Embed Size (px)
Citation preview

Bioelectromagnetics 32:140^147 (2011)
Effects of Static Magnetic Fields on theGrowth of VariousTypes of Human Cells
Katherine Sullivan, Arthur K. Balin,* and Robert G. AllenThe LongevityAchievement Foundation, Media, Pennsylvania
The effects of a static magnetic field (SMF) on the proliferation of various types of human cells weredetermined. All cultures were maintained at 37 8C throughout the experiment. SMF was generated byplacing two magnets oppositely oriented on either side of a T25 flask. The flux density in the flaskranged from35 to 120mT.Growth curveswere constructed by plotting cell number at 18 h and 4, 7, 11,and 14 days after seeding, with the 18-h point being a measure of attachment efficiency. Exposure toSMF significantly decreased initial attachment of fibroblasts and decreased subsequent growthcompared to sham-exposed control. Significant effects were observed in both fetal lung (WI-38) andadult skin fibroblasts, but theywere generally larger in the fetal lung fibroblast line. SMF did not affectattachment of human melanoma cells, but inhibited their growth by 20% on day 7. SMF producedno effects in a human adult stem cell line. Oxidant production increased 37% inWI-38 cells exposed toSMF (230–250mT) during the first 18 h after seeding, when cell attachment occurs. Conversely, noelevation in oxidant levels was observed after a prolonged 5-day exposure. These results indicate thatexposure to SMF has significant biological effects in some, but not all types of human cells.Bioelectromagnetics 32:140–147, 2011.� 2010 Wiley-Liss, Inc.
Key words: adult stem cells; cell growth; fibroblasts; melanoma; static magnetic field
INTRODUCTION
Exposures to lowandmoderate intensitymagneticfields in the workplace have increased progressivelyduring the last several decades. This has raised publichealth concerns. Moderate intensity static magneticfields (SMF) have been reported to influence some typesof human cells as well as cells from several otherspecies [Leszczynski, 2005; Miyakoshi, 2005; Saun-ders, 2005]; however, an examination of the existingliterature reveals little consensus on whether magneticfields influence the growth of human cells. Some typesof cells appear to be more resistant to the influence ofmagnetic fields. For example, HeLa cells are reported tobe essentially unaffected by exposure to a gradient SMF[Sato et al., 1992], while at least a transient effect isobserved in human skin fibroblasts [Pacini et al., 2003].Conversely, Raylman et al. [1996] reported diminishedgrowth in lymphoma, melanoma, and ovarian carci-noma placed in a strong SMF. The parameters used toquantify SMF effects on living cells vary greatly.
It is noteworthy that few studies that reportcell growth as a parameter of SMF effects actuallydirectly measure changes in cell number per unit time.Instead, thymidine incorporation [McDonald, 1993;Pacini et al., 2003], bromodeoxyuridine (BrdU) uptake[Aldinucci et al., 2003], colony formation [Pacini et al.,2003], and indicator dye conversion [Tenuzzo et al.,2006] have been used as indicators of cell growth. Sato
et al. [1992] counted cells but normalized the number ofattached cells at the time the cultures were exposed to amagnetic gradient. This suggests that cells were alreadyattached prior to exposure to magnetic fields and thusany effects on attachment could not be observed.Although all of these experiments achieved their goalswith the parameters used, a more complete analysis ofthe changes in the growth kinetics of cells stimulated bySMF may provide further insight into the underlyingnature of SMF effects on living cells.
The underlying causes ofmagnetic field effects oncell growth, when they occur, remain unclear. Changesin Ca2þ have been observed in Jurkat cells exposed
�2010Wiley-Liss, Inc.
——————Grant sponsors: Pennsylvania Department of Health, SAPnumber: 4100037834; Longevity Achievement Foundation(Media, PA)
The Department specifically disclaims responsibility for anyanalyses, interpretations or conclusions.
*Correspondence to: Arthur K. Balin, The Longevity Achieve-ment Foundation, 110 Chesley Drive, Media, PA 19063.E-mail: [email protected]
Received for review 18 January 2010; Accepted 15 September2010
DOI 10.1002/bem.20624Published online 17 November 2010 in Wiley Online Library(wileyonlinelibrary.com).

to SMF [Aldinucci et al., 2003]. Several reports havelinked the effects of SMF to oxidative damage [Koanaet al., 1997; Suzuki et al., 2001; Zhang et al., 2003]. Ithas also been reported that electromagnetic field (EMF)or extremely low frequency (ELF)-EMF stimulateschanges in cellular antioxidant defenses and redoxpotential as well as increasing oxidative damage [Wolfet al., 2005; Zwirska-Korczala et al., 2005; Simko,2007; Fitzsimmons et al., 2008]. This suggests thatmagnetic fields increase the rate at which cells generateoxygen free radicals and peroxide, collectively termedreactive oxygen species (ROS).
