Embed Size (px)
Citation preview

- Effects of temperature on spring bloom dynamics of diatom communities - 599
Effects of temperature on spring bloom dynamics of epilithicdiatom communities in the Gulf of Bothnia
Snoeijs, Pauli J. M.
Department of Ecological Botany, Uppsala University, Box 559, S-751 22 Uppsala, Sweden;Tel. +46 18 182869; Fax +46 18 553419; E-mail [email protected]
Keywords: Baltic Sea; Biomass; Climatic change; Constrain-ed ordination; Microphytobenthos; Thermal discharge.
Nomenclature: Hartley (1986); Williams & Round (1986,1987, 1988); Snoeijs (1989a).
Introduction
Recent reports predict that a man-made global tem-perature increase will occur within the next few decades,as increasing concentrations of carbon dioxide and othergases originating from fossil fuel burning trap outgoingradiation in the lower atmosphere (Gregory 1988; Bach1989; Budyko 1989). High latitude areas, including theBaltic Sea, are considered to be especially sensitive tothis ‘greenhouse effect’ (Bach 1989; Alenius 1989).Cooling water discharge areas provide field conditionsfor research and predictions on the effects of possibleglobal changes on aquatic ecosystems.
Diatoms are amongst the first colonizers of the rockysubstrates in the upper littoral zone of the Baltic Sea thatare left practically bare in spring after the abrading effectof ice-break. This study presents an ecological analysison epilithic diatom spring-bloom communities near theForsmark nuclear power plant (Swedish Baltic Sea coast).During previous investigations in the area, involvingone-year-round studies of diatoms (Snoeijs 1989b) andmacroalgae including diatoms in macroscopic colonies(Snoeijs & Prentice 1989), it was found that the diatomsreact to higher water temperature throughout the year,but especially in spring by species shifts and increasedbiomass. Therefore the diatom spring bloom was herestudied in more detail from 1983 to 1989 to assessfluctuations between years.
This period includes a large natural variation in tem-perature conditions with the 1988 / 1989 winter beingextremely mild, so that hardly any ice occurred in thesouthern Gulf of Bothnia. The samples analysed repre-sent natural temperatures and a range of temperatureanomalies from 1 to 10 °C caused by the high tempera-ture of the cooling water from the power plant. Someenvironmental shifts related to raised temperature alsooccur: normally an ice cover is present in this part of theBaltic Sea during ca. 100 days each year (January -April), but not in sites with water temperature > 0 °C.
Abstract. In order to forecast consequences of climaticchanges for littoral algae in coastal ecosystems, the ef-fects of temperature increase on spring bloom dynamicsof epilithic diatom communities were studied in twoways: (1) Communities were compared from sites re-ceiving different amounts of cooling water dischargefrom a nuclear power plant. (2) Communities from theexceptionally warm spring of 1989 were compared withthose from the same sites in the preceding normal years.The studies were carried out in and around the ForsmarkBiotest Basin, an artificial waterbody that receives brack-ish cooling water from the Forsmark nuclear power planton the Swedish east coast. Species composition andbiomass data from 200 epilithic diatom samples taken inthe period January-May from 1983 to 1989 were ana-lysed.
Multiple regression analysis was used to show theresponses of diatom cover, community diversity andabundances of individual taxa to water temperature andother environmental factors. Constrained ordination(CCA) was used to display the relationship of overallcommunity composition to environment. Increased wa-ter temperature resulted in higher biomass through achain of ecological effects. Strong reduction of ice coverwas crucial in this chain, resulting in reduced loss ofdiatom cells by reduced abrading and higher primaryproduction by higher availability of light, nutrients andsubstrate.
The conclusion is that large blooms of fast-growingepilithic diatoms in large colonies can occupy the nichethat emerges when the ice-free season is prolonged in thenorthern Baltic Sea and areas similar in salinity andclimatic conditions. Dramatic species shifts may only beexpected if the winter ice cover is totally absent.
Journal of Vegetation Science 1: 599-608, 1990© IAVS; Opulus Press Uppsala. Printed in Sweden

Snoeijs, Pauli J. M.600
temperature inevitably vary with time. Other importantfactors affecting microphytobenthic communities in theBaltic Sea are substrate type and salinity. These factorswere held invariable by sampling from a uniform sub-strate and within an area of limited salinity fluctuations.The simultaneous involvement of several environmen-tal factors requires a multivariate approach for dataanalysis to reveal the impact of temperature.
Study area
Forsmark is situated on the east coast of Sweden atthe southern end of the Gulf of Bothnia, ca. 130 kmnorth of Stockholm (60° 25' 80" N, 18° 11' 14" E).Salinity in the area is 0.5 - 0.6 %. The cooling waterfrom the Forsmark nuclear power plant is led through anartificial enclosure, the Forsmark Biotest Basin, beforeit is returned to the Baltic Sea. The basin was construct-ed especially for environmental impact studies (Grimås1979). It has an area of ca. 1 km2 and provides a rangeof environments, with flow rates from sluggish (in thebackwaters) to fast, and temperatures increased by vari-ous amounts depending on location within the basin. Anice cover is never formed inside the Biotest Basin. Thesurroundings of the basin provide a complementary setof environments, with and without ice cover and withvarying degrees of heating (e.g. by leakage from thebasin) and varying exposure to natural waves. Forfurther area description see Snoeijs & Prentice (1989).
Methods
Sampling and data collection
Samples were taken on 23 dates in the years 1983 -1987 and 1989 from 13 sites in and outside the BiotestBasin (in some cases fewer sites). For each of the 200samples taken, eight more-or-less flat-surfaced stones(diameter 7 - 12 cm) were taken from a depth of 20 - 50cm. The same type of stones occurred in all samplingsites, and were derived from the dynamiting during theconstruction of the basin (in 1977). In the years 1983,1984, 1985 and 1989 three to five additional samples persite were taken to measure the ash-free dry weight of thetotal microphytobenthos (mainly diatoms).
For each site / date water temperature was measured.Nutrient concentrations (at six sites) and insolation (PAR= photosynthetically active radiation) were measured in1983, 1984 and 1985, but not in the other years. Fromthe mean values of the available measurements general-ized curves were constructed, so that for all six years atleast the seasonal trend of nutrient concentrations and
Table 1. Cover scales used for diatoms (D) andmacroalgae. S = stone surface; vis. = visible; col. =colonies; Ind. = individuals.
Diatoms MacroalgaeScore Cover % Ind.
