Embed Size (px)
Citation preview

1 3
Mar Biol (2016) 163:3 DOI 10.1007/s00227-015-2797-3
ORIGINAL PAPER
Effects of starvation on energy metabolism and crustacean hyperglycemic hormone (CHH) of the Atlantic ghost crab Ocypode quadrata (Fabricius, 1787)
A. S. Vinagre1,2 · J. Sook Chung2
Received: 28 July 2015 / Accepted: 7 December 2015 © Springer-Verlag Berlin Heidelberg 2016
OcqCHH cDNA sequence has the typical structure of CHHs, and its expression did not change by starvation. Starvation decreased hemolymphatic glucose, muscular glycogen and hepatopancreatic lipids. This implies that hepatopancreas lipolysis followed by β-oxidation, hepato-pancreas gluconeogenesis and muscular glycogenolysis may be responsible for the energy requirements of O. quad-rata during nutritional stress. Taken together, these results suggest that O. quadrata may be adapted to nutritional dep-rivation for an extended period of time.
Introduction
Crabs of the genus Ocypode (Family Ocypodidae), known as ghost crabs, are conspicuous inhabitants of tropical and subtropical sandy beaches worldwide (Schlachter et al. 2013). Ocypode quadrata (Fabricius 1787) is commonly found on sandy beaches along the Western Atlantic coast, from the state of Rhode Island in the northeastern USA (42°N and 70°W) to the state of Rio Grande do Sul (30°S and 50°W) in Southern Brazil (Melo 1999).
Ocypode quadrata is a top carnivore in a simple filter-feeding-based food web (Fales 1976; Branco et al. 2010). As a facultative scavenger, O. quadrata consumes fresh beached fish for example, and as an active predator, it feeds on live prey such as the mole crabs Emerita talpoidea, coquina clams Donax variabilis and sea turtles eggs/hatch-lings at US sandy beaches (Fales 1976; Wolcott 1978). This flexibility of feeding behavior is also characteristic of other crabs of the genus Ocypode and is influenced by the abun-dance and type of food available at studied sites (Branco et al. 2010; Schlachter et al. 2013).
In temperate beaches, the activities of O. quadrata are related to the seasonal variations, especially in air
Abstract Ocypode quadrata, a crab species found in sandy beaches along the western Atlantic coast, spends the winter months underground without feeding. The main objective of this work was to determine whether O. quad-rata is naturally adapted to food deprivation for a period of time. The effects of starvation on the energy metabolism and the expression levels of crustacean hyperglycemic hor-mone (CHH), known to have an adaptive role in response to many types of environmental stresses, were investi-gated. First, we isolated the full-length cDNA sequence of OcqCHH and localized the cells producing CHH neu-ropeptide in eyestalk ganglia by immunostaining. Second, the levels of OcqCHH transcripts were determined in the fed and starved (15 days) intermolt crabs using qRT-PCR assay. The concentration of carbohydrate and lipids in the hemolymph, muscle and hepatopancreas was measured.
Responsible Editor: H.O. Pörtner.
Reviewed by undisclosed experts.
Electronic supplementary material The online version of this article (doi:10.1007/s00227-015-2797-3) contains supplementary material, which is available to authorized users.
* A. S. Vinagre [email protected]
1 Programa de Pós-Graduação em Ciências Biológicas: Fisiologia, Laboratório de Metabolismo e Endocrinologia Comparada (LAMEC), Department of Physiology, Instituto de Ciências Básicas da Saúde (ICBS), Universidade Federal do Rio Grande do Sul (UFRGS), Rua Sarmento Leite, 500, Porto Alegre, RS CEP 90050-170, Brazil
2 Institute of Marine and Environmental Technology, Columbus Center, University of Maryland Center for Environmental Science, 701 E. Pratt Street, Baltimore, MD 21202, USA

Mar Biol (2016) 163:3
1 3
3 Page 2 of 11
temperature. Whenever the temperature is below 16 °C, the crabs remain inside their burrows (Milne and Milne 1946; Haley 1972; Alberto and Fontoura 1999; Antunes et al. 2010; Corrêa et al. 2014). Since the winter also affects food availability, it was suggested that O. quadrata starves during this period (Wolcott 1978). According to McCue (2010), starvation and fasting are distinct physiological conditions: Fasting is a post-absorptive period, when ani-mals are involved in various activities such as avoiding predators, territorial defense and reproduction; while star-vation is also a post-absorptive status, it occurs when the animal is willing or able to feed; however, there are limita-tions in food resources. In crustaceans, nutritional depriva-tion can occur in their life for different reasons. They can fast as the result of winter torpor or behavioral modifica-tions during spawning/mating and ecdysis despite sufficient food supply, or they can face periods of starvation when there are limitations in food resources (Vinagre and Da Silva 1992; Hervant et al. 1999; Hervant and Renault 2002; Pellegrino et al. 2008, 2013).
In decapod crustaceans, the main hormone responsible for glucose homeostasis is the crustacean hyperglycemic hormone (CHH). This hormone is mainly secreted from the eyestalk X-organ-sinus gland system, particularly in response to stress conditions such as hypoxia, hypo- and hyperthermia, hyper- and hypo-salinity, infection, exer-cise or walking (Chang et al. 1998; Stentiford et al. 2001; Chung and Webster 2005; Chung et al. 2010; Morris et al. 2010; Webster et al. 2012). The increase in the circulating levels of CHH is followed by hyperglycemia possibly via glycogenolysis in muscle and hepatopancreas (Chung et al. 2010; Nagai et al. 2011; Webster et al. 2012; Katayama et al. 2013). This adaptive role of CHH in response to these environmental stresses is well described, whereas few stud-ies have been carried out to define the role of CHH during starvation.
Given that O. quadrata has been considered as a star-vation-adapted (tolerant) species, we aimed to investigate the effects of starvation on energy metabolism and the expression levels of CHH. Therefore full-length cDNA of O. quadrata CHH (OcqCHH, GenBank accession no.
KM052164) was isolated. Then, the levels of OcqCHH transcripts in the eyestalk ganglia and the concentrations of metabolites in hemolymph, muscle and hepatopancreas were measured in starved and fed crabs.