The present study was conducted to more system-atically analyze the effects of SMF on the growth ofseveral types of human cells. The analysis includes bothinitial cell attachment and subsequent growth rate.Because it was previously implicated as an underlyingcause of magnetic field effects on cells, we also analyz-ed the effects of SMF on the rate of ROS generation infetal human lung fibroblast cells (WI-38).
METHODS
Cell Lines
The human diploid embryonic lung fibroblast cellline WI-38 was obtained from Dr. Vincent J. Cristofaloof the Lankenau Center for Medical Research, Wynne-wood, PA. The adult skin fibroblast line AG11020 wasobtained from the National Institute of Aging Cell Re-pository at the Institute forMedical Research, Camden,NJ. The adult adipose stem cell line SBMC046 wasestablished at the Sally Balin Medical Center, Media,PA from adipose tissue discarded following a standardliposuction procedure using the method of Zuk et al.[2001]. Briefly, the adipose tissue was washed withPBS and then treated with collagenase to release theembedded stem cells. Stem cells were collected bycentrifugation. The human melanoma line LIDRU 80was obtained from the Laboratory for InvestigativeDermatology, Rockefeller University, New York, NY.
Cell Culture
All cell lines used in this study were routinelytested for mycoplasma infection using a kit from IntronBiotechnology (distributed by Abbott Laboratories,Abbott Park, IL) and were found to be uncontaminated.Cell viability is routinely tested in our laboratory usingtrypan blue dye exclusion [Freshney, 1994]. All mortalcell lines used in this study had completed <50% oftheir lifespan when the experiments were performed.WI-38 cells were at population density levels (PDLs) of19–22 (of about 65 possible); AG11020 were at PDLsof 17–19 (the lifespan of this line varies between 48 and
66); and stem cells were at PDLs of 2–3, but theyseldom double more than 22 times. Unless otherwisestated, all reagents and media were purchased fromSigma–Aldrich (St. Louis, MO). Stem cells weregrown in Dulbecco’s modified Eagle’s medium(DMEM, 1 g/L glucose) [Morton, 1970] supplementedwith 10% fetal bovine serum (FBS; Atlanta Bio-logicals, Lawrenceville, GA). AG11020 and WI-38cells were grown in Minimum Essential Medium(MEM). Immediately before use, the medium wassupplemented with L-glutamine (2mM; Flow Labora-tories, Rockville, MD) and FBS (10%, v/v). In eachexperiment, all of the flasks for each line were seededfrom a single pool of cells to increase uniformity. In allcases, medium was equilibrated with a gas mixture thatcontained 5% O2, 5% CO2, and 90% N2. The partialpressure of oxygen and carbon dioxide in the mediumin each flask was measured at harvest with a bloodgas analyzer (Model 113, Instrumentation Laboratory,Lexington,MA). The average oxygen tension at harvestwas 40mm Hg or 4.7% oxygen in WI-38 cultures.38mmHg in melanoma cultures, 42mmHg in culturesof SBMC046, and 37mm Hg in cultures of AG11020.Antibiotics were not used. Cultures were grown at37 8C in 25 cm2 sealed polystyrene flasks (No. 3024,Falcon Labware, Oxnard, CA) containing 10mlmedium.The atmosphere of themammalian cell cultureincubator was humidified and contained 5% CO2. Thecells were released from the plastic with trypsin(0.25%) in Ca2þ- and Mg2þ-free MEM. After suspen-sion in medium containing 10% FBS, the cells werecounted and inoculated into appropriate vessels at adensity of 104 cells/cm2. A Coulter Counter (CoulterElectronics, Hialeah, FL) was used for cell counts.
Cell Attachment
Increases in the number of cells anchored to thegrowing surface during the initial period followingseeding are the result of physical attachment ratherthan growth [Freshney, 1994]. Depending on the celltype, most of the cells attach to the flask within 6 h afterseeding; however, some cells may require longerperiods of time. The level of cellular attachment isusually determined by counting the number of attachedcells at 18 h after seeding [Freshney, 1994; Cristofaloet al., 2000]. In this study, the initial cell count taken18 h after seeding was used as the measure of attach-ment.
