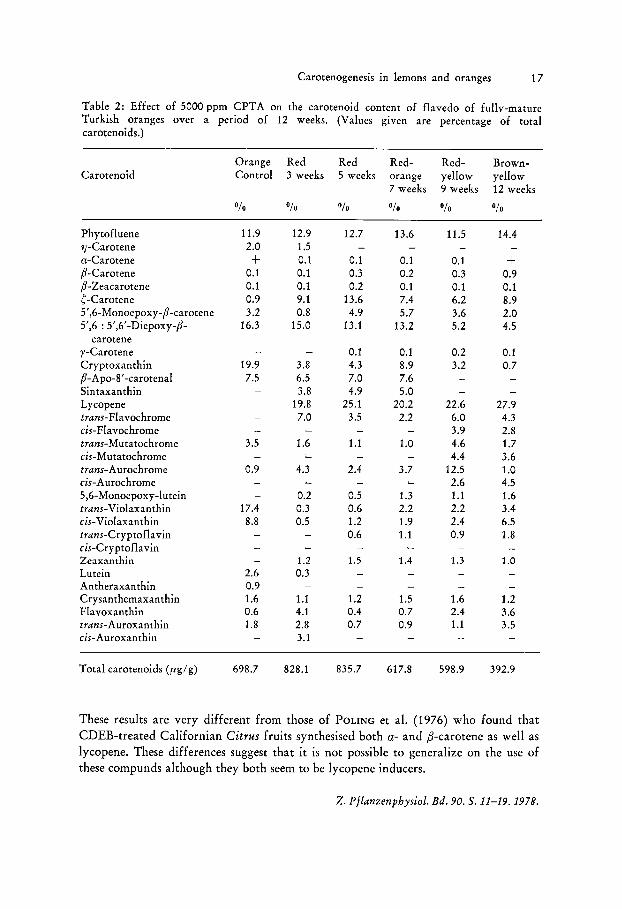
Carotenogenesis in lemons and oranges 17
Table 2: Effect of 5000 ppm CPTA on the carotenoid content of flavedo of fully-mature Turkish oranges over a period of 12 weeks. (Values given are percentage of total carotenoids.)
Carotenoid
Phytofluene 11-Carotene a-Carotene p-Carotene p-Zeacarotene ~-Carotene 5',6-Monoepoxy-p-carotene 5',6 : 5',6' -Diepoxy-p-
carotene y-Carotene Cryptoxanthin p -A po-8' -carotenal Sintaxanthin Lycopene trans-Fla vochrome cis-Fla vochrome trans-Mutatochrome cis-Mutatochrome trans-Aurochrome cis-Aurochrome 5,6-Monoepoxy-Iutein trans-Violaxanthin cis-Violaxanthin trans-Cryptofla vin cis-Cryptoflavin Zeaxanthin Lutein Antheraxanthin Crysanthemaxan thin Flavoxanthin trans-Auroxanthin cis-Auroxanthin
Total carotenoids Cug/g)
Orange Red Red Red-Control 3 weeks 5 weeks orange
7 weeks
11.9 2.0
+ 0.1 0.1 0.9 3.2
16.3
19.9 7.5
3.5
0.9
17.4 8.8
2.6 0.9 1.6 0.6 1.8
698.7
0/0
12.9 1.5 0.1 0.1 0.1 9.1 0.8
15.0
3.8 6.5 3.8
19.8 7.0
1.6
4.3
0.2 0.3 0.5
1.2 0.3
1.1 4.1 2.8 3.1
828.1
0/0
12.7
0.1 0.3 0.2
13.6 4.9
13.1
0.1 4.3 7.0 4.9
25.1 3.5
1.1
2.4
0.5 0.6 1.2 0.6
1.5
1.2 0.4 0.7
835.7
0/0
13.6
0.1 0.2 0.1 7.4 5.7
13.2
0.1 8.9 7.6 5.0
20.2 2.2
1.0
3.7
1.3 2.2 1.9 1.1
1.4
1.5 0.7 0.9
617.8
Redyellow 9 weeks
Ofo
11.5
0.1 0.3 0.1 6.2 3.6 5.2
0.2 3.2
22.6 6.0 3.9 4.6 4.4
12.5 2.6 1.1 2.2 2.4 0.9
1.3
1.6 2.4 1.1
598.9
Brownyellow 12 weeks
Ofo
14.4
+ 0.9 0.1 8.9 2.0 4.5
0.1 0.7
27.9 4.3 2.8 1.7 3.6 1.0 4.5 1.6 3.4 6.5 1.8
1.0
1.2 3.6 3.5
392.9
These results are very different from those of POLING et al. (1976) who found that CDEB-treated Californian Citrus fruits synthesised both a- and p-carotene as well as lycopene. These differences suggest that it is not possible to generalize on the use of these compunds although they both seem to be lycopene inducers.