Materials and methods
Animals
Ocypode quadrata were collected in Wachapreague (37.61°N and 75.69°W, VA, USA) and kept overnight at the Eastern Shore Laboratory, Virginia Institute of Marine Sciences (VIMS). The crabs were transported to the Aqua-culture Research Center (ARC), Institute of Marine and Environmental Technology, Baltimore, MD, USA. The crabs were kept in aquaria (19 L) filled with sterilized sand (to eliminate possible contamination of microorgan-isms), provided with a bowl containing artificial seawater at 30 ppt salinity, 12L/12D photoperiod, 25 ± 2 °C. In order to mimic their natural diet, the crabs were fed with clams, krill, shrimp or silversides (Table 1) every other day around 6 p.m. (Weinstein and Full 1998; Tullis and Andrus 2011).
Molecular isolation of the full‑length OcqCHH (Ocypode quadrata crustacean hyperglycemic hormone)
Molecular cloning and sequencing
Eyestalk ganglia were microscopically dissected from ice-chilled crabs and frozen immediately on dry ice. Total RNAs were extracted using Qiazol Reagent (Qiagen), and their integrity and quantity were evaluated using a Nan-oDrop spectrophotometer (Thermo Scientific). One to three micrograms of total RNAs was subject to 5′ and 3′ RACE cDNA syntheses using SMART™ cDNA Amplifi-cation kit (BD Bioscience) by following the manufacturer’s instructions. A two-step PCR assay was carried out using three CHH degenerate primers described in Chung et al. (2009, 2015) (Table 2) by following the procedures as stated (Chung et al. 2009, 2015). In the first step, touch-down (TD) PCR, the annealing temperatures ranged from 54 to 50 °C for eight cycles and 30 cycles at 55 °C, exten-sion temperature 68 °C. In the second step, nested PCR, the annealing temperature was 59 °C and extension tem-perature 72 °C for 35 cycles. The identified PCR band was subject for sequencing as described (Chung et al. 2009, 2015). Based on the initial sequence of OcqCHH, gene-specific primers for obtaining the full-length cDNA were generated (Table 2) and amplified using the procedures described in Chung et al. (2009, 2015), except the anneal-ing temperature (59 °C). The remaining cloning procedures were as stated (Chung et al. 2009, 2015). The putative open
Table 1 Composition of the diet fed to O. quadrata (adapted from the manufactures’ information)
Name Brand Protein% Fat% Fiber% Moisture%
Krill PROSACT 13.9 3.3 0.3 83.6
Silversides PROSACT 13.9 2.8 0.2 78.5
Mysis shrimp HIKARI 10.5 1.0 2.0 85.0
Clam on a half shell
HIKARI 12.8 1.8 1.6 88.7
Average 12.9 2.2 1.0 83.9

Mar Biol (2016) 163:3
1 3
Page 3 of 11 3
reading frame (ORF) sequence of OcqCHH was further confirmed by sequencing the colonies (n = 18) containing the insert that was amplified using a High Fidelity Taq (BD Biosciences).
Sequence analysis of OcqCHH
The nucleotide sequence of OcqCHH cDNA was exam-ined using BLAST (http://blast.ncbi.nlm.nih.gov/). The ORF was found with ORF finder (http://ncbi.nlm.nih.gov/projects/gorf). The presence of the signal peptide of the deduced aa of preproOcqCHH was examined using Sig-nalP 4.0 Server (http://www.cbs.dtu.dk/services/SignalP/). The sequence similarity of the deduced aa sequence was aligned with eyestalk CHHs from other species of crabs deposited in GenBank using clustalδ (http://www.ebi.ac.uk/Tools/msa/clustalo/). A cladogram of putative crabs CHHs was generated using Phylogeny.fr (www.phylogeny.fr) (Dereeper et al. 2008).
Immunolocalization of CHH neuropeptide in O. quadrata eyestalk ganglia
The presence of CHH in the eyestalk ganglia was examined using whole-mount immunohistochemistry as described (Chung et al. 2012). Animals were placed on ice for 10 min prior to dissection of the eyestalk ganglia. The eyestalk ganglia were fixed immediately in Drosophila fixative (Chung et al. 2012) and were incubated with anti-Carcinus CHH serum (Chung and Webster 2004) at a final dilution of 1:1000 in PBS containing 0.5 % Triton X for 1 day at room
temperature. Subsequently, these ganglia were washed, incubated with the goat-antirabbit-IgG labeled with Alexa 448 (Invitrogen) at a 1:100 dilution. The samples were examined using a confocal microscope (Bio-Rad), and images were collected using Z projections (Chung and Zmora 2008; Chung et al. 2012).
Effects of starvation on the metabolism of O. quadrata
Experimental setup
The crabs were individually housed in same size aquaria (19 L) partially filled with sterilized sand as described above (Supplementary Figure 1). During an acclimation period of 10 days, the crabs were fed with clams, krill, shrimp or silversides (Table 1) every other day around 6 p.m. Each day a different food item was fed to the crabs. On the following morning after feeding, the remaining food was removed, fresh seawater was provided, and ~50 % of the sand was replaced with freshly sterilized sand.
After acclimation, five crabs were randomly chosen as the initial group with carapace length (CL) 3.52 ± 0.35 and carapace width (CW) 2.82 ± 0.46. They were placed on ice for 15 min, and hemolymph and tissues were collected. These samples were immediately frozen on dry ice. The rest of the crabs were divided into two groups: fed (n = 11, CL 3.59 ± 0.64; CW 2.98 ± 0.68) and starved (n = 11, CL 3.67 ± 0.50; CW 3.08 ± 0.55). The fed group was kept on the same diet regimen as the acclimation period, while the starved group was kept in the same conditions, but without food. After a 15-day experimental period, the crabs were killed for sample collection. The hemolymphs were col-lected for determining the concentration of glucose, lactate and triglycerides. The hepatopancreas samples were used for measuring the levels of glycogen, triglycerides and glycerol. Muscle samples collected from the meropodite of the second leg (a mixture of white and red muscle fib-ers) were used to determine glycogen concentration. All the crabs used in this study were collected from late spring to early autumn and were at the intermolt stage (Drach and Tchernigovtzeff 1967).
