Embed Size (px)
Citation preview

Effects of Pituitary AdenylateCyclase-Activating Polypeptide,Vasoactive Intestinal Polypeptide, andSomatostatin on the Release ofThyrotropin from the BullfrogPituitary
REIKO OKADA,a KAZUTOSHI YAMAMOTO,a YOICHI ITO,a
NICOLAS CHARTREL,b JEROME LEPRINCE,b ALAIN FOURNIER,c
HUBERT VAUDRY,b AND SAKAE KIKUYAMAa
aDepartment of Biology, School of Education, Waseda University,Tokyo 169-8050, JapanbINSERM U413, Laboratory of Cellular and Molecular Neuroendocrinology,European Institute for Peptide Research (IFRMP 23), University of Rouen,76821 Mont-Saint-Aignan, FrancecINRS–Institut Armand Frappier, Montreal, Canada
ABSTRACT: The recent development of a specific radioimmunoassay foramphibian (bullfrog, Rana catesbeiana) thyrotropin (TSH) has made itpossible to study the effects of various neuropeptides on the release ofTSH from the pituitary in vitro. Up to now, corticotropin-releasing factorof bullfrog origin has been shown to have a potent TSH-releasing activity,whereas gonadotropin-releasing hormone and TSH-releasing hormoneexhibit a moderate TSH-releasing effect on the adult, but not larval,pituitary. In the present study, the effects of pituitary adenylate cyclase-activating polypeptide (PACAP), vasoactive intestinal polypeptide (VIP),and somatostatin (SS) on the in vitro release of TSH from the bull-frog pituitary were investigated. Both frog (R. ridibunda) PACAP-38 andPACAP-27 caused a concentration-dependent stimulation of the releaseof TSH from dispersed pituitary cells during a 24-h culture. The PACAP-38- and PACAP-27-induced TSH release was suppressed by a simulta-neous application of PACAP6–38. Application of high concentrations ofPACAP6–38 alone caused a slight but significant stimulatory effect onthe release of TSH. Frog VIP also stimulated TSH release from pituitary
Address for correspondence: Reiko Okada, Department of Biology, School of Education, WasedaUniversity, Nishiwaseda 1-6-1, Shinjuku-ku, Tokyo 169-8050, Japan. Voice: +81-3-5286-1517;fax: +81-3-3207-9694.
e-mail: [email protected]
Ann. N.Y. Acad. Sci. 1070: 474–480 (2006). C© 2006 New York Academy of Sciences.doi: 10.1196/annals.1317.064
474

OKADA et al.: RELEASE OF THYROTROPIN FROM THE BULLFROG PITUITARY 475
cells concentration-dependently. Frog SS1 (homologous to mammaliansomatostatin-14) and SS2 (homologous to mammalian cortistatin) didnot affect the basal release of TSH but caused a concentration-dependentsuppression of the PACAP-38-induced release of TSH. These results sug-gest the involvement of multiple neuropeptides in the regulation of therelease of TSH from the amphibian pituitary.
KEYWORDS: TSH; PACAP; VIP; somatostatin; amphibia
INTRODUCTION
Until recently, a radioimmunoassay (RIA) for amphibian thyrotropin (TSH)was not available because highly purified TSH preparations, necessary forantibody production, had never been obtained. We have recently developedan RIA for frog TSH using an antiserum against the C-terminal peptide ofthe bullfrog (Rana catesbeiana) TSH-� subunit deduced from the nucleotidesequence of the TSH-� complementary DNA.1,2 Using this RIA, we haveshown that corticotropin-releasing factor (CRF) stimulates the release of TSHfrom bullfrog pituitary cells more potently than thyrotropin-releasing hormone(TRH) and gonadotropin-releasing hormone (GnRH), and we have found thatendogenous CRF accounts for approximately 50% of the total TSH-releasingactivity in the frog hypothalamus.3
Pituitary adenylate cyclase-activating polypeptide (PACAP) and vasoactiveintestinal polypeptide (VIP) belong to the secretin/glucagon/growth hormone(GH)-releasing hormone superfamily.4 In amphibians as in mammals, PACAPexists in two molecular forms with 38 (PACAP-38) and 27 (PACAP-27) aminoacids.5–8 Three types of receptors for PACAP and VIP have been character-ized so far: the PACAP-specific receptor, PAC1-R, that binds PACAP-38 andPACAP-27 with high affinity, and the PACAP/VIP mutual receptors, VPAC1-Rand VPAC2-R, that bind PACAP-38, PACAP-27, and VIP with similar affin-ity.9 PACAP and VIP have been shown to stimulate the release of variouspituitary hormones in vertebrates.10
Somatostatin was first identified from the ovine hypothalamus on the basis ofits GH release-inhibiting activity.11 Two molecular forms of somatostatin (SS)have been characterized in the frog brain, that is, a variant called SS1 that pos-sesses the same sequence as mammalian somatostatin-14, and a variant calledSS2 ([Pro2,Met13]somatostatin-14) that is orthologous to cortistatin.12–14 Ev-idence has recently shown that somatostatin inhibits the release of TSH fromrat15 and chicken16 pituitaries.