Z. Pjlanzenphysiol. Bd. 90. S. 11-19. 1978.
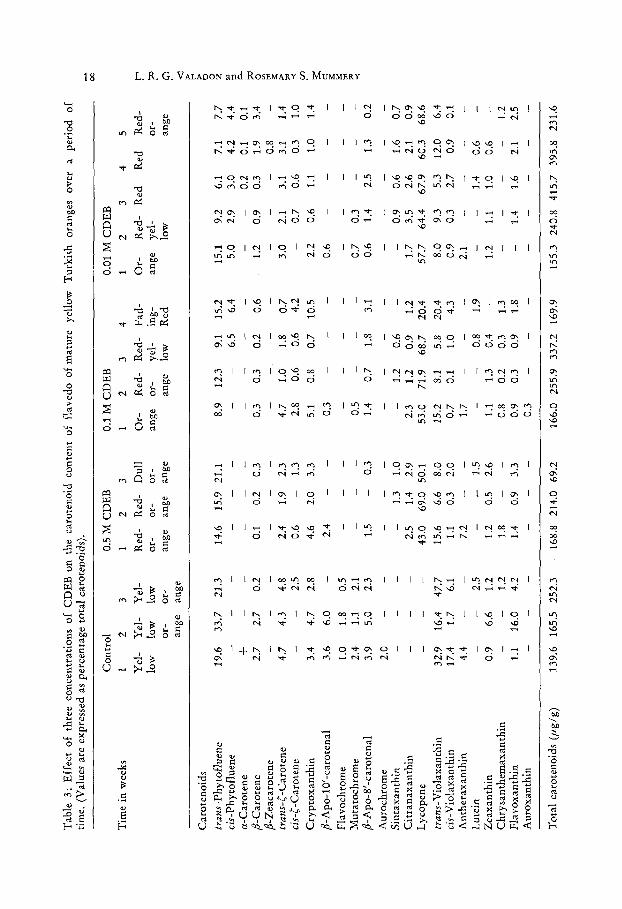
Tab
le 3
: E
ffec
t o
f th
ree
con
cen
trat
ion
s o
f C
DE
B o
n t
he
caro
teno
id c
on
ten
t o
f fl
aved
o o
f m
atu
re
yell
ow
Tu
rkis
h
oran
ges
ov
er
a p
erio
d
of
tim
e. (
Val
ues
are
expr
esse
d as
per
cen
tag
e to
tal
caro
teno
ids)
. >
-'
00
Co
ntr
ol
0.5
M C
DE
B
0.1
M C
DE
B
0.01
M C
DE
B
Tim
e in
wee
ks
2 3
2 3
2 3
4 2
3 4
5
Yel
-Y
el-
Yel
-R
ed-
Red
-D
ull
O
r-R
ed-
Red
-F
ad-
Or-
Red
-R
ed
Red
R
ed-
r lo
w
low
lo
w
or-
or-
or-
ange
o
r-ye
l-in
g-an
ge
yel-
or-
?'
or-
or-
ange
an
ge
ange
an
ge
low
R
ed
low
an
ge
0 an
ge
ange
<
>
....