Growth Curves
Growth curves were constructed by counting cellsafter attachment at 18 h, and 4, 7, 11, and 14 days, andplotting log of cell number per cm2 versus time. Lidru80 (melanoma) cells were counted only at 18 h and
SMFand Growth of Human Cells 141
Bioelectromagnetics

days 4 and 7 because beyond this time cells tended tobecome unattached and skew results. The 18 h attach-ment point is used as the reference for growth ratherthan the number of cells seeded because not all of thecells attach. Three flasks treated with SMF and threesham-treated controls were counted at each time pointand the averages were compared statistically in eachexperiment. Controls were sham-exposed simultane-ously in an identical incubation chamber and treatedexactly the same way as exposed cells, except thatmagnets were not applied to these flasks. Additionally,each experiment was repeated three times (total of four)with similar results. Variation within one experimentusing one subclone was usually <5%, while the varia-tion between experiments was usually no more than10%.
Population Doubling Level (PDL)
In one case, the population culture’s age wasexamined. Cells were counted and the fold increasewas calculated as the number of cells obtained from theculture vessel divided by the number of cells seeded.The change in population doubling level (DPDL) wascalculated as DPDL¼ log (fold increase in cellnumber)� (log 2). The cumulative change in popula-tion doubling level throughout the lifespan of thecultures is a measure of the proliferative lifespan.
Proliferative Senescence
During the expansion that produced cultures thathad progressed to different points in the proliferativelifespan, cultures that had progressed to the desiredpoint were not further subcultivated and were insteadrefed with MEM that contained 0.5% serum. Thismaintained the cells in a state of growth arrest until all ofthe cultures at various points in their lifespanwere readyfor assay. At least 24 h prior to use the cells were refedwith MEM that contained 10% FBS.
Magnetic Fields
Magnetic fields were generated by placing onemagnet above the flask and one below, with theirmagnetic poles oppositely oriented (Part No. BY0X02,K& JMagnetics, Jamison, PA). The attraction betweenthe two magnets was great enough to hold the magnetsin place without further support. The dimensions of themagnets used in this study were 5.1 cm� 2.5 cm�0.3 cm; the surface area of each large flat side was thus12.75 cm2. These magnets were fabricated so that theentire flat surface of each side was a pole. North andsouth poles were determined using a magnetic poleidentifier (K& JMagnetics). The magnetic flux densitynear the lower growth surface inside the flask wasmeasured at 120mT using a DC magnetometer
(AlphaLab, Salt Lake City, UT). The growth surfaceof the flasks used was 4.8 cm wide. Because the flaskswerewider than the magnets, some of the cells attachedand grew in areas not directly over the magnet. Thefield strength in these areas (12.25 cm2) ranged from35 to 52mT. In one study, the magnetic field near thegrowth surface was decreased by placing a secondflask between the magnet and the culture vessel, thusincreasing the distance between the magnets and thecells. Magnetic flux at the growth surface was 38mTover the magnet in those flasks, and ranged from 21 to35mT in the growth area not directly over the magnet.The orientation of the north and south poles was variedto determine whether any differences existed in theireffects on cell growth. Magnets pairs were placed sothat half of the flasks had the north pole near the celllayer and half had the south pole near the cell layer. Instudies that did not compare the effects of north andsouth poles on cell proliferation, magnet pairs wereoriented with the north pole nearest to the growthsurface of the culture vessel.
Magnetic Fields for 96-Well Culture Plates
Twelve 5.1 cm� 2.5 cm� 0.3 cm magnets werefixed with epoxy to two steel plates (six magnets perplate) to permit exposure of cells in 96-well plates to anSMF. The poles were alternated between north andsouth to make it possible for the magnets to be alignedon the steel plate and not repel one another. The 96-wellplate was placed over the magnets, and a second steelplate with magnets oppositely oriented to the first platewas placed on top to create an SMF between the twosteel plates. The dimension of the magnetic surfacewas7.5 cm� 10 cm. This was not enough to cover the entire96-well plate; however, onlywells thatwere completelycovered by the magnets and that gave consistentreadings were used in the study. Ultimately, measure-ments were taken from only 72 of the 96 wells. Theprobe end measured 2.2 cm� 0.5 cm� 0.1 cm but itwas flexible enough to allow it to be shaped to lie onthe well floor (final dimensions of the probe tip were0.5 cm� 0.4 cm� 0.1 cm). Reshaping the probe de-creased readings by 15% and the values reported herehave been corrected for this. The magnetic flux densitynear the lower growth surface inside the plate wasmeasured at 232–252mT using a DC magnetometer.