In order to elucidate the possible involvement of hypothalamic peptides otherthan CRF, TRH, and GnRH in the release of TSH in amphibians, the effects ofPACAP, VIP, and somatostatins on the release of TSH from bullfrog pituitarycells were investigated.

476 ANNALS NEW YORK ACADEMY OF SCIENCES
MATERIALS AND METHODS
Culture of dispersed distal lobe pituitary cells of adult bullfrogs was per-formed as described elsewhere.1 Pituitary cells were suspended in 70% medium199 (M199) containing 0.1% bovine serum albumin. An aliquot of the cellsuspension was adjusted so that 1 mL contained 350,000 cells. The pituitarycells were incubated at 23◦C in a humidified atmosphere of 95% air–5% CO2
for 24 h. After preincubation, the culture medium was replaced with freshmedium containing frog (R. ridibunda) PACAP-38 (f PACAP-38),7 PACAP-27, PACAP6–38, frog VIP (f VIP),17 and/or frog SS (SS1 or SS2).11,12 Incubationwas continued for 24 h. After incubation, the medium was collected from eachwell and centrifuged, and the supernatant was subjected to RIA for bullfrogTSH.1 All experiments were approved by the Steering Committee for AnimalExperimentation at Waseda University.
RESULTS
The release of TSH from dispersed pituitary cells was stimulated by bothf PACAP-38 and PACAP-27 in a concentration (1–1000 nM)-dependent man-ner. During a 24-h incubation, a significant enhancement of the release ofTSH (226–358% of the control value) was observed following applicationof 10, 100, and 1000 nM f PACAP-38. Likewise, 100 and 1000 nM PACAP-27 stimulated the release of TSH (420–568% of the control value) signifi-cantly. f VIP also exhibited a potent TSH-releasing activity: at concentrations of1–100 nM, f VIP enhanced the release of TSH by 340–712% of the controlvalue; at the concentration of 100 nM, the response of pituitary cells to f VIPwas maximum (TABLE 1). On the other hand, the release of TSH enhanced
TABLE 1. Effects of frog PACAP-38 (f PACAP-38), PACAP-27, and frog VIP (f VIP) onthe release of TSH from dispersed distal lobe pituitary cells of the bullfrog
TSH released (% of control)
Concentration (nM)
Treatment 0 1 10 100 1000
f PACAP-38 100 ± 8.6a 138 ± 14.9a,b 226 ± 23.8b,c 236 ± 19.2c,d 358 ± 34.3d
PACAP-27 100 ± 7.7a 136 ± 12.1a 178 ± 28.1a 420 ± 35.7b 568 ± 33.5b
f VIP 100 ± 7.0a 340 ± 46.5b 503 ± 26.5c 712 ± 34.5d 705 ± 39.5d
The amounts of TSH secreted into the medium by dispersed pituitary cells during a 24-h incubationare expressed as percentages of the mean control value. The values are expressed as means ± SEM ofseven determinations. The mean values for TSH released in the absence of f PACAP-38, PACAP-27,and f VIP were 413 ± 36, 341 ± 26, and 356 ± 25 pg/10,000 cells, respectively. Within each treatment,values without a common letter in the superscripts are significantly different at the 5% level (ANOVAand Scheffe’s test).