qRT‑PCR assay of OcqCHH expression in eyestalk ganglia
Total RNAs from eyestalk samples were prepared as described. One to two micrograms of total RNAs was sub-jected to cDNA synthesis using PrimeScript™ RT reagent Kit with gDNA Eraser (BD Bioscience) by following the manufacturer’s instructions. The expression of OcqCHH in these cDNA samples was estimated using Fast SYBR® Green Master Mix (Life Technologies) for RT-PCR assay on an Applied Biosystems 7500 Real-Time PCR System
Table 2 Primer sequences used for isolating the full-length CHH cDNA sequence of O. quadrata and qRT-PCR assay
d Degenerate primers
Primer sequences (5′–3′)
CHH dF1 TGYAARGGNGTNTAYGA
CHH dF2 CATRCAYTGNCKRAANAC
CHH dR1 GAYTTYATHGCNGCNGGNAT
Ocq CHH 3F1 CCTCTTCAGCAAACTGGAACACGTTTG
Ocq CHH 3F2 ACTGGAACACGTTTGCGACGACT
Ocq CHH 3F3 CGTACCTCATATGTGGCCAGCGC
Ocq CHH 3F4 (start) ATGACTTCCAGAATGACCTCCG-TAGCAGT
Ocq CHH 5R1 AGGTTGCTGTAGCAGTTTGACCTG
Ocq CHH 5R2 CTGCATGCGCTGGCCACATATGAGGT
Ocq CHH 5R3 CAAACGTGTTCCAGTTTGCTGAAGAGG
Ocq CHH 5R4 (end) TTACTTCTTCCTGCCGACGACTTGTATAG
Ocq RL10 3F1 ACGTGCGGGTGCGCCTTCACCCATTC-CACG
Ocq RL10 5R1 GCGAGAGACGTAAATCTTCTGCCGGC-CAGGG

Mar Biol (2016) 163:3
1 3
3 Page 4 of 11
(Chung et al. 2009, 2015). Each sample cDNA contain-ing 25 ng of total RNA equivalent was assayed in trip-licate, using primers OcqCHHF1 and OcqCHHR4 as template (Table 2). As a reference gene, the levels of ribo-somal protein L10 (OcqRPL10) (GenBank accession no. JX894250.1) were determined in the same sample cDNAs. Both standards of OcqCHH and OcqRL10 were produced similarly, as described in Chung et al. (2009). The data were expressed as copies × µg total RNA−1.
Metabolites analysis
Carbohydrates
Glucose was measured using a hexokinase glucose assay kit (Sigma-Aldrich), and results were expressed as mmol glucose × mL hemolymph−1 (Chung and Zmora 2008; Shi and Chung 2014). The concentration of glycogen in the tis-sue samples was determined as described in Shi and Chung (2014). Briefly, the tissue samples were first hydrolyzed with 2 N HCl, then neutralized with 2 N NaOH and finally diluted with 0.2 M phosphate buffer. The levels of glycogen were determined as glucose equivalents using the same glu-cose kit and expressed as mg glycogen × g tissue−1 (wet weight).
Lactate
Lactate was measured using a Trinity Biotech assay kit (Chung and Zmora 2008), and results were expressed as mmol lactate × mL hemolymph−1.
Lipids
Hepatopancreas triglycerides were determined with a Cay-man Triglyceride Colorimetric Assay kit by following the manufacturer’s protocol. First, the samples were homog-enized and centrifuged (10,000g, 4 °C, 10 min). The super-natant was used to measure triglycerides and glycerol. Glycerol concentration was determined with a Cayman Glycerol Colorimetric Assay kit by following the manu-facturer’s protocol. The data were expressed as mg × dL−1 (hemolymph) and mg × g−1 (wet weight hepatopancreas).
Statistical analysis
Results were expressed as the mean ± standard error of the mean (SE). Before applying the ANOVA test, the homoge-neity of the data was examined with Levene test. Analysis of variance (ANOVA) in one way was performed using the software PASW Statistics version 18.0 compatible with Windows 7 and was followed by Bonferroni post hoc test. OcqRL10 expression and OcqCHH expression were
compared with independent samples t test. Statistical sig-nificance was accepted at P < 0.05 and noted with different letters.
Results
Isolation of the full‑length OcqCHH cDNA
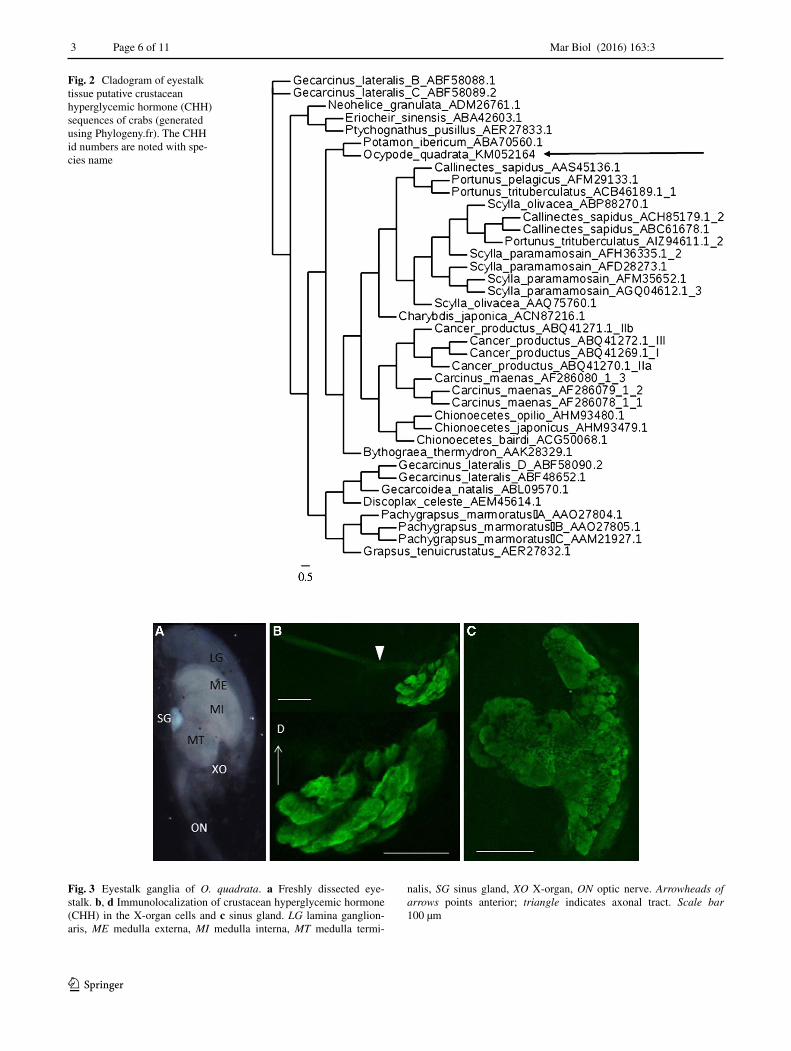
The full-length cDNA (953 nucleotides) sequence of O. quadrata CHH (GenBank accession no. KM052164) is described in Fig. 1. The ORF sequence has 136 amino acids (106–517 nucleotides) and encodes a signal peptide (27 aa); a putative CHH precursor-related peptide (CPRP) (31 aa); a dibasic cleavage site (KR) and the putative CHH (72 aa) with an amidation site (G), a dibasic cleavage site (KK) and a stop codon (taa). As shown in Fig. 2, the puta-tive OcqCHH sequence is grouped with the sequences of other crab species and is most closely related to Potamun ibericum CHH. The alignment of these structures can be consulted in Supplementary File 1.