Reactive Oxygen Species (ROS) Generation
20,70-dichlorofluorescin diacetate (DCFH-DA)was initially solubilized in 250 ml dimethyl sulfoxide(DMSO) and then added to 150ml phenol red—serum-free MEM. WI-38 cells were seeded in a black 96-wellviewplate (Perkin Elmer, Waltham, MA) at 10000 cellsper well. Steel plates with attached magnets were then
142 Sullivan et al.
Bioelectromagnetics

placed above and below the culture plate. The magnets’surface, not the steel plate, was always placed nearestthe cells. Cells were returned to the incubator andallowed either to attach for 18 h or to grow for 5 daysprior to measuring ROS generation. The cells werewashed once with phenol red—serum-free MEM andwere then treated with 100 ml medium containingDCFH-DA. The plates were then incubated at 37 8Cfor 1 h. Following incubation, the cells were washedonce with the wash buffer, and read (ex 494, em 518)with a Victor 2 plate reader (Perkin Elmer). Filters usedwere ex 485 (PerkinElmer Filter 11440022) and em535(Perkin Elmer Filter 11440023). Protein in the sampleswas determined using Bradford protein assay reagent(Bio-Rad, Hercules, CA). The reagent was diluted 1:5with ultrapure deionized water and filtered prior totreating samples. The Victor 2 plate reader was used todetermine absorbance at 595 nm. Standard curves wereprepared for each assay from known concentrations of20,70-dichlorofluorescein (DCF) and protein. Theseexperiments were repeated twice (total of three).
Statistics
Data were compared using either Student’s t-testor analysis of variance (ANOVA). In all cases, P< 0.05was considered statistically significant.
RESULTS
Cell Attachment and Growth
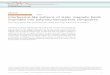
Exposure of WI-38 cells to magnetic fields atseeding (104 cells/cm2) significantly decreased thenumber of cells that were attached to the flask after18 h (ANOVA, P< 0.001; Fig. 1A). Cell numbers incultures treated with SMF also significantly laggedbehind the sham-treated control on all days subsequentto attachment (ANOVA, P< 0.001 in all cases;Fig. 1B). Trypan blue exclusion revealed no differencesin viability between sham-treated controls and SMF-treated cultures. Because exposure to SMF decreasedattachment, it was possible that the lower cell countsobserved after the initial 18 h measurement wereactually the result of the growth phase of cultures start-ing from smaller populations rather than a manifes-tation of a slower growth rate. In order to resolve this,cultureswere not exposed to SMFuntil after attachmenthad occurred. In this case, cells were seeded andallowed to attach for 18 h prior to application of themagnets to the flasks. When magnets were not appliedto cultures until after cell attachment, growth rate wasalso diminished (ANOVA,P< 0.02 at all points includ-ing days 4, 7, 11, and 14; data not shown). In bothstudies, the orientation of the magnets, that is, whichpole was nearest the cells, was irrelevant (Fig. 1B).
Similar effects were observed in the adult skinfibroblast line AG11020, although treated culturesultimately reached cell densities similar to controlsby day 14 (Table 1). The fact that SMF-treated adultskin fibroblast cultures ultimately reached control celldensities was not due to changes in growth rate; instead,the control cultures that exhibited a more rapid initialgrowth rate became contact inhibited and stoppedgrowing 2–3 days before treated cultures becamecontact inhibited. No effects were observed on theattachment of non-pigmented melanoma cells (LIDRU80), but subsequent growth of this line was greatlydiminished. By day 7, SMF-treated cultures had 20%fewer cells than controls (Table 1). No effect on eitherattachment or growth was observed in adipose-derivedadult stem cells (SBMC046; Table 1).
Proliferative Age
Factors other than cell typemay also affect results.Mortal cells such asWI-38 cells maintained in a culture
0
2000
4000
6000
8000
Control North South
Cel
ls/c
m2
* *
18 hr. day 4 day 7 day 11 day 14
0
40000
80000
120000Control
SouthNorth
Cel
ls/c
m2
*
* **
*
* *
*
* **
*
A
B
Fig. 1. EffectsofstaticmagneticfieldsonattachmentandgrowthofWI-38 cells. Cells were exposed to magnetic fields from the timethey were seeded.A: Cell attachment after18 h exposure to SMF.B: Growth curves resulting from application of a strong staticmagnetic fieldtocellsat thetimeofseeding.Statisticallysignificantdifferences between control and treated cultures are indicatedby (*). The pole nearest the bottom flask surface where cellsattachedisindicated.