1 S vis., D not vis. < 1 1 - 52 S vis., D vis. on part of stone < 1 5 - 103 S vis., D vis. on whole stone < 1 > 104 S not vis., D layer < 1 mm 1 - 55 S not vis., D layer 1 - 2 mm 6 - 106 S not vis., D layer 3 - 5 mm 11 - 257 D in upright col. 0.5 - 5 cm 26 - 508 D in upright col. 6 - 10 cm 51 - 759 D in upright col. > 10 cm 76 -100
light could be used in the data analyses.Water movement varied mainly with sampling site.
The sites could easily be divided into three groupsaccording to water movement conditions. Stagnantwater means little water movement, even when strongwinds are blowing. Rough water includes differentdegrees of water movement caused by wind. Flowingwater occurs only as a result of cooling water intake ordischarge; here the water flow is unidirectional (inde-pendent of the prevailing winds) and relatively constant,but more or less fast depending on the water volumedischarged and the situation of the site. The environ-mental factor ‘water movement’ was used in the analy-ses either as a three-class variable or as an ordinalvariable for the amount of water movement: 1 = stag-nant; 2 = rough; 3 = flowing.
Three sites with different water movement condi-tions (stagnant, rough and flowing water) were consid-ered reference sites for temperature, all other sites wereto some extent artificially heated. The ‘ temperatureanomaly’ for each sample was calculated as the differ-ence in water temperature between the sampling site anda reference site with the same water-movement condi-tions. When ice cover occurred at the reference siteswith stagnant and rough water, no samples could betaken. The reference site with flowing water was neverice-covered because of the strong water flow there.
In the laboratory the algae were brushed off thestones, and the amount of diatoms and macroalgae wasestimated according to a cover scale (Table 1). In fouryears microphytobenthic ash-free dry weight was alsodetermined. The samples for the determination of spe-cies composition were treated with hydrogen peroxideand potassium dichromate and mounted in Hyrax. Persample 1000 diatom valves - 500 from each of twosubsamples - were identified and counted along a tran-sect of the cover glass at magnification × 1000 using oilimmersion. For each species a relative abundance wascalculated as a percentage of the sample. Taxonomical
- Effects of temperature on spring bloom dynamics of diatom communities - 601
Multiple regression analysis was applied to testdiatom cover, Shannon-Weaver diversity and relativeabundances of individual diatom taxa for the effects ofdifferent environmental variables acting in combination(Jongman, ter Braak & van Tongeren 1987; ter Braak& Prentice 1988). A significant regression coefficientimplies that the variable has a definite effect when theother variables are held constant. Recently the problemof autocorrelation occurring in most if not allenvironmental data has been discussed (Legendre &Fortin 1989). Autocorrelation biases the test ofsignificance of regression coefficients in many statisticalmethods used in ecology. Therefore the correlationsthat are only marginally significant in the multipleregression analysis are considered less significant thanthe p-values indicate. The Shannon-Weaver index (H',log base = e) was computed for all samples.
Results
Environmental factors
Spring water temperatures at Forsmark naturallyvary with the time of year and from year to year dependingon air temperatures and severity of the preceding winter.In the cooling water discharge area there are also spatialwater temperature variations, depending on the degreeof heating of the water at the sampling site. FromJanuary to April the reference sites had water tempera-tures of ca. 0 °C and the most heated sites ca. 10 °C. Bythe end of April temperatures started rising in line withthe seasonal cycle, and by the end of May the lowestwater temperature was ca. 6 °C and the highest ca. 19 °C.All possible temperature anomalies between 1 and 10 °Care well represented by the samples in this study.