Immunolocalization of CHH neuropeptide in O. quadrata
Anti-Carcinus CHH serum detected the presence of CHH in the cells (30–50 µm diameter) located in the X-organ, axonal tract (400–500 µm length) and sinus gland (300–400 µm diameter) (Fig. 3). The immunostaining intensity of CHH in the sinus gland (Fig. 3c) appeared higher than that of X-organ neurosecretory cells (Fig. 3d).
Effects of starvation on the metabolism of O. quadrata
No mortalities were recorded during the starvation period.
Expression of OcqCHH in eyestalk ganglia
According to one-way ANOVA, the expression levels of OcqCHH were not affected by starvation [F(1,27) = 1.359, P = 0.276] (Fig. 4). The expression of ribosomal pro-tein 10 (OcqRPL10) showed significant differences [F(1,27) = 6.932, P = 0.004]. Fed crabs had significantly higher amounts of OcqRPL10 than the control group. In the starved crabs, expression of OcqRPL10 was similar to that of control group. When OcqRPL10 and OcqCHH were compared at each treatment, the only difference was between the fed groups (t = −3.815, P = 0.001).
Carbohydrates
The level of glucose in the hemolymph of the starved group decreased in relation to the initial group [F(1,27) = 3.400,

Mar Biol (2016) 163:3
1 3
Page 5 of 11 3
P = 0.047] (Fig. 5a). Although the concentration of gly-cogen in the hepatopancreas (Fig. 6a) decreased by 40 % in the starved group in relation to the initial values, it was not significant [F(1,24) = 3.340, P = 0.055]. In the muscle of the starved crabs, glycogen concentration decreased by 50 % in relation to the initial group [F(1,27) = 4.536, P = 0.021] (Fig. 6b).
Lactate
The concentration of lactate in the hemolymph (Fig. 5b) of the starved crabs did not change in relation to the initial values or the fed group [F(1,27) = 0.050, P = 0.951].
Lipids
The concentration of triglycerides in the hemolymph of the starved crabs (Fig. 5c) did not change in relation to the initial values or the fed group [F(1,25) = 0.574, P = 0.572].
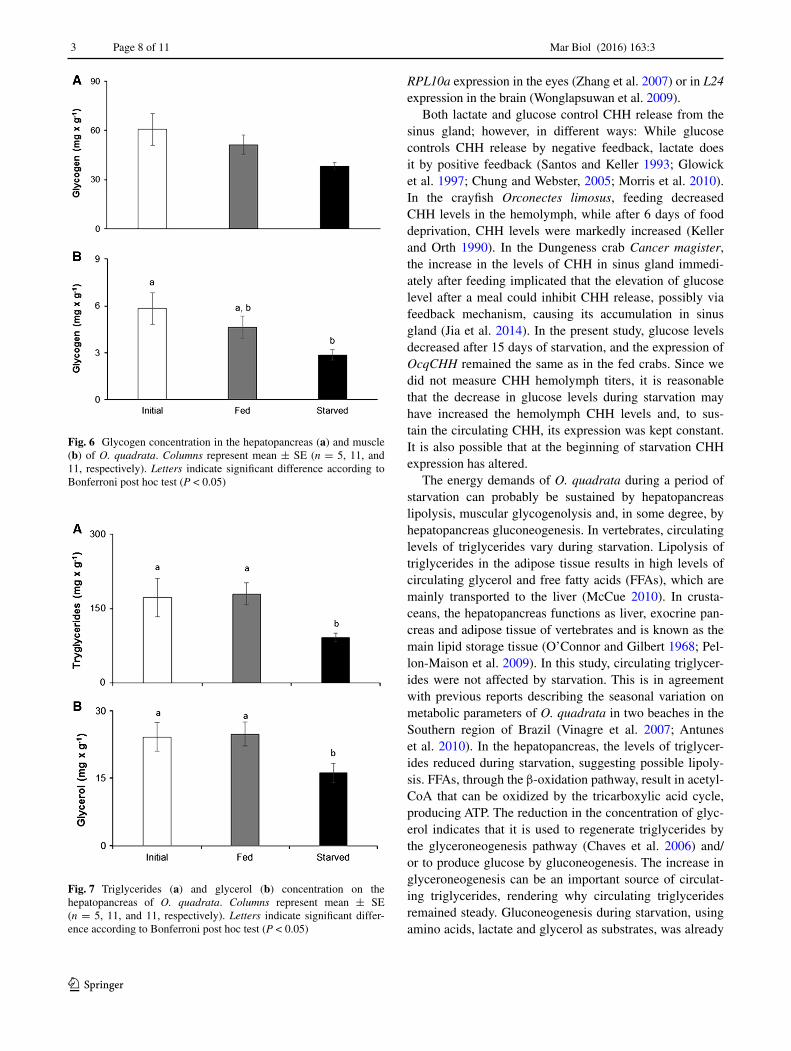
In the hepatopancreas, the concentration of triglycerides [F(1,25) = 5.247, P = 0.014] and glycerol [F(1,27) = 5.191, P = 0.013] (Fig. 7) decreased in the starved crabs in rela-tion to both the initial and fed crabs. In the starved crabs, triglycerides concentration reduced by 50 % in relation to initial values, while the glycerol reduction was by 40 %.
Discussion
The present work describes the effect of 15 days of food deprivation on the metabolism of O. quadrata and proposes that this crab species may be naturally adapted to fasting or starvation. The full-length cDNA sequence of O. quadrata CHH and CHH neuropeptide localization in the eyestalk ganglia are stated, while OcqCHH expression levels after this period of starvation are also described.