SMFand Growth of Human Cells 143
Bioelectromagnetics

environment exhibit a limited proliferative capacity.Furthermore, cellular responses to various stimuli maychange as cultures progress through their proliferativelifespan. In order to determine if the responses ofWI-38cells were consistent throughout their proliferativelifespan, we exposed cultures from a single subclone ofWI-38 cells that had been subcultured to various pointsin their lifespan and determined the effects of SMF onattachment. The effects of SMF exposure increased ascell populations progressed through their proliferativelifespan (Table 2). SMF exposure decreased cellattachment by <10% in relatively young cultures andby more than 60% in later passage cultures.
Seeding Density
The effects of SMF on attachment were greaterwhen seeding density was decreased. Figure 2 depictsattachment and growth of WI-38 cells seeded at adensity of 103 cells/cm2. Attachment was decreased
by 52% in SMF-exposed cultures (Student’s t-test,P< 0.002), but the subsequent growth rate was slightlygreater in SMF-exposed cultures. By day 11, cell countsin control and SMF-treated populations were similar.
ROS Generation
The cellular steady-state level of ROS (primarilyH2O2) was determined using DCFH-DA. Because it isnon-polar, cell membranes are permeable to DCFH-DA[Bass et al., 1983]; once in cells, the compound isdesacetylated to 20,70-dichlorofluorescin (DCFH).DCFH is polar and is thus trapped in cells. DCFH isoxidized by ROS to the highly fluorescent DCF [Basset al., 1983].WI-38 cells have been previously analyzedfor their growth characteristics, antioxidant defenselevels, and rates of ROS generation under differentconditions [Allen et al., 1999; Cristofalo et al., 2000]and were therefore selected for this experiment.Analysis of WI-38 cells treated with magnets contin-uously from the time they were seeded revealed a sharpincrease in ROS levels (Student’s t-test, P< 0.00007)after 18 h exposure. The increase was transitory;cultures continuously exposed to SMF for 5 daysexhibited no difference with the control in ROS levels(Fig. 3).
DISCUSSION
Both attachment and growth characteristics ofhuman fibroblast cells can be modulated by exposure toSMF. SMF also affected growth rate of human mela-noma cells, but had no effect on adult adipose stem cellattachment or growth. Although the growth rate of adult
TABLE 1. Effect of a Strong Static Magnetic Field on the Growth of Various Human CellLines
Cell line Time CTRL 35–120mT
Lidru 80 (melanoma) 18 h 1157� 37 1250� 35**Day 4 1655� 184 1367� 84***Day 7 1988� 183 1630� 38*
SBMC046 (adult adipose stem cell line) 18 h 9540� 134 9314� 140Day 4 17042� 571 16282� 406Day 7 18713� 1091 18080� 1858Day 11 19612� 339 19301� 483Day 14 21992� 1249 23291� 580
AG11020 (adult skin fibroblasts) 18 h 9543� 410 8802� 76****Day 4 18716� 281 17673� 250***Day 7 21630� 707 19835� 371*Day 11 21335� 205 20343� 263***Day 14 20482� 647 19947� 518
Number of cells/cm2 of growing surface� standard deviation; Student’s t-test.*P< 0.01.**P< 0.02.***P< 0.03.****P< 0.04.
TABLE 2. Effects of Proliferative Age (Population DoublingLevel) and SMF on Cell Attachment
Population doublinglevel (PDL)
% Lifespancompleteda
% Controlattachmentb
29 47 9133 53 8736 58 8643 69 6953 85 37
aMaximum PDL for this subclone was 62.bThe number of attached cells was counted after 18 h at variousproliferative ages. Results reflect comparison to sham-treatedcontrols at the same proliferative age.
144 Sullivan et al.
Bioelectromagnetics

fibroblasts was affected by SMF exposure, the effectwas smaller than the effects observed in either fetalfibroblasts or melanoma cells. Seeding density andproliferative age strongly influenced the attachment ofWI-38 cells exposed to SMF.
As noted above, few studies that report cell growthas a parameter of SMFeffects actuallymeasure changesin cell number per unit time, and most of those that didgave little attention to effects of magnetic fields oninitial attachment. Most have relied on various indirect
cell or biochemical assays to quantify changes in cellnumbers [McDonald, 1993; Aldinucci et al., 2003;Pacini et al., 2003; Tenuzzo et al., 2006]. Similarly,investigators who have examined the effects of EMF orELF-EMF on growth have usually either indirectlydetermined growth rate (via dye uptake) [Loberg et al.,2000] or failed to examine effects on early parameterssuch as cell attachment [Schwenzer et al., 2007; Denaroet al., 2008]. One notable exception was a studyreported by Nakahara et al. [2002], who exposed CHO-K1 cells to a 10T SMF immediately after cells wereseeded and counted cells on each of the first 4 daysfollowing seeding, but they found no effect on attach-ment or growth in this type of cell. They also examinedcultures for the distribution of cells in the phases of cellcycle, but observed no effects from exposure to SMF.