Heated sites with flowing water generally had highertemperature anomalies than those with stagnant or roughwater. The general pattern of temperature anomalybetween sites in winter and early spring changed in Maydepending on water movement conditions; sites withshallow stagnant water were heated up by the sun moreeasily than sites with fast flow. For example, on 23 May1984 a site with fast flowing artificially heated water hada temperature of 19.5 °C, which was hardly more thanthe reference site with stagnant water (19.3 °C).
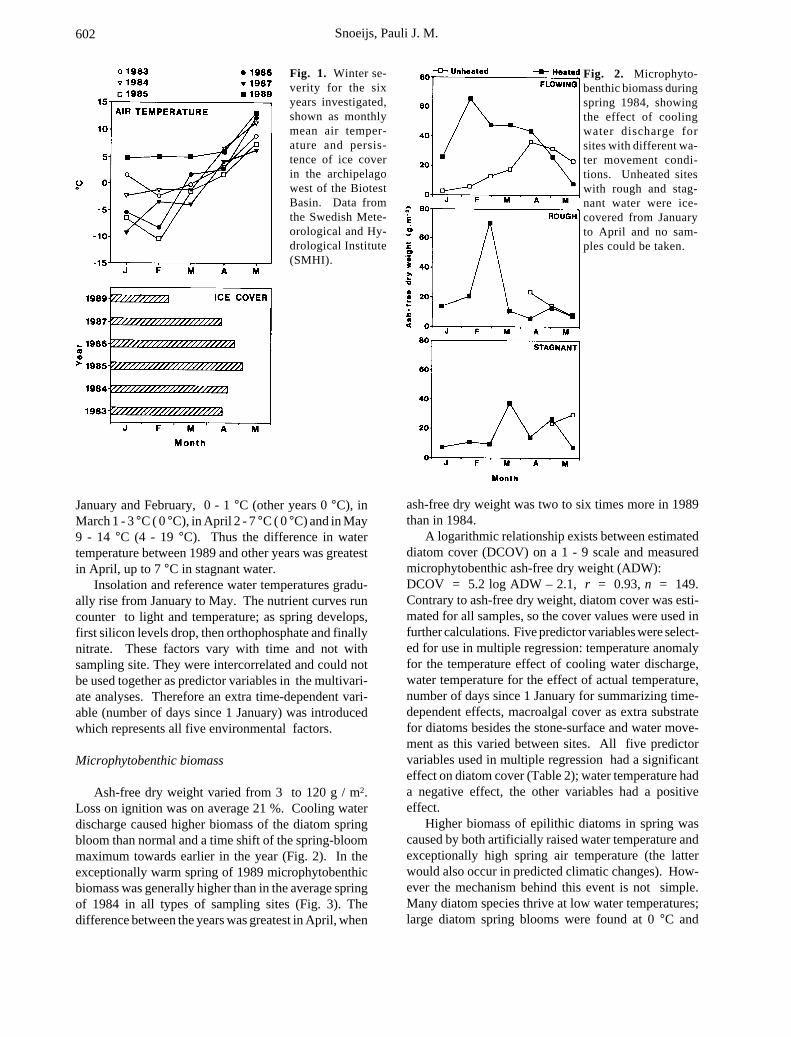
Air temperatures and persistence of ice cover for thesix years investigated are given in Fig. 1. In 1989 airtemperatures were 9, 10 and 6 °C higher than the meansduring the other five years in January, February andMarch respectively. Therefore only a very thin ice coverwas locally formed in the area in 1989, which disap-peared as early as February instead of April. In thereference sites water temperatures in 1989 were, during
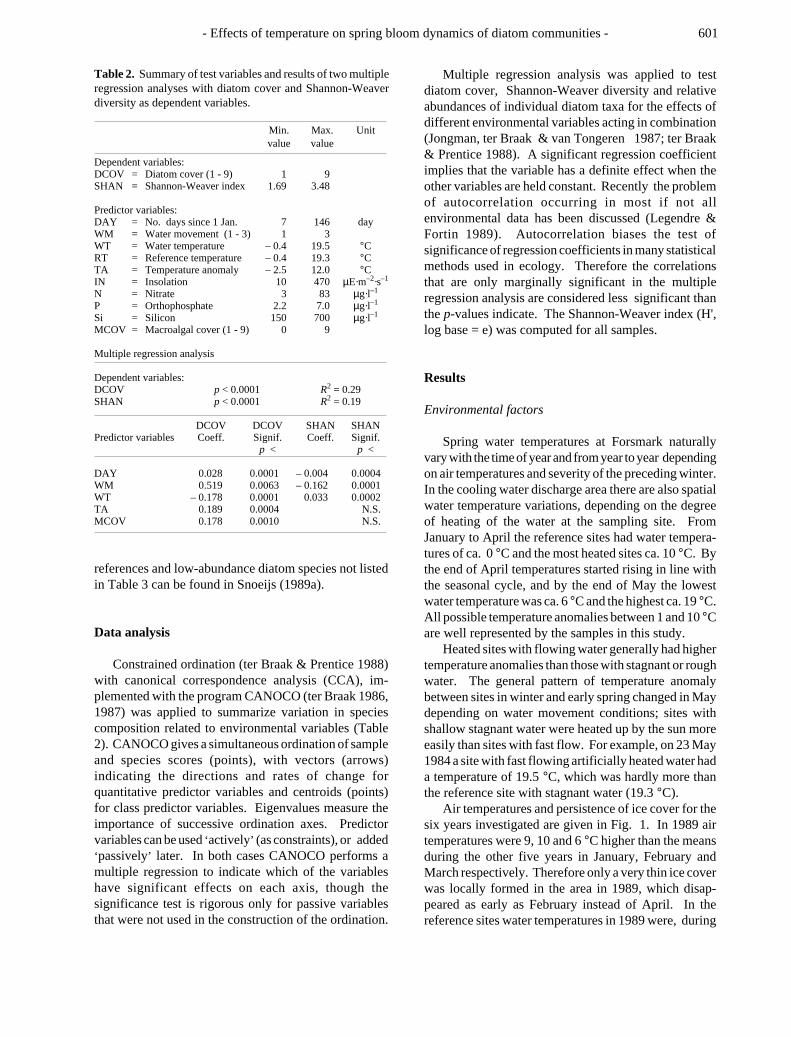
Table 2. Summary of test variables and results of two multipleregression analyses with diatom cover and Shannon-Weaverdiversity as dependent variables.
Min. Max. Unitvalue value
Dependent variables:DCOV = Diatom cover (1 - 9) 1 9SHAN = Shannon-Weaver index 1.69 3.48
Predictor variables:DAY = No. days since 1 Jan. 7 146 dayWM = Water movement (1 - 3) 1 3WT = Water temperature – 0.4 19.5 °CRT = Reference temperature – 0.4 19.3 °CTA = Temperature anomaly – 2.5 12.0 °CIN = Insolation 10 470 µE·m–2·s–1
N = Nitrate 3 83 µg·l–1
P = Orthophosphate 2.2 7.0 µg·l–1
Si = Silicon 150 700 µg·l–1
MCOV = Macroalgal cover (1 - 9) 0 9
Multiple regression analysis
Dependent variables:DCOV p < 0.0001 R2 = 0.29SHAN p < 0.0001 R2 = 0.19
DCOV DCOV SHAN SHANPredictor variables Coeff. Signif. Coeff. Signif.
p < p <
DAY 0.028 0.0001 – 0.004 0.0004WM 0.519 0.0063 – 0.162 0.0001WT – 0.178 0.0001 0.033 0.0002TA 0.189 0.0004 N.S.MCOV 0.178 0.0010 N.S.
references and low-abundance diatom species not listedin Table 3 can be found in Snoeijs (1989a).
Data analysis
Constrained ordination (ter Braak & Prentice 1988)with canonical correspondence analysis (CCA), im-plemented with the program CANOCO (ter Braak 1986,1987) was applied to summarize variation in speciescomposition related to environmental variables (Table2). CANOCO gives a simultaneous ordination of sampleand species scores (points), with vectors (arrows)indicating the directions and rates of change forquantitative predictor variables and centroids (points)for class predictor variables. Eigenvalues measure theimportance of successive ordination axes. Predictorvariables can be used ‘actively’ (as constraints), or added‘passively’ later. In both cases CANOCO performs amultiple regression to indicate which of the variableshave significant effects on each axis, though thesignificance test is rigorous only for passive variablesthat were not used in the construction of the ordination.
Snoeijs, Pauli J. M.602
Fig. 1. Winter se-verity for the sixyears investigated,shown as monthlymean air temper-ature and persis-tence of ice coverin the archipelagowest of the BiotestBasin. Data fromthe Swedish Mete-orological and Hy-drological Institute(SMHI).
Fig. 2. Microphyto-benthic biomass duringspring 1984, showingthe effect of coolingwater discharge forsites with different wa-ter movement condi-tions. Unheated siteswith rough and stag-nant water were ice-covered from Januaryto April and no sam-ples could be taken.
ash-free dry weight was two to six times more in 1989than in 1984.