The absence of mortality found in this study suggests that this crab is naturally adapted to extended periods
Fig. 1 a The full-length crus-tacean hyperglycemic hormone (CHH) cDNA sequence of O. quadrata (GenBank accession no. KM052164). The numbers on the left-hand side are for nucleotide sequence, and those on the right-hand side are for the putative amino acid sequence of CHH. 5′ UTR (1–105) and 3′ UTR (518–955) are italicized, while open read-ing frame (ORF) is in regular-type letters. Signal peptide (27 aa) is in bold and italicized, the putative CPRP (31 aa) is underlined, the dibasic cleavage site (KR) is boxed, the putative CHH (72 aa) is in bold, as well as the amidation site (G, cir-cled) and a tribasic cleavage site (RKK, underlined). b A sche-matic diagram of the putative preproCHH. Numbers represent the position of the aa: the signal peptide (1–27), CPRP (28–58), dibasic cleavage site (59–60), CHH (61–132), amidation site (133) and tribasic cleavage site (135–136). CPRP crustacean hyperglycemic hormone-related peptide, aa amino acids
Mar Biol (2016) 163:3
1 3
3 Page 6 of 11
Fig. 2 Cladogram of eyestalk tissue putative crustacean hyperglycemic hormone (CHH) sequences of crabs (generated using Phylogeny.fr). The CHH id numbers are noted with spe-cies name
Fig. 3 Eyestalk ganglia of O. quadrata. a Freshly dissected eye-stalk. b, d Immunolocalization of crustacean hyperglycemic hormone (CHH) in the X-organ cells and c sinus gland. LG lamina ganglion-aris, ME medulla externa, MI medulla interna, MT medulla termi-
nalis, SG sinus gland, XO X-organ, ON optic nerve. Arrowheads of arrows points anterior; triangle indicates axonal tract. Scale bar 100 µm

Mar Biol (2016) 163:3
1 3
Page 7 of 11 3
of nutritional deprivation. The ability to withstand and recover from periods of such nutritional stress is an impor-tant adaptation for survival of any organism that must spo-radically endure periods of limited food supply (Comoglio et al. 2005; McCue 2010). The absence of mortality dur-ing an equivalent period of starvation is also described in other decapods such as Astacus lepdodactylus (Malev et al. 2010), Litopenaeus vannamei (Pascual et al. 2006) Paralo-mis granulosa (Comoglio et al. 2005), Lithodes santolla (Comoglio et al. 2008) and Neohelice granulata (Vinagre and Da Silva 1992). The capacity to survive prolonged periods of starvation (180 days) is particularly well devel-oped in groundwater crustaceans (Hervant and Renault 2002; Hervant et al. 1999).
The full-length cDNA sequence of O. quadrata CHH (Fig. 1a) (GenBank accession no. KM052164) contains the typical structural elements described in other decapod pre-proCHHs (Fig. 1b): an ORF including a signal peptide; a putative CPRP, a dibasic cleavage site, the putative CHH, an amidation site and a tribasic cleavage site (Webster et al. 2012; Katayama et al. 2013; Chung et al. 2015). The length and localization of these conserved elements in prepro OcqCHH sequence are in agreement with the analysis of 38 sequences of eyestalk CHHs from other brachyuran species deposited in GenBank (Fig. 2; Supplementary file 1).
CHH is synthesized by neurosecretory somata in the medula terminalis X-organ of the eyestalk ganglia and transported through the axonal tract to the sinus gland, where it is stored until release (Olivaux and Soyez 2000; Chung et al. 2010; Webster et al. 2012). The positive immunostaining with anti-Carcinus CHH serum (Chung and Webster 2004, 2005) was higher in the sinus gland than in the X-organ, suggesting the accumulation of CHH in the sinus gland, as shown also in other decapods (Webster et al. 2012; Chung et al. 2015).
For the qRT-PCR assay, ribosomal protein 10 (RPL10) was used as a reference gene because ribosomal genes are considered housekeeping genes (Zhu et al. 2014; Leone et al. 2015). However, OcqRPL10 expression increased in the fed animals and returned to initial levels after starvation. As shown in other crustaceans, several genes used as refer-ence for qRT-PCR studies also varied during physiological conditions, e.g., molting and reproduction or during hypoxic stress (Chung and Webster 2003; Zmora et al. 2007; Chung and Zmora 2008). In female shrimps Marsupenaeus japoni-cus (Zhang et al. 2007) and Fenneropenaeus merguien-sis (Wonglapsuwan et al. 2009), the expression of RPL24 and RPL10a, respectively, increased in the ovaries during vitellogenesis. However, no differences were recorded in
Fig. 4 Expression levels of O. quadrata crustacean hyperglyce-mic hormone (OcqCHH) (open bars) and ribosomal protein 10 (OcqRPL10) (closed bars). Columns represent mean ± SE (n = 5, 11 and, 11, respectively). Letters indicate significant difference according to ANOVA one-way followed by Bonferroni post hoc test (P < 0.05). Asterisk indicates difference between genes according to t test (P < 0.01)
Fig. 5 Concentration of glucose (a), lactate (b) and c triglycerides in the hemolymph of O. quadrata. Columns represent mean ± SE (n = 5, 11, and 11, respectively). Letters indicate significant differ-ence according to Bonferroni post hoc test (P < 0.05)
Mar Biol (2016) 163:3
1 3
3 Page 8 of 11
RPL10a expression in the eyes (Zhang et al. 2007) or in L24 expression in the brain (Wonglapsuwan et al. 2009).
Both lactate and glucose control CHH release from the sinus gland; however, in different ways: While glucose controls CHH release by negative feedback, lactate does it by positive feedback (Santos and Keller 1993; Glowick et al. 1997; Chung and Webster, 2005; Morris et al. 2010). In the crayfish Orconectes limosus, feeding decreased CHH levels in the hemolymph, while after 6 days of food deprivation, CHH levels were markedly increased (Keller and Orth 1990). In the Dungeness crab Cancer magister, the increase in the levels of CHH in sinus gland immedi-ately after feeding implicated that the elevation of glucose level after a meal could inhibit CHH release, possibly via feedback mechanism, causing its accumulation in sinus gland (Jia et al. 2014). In the present study, glucose levels decreased after 15 days of starvation, and the expression of OcqCHH remained the same as in the fed crabs. Since we did not measure CHH hemolymph titers, it is reasonable that the decrease in glucose levels during starvation may have increased the hemolymph CHH levels and, to sus-tain the circulating CHH, its expression was kept constant. It is also possible that at the beginning of starvation CHH expression has altered.