Car
ote
no
ids
>
0
tra
ns-
Ph
yto
flu
ene
19.6
33
.7
21.3
14
.6
15.9
21
.1
8.9
12.3
9.
1 15
.2
15.1
9.
2 6.
1 7.
1 7.
7 0 z
cis-
Phy
tofl
uene
6.
5 60
4 5.
0 2.
9 3.
0 4.
2 40
4 ., ::;
a-C
aro
ten
e +
0.
2 0.
1 0.
1 0
-
fJ-C
arot
ene
2.7
2.7
0.2
0.1
0.2
0.3
0.3
0.3
0.2
0.6
1.2
0.9
0.3
1.9
304
~
Sl fl
-Zea
caro
tene
0.
8 '" is:
tran
s-C
-Car
ote
ne
4.7
4.3
4.8
204
1.9
2.3
4.7
1.0
1.8
0.7
3.0
2.1
3.1
3.1
104
> "
cis-
C -C
aro
ten
e 2.
5 0.
6 1.
3 2.
S 0.
6 0.
6 4.
2 0.
7 0.
6 0.
3 1.
0 -< C
/l C
ryp
tox
anth
in
304
4.7
2.S
4.6
2.0
3.3
5.1
O.S
0.
7 10
.5
2.2
0.6
1.1
1.0
104
~
fl-A
po
-l 0
' -ca
rote
nal
3.
6 6.
0 20
4 0.
3 0.
6 c:::
is:
F
lavo
chro
me
1.0
1.S
0.5
is: '"
Mu
tato
chro
me
2.4
1.1
2.1
0.5
0.7
0.3
" -< fl
-Ap
o-S
' -ca
rote
nal
3.
9 5.
0 2.
3 1.
5 0.
3 10
4 0.
7 1.
8 3.
1 0.
6 10
4 2.
5 1.
3 0.
2
Aur
ochr
ome
2.0
Sin
tax
anth
in
1.3
1.0
1.2
0.6
0.9
0.6
1.6
0.7
Cit
ran
axan
thin
2.
5 10
4 2.
9 2.
3 1.
2 0.
9 1.
2 1.
7 3.
5 2.
6 2.
1 0.
9 L
ycop
ene
43.0
69
.0
50.1
53
.0
71.9
68
.7
2004
57
.7
6404
67
.9
60.3
68
.6
tran
s-V
iola
xan
thin
32
.9
1604
47
.7
15.6
6.
6 8.
0 15
.2
8.1
5.8
2004
8.
0 9.
3 5.
3 12
.0
604
cis-
Vio
lax
anth
in
1704
1.
7 6.
1 1.
1 0.
3 2.
0 0.
7 0.
1 1.
0 4.
3 0.
9 0.
3 2.
7 0.
9 0.
1 A
nth
erax
anth
in
404
7.2
1.7
2.1
Lut
ein
2.5
1.5
0.8
1.9
104
0.6
Zea
xan
thin
0.
9 6.
6 1.
2 1.
2 0.
5 2.
6 1.
1 1.
3 00
4 1.
2 1.
1 1.
0 0.
6 C
hry
san
them
axan
thin
1.
2 1.
S O
.S
0.2
0.3
1.3
1.2
Fla
vo
xan
thin
1.
1 16
.0
4.2
1.4
0.9
3.3
0.9
0.3
0.9
1.8
1.4
1.6
2.1
2.5
Au
rox
anth
in
0.3
To
tal
caro
teno
ids
(,ug
/ g)
139.
6 16
5.5
252.
3 16
8.8
214.
0 69
.2
166.
0 25
5.9
337.
2 16
9.9
155.
3 24
0.S
41
5.7
395.
S
231.