The results of the present study demonstrated astatistically significant increase in ROS production innewly seeded WI-38 cultures that were immediatelyexposed to a SMF. When cells were continuouslyexposed to the SMF for several days, no difference inROS levels were observed. The initial increase in ROSgenerationmay account for the relatively large decreasein attachment observed in WI-38 cells. This cell lineis extremely sensitive to oxidative stress, and lowerseeding densities exacerbate the toxic effects of oxygen[Balin et al., 1984]. Thus, the observation that SMFmore strongly affected cells seeded at lower density alsosupports the hypothesis that increased ROS generationwas the underlying cause.
A number of other investigators reported thatexposure to either SMF or ELF-EMF stimulated ROSgeneration [Suzuki et al., 2001; Zhang et al., 2003;Wolf et al., 2005]. Wolf et al. [2005] also reportedthat ELF-EMF stimulated a spike in ROS generation inWI-38 cells as well as rat 1 and HeLa cells. Exposure toELF-EMF has also been observed to decrease anti-oxidant defense levels [Zwirska-Korczala et al., 2005],which would be expected to result in a rise in steady-state ROS levels.
Exposure to magnetic fields stimulates the accu-mulation of oxidative damage. Wolf et al. [2005]observed that exposure to a 50Hz ELF magnetic fieldinduced DNA damage. Similarly, Zhang et al. [2003]found that exposure of E. coli to a strong SMF of 9Tinduced statistically significant levels of DNA damage.Suzuki et al. [2001] found statistically significantincreases in micronucleus formation in mouse bonemarrow following exposure of mice to a SMF of 4.7 T.DNAdamage observed following exposure tomagneticfields results from increased oxidation reactions sincethe antioxidant tocopherol can effectively block theseeffects [Koana et al., 1997; Wolf et al., 2005]. Faloneet al. [2007] discovered that exposure of neuroblastoma
Fig. 2. Low seeding density modulates the effects of static mag-netic fieldsonattachmentandgrowthofWI-38cells.Thenorthpolewas placed on the bottom flask surface where cells attached.Magnets were added at the time of seeding. Attachment is indi-cated by the measurement at 18 h. Each point is the average ofmeasurementsmade on three independent cultures. Statisticallysignificant differencesare indicatedby (*).
*
MagnetControlMagnetControl0.0
0.1
0.2
0.3
0.4
5 day18 hr
RO
S (n
mol
/µg
prot
ein)
Fig. 3. Effects of static magnetic fields on oxidant generation inWI-38cells.Cellswereexposedtomagneticfieldsfor18 hor5days,and ROS were measured after 18 h or 5 days. Each point is anaverage of measurements on 72 wells. Statistically significantdifferencesisindicatedby (*).Errorbarsarestandarddeviations.
SMFand Growth of Human Cells 145
Bioelectromagnetics

cells to ELF-EMF increased glutathione, which causeda shift in the cellular environment toward a morereduced state. However, mortal (non-cancer) cellsfrequently respond to oxidative stress by increasingglutathione synthesis [Allen et al., 1999]. In fact, amorerecent study of the effects of ELF-EMF on developingrat cortical neurons revealed a positive correlationbetween oxidant generation and reduced glutathioneconcentration [Di Loreto et al., 2009]. Thus, the resultsreported by Falone et al. [2007] are consistent withincreased oxidative stress.
Our results showed considerable variation in themagnitude of the response of different cell types to aSMF. Slower growing cell lines tended to be lessaffected. The fact that different types of cells responddifferently to SMF exposure probably results frominherent differences in their physiology. Simko [2007]postulated that it was differences in the cellular redoxstate that accounted for variations observed when dif-ferent types of cells are exposed to magnetic fields. Infact, large differences in the cellular redox state exist indifferent types of cells. Undifferentiated and immorta-lized cells both tend to have higher glutathione con-centrations than mortal differentiated cells [Allen andBalin, 1989, 2003].More rapidly growing cell lines alsotend to exhibit greater concentrations of glutathione[Allen and Balin, 1989; Huang et al., 2001]. In thepresent study, the growth rates of fetal fibroblasts andmelanoma were the most strongly affected by SMF,while mortal cell lines derived from adults were leastaffected. On the other hand, cell attachment wasaffected only in fibroblasts including both fetal andadult cells. Glutathione levels were extremely high inmelanoma cells compared to either of the fibroblastlines examined [Allen and Balin, 2003]. Since attach-ment of melanoma cells was unaffected by exposure toSMF, it is possible that the more reducing environmentof these cells was sufficient to protect them fromincreased ROS generation.