The energy demands of O. quadrata during a period of starvation can probably be sustained by hepatopancreas lipolysis, muscular glycogenolysis and, in some degree, by hepatopancreas gluconeogenesis. In vertebrates, circulating levels of triglycerides vary during starvation. Lipolysis of triglycerides in the adipose tissue results in high levels of circulating glycerol and free fatty acids (FFAs), which are mainly transported to the liver (McCue 2010). In crusta-ceans, the hepatopancreas functions as liver, exocrine pan-creas and adipose tissue of vertebrates and is known as the main lipid storage tissue (O’Connor and Gilbert 1968; Pel-lon-Maison et al. 2009). In this study, circulating triglycer-ides were not affected by starvation. This is in agreement with previous reports describing the seasonal variation on metabolic parameters of O. quadrata in two beaches in the Southern region of Brazil (Vinagre et al. 2007; Antunes et al. 2010). In the hepatopancreas, the levels of triglycer-ides reduced during starvation, suggesting possible lipoly-sis. FFAs, through the β-oxidation pathway, result in acetyl-CoA that can be oxidized by the tricarboxylic acid cycle, producing ATP. The reduction in the concentration of glyc-erol indicates that it is used to regenerate triglycerides by the glyceroneogenesis pathway (Chaves et al. 2006) and/or to produce glucose by gluconeogenesis. The increase in glyceroneogenesis can be an important source of circulat-ing triglycerides, rendering why circulating triglycerides remained steady. Gluconeogenesis during starvation, using amino acids, lactate and glycerol as substrates, was already
Fig. 6 Glycogen concentration in the hepatopancreas (a) and muscle (b) of O. quadrata. Columns represent mean ± SE (n = 5, 11, and 11, respectively). Letters indicate significant difference according to Bonferroni post hoc test (P < 0.05)
Fig. 7 Triglycerides (a) and glycerol (b) concentration on the hepatopancreas of O. quadrata. Columns represent mean ± SE (n = 5, 11, and 11, respectively). Letters indicate significant differ-ence according to Bonferroni post hoc test (P < 0.05)

Mar Biol (2016) 163:3
1 3
Page 9 of 11 3
demonstrated in other crustaceans (Hervant et al. 1999; Hervant and Renault 2002; Vinagre and Da Silva 2002; Oliveira et al. 2004; Pellegrino et al. 2008). The use of glu-coneogenic amino acids such as alanine or glycine cannot be ruled out, because O. quadrata diet had a good source of protein. During 3 weeks of starvation, the crab N. gran-ulata previously fed with a high-protein diet, kept a high gluconeogenic activity using both glycine (Vinagre and Da Silva 2002) and alanine as precursors (Oliveira et al. 2004).
Lactate is the main end product of anaerobiosis in crus-taceans (Van Aardt 1988; Maciel et al. 2008). Hemolym-phatic lactate levels increase when crustaceans are exposed to hypoxic/anoxic environments (Van Aardt 1988; Chung and Zmora 2008; Maciel et al. 2008) or exercise (Henry et al. 1994; Weinstein and Full 1998; Morris et al. 2010). During starvation, circulating lactate levels can remain constant or oscillate (Pascual et al. 2006; Pellegrino et al. 2008; Malev et al. 2010). In A. leptodactillus, lactate levels in the hemo-lymph decreased after 7 days of starvation but returned to ini-tial levels after 14 days of starvation (Malev et al. 2010). In the present study, circulating lactate levels were not affected by starvation, while muscular glycogen decreased, suggest-ing that glycogenolysis may be important to sustain muscular energy requirements during food deprivation. Muscular gly-cogen levels also decreased during starvation in N. granulata previously fed with a high-protein diet (Vinagre and Da Silva 1992). It was later demonstrated that glycogen synthesis from 14C-glucose remained similar to fed state, while 14C-gly-cine conversion to glycogen decreased, suggesting that the decrease in glycogen concentration can be related to muscu-lar glycogenolysis (Vinagre and Da Silva 2002).
In summary, OcqCHH contains the typical structure of CHHs and its expression levels remain the same after starvation. The results indicate that O. quadrata may be adapted to starvation for a certain period of time during which it probably relies on hepatopancreas lipolysis and gluconeogenesis and on muscular glycogenolysis to sustain its energetic requirements. In a future study, protein metab-olism and the hormonal role of CHH should be investigated by measuring its circulating levels in crabs experiencing different lengths of starvation period.
Acknowledgments The authors gratefully acknowledge Coorde-nação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Brazil, for the senior fellowship to A. S. Vinagre and US National Science Foundation program award to J. S. Chung (No. 1146774). The authors thank P.G. Ross, Richard A. Snyder and all the staff of the Eastern Shore Laboratory, Virginia Institute of Marine Sciences (VIMS), for their assistance with animal collection and all the staff of the Aquaculture Research Center (ARC), Institute of Marine and Environmental Technology (IMET), for the help with tank sys-tem. A.S. Vinagre acknowledges Professor Richard W. Hill for the introduction to VIMS team and Juliana Batista Graceli and Bettina Koepke, for the help during the field trips. We also thank Leah Maurer for revising the manuscript.