OKADA et al.: RELEASE OF THYROTROPIN FROM THE BULLFROG PITUITARY 477
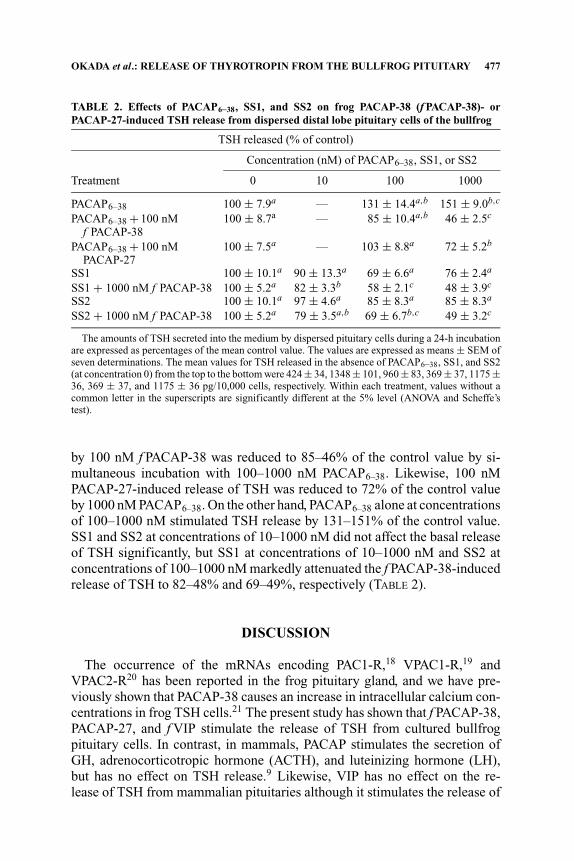
TABLE 2. Effects of PACAP6–38, SS1, and SS2 on frog PACAP-38 (f PACAP-38)- orPACAP-27-induced TSH release from dispersed distal lobe pituitary cells of the bullfrog
TSH released (% of control)
Concentration (nM) of PACAP6–38, SS1, or SS2
Treatment 0 10 100 1000
PACAP6–38 100 ± 7.9a — 131 ± 14.4a,b 151 ± 9.0b,c
PACAP6–38 + 100 nM 100 ± 8.7a — 85 ± 10.4a,b 46 ± 2.5c
f PACAP-38PACAP6–38 + 100 nM 100 ± 7.5a — 103 ± 8.8a 72 ± 5.2b
PACAP-27SS1 100 ± 10.1a 90 ± 13.3a 69 ± 6.6a 76 ± 2.4a
SS1 + 1000 nM f PACAP-38 100 ± 5.2a 82 ± 3.3b 58 ± 2.1c 48 ± 3.9c
SS2 100 ± 10.1a 97 ± 4.6a 85 ± 8.3a 85 ± 8.3a
SS2 + 1000 nM f PACAP-38 100 ± 5.2a 79 ± 3.5a,b 69 ± 6.7b,c 49 ± 3.2c
The amounts of TSH secreted into the medium by dispersed pituitary cells during a 24-h incubationare expressed as percentages of the mean control value. The values are expressed as means ± SEM ofseven determinations. The mean values for TSH released in the absence of PACAP6–38, SS1, and SS2(at concentration 0) from the top to the bottom were 424 ± 34, 1348 ± 101, 960 ± 83, 369 ± 37, 1175 ±36, 369 ± 37, and 1175 ± 36 pg/10,000 cells, respectively. Within each treatment, values without acommon letter in the superscripts are significantly different at the 5% level (ANOVA and Scheffe’stest).
by 100 nM f PACAP-38 was reduced to 85–46% of the control value by si-multaneous incubation with 100–1000 nM PACAP6–38. Likewise, 100 nMPACAP-27-induced release of TSH was reduced to 72% of the control valueby 1000 nM PACAP6–38. On the other hand, PACAP6–38 alone at concentrationsof 100–1000 nM stimulated TSH release by 131–151% of the control value.SS1 and SS2 at concentrations of 10–1000 nM did not affect the basal releaseof TSH significantly, but SS1 at concentrations of 10–1000 nM and SS2 atconcentrations of 100–1000 nM markedly attenuated the f PACAP-38-inducedrelease of TSH to 82–48% and 69–49%, respectively (TABLE 2).