6

Carotenogenesis in lemons and oranges 19
Acknowledgements
We thank Dr. H. YOKOYAMA, Pasadena, California for a generous gift of CDEB and for general help in these studies. We also thank the firm Amchem Products, Ambler, Pasadena, U.S.A. for a gift of CPTA, and Mr. A. K. CETIN for providing us with the oranges and lemons.
References
COGGINS, Jr. C. W., G. L. HENNING, and H. YOKOYAMA: Lycopene accumulation induced by 2-( 4-chlorophenylthio )-triethylamine hydrochloride. Sc:ence 168, 1589-1591 (1970).
CURL, A. L.: The carotenoids of red bell peppers. Agric. Food Chern. 10, 504-509 (1962). DAVIES, B. H. Analysis of carotenoid pigments. Chemistry and Biochemistry of Plant Pig
ments (T. W. GOODWIN, ed.). Acad. Press, London. 489-532 (1965). DAVIS, J. B., L. M. JACKMAN, P. T. SIDDONS, and B. C. L. WEEDON: The structures of phy
toene, phytofluene, ~-carotene and neurosporene. Proc. Chern. Soc. 261-263 (1961). ELAHI, M., T. H. LEE, K. L. SIMPSON, and C. O. CHICHESTER: Effects of CPTA and cyco
cel on the biosynthesis of carotenoids by Phycomyces blakesleeanus mutants. Phytochemistry 12,1633-1639 (1973).
Hsu, W. J., S. M. POLING, and H. YOKOYAMA: Effects of amines on the carotenogenesis in Blakeslea trispora. Phytochemistry 13, 415-419 (1974).
JUNGALWALA, F. B. and H. R. CAMA: Carotenoids in Delonix regia (Guhl Mohr) flowers. Biochem. J. 85,1-8 (1962).
LIAAEN-JENSEN, S.: Isolation Reactions. Carotenoids, ISLER, O. (Ed.) p. 169. Birkhiiuser Verlag, Basel, 1971.
POLING, S. M., W. J. Hsu, and H. YOKOYAMA: Structure-activity relationships of chemical inducers of carotenoid biosynthesis. Phytochemistry 14, 1933-1938 (1975).
- - - Synthetic regulators of carotenoid biosynthesis in Citrus paradisi. Phytochemistry 15, 1685-1687 (1976).
POLING, S. M., W. J. Hsu, F. J. KOEHRN, and H. YOKOYAMA: Chemical induction of fJ-carotene biosynthesis. Phytochemistry 16, 551-555 (1977).
RAYMUNDO, L. C. and K. L. SIMPSON: The isolation of a poly-cis ~-carotene from the tangerine tomato. Phytochemistry 11, 397-400 (1972).
SIMPSON, D. J., F. M. M. RAHMAN, K. A. BUCKLE, and T. H. LEE: Chemical regulation of plastid development. Aust. J. Plant Physio!. 1, 135-147 (1974).
VALADON, L. R. G. and R. S. MUMMERY: Carotenoids of floral parts and of the spadix of Arum macula tum. Z. Pflanzenphysio!. 75, 88-94 (1975).
- - Carotenoids of lilies and of red pepper. Z. Pflanzenphysiol. 82, 407-416 (1977). YOKOYAMA, H. and M. J. WHITE: Carotenoids in the flavedo of Marsh seedless grapefruits.
J. agr. Food Chern. 15,693-696 (1967). YOKOYAMA, H., C. W. COGGINS Jr., G. L. HENNING, and C. DE BENEDICT: The effect of
CPTA on the formation of carotenoids in citrus. Phytochemistry 11, 1721-1724 (1972).
L. R. G. VALADON, Department of Botany, Royal Holloway College, University of London, Egham Hill, Egham, Surrey TW20 OEX, England.
Z. PJlanzenphysiol. Bd. 90. S. 11-19. 1978.





