These studies reveal that exposure of humanfibroblasts to SMF causes diminished attachment andgrowth. Factors such as proliferative age and seedingdensity strongly influence the effect of SMF on attach-ment. The effects of SMF are not universal. SMF haslittle effect on attachment in melanoma cells, butgrowth rate is most strongly affected in these cells.Adult fibroblasts are less affected than fetal fibroblasts,and adult adipose stem cells are unaffected by SMF.SMF exposure stimulates ROS production in WI-38cells during the first 18-h period when cells are attach-ing to the culture vessel. These results support the hy-pothesis that increased ROS formation may account forSMF effects on attachment. However, SMF decreasesgrowth inWI-38 cells long after the increase inROShas
abated, suggesting that other mechanisms account forSMF effects on growth.
REFERENCES
Aldinucci C, Garcia JB, Palmi M, Sgaragli G, Benocci A, Meini A,Pessina F, Rossi C, Bonechi C, Pessina GP. 2003. The effectof strong static magnetic field on lymphocytes. Bioelectro-magnetics 24(2):109–117.
Allen RG, Balin AK. 1989. Oxidative influence on developmentand differentiation: An overview of a free radical theory ofdevelopment. Free Radic Biol Med 6(6):631–661.
Allen RG, Balin AK. 2003. Effects of oxygen on the antioxidantresponses of normal and transformed cells. Exp Cell Res289(2):207–316.
Allen RG, Tresini M, Keogh BP, Doggett DL, Cristofalo VJ. 1999.Differences in electron transport potential, antioxidantdefenses, and oxidant generation in young and senescentfetal lung fibroblasts (WI-38). J Cell Physiol 180(1):114–122.
Balin AK, Fisher AJ, Carter DM. 1984. Oxygen modulates growthof human cells at physiologic partial pressures. J Exp Med160:152–166.
Bass DA, Parce JW, DeChatelet LR, Szejda P, Seeds MC, ThomasM. 1983. Flow cytometric studies of oxidative productformation by neutrophils: A graded response to membranestimulation. J Immunol 130:1910–1917.
CristofaloVJ,VolkerC,AllenRG. 2000.Use of the fibroblastmodelin the study of cellular senescence. In: Barnett Y, Barnett CR,editors. Aging methods and protocols. Totowa, New Jersey:Humana Press. pp. 23–52.
DenaroV, PapapietroN, SgambatoA,Barnaba SA,Ruzzini L, PaolaBD, Rettino A, Cittadini A. 2008. Periprosthetic electro-chemical corrosion of titanium and titanium-based alloys as acause of spinal fusion failure. Spine 33(1):8–13.
Di Loreto S, Falone S, Caracciolo V, Sebastiani P, D’AlessandroA, Mirabilio A, Zimmitti V, Amicarelli F. 2009. Fifty hertzextremely low-frequency magnetic field exposure elicitsredox and trophic response in rat-cortical neurons. J CellPhysiol 219(2):334–343.
Falone S, Grossi MR, Cinque B, D’Angelo B, Tettamanti E, CiminiA, Di Ilio C, Amicarelli F. 2007. Fifty hertz extremely low-frequency electromagnetic field causes changes in redox anddifferentiative status in neuroblastoma cells. Int J BiochemCell Biol 39(11):2093–2106.
Fitzsimmons RJ, Gordon SL, Kronberg J, Ganey T, Pilla AA.2008. A pulsing electric field (PEF) increases humanchondrocyte proliferation through a transduction pathwayinvolving nitric oxide signaling. J Orthop Res 26(6):854–859.
Freshney RI. 1994. Culture of animal cells. New York: John Wileyand Sons. 486 p.
Huang ZZ, Chen C, Zeng Z, Yang H, Oh J, Chen L, Lu SC. 2001.Mechanism and significance of increased glutathione level inhuman hepatocellular carcinoma and liver regeneration.FASEB J 15(1):19–21.
Koana T,OkadaMO, IkehataM,NakagawaM. 1997. Increase in themitotic recombination frequency inDrosophilamelanogasterby magnetic field exposure and its suppression by vitamin Esupplement. Mutat Res 373(1):55.