A logarithmic relationship exists between estimateddiatom cover (DCOV) on a 1 - 9 scale and measuredmicrophytobenthic ash-free dry weight (ADW):DCOV = 5.2 log ADW – 2.1, r = 0.93, n = 149.Contrary to ash-free dry weight, diatom cover was esti-mated for all samples, so the cover values were used infurther calculations. Five predictor variables were select-ed for use in multiple regression: temperature anomalyfor the temperature effect of cooling water discharge,water temperature for the effect of actual temperature,number of days since 1 January for summarizing time-dependent effects, macroalgal cover as extra substratefor diatoms besides the stone-surface and water move-ment as this varied between sites. All five predictorvariables used in multiple regression had a significanteffect on diatom cover (Table 2); water temperature hada negative effect, the other variables had a positiveeffect.
Higher biomass of epilithic diatoms in spring wascaused by both artificially raised water temperature andexceptionally high spring air temperature (the latterwould also occur in predicted climatic changes). How-ever the mechanism behind this event is not simple.Many diatom species thrive at low water temperatures;large diatom spring blooms were found at 0 °C and
January and February, 0 - 1 °C (other years 0 °C), inMarch 1 - 3 °C ( 0 °C), in April 2 - 7 °C ( 0 °C) and in May9 - 14 °C (4 - 19 °C). Thus the difference in watertemperature between 1989 and other years was greatestin April, up to 7 °C in stagnant water.
Insolation and reference water temperatures gradu-ally rise from January to May. The nutrient curves runcounter to light and temperature; as spring develops,first silicon levels drop, then orthophosphate and finallynitrate. These factors vary with time and not withsampling site. They were intercorrelated and could notbe used together as predictor variables in the multivari-ate analyses. Therefore an extra time-dependent vari-able (number of days since 1 January) was introducedwhich represents all five environmental factors.
Microphytobenthic biomass
Ash-free dry weight varied from 3 to 120 g / m2.Loss on ignition was on average 21 %. Cooling waterdischarge caused higher biomass of the diatom springbloom than normal and a time shift of the spring-bloommaximum towards earlier in the year (Fig. 2). In theexceptionally warm spring of 1989 microphytobenthicbiomass was generally higher than in the average springof 1984 in all types of sampling sites (Fig. 3). Thedifference between the years was greatest in April, when

- Effects of temperature on spring bloom dynamics of diatom communities - 603
Fig. 3. Microphytobenthicbiomass during the exception-ally warm spring of 1989 com-pared with the average springof 1984 for sites with differ-ent water movement condi-tions. The samples were takenin the middle of each month.* = no sample taken becauseof ice cover.
multiple regression analysis (Table 2) showed a nega-tive correlation between water temperature and diatombiomass, because water temperature was highest in Maywhen biomass had declined after the maximum in April.Fig. 4 shows how the different environmental factorsmight affect each other, with higher diatom biomass asthe ultimate result. Ice cover is crucial in the chain ofeffects. No ice cover implies no abrading effect of ice-break in spring (diatoms are not rubbed off), higher lightavailability throughout winter (light is not inhibited byice and snow) and more water movement is possible (byprevailing winds). No abrading also means that macro-algae are not rubbed off (larger substrate area fordiatoms to colonize on). The factors directly affectingdiatom growth were here identified as reduced loss byreduced abrading and higher production by higher avail-ability of light, nutrients and substrate.
Cooling water discharge and community composition
The 200 samples contained 251 diatom taxa. Thesame five factors (temperature anomaly, water tem-perature, water movement, number of days since 1January, and macroalgal cover) were used as constraintsin CCA. Other variables were added passively. Thefour CCA axes had eigenvalues 0.18, 0.12, 0.09, and0.03). Multiple regression of the environmental variableson ordination axes 1 and 2 showed significance (p <0.05) for all variables on one or both axes.
The vectors for environmental variables are relatedto the correlations between each variable and the axes(Fig. 5); they point in the direction of maximum increaseof the variables. Water movement is related to axis 2,
Fig 4. Hypothetical synthesis of interactions between envi-ronmental factors that lead to higher diatom biomass.
with increasing water movement down the axis. Time isrelated to axis 1, with highest nutrient concentrations inJanuary to the right and highest insolation and referencewater temperatures in May to the left. The vector fortemperature anomaly points towards the lower-rightquadrant, indicating that the temperature effect of coolingwater discharge is greatest during the first months of thesampling period in sites with flowing water. The factorwater temperature is composed of reference temperature(dependent on time and water movement conditions)and temperature anomaly, which have vectors in oppositedirections in the ordination. The vector for watertemperature points in a direction intermediate to referencetemperature and temperature anomaly, and has theshortest arrow (least effect). The vectors for macroalgalcover and diatom cover point to the lower part of theordination; that for Shannon-Weaver diversity points inthe opposite direction of diatom cover.
The species points (Table 3) summarize the distri-butions of the diatom taxa by time and environment.E.g. the lower-right quadrant contains the taxa mostfavoured early in the year by the cooling water discharge(e.g. Melosira spp. and Navicula perminuta), and theupper half of the ordination shows taxa that occur morein stagnant water (e.g. Fragilariaceae). CCA sum-marizes the effects of specified variables on communitycomposition, but it can give only limited informationabout the specific responses of individual taxa. Table 3shows the results of multiple regressions and deter-mination of temperature optima (n = 200) from graphi-cal analysis for the 47 most abundant taxa. Most taxashow responses to one or more test-variables. Log-transformation of abundances did not affect the results.
Ecological groups
The overall response patterns of individual speciesto temperature anomaly (TA), water movement (WM),time (DAY) and macroalgal cover (MCOV) are largely

Snoeijs, Pauli J. M.604
Fig. 5. CCA ordination (axes 1 and 2) of 200 diatomsamples, constrained by five predictor variables (thickarrows). The other variables were added passively (thinarrows). For abbreviations see Table 2.