DISCUSSION
The occurrence of the mRNAs encoding PAC1-R,18 VPAC1-R,19 andVPAC2-R20 has been reported in the frog pituitary gland, and we have pre-viously shown that PACAP-38 causes an increase in intracellular calcium con-centrations in frog TSH cells.21 The present study has shown that f PACAP-38,PACAP-27, and f VIP stimulate the release of TSH from cultured bullfrogpituitary cells. In contrast, in mammals, PACAP stimulates the secretion ofGH, adrenocorticotropic hormone (ACTH), and luteinizing hormone (LH),but has no effect on TSH release.9 Likewise, VIP has no effect on the re-lease of TSH from mammalian pituitaries although it stimulates the release of

478 ANNALS NEW YORK ACADEMY OF SCIENCES
prolactin (PRL), LH, and GH.22 The fact that f PACAP and f VIP could both en-hance TSH secretion in frog strongly suggests the involvement of the VPAC1-Rand/or the VPAC2-R in the response of thyrotrope cells to the peptide.
We have previously shown that the two frog somatostatin isoforms, SS1and SS2, inhibit growth hormone-releasing hormone (GHRH)-induced GHsecretion from bullfrog pituitary cells.23 Here, we show that both SS1 and SS2inhibit f PACAP-38-evoked TSH release. It thus appears that the two somato-statin isoforms may control the activity of both somatotrope and thyrotropecells in amphibians. Consistent with this notion, it has been recently reportedthat the gene encoding the SS2 precursor is actively expressed in the pars in-termedia of the frog pituitary,24 suggesting that SS2, released by melanotropecells, can diffuse toward the distal lobe to regulate the secretion of variousadenohypophysial hormones.
In conclusion, the release of TSH in the amphibian pituitary is controlled bymultiple hypothalamic neurohormones. The marked differences, between am-phibians and mammals, in the hypothalamic neuropeptides regulating hormonesecretion from each adenohypophysial cell type demonstrate the existence ofphylogenetic variations in the expression pattern of the receptors for the hy-pothalamic factors in the different pituitary cells.
ACKNOWLEDGMENTS
This work was supported by Inamori Foundation (H17) JSPS (16370031,15207007), Waseda University (2004B-839, 2003C-009), INSERM (U413),the European Institute for Peptide Research (IFRMP 23), and the ConseilRegional de Haute-Normandie.
REFERENCES
1. OKADA, R., K. YAMAMOTO, A. KODA, et al. 2004. Development of radioimmunoas-say for bullfrog thyroid-stimulating hormone (TSH): effects of hypothalamic re-leasing hormones on the release of TSH from the pituitary in vitro. Gen. Comp.Endocrinol. 135: 42–50.
2. OKADA, R., Y. ITO, M. KANEKO, et al. 2005. Frog corticotropin-releasing hormone(CRH): isolation, molecular cloning, and biological activity. Ann. N. Y. Acad.Sci. 1040: 150–155.
3. ITO, Y., R. OKADA, H. MOCHIDA, et al. 2004. Molecular cloning of bullfrogcorticotropin-releasing factor (CRF): effect of homologous CRF on the releaseof TSH from pituitary cells in vitro. Gen. Comp. Endocrinol. 138: 218–227.
4. ARIMURA, A. 1998. Perspectives on pituitary adenylate cyclase-activating polypep-tide (PACAP) in the neuroendocrine, endocrine, and nervous systems. Jpn. J.Physiol. 48: 301–331.
5. MIYATA, A., A. ARIMURA, R.R. DAHL, et al. 1989. Isolation of a novel 38 residue-hypothalamic polypeptide which stimulates adenylate cyclase in pituitary cells.Biochem. Biophys. Res. Commun. 164: 567–574.