Leszczynski D. 2005. Rapporteur report: Cellular, animal andepidemiological studies of the effects of staticmagnetic fieldsrelevant to human health. Prog Biophys Mol Biol 87(2–3):247–253.
146 Sullivan et al.
Bioelectromagnetics

Loberg LI, Engdahl WR, Gauger JR, McCormick DL. 2000. Cellviability and growth in a battery of human breast cancer celllines exposed to 60Hz magnetic fields. Radiat Res 153(5 Pt2):725–728.
McDonald F. 1993. Effect of static magnetic fields on osteoblastsand fibroblasts in vitro. Bioelectromagnetics 14(3):187–196.
Miyakoshi J. 2005. Effects of static magnetic fields at the cellularlevel. Prog Biophys Mol Biol 87(2–3):213–223.
Morton HJ. 1970. A survey of commercially available tissue culturemedia. In Vitro 6(2):89–108.
Nakahara T, Yaguchi H, Yoshida M, Miyakoshi J. 2002. Effects ofexposure of CHO-K1 cells to a 10-T static magnetic field.Radiology 224(3):817–822.
Pacini S, Gulisano M, Peruzzi B, Sgambati E, Gheri G, Gheri BrykS, Vannucchi S, Polli G, Ruggiero M. 2003. Effects of 0.2 Tstatic magnetic field on human skin fibroblasts. CancerDetect Prev 27(5):327–332.
Raylman RR, Clavo AC, Wahl RL. 1996. Exposure to strong staticmagnetic field slows the growth of human cancer cellsin vitro. Bioelectromagnetics 17(5):358–363.
Sato K, Yamaguchi H, Miyamoto H, Kinouchi Y. 1992. Growthof human cultured cells exposed to a non-homogeneousstatic magnetic field generated by Sm-Co magnets. BiochimBiophys Acta 1136(3):231–238.
Saunders R. 2005. Static magnetic fields: Animal studies. ProgBiophys Mol Biol 87(2–3):225–239.
Schwenzer NF, Bantleon R, Maurer B, Kehlbach R, Herberts T,Claussen CD, Rodegerdts E. 2007. In vitro evaluation ofmagnetic resonance imaging at 3.0 tesla on clonogenicability, proliferation, and cell cycle in human embryonic lungfibroblasts. Invest Radiol 42(4):212–217.
Simko M. 2007. Cell type specific redox status is responsiblefor diverse electromagnetic field effects. Curr Med Chem14(10):1141–1152.
Suzuki Y, IkehataM,Nakamura K,NishiokaM,AsanumaK, KoanaT, Shimizu H. 2001. Induction of micronuclei in miceexposed to static magnetic fields. Mutagenesis 16(6):499–501.
Tenuzzo B, Chionna A, Panzarini E, Lanubile R, Tarantino P, DiJeso B, Dwikat M, Dini L. 2006. Biological effects of 6 mTstatic magnetic fields: A comparative study in different celltypes. Bioelectromagnetics 27(7):560–577.
Wolf FI, Torsello A, Tedesco B, Fasanella S, Boninsegna A,D’Ascenzo M, Grassi C, Azzena GB, Cittadini A. 2005.50-Hz extremely low frequency electromagnetic fieldsenhance cell proliferation and DNA damage: Possibleinvolvement of a redox mechanism. Biochim Biophys Acta1743(1–2):120–129.
ZhangQM,TokiwaM,Doi T,Nakahara T, Chang PW,NakamuraN,Hori M, Miyakoshi J, Yonei S. 2003. Strong static magneticfield and the induction of mutations through elevated pro-duction of reactive oxygen species in Escherichia coli soxR.Int J Radiat Biol 79(4):281–286.
Zuk PA, ZhuM,Mizuno H, Huang J, Futrell JW, Katz AJ, BenhaimP, Lorenz HP, Hedrick MH. 2001. Multilineage cells fromhuman adipose tissue: Implications for cell-based therapies.Tissue Eng 7(2):211–228.
Zwirska-Korczala K, Jochem J, Adamczyk-Sowa M, Sowa P,Polaniak R, Birkner E, LatochaM, PilcK, Suchanek R. 2005.Effect of extremely low frequency electromagnetic fields oncell proliferation, antioxidative enzyme activities and lipidperoxidation in 3T3-L1 preadipocytes—An in vitro study. JPhysiol Pharmacol 56 (Suppl 6):101–108.
SMFand Growth of Human Cells 147
Bioelectromagnetics