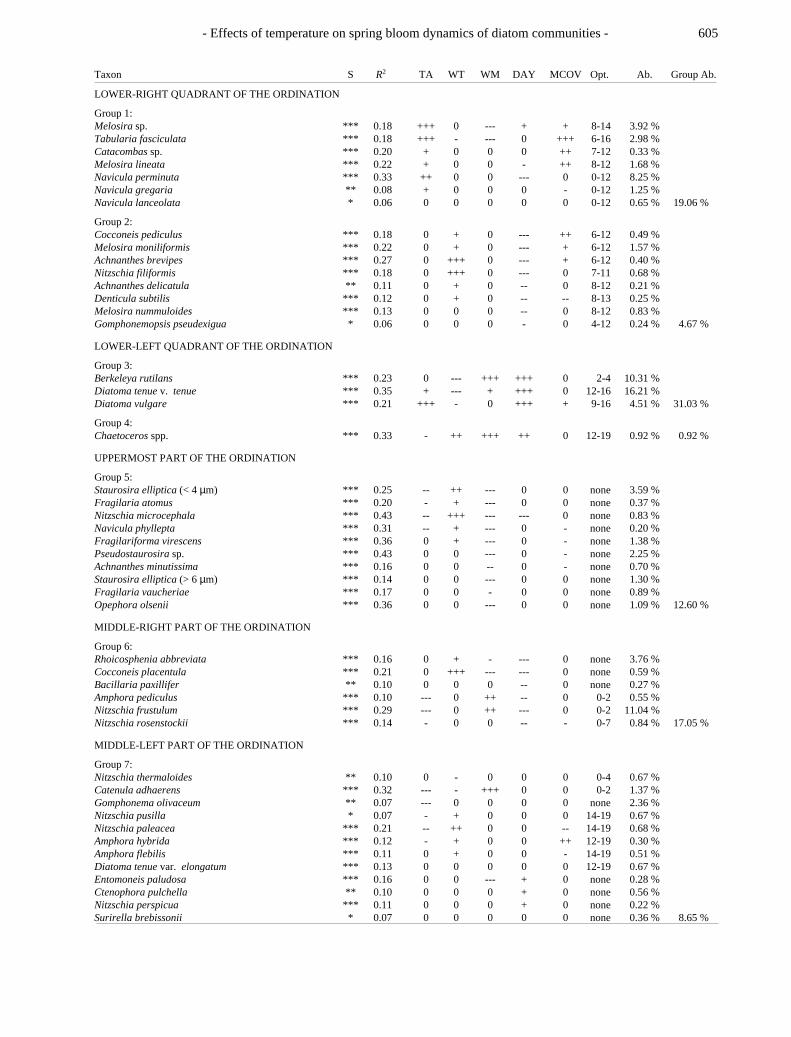
Table 3. Results of multiple regression analysis (n = 200)and graphical analysis of temperature optima for the 47most abundant diatom taxa (mean abundance > 0.20 %;total abundance 94 %). The taxa are grouped according totheir scores in the CCA ordination (Figs. 5 - 6), and theresults of multiple regression. opt = optimum (°C); Ab. =abundance; for further abbreviations see Table 2.S = Significance levels (*** = p < 0.001; ** = p < 0.01; * = p <0.05) Responses to predictor variables in multiple regressionanalysis: Positive coefficient: +++ = p < 0.001; ++ = p < 0.01; += p < 0.05; Negative coefficient: --- = p < 0.001; -- = p < 0.01; -= p < 0.05; 0 = no response; none = no temperature optimum wasfound by graphical analysis.
Fig. 6. Taxon ordination from the same analysis as Fig. 5 forthe taxa with mean relative abundance > 0.20 %. A-E: Thesymbols + and - indicate the sign of the (significant) regressioncoefficients for each taxon with respect to the five predictorvariables that were used as constraints. Non-significant (p >0.05) relationships are left out. Each arrow indicates thedirection and rate of change of the investigated environmentalvariable. For abbreviations see Table 2. F: The numbersindicate the ecological groups of taxa according to Table 3.
in agreement with the same species' scores in the com-munity analysis with CCA, but water temperature (WT)shows a deviating pattern (Fig. 6). In Table 3 the taxa aredivided into ‘ecological groups’ according to the resultsof CCA and multiple regression. The taxa of each groupreact in a different manner to environmental conditions.- Group 1 taxa have a positive relation to temperatureanomaly caused by cooling water discharge, some alsoto macroalgal cover. Although Melosira sp. also reactsstrongly to water movement and Navicula perminuta totime as shown by multiple regression, their position inthe ordination depends on temperature anomaly. - Group 2: time is the most important factor; taxa occurearly in the year (January-February). But as seasonaltemperature is lowest early in the year and the reaction towater temperature for most of them is positive, thesetaxa can also be considered reacting positively to cool-ing water discharge; they are situated in the direction oftemperature anomaly rather than reference temperature.- Group 3 taxa occur abundantly towards the end of theinvestigation period (March-April). Normally Berke-leya rutilans and Diatoma tenue are typical of the springbloom in the littoral of this part of the Baltic Sea. B.rutilans and Diatoma spp. have quite different tempera-ture optima. B. rutilans prefers water temperatures be-low 4 °C. D. tenue reacts positively to cooling waterdischarge by being abundant earlier in the year thannormal. D. vulgare is almost totally absent in unheatedsites.- Group 4 is represented by the pelagic diatom genusChaetoceros spp. only. Its occurence in the littoraldepends on the time of the phytoplankton spring bloom(April-May).- Group 5 taxa are found in the uppermost part of theordination; they show strong affinity for stagnant water.The abundance of most taxa depends on the amount ofsilt on the stones, which is highest in sites with stagnantwater. Some taxa also react to temperature, but for none
of them could a clear temperature optimum be defined;water movement is the dominant factor here.- Group 6: time is important; taxa occur early in theyear. Otherwise this group is quite heterogeneous.Amphora pediculus, Nitzschia frustulum and N.rosenstockii react negatively to temperature anomalyand have low temperature optima.- Group 7 is also heterogeneous, some taxa reactnegatively to temperature anomaly, either with low tem-perature optima (e.g. Catenula adhaerens), or (‘summer
- Effects of temperature on spring bloom dynamics of diatom communities - 605
Taxon S R2 TA WT WM DAY MCOV Opt. Ab. Group Ab.