References
Aardt Van (1988) Lactate metabolism and glucose patterns in the river crab, Potamonautes warren (Calman), during anoxia and subse-quent recovery. Comp Biochem Physiol A 91:299–304
Alberto RMF, Fontoura NF (1999) Distribuição e Estrutura Etária de Ocypode quadrata (Fabricius, 1787) (Crustacea, Decapoda, Ocy-podidae) em Praia Arenosa do Litoral Sul do Brasil. Braz J Biol 59:95–108
Antunes GF, Amaral APN, Ribarcki FP, Wiilland EF, Zancan DM, Vinagre AS (2010) Seasonal variations in the biochemical composition and reproductive cycle of the ghost crab Ocypode quadrata (Fabricius, 1787) in Southern Brazil. J Exp Zool 313A:280–291
Branco J, Hilleshein JC, Fracasso HAA, Christoffersen ML, Evange-lista C (2010) Bioecology of the ghost crab Ocypode quadrata (Fabricius, 1787) (Crustacea: Brachyura) compared with other intertidal crabs in the southwestern Atlantic. J Shellfish Res 29(2):503–512
Chang ES, Keller R, Chang SA (1998) Quantification of crustacean hyperglycemic hormone by ELISA in hemolymph of the lobster, Homarus americanus, following various stresses. Gen Comp Endocrinol 111:359–366
Chaves VE, Frasson D, Martins-Santos MES, Boschini RP, Garófalo MAR, Festuccia WTL, Kettelhut IC, Migliorini RH (2006) Glyc-eroneogenesis is reduced and glucose uptake is increased in adi-pose tissue from cafeteria diet-fed rats independently of tissue symphathetic innervation. J Nutr 136:2475–2480
Chung JS, Webster SG (2003) Moult cycle-related changes in biologi-cal activity of moult-inhibiting hormone (MIH) and crustacean hyperglycemic hormone (CHH) in the crab, Carcinus maenas: from target to transcript. Eur J Biochem 270:3280–3288
Chung JS, Webster SG (2004) Expression and release patterns of neu-ropeptides during embryonic development and hatching in the green crab Carcinus maenas. Development 131:4751–4761
Chung JS, Webster SG (2005) Dynamics of in vivo release of molt-inhibiting hormone and crustacean hyperglycemic hormone in the shore crab, Carcinus maenas. Endocrinology 146:5545–5551
Chung JS, Zmora N (2008) Functional studies of crustacean hyper-glycemic hormones of the blue crab, Callinectes sapidus—the expression and release of CHH in the eyestalk and pericardial organ in response to environmental stress. FEBS J 275:693–704
Chung JS, Bembe S, Tamone S, Andrews E, Thomas H (2009) Molec-ular cloning of the crustacean hyperglycemic hormone precursor from the X-organ and the identification of the neuropeptide from sinus gland of the Alaskan Tanner crab, Chionoectes bairdi. Gen Comp Endocrinol 162:129–133
Chung JS, Zmora N, Katayama H, Tsutsui N (2010) Crustacean hyperglycemic hormone (CHH) neuropeptides family: func-tions, titer, and binding to target tissues. Gen Comp Endocrinol 166:447–454
Chung JS, Katayama H, Dircksen H (2012) New functions of arthro-pod bursicon: inducing deposition and thickening of new cuticle and hemocyte granulation in the blue crab Callinectes sapidus. PLoS One 7:e496299
Chung JS, Ahn IS, Yu OH, Kim DS (2015) Crustacean hyperglycemic hormones of two cold water crab species, Chionoecetes opilio and C. japonicus: isolation of cDNA sequences and localization of CHH neuropeptide in eyestalk ganglia. Gen Comp Endocrinol 24:177–185
Comoglio L, Smolko L, Amin O (2005) Effects of starvation on oxy-gen consumption, ammonia excretion and biochemical composi-tion of the hepatopancreas on adult males of the false southern king crab Paralomis granulosa (Crustacea, Decapoda). Comp Biochem Physiol B 140:411–416

Mar Biol (2016) 163:3
1 3
3 Page 10 of 11
Comoglio L, Goldsmit J, Amin O (2008) Starvation effects on physi-ological parameters and biochemical composition of the south-ern king crab Lithodes santolla (Molina, 1782). Rev Biol Mar Oceanogr 43(2):245–353
Corrêa MODA, Andrade LS, Costa RC, Castilho AL, Bertini G, Fran-sozo A (2014) Vertical distribution by demographic groups of ghost crab Ocypode quadrata (Crustacea: Brachyura). Biologia 69(7):905–915
Dereeper A, Guignon V, Blanc G, Audic S, Buffer S, Chevenet F, Dufayard JF, Guindon S, Lefort V, Lecot M, Claverie JM, Gas-cuel O (2008) Phylogeny.fr: robust phylogenetic analysis for the non-specialist. Nucleic Acids Res 36:W465–W469
Drach P, Tchernigovtzeff M (1967) Sur la méthode de détermination des stades d’intermue et son application générale aux crustacés. Vie Millieu 18:595–607
Fales RR (1976) Apparent predation on the mole crab Emerita tal-poida (Say) by the ghost crab Ocypode quadrata Fabricius. Che-sap Sci 17:65
Glowick RM, Golowash R, Keller R, Marder E (1997) D-glucose-sensitive neurosecretory cells of the crab Cancer borealis and negative feedback regulation of blood glucose level. J Exp Biol 200:1421–1431
Haley SR (1972) Reproductive cycling in the ghost crab, Ocy-pode quadrata (Fabr.) (Brachyura, Ocypodidae). Crustaceana 23(1):1–11
Henry RP, Booth CE, Lalier F, Walsh P (1994) Post-exercise lactate production and metabolism in three species of aquatic and ter-restrial decapod crustaceans. J Exp Mar Biol Ecol 186:215–234
Hervant F, Renault D (2002) Long-term fasting and realimentation in hypogean and epigean isopods: a proposed adaptive strategy for groundwater organisms. J Exp Biol 205:2079–2087
Hervant F, Mathieu J, Barré H (1999) Comparative study on the meta-bolic responses of subterranean and surface-dwelling amphipods to long-term starvation and subsequent refeeding. J Exp Biol 202:3587–3595
Jia C, Yu Q, Wang J, Li L (2014) Qualitative and quantitative top-down mass spectral analysis of crustacean hyperglycemic hor-mones in response to feeding. Proteomics 14(10):1185–1194
Katayama H, Ohira T, Nagasawa H (2013) Crustacean peptide hor-mones: structure, gene expression and function. Aqua Biosci Monogr 6(2):49–90
Keller R, Orth HP (1990) Hyperglycemic neuropeptides in crus-tacean. In: Epple A, Scanes CG, Stetson MH (eds) Progress in comparative endocrinology. Wiley-Liss, New York, pp 265–271
Leone FA, Garçon DP, Lucena MN, Faleiros RO, Azevedo SV, Pinto MR, McNamara JC (2015) Gill-specific (Na+, K+)-ATPase and α-subunit mRNA expression during low-salinity acclimation of the ornate blue crab Callinectes ornatus (Decapoda, Brachyura). Comp Biochem Physiol B 186:59–67