LOWER-RIGHT QUADRANT OF THE ORDINATION
Group 1:Melosira sp. *** 0.18 +++ 0 --- + + 8-14 3.92 %Tabularia fasciculata *** 0.18 +++ - --- 0 +++ 6-16 2.98 %Catacombas sp. *** 0.20 + 0 0 0 ++ 7-12 0.33 %Melosira lineata *** 0.22 + 0 0 - ++ 8-12 1.68 %Navicula perminuta *** 0.33 ++ 0 0 --- 0 0-12 8.25 %Navicula gregaria ** 0.08 + 0 0 0 - 0-12 1.25 %Navicula lanceolata * 0.06 0 0 0 0 0 0-12 0.65 % 19.06 %
Group 2:Cocconeis pediculus *** 0.18 0 + 0 --- ++ 6-12 0.49 %Melosira moniliformis *** 0.22 0 + 0 --- + 6-12 1.57 %Achnanthes brevipes *** 0.27 0 +++ 0 --- + 6-12 0.40 %Nitzschia filiformis *** 0.18 0 +++ 0 --- 0 7-11 0.68 %Achnanthes delicatula ** 0.11 0 + 0 -- 0 8-12 0.21 %Denticula subtilis *** 0.12 0 + 0 -- -- 8-13 0.25 %Melosira nummuloides *** 0.13 0 0 0 -- 0 8-12 0.83 %Gomphonemopsis pseudexigua * 0.06 0 0 0 - 0 4-12 0.24 % 4.67 %
LOWER-LEFT QUADRANT OF THE ORDINATION
Group 3:Berkeleya rutilans *** 0.23 0 --- +++ +++ 0 2-4 10.31 %Diatoma tenue v. tenue *** 0.35 + --- + +++ 0 12-16 16.21 %Diatoma vulgare *** 0.21 +++ - 0 +++ + 9-16 4.51 % 31.03 %
Group 4:Chaetoceros spp. *** 0.33 - ++ +++ ++ 0 12-19 0.92 % 0.92 %
UPPERMOST PART OF THE ORDINATION
Group 5:Staurosira elliptica (< 4 µm) *** 0.25 -- ++ --- 0 0 none 3.59 %Fragilaria atomus *** 0.20 - + --- 0 0 none 0.37 %Nitzschia microcephala *** 0.43 -- +++ --- --- 0 none 0.83 %Navicula phyllepta *** 0.31 -- + --- 0 - none 0.20 %Fragilariforma virescens *** 0.36 0 + --- 0 - none 1.38 %Pseudostaurosira sp. *** 0.43 0 0 --- 0 - none 2.25 %Achnanthes minutissima *** 0.16 0 0 -- 0 - none 0.70 %Staurosira elliptica (> 6 µm) *** 0.14 0 0 --- 0 0 none 1.30 %Fragilaria vaucheriae *** 0.17 0 0 - 0 0 none 0.89 %Opephora olsenii *** 0.36 0 0 --- 0 0 none 1.09 % 12.60 %
MIDDLE-RIGHT PART OF THE ORDINATION
Group 6:Rhoicosphenia abbreviata *** 0.16 0 + - --- 0 none 3.76 %Cocconeis placentula *** 0.21 0 +++ --- --- 0 none 0.59 %Bacillaria paxillifer ** 0.10 0 0 0 -- 0 none 0.27 %Amphora pediculus *** 0.10 --- 0 ++ -- 0 0-2 0.55 %Nitzschia frustulum *** 0.29 --- 0 ++ --- 0 0-2 11.04 %Nitzschia rosenstockii *** 0.14 - 0 0 -- - 0-7 0.84 % 17.05 %
MIDDLE-LEFT PART OF THE ORDINATION
Group 7:Nitzschia thermaloides ** 0.10 0 - 0 0 0 0-4 0.67 %Catenula adhaerens *** 0.32 --- - +++ 0 0 0-2 1.37 %Gomphonema olivaceum ** 0.07 --- 0 0 0 0 none 2.36 %Nitzschia pusilla * 0.07 - + 0 0 0 14-19 0.67 %Nitzschia paleacea *** 0.21 -- ++ 0 0 -- 14-19 0.68 %Amphora hybrida *** 0.12 - + 0 0 ++ 12-19 0.30 %Amphora flebilis *** 0.11 0 + 0 0 - 14-19 0.51 %Diatoma tenue var. elongatum *** 0.13 0 0 0 0 0 12-19 0.67 %Entomoneis paludosa *** 0.16 0 0 --- + 0 none 0.28 %Ctenophora pulchella ** 0.10 0 0 0 + 0 none 0.56 %Nitzschia perspicua *** 0.11 0 0 0 + 0 none 0.22 %Surirella brebissonii * 0.07 0 0 0 0 0 none 0.36 % 8.65 %

Snoeijs, Pauli J. M.606
Diversity
The Shannon-Weaver index varied from 1.69 to3.48, and was correlated to both evenness (r = 0.96, n =200) and species richness (r = 0.82, n = 200). Coolingwater discharge or exceptionally high spring airtemperatures did not affect diatom species diversity.Multiple regression (Table 2) showed that the Shannon-Weaver index was negatively related to number of dayssince 1 January and water movement, positively to watertemperature and not to temperature anomaly ormacroalgal cover. The signs of the environmental factorsthat both affected diversity and diatom cover are oppositefor these two community parameters in multipleregression, and the vectors for diatom cover and Shannon-Weaver diversity are opposite in the CCA ordination(Fig. 5); no linear relationship occurred between them.
Discussion
Comparison of diatoms with other algae
Compared with other algal groups, diatoms are quan-titatively most favoured by the cooling water dischargein Forsmark. The diatoms profit most from high winter-nutrient concentrations and enhanced insolation by lackof ice cover because they have relatively low tempera-ture optima and light requirements (Snoeijs & Kautsky1989), fast turnover and immigration rates, and they areecologically extremely diverse; > 300 epilithic speciesoccur at Forsmark (Snoeijs 1989a) with a great varietyof growth forms and life-history strategies (Snoeijs1989b). Widely varying responses shown by diatomsare found in Table 3. Green and blue-green algae inForsmark prefer higher water temperatures than mostdiatom species (>13 °C). Red and brown algae prefer(like diatoms) low water temperatures, but they haveslower growth rates and, in this part of the Baltic Sea, asmall species pool because of the low salinity (Wærn1952; Snoeijs & Prentice 1989).
Biomass and species composition
The diatoms adding most to the microphytobenthicbiomass are medium to large-sized species (from vol-ume ca. 500 µm3 , e.g. Berkeleya rutilans, up to volumeca. 50 000 µm3, e.g. Melosira moniliformis). Severalsmaller diatoms that in some samples were very abun-dant e.g. Fragilariaceae and Nitzschia frustulum (< 50µm3) were of less importance with respect to biomass.The diatoms mainly responsible for the high spring-bloom biomass under natural conditions are Berkeleyarutilans, Diatoma tenue, Gomphonema olivaceum andNavicula spp., and those dominating under artificial
Fig. 7. CCA ordination (axes 1 and 2) of 32 diatom samplesfrom unheated sites, showing centroids (circles) for watermovement * year(s) and taxon scores (squares) for the taxawith mean abundance > 2 % in the dataset. BER RUTI =Berkeleya rutilans; DIA TENU = Diatoma tenue var. tenue;FRA VAUC = Fragilaria vaucheriae; FFO VIRE =Fragilariforma virescens; GOM OLIV = Gomphonemaolivaceum; NAV PERM = Navicula perminuta; NIT FRUS =Nitzschia frustulum; PSE SPEC = Pseudostaurosira sp.; RHOABBR = Rhoicosphenia abbreviata; SRA ELMI = Staurosiraelliptica (<4 µm).
species’) with high ones (e.g. Nitzschia paleacea).
Winter severity and community composition
CCA and multiple regression show the effect ofheating by cooling-water discharge (variable TA), butthe effect of winter severity on community compositionwas not shown. In an additional CCA analysis only the32 samples from unheated sites that have an ice cover inwinter were used. Four class variables for watermovement * year(s) were used as constraints. The fourCCA axes had eigenvalues 0.19, 0.11, 0.04, and 0.03. Inthis analysis the exceptionally warm spring of 1989 wascompared with the more normal ones in 1983-1987.Melosira spp. that were shown to profit from thermalinput by cooling water discharge did not appear abun-dantly in 1989 in the reference sites either (but theybecame even more abundant in artificially heated sites in1989). Instead some diatom species that normally occurin the spring bloom in this part of the Baltic Sea reactedin the reference sites: Berkeleya rutilans increased verymuch in relative abundance in April in sites with roughwater and Diatoma tenue occurred earlier in the year;also Navicula perminuta was enhanced in 1989. Thesetaxa together accounted for a large part of the extremelyhigh biomass in 1989 (Fig. 3). Some taxa common in1983-1987 (Gomphonema olivaceum in reference sites,Diatoma vulgare in heated sites) decreased greatly inrelative abundance in 1989. Also Fragilaria vaucheriaeand Nitzschia frustulum decreased in 1989. Some smallFragilariaceae were not affected.

- Effects of temperature on spring bloom dynamics of diatom communities - 607
heating with total absence of ice cover are Melosira spp.,Diatoma vulgare, D. tenue and Navicula spp. All ofthese are colony-forming taxa (Snoeijs 1989b), and thusable to grow fast by vertical expansion towards betterlight and nutrient conditions.
Ice cover was identified as a crucial factor control-ling biomass in spring (Fig. 4). Earlier ice-break in1989 resulted in higher abundances of some species thatnaturally also dominate the spring bloom in the upperlittoral, in spite of up to 7 °C higher water temperature inApril than in other years. In some heated sites withtemperature anomaly 5 - 6 °C the spring bloom wastotally dominated by Melosira spp. Melosira spp. werealso found to dominate at other sites in the Baltic Seawhere cooling water discharge prevents an ice coverfrom forming (Kuylenstierna 1982; Keskitalo & Heitto1987) and usually occur in the cold season in marinesites with higher salinity where an ice cover is neverformed (Hendey 1964). It seems that ice cover also hasan effect in interspecific competition. B. rutilans has aheteromorphic growth form, either colonial or as indi-vidual cells (Hudon & Legendre 1987), and is thereforea good early colonizer of bare rock in its solitary motileform. But when the rocks are not bare in spring, autumnand winter species are allowed to persist, either as (low-abundant) ordinary cells or vegetative resting stadia(Stoermer 1986). This might be the reason for domi-nance of Melosira spp., which may also be promoted bythe persistence of basal parts of some epilithic macro-algae (notably Cladophora glomerata) through winterwhen there is no ice cover.
Diversity
Community diversity would be expected to decreasewhen biomass increases through dominance of one orfew fast-growing colonial diatom species which wasfound to be an effect of raised temperature. But no linearcorrelation was found between diatom cover and Shan-non-Weaver diversity. This is partly explained by thefact that some of the samples were taken at an earlysuccessional stage: primary colonization of the barestones after ice-break. These samples had low cover andfew species. The mean effect of all samples is howeverthat expected; in the ordination (Fig. 5) the vector forcover is exactly opposite to that of diversity.
Temperature optima
In contrast to recent studies determining pH optimaof diatoms (ter Braak & van Dam 1989; Oksanen et al.1990), the temperature optima given in Table 3 shouldbe interpreted with great care. They are extracted froma dataset from a limited area with extremely smallsalinity fluctuations, and other environmental factors
than temperature are (more) important in the distribu-tion of some species.
Effects of a global temperature increase
This study shows that temperature input by reducingthe ice season promotes the growth of benthic diatoms inspring in a shallow coastal ecosystem with low salinity.In case of a global temperature increase >20 cm thickslimy brown layers of diatoms (as in 1989) might occureach year along the Baltic coast in spring. Generallydiatoms become more important compared with otherlittoral algae when salinity decreases, so in the northern-most part of the Baltic Sea the described effect would begreatest.
Surface water temperature has risen about 1 °C sincethe beginning of this century (Launiainen & Koljonen1981) and the length of the ice season (largely governedby air temperature) has decreased by 20-30 days over thesame period (Leppäranta & Sein 1985). Leppäranta(1989) calculated that a rise of 1 °C in the air tempera-ture level would delay the freezing date by six days,decrease ice-thickness by 8 cm and make the break-update of 50 - cm ice eight days earlier. In 1989 ice-breakoccurred about six weeks earlier than in the other fiveyears investigated, caused by 8 °C higher air tempera-ture in January - March. Compared with a longer timeseries (1944-1981: Eriksson 1982) the mean air tem-perature in January-March was also 8 °C higher. Com-plete absence of ice cover in this part of the Baltic Seawould thus require even higher air temperature risesthan ca. 8 °C. The predicted sea level rise (Bach 1989;Coker et al. 1989; Lindh, Hansson & Larsson 1989)resulting from the ‘greenhouse effect’ will not affectepilithic diatom communities as colonization is fast anddoes not depend on gradual changes in water level.
Large diatom blooms can cause nuisance, e.g. fish-ing nets and the under-water parts of boats get over-grown by a thick slimy layer. If climatic change wouldgo as far as preventing an ice cover from formingcompletely, Melosira spp. could take over in areas withlow salinity as early in the year as February. This wouldmean that the spring bloom would not be restricted to asubstrate anymore, as Melosira colonies have a looseattachment to the substrate and therefore possess somemobility as whole colonies. Large loose colonies canentrap air bubbles and float close to the water surfaceallowing them to make full use of available light for theirfast growth. Many such floating colonies can cause lightinhibition for the benthic and pelagic systems, and oxy-gen depletion when they die off.