Maciel JE, Souza F, Valle S, Kucharski LC, Da Silva RSM (2008) Lactate metabolism in the muscle of the crab Chasmagnathus granulatus during hypoxia and post-hypoxia recovery. Comp Biochem Physiol A 151:61–65
Malev O, Šrut M, Maguire I, Štambuk A, Ferrero EA, Lorenzon S, Klobucar G (2010) Genotoxic, physiological and immunologi-cal effects caused by temperature increase, air exposure or food deprivation in freshwater crayfish Astacus leptodactylus. Comp Biochem Physiol C 152:433–443
McCue MD (2010) Starvation physiology: reviewing the different strategies animals use to survive a common challenge. Comp Biochem Physiol A 156:1–18
Melo GAS (1999) Infraordem Brachyura (Siris e caranguejos: espé-cies marinhas e de água doce). In: Buckup L, Bond-Buckup G (eds) Os Crustáceos do Rio Grande do Sul. Editora da Universi-dade UFRGS, Porto Alegre, pp 415–485
Milne LJ, Milne MJ (1946) Notes on the behavior of the ghost crab. Am Nat 80:362–380
Morris S, Postel U, Mrinalini LM, Turner L, Palmer J, Webster SG (2010) The adaptive significance of crustacean hyperglycemic hormone (CHH) in daily and seasonal migratory activities of the Christmas Island red crab Gecarcoidea natalis. J Exp Biol 213:3062–3073
Nagai C, Nagata S, Nagasawa H (2011) Effects of crustacean hyper-glycemic hormone (CHH) on the transcript expression of carbo-hydrate metabolism-related enzyme genes in the Kuruma prawn Marsupenaeus japonicus. Gen Comp Endocrinol 172:293–304
O’Connor JD, Gilbert LI (1968) Aspects of lipid metabolism in crus-taceans. Am Zool 8:529–539
Olivaux C, Soyez D (2000) Dynamics of biosynthesis and release of crustacean hyperglycemic hormone in the X-organ—sinus gland complex of the crayfish Orconectes limosus. Eur J Biochem 267:5106–5114
Oliveira GT, Rossi IC, Kucharski LC, Da Silva RSM (2004) Hepato-pancreas gluconeogenesis and glycogen content in crabs previ-ously maintained on a high-protein or carbohydrate-rich diet. Comp Biochem Physiol A 137:383–390
Pascual C, Sánchez A, Zenteno E, Cuzon G, Gaxiola G, Brito R, Gelabert R, Hidalgo E, Rosas C (2006) Biochemical, physiologi-cal, and immunological changes during starvation in juveniles of Litopenaeus vannamei. Aquaculture 251:416–429
Pellegrino R, Kucharski LC, Da Silva RSM (2008) Effect of starvation and refeeding on gluconeogenesis and glyconeogenesis in the mus-cle of the crab Chasmagnathus granulatus previously fed a protein- or carbohydrate-rich diet. J Exp Mar Biol Ecol 358:144–150
Pellegrino R, Martins TL, Pinto CB, Schein V, Kucharski LC, Da Silva RSM (2013) Effect of starvation and refeeding on amino acid metabolism in the muscle of the crab Neohelice granulata previously fed protein- or carbohydrate-rich diets. Comp Bio-chem Physiol A 164(1):29–35
Pellon-Maison M, Garcia CF, Cattaneo ER, Coleman RA, Gonzales-Baro MR (2009) Macobrachium borelli hepatopancreas contains a mitochondrial glycerol-3-phosphate acyltransferase which ini-tiates triacylglycerol biosynthesis. Lipids 44:337–344
Santos EA, Keller R (1993) Effect of exposure to atmospheric air and blood glucose and lactate concentrations in two crustacean spe-cies: a role of the crustacean hyperglycemic hormone (CHH). Comp Biochem Physiol A 106:343–347
Schlachter TA, Strydon S, Connolly RM, Schoeman D (2013) Donor-control of scavenging food webs at the land–ocean interface. PLoS One 8(6):e68221
Shi Q, Chung JS (2014) Trehalose metabolism in the blue crab Cal-linectes sapidus: isolation of multiple structural cDNA isoforms of trehalose-6-phosphate synthase and their expression in mus-cles. Gene 536:105–113
Stentiford GD, Chang ES, Chang SA, Neil DM (2001) Carbohydrate dynamics and the crustacean hyperglycemic hormone (CHH): effects of parasitic infection in Norway lobsters (Nephrops nor-vergicus). Gen Comp Endocrinol 121:13–22
Tullis A, Andrus SC (2011) The cost of incline locomotion in ghost crabs (Ocypode quadrata) of different sizes. J Comp Physiol B 181:873–881
Vinagre AS, Da Silva RSM (1992) Effects of starvation on the carbo-hydrate and lipid metabolism in crabs previously maintained on a high-protein or carbohydrate-rich diet. Comp Biochem Physiol A 102:579–583
Vinagre AS, Da Silva RSM (2002) Effects of starvation and refeeding on metabolic processes in the crab Chasmagnathus granulatus (Dana, 1851). Can J Zool 80:1413–1421
Vinagre AS, Amaral APN, Ribarcki FP, Silveira EF, Périco E (2007) Seasonal variation of energy metabolism in ghost crab Ocypode

Mar Biol (2016) 163:3
1 3
Page 11 of 11 3
quadrata at Siriú Beach (Brazil). Comp Biochem Physiol A 146:514–519
Webster SG, Keller R, Dircksen H (2012) The CHH-superfamily of multifunctional peptide hormones controlling crustacean metab-olism, osmoregulation, moulting and reproduction. Gen Comp Endocrinol 175:217–233
Weinstein RB, Full RJ (1998) Performance limits of low-temperature, continuous locomotion are exceeded when locomotion is inter-mittent in the ghost crab. Physiol Zool 71(3):274–284
Wolcott TG (1978) Ecological role of ghost crab Ocypode quadrata (Fabricius) on an ocean beach: scavengers or predators? J Exp Mar Biol 31:67–82
Wonglapsuwan M, Phongdara A, Chotigeat W (2009) Dynamic changes in gene expression during vitellogenic stages of the white shrimp: Fenneropenaeus merguiensis de Man. Aquac Res 40:633–643
Zhang Z, Wang Y, Jiang Y, Lin P, Jia X, Zou Z (2007) Ribossomal protein L24 is differentially expressed in ovary and testis of the marine shrimp Marsupenaeus japonicus. Comp Biochem Phys-iol B 147:466–474
Zhu X, Yuan M, Shakeel M, Zhang Y, Wang S, Zhan S, Kang T, Li J (2014) Selection and evaluation of reference genes for expres-sion analysis using qRT-PCR in the beet armyworm Spodop-tera exigua (Hubner) (Lepidoptera: Noctuidae). PLoS One 9(1):e84730
Zmora N, Trant J, Chen SM, Chung JS (2007) Vitellogenin and its Messenger RNA during ovarian development in female blue crab, Callinectes sapidus: gene expression, synthesis, transport, and cleavage. Biol Reprod 77:138–146





