Acknowledgements: Florence Forsell assisted in the fieldand the laboratory. Ulf Grimås, head of the Forsmarkproject at the Swedish Environmental Protection Agency

Snoeijs, Pauli J. M.608
(SNV), helped in many practical ways. Jan Anderssonsupplied air temperature-, insolation- and ice cover datafrom the Swedish Meteorological and HydrologicalInstitute (SMHI). Eddy van der Maarel, Pierre Legendre,Olof Sandström and two reviewers commented on themanuscript. The research was financed by the SwedishEnvironmental Protection Agency (SNV).
References
Legendre, P. & Fortin, M.-J. 1989. Spatial pattern andecological analysis. Vegetatio 80: 107-138.
Leppäranta, M. 1989. On climatic variations of the BalticSea ice conditions. In: Conference on climate andwater. Publ. Acad. Finland 9, Vol. 1, pp. 63-72.
Leppäranta, M. & Sein, A. 1985. Freezing, maximumannual ice thickness and break-up of ice on the Finnishcoast during 1830-1984. Geophysica 21: 87-104.
Lindh, G., Hanson, H. & Larson, M. 1989. Impact of sealevel rise on coastal zone management in southernSweden. In: Conference on climate and water. Publ.Acad. Finland 9, Vol. 2, pp. 129-147.
Oksanen, J., Läära, E., Huttonen, P. & Meriläinen, J. 1990.Maximum likelihood prediction of lake acidityaccording to sedimental diatoms. J. Veg. Sci. 1: 49-56.
Snoeijs, P. J. M. 1989a. A check-list of the benthicdiatoms at Forsmark (northern Baltic Sea). 1. Epilithicand epiphytic taxa. Ann. Bot. Fenn. 26: 427-439.
Snoeijs, P. J. M. 1989b. Ecological effects of coolingwater discharge on hydrolittoral epilithic diatomcommunities in the northern Baltic Sea. Diatom Res. 4:373-398.
Snoeijs, P. J. M. & Kautsky, U. 1989. Effects of ice-breakon the structure and dynamics of a benthic diatomcommunity in the northern Baltic Sea.Bot. Mar. 32: 547-562.
Snoeijs, P. J. M. & Prentice, I. C. 1989. Effects ofcooling water discharge on the structure and dynamicsof epilithic algal communities in the northern Baltic.Hydrobiologia 184: 99-123.
Stoermer, E. F. 1986. Some perspectives on diatom ecology.In: Ricard, M. (ed.), Proc. 8. Int. Diatom Symposium,pp. 315-322.
ter Braak, C. J. F. 1986. Canonical correspondence analysis:a new eigenvector technique for multivariate directgradient analysis. Ecology 67: 1167-1179.
ter Braak, C. J. F. 1987. CANOCO - a FORTRAN programfor canonical community ordination by [partial][detrended] [canonical] correspondence analysis,principal components analysis and redundancy analysis(version 2.1). TNO Institute of Applied ComputerScience, Wageningen.
ter Braak, C. J. F. & Prentice, I. C. 1988. A theory ofgradient analysis. Adv. Ecol. Res. 18: 271-317.
ter Braak, C. J. F. & van Dam, H. 1989. Inferring pH fromdiatoms: a comparison of old and new calibrationmethods. Hydrobiologia 178: 209-223.
Wærn, M. 1952. Rocky-shore algae in the Öregrundarchipelago. Acta Phytogeogr. Suec. 30: 1-298.
Williams, D. M. & Round, F. E. 1986. Revision of thegenus Synedra Ehrenb. Diatom Res. 1: 313-339.
Williams, D. M. & Round, F. E. 1987. Revision of thegenus Fragilaria . Diatom Res. 2: 267-288.
Williams, D. M. & Round, F. E. 1988. Fragilariforma,nom. nov., a new generic name for NeofragilariaWilliams & Round. Diatom Res. 3: 265-266.
Received 11 January 1990;Revision received 26 June 1990;
Accepted 29 June 1990.
Alenius, P. 1989. Variations of the sea temperature aroundthe coasts of Finland. In: Conference on climate andwater. Publ. Acad. Finland 9, Vol. 1, pp. 51-62.
Bach, W. 1989. Projected climatic changes and impacts inEurope due to increased CO2. In: Conference onclimate and water. Publ. Acad. Finland 9, Vol. 1, pp.31-50.
Budyko, M. I. 1989. Climatic conditions of the future. In:Conference on climate and water. Publ. Acad. Finland9, Vol. 1, pp. 9-30.
Coker, A. M., Thompson, P. M., Smith, D. I. & Penning-Rowsell, E. C. 1989. The impact of climate change oncoastal zone management in Britain: a preliminaryanalysis. In: Conference on climate and water. Publ.Acad. Finland 9, Vol. 2, pp. 148-160.
Eriksson, B. 1982. Data rörande Sverigestemperaturklimat. SMHI Reports Meteorology andClimatology RMK 39. Stockholm.
Gregory, S. (ed.) 1988. Recent climatic changes. BelhavenPress, London.
Grimås, U. 1979. The Biotest Basin of the Forsmark nuclearpower plant, Sweden. In: Methodology for assessingimpacts of radioactivity on aquatic ecosystems.Technical Reports Series No. 190, pp. 217-226. IAEA-AG-134/10, IAEA, Vienna.
Hartley, B. 1986. A check-list of the freshwater, brackishand marine diatoms of the British isles and adjoiningcoastal waters. J. Mar. Biol. Ass. U. K. 66: 531-610.
Hendey, N. I. 1964. An introductory account of the smalleralgae of Bri t ish coastal waters. Part V:Bacillariophyceae (diatoms). Fishery Investig. Ser.IV: 1-317.
Hudon, C. & Legendre, P. 1987. The ecologicalimplications of growth forms in epibenthic diatoms.J. Phycol. 23: 434-441.
Jongman, R. H. G., ter Braak, C. J. F. & van Tongeren, O.F. R. 1987. Data analysis in community and landscapeecology. Pudoc, Wageningen.
Keskitalo, J. & Heitto, L. 1987. Overwintering of benthicvegetation outside the Olkiluoto nuclear power station,west coast of Finland. Ann. Bot. Fenn. 24: 231-243.
Kuylenstierna, M. 1982. Massuppträdande av kiselalger ikylvattenutsläppet vid Ringhals kärnkraftverk.Rep. Dept. Quaternary Geology, Lund 22: 1-13.
Launiainen, J. & Koljonen, J. 1981. Variations oftemperature and salinity at Finnish fixed hydrographicstations in the Baltic Sea. 69th ICES Statutory Meeting,5-14 October 1981, Woods Hole, USA, C.M. 1981/C:25. (mimeogr.)