OKADA et al.: RELEASE OF THYROTROPIN FROM THE BULLFROG PITUITARY 479
6. MIYATA, A., L. JIANG, R.D. DAHL, et al. 1990. Isolation of a neuropeptide cor-responding to the N-terminal 27 residues of the pituitary adenylate cyclase-activating polypeptide with 38 residues (PACAP38). Biochem. Biophys. Res.Commun. 170: 643–648.
7. CHARTREL, N., M.C. TONON, H. VAUDRY & J.M. CONLON. 1991. Primarystructure of frog pituitary adenylate cyclase-activating polypeptide (PACAP)and effects of ovine PACAP on frog pituitary. Endocrinology 129: 3367–3371.
8. YON, L., M. FEUILLOLEY, N. CHARTREL, et al. 1992. Immunohistochemical distribu-tion and biological activity of pituitary adenylate cyclase-activating polypeptide(PACAP) in the central nervous system of the frog Rana ridibunda. J. Comp.Neurol. 324: 485–499.
9. VAUDRY, D., B.J. GONZALEZ, M. BASILLE, et al. 2000. Pituitary adenylate cyclase-activating polypeptide and its receptors: from structure to function. Pharmacol.Rev. 52: 269–324.
10. RAWLINGS, S.R. & M. HEZAREH. 1996. Pituitary adenylate cyclase-activatingpolypeptide (PACAP) and PACAP/VIP receptors: actions on the anteriorpituitary gland. Endocr. Rev. 17: 4–29.
11. BRAZEAU, P., W. VALE, R. BURGUS, et al. 1973. Hypothalamic polypeptide thatinhibits the secretion of immunoreactive pituitary growth hormone. Science 179:77–79.
12. VAUDRY, H., N. CHARTREL & J.M. CONLON. 1992. Isolation of[Pro2,Met13]somatostatin-14 and somatostatin-14 from the frog brain reveals theexistence of a somatostatin gene family in a tetrapod. Biochem. Biophys. Res.Commun. 188: 477–482.
13. TOSTIVINT, H., I. LIHRMANN, C. BUCHARLES, et al. 1996. Occurrence of two so-matostatin variants in the frog brain: characterization of the cDNAs, distributionof the mRNAs and receptor-binding affinities of the peptides. Proc. Natl. Acad.Sci. USA 93: 12605–12610.
14. DE LECEA, L., J.R. CRIADO, O. PROSPERO-GARCIA, et al. 1996. A cortical neu-ropeptide with neuronal depressant and sleep-modulating properties. Nature 381:242–245.
15. REICHLIN, S. 2001. Neuroendocrine control of thyrotropin secretion. In The ThyroidL. Braverman, & R.D. Utiger, Eds.: 241–266. Lippincott Williams & Wilkins.Philadelphia.
16. GERIS, K.L., B. DE GROEF, E.R. KUHN & V.M. DARRAS. 2003. In vitro studyof corticotropin-releasing hormone-induced thyrotropin release: ontogeny andinhibition by somatostatin. Gen. Comp. Endocrinol. 132: 272–277.
17. CHARTREL, N., Y. WANG, A. FOURNIER, et al. 1995. Frog vasoactive intestinalpolypeptide and galanin: primary structures and effects on pituitary adenylatecyclase. Endocrinology 136: 3079–3086.
18. ALEXANDRE, D., H. VAUDRY, V. TURQUIER, et al. 2002. Novel splice variants of type Ipituitary adenylate cyclase-activating polypeptide receptor in frog exhibit alteredadenylate cyclase stimulation and differential relative abundance. Endocrinology143: 2680–2692.
19. ALEXANDRE, D., Y. ANOUAR, S. JEGOU, et al. 1999. A cloned frog vasoac-tive intestinal polypeptide/pituitary adenylate cyclase-activating polypeptidereceptor exhibits pharmacological and tissue distribution characteristics ofboth VPAC1 and VPAC2 receptors in mammals. Endocrinology 140: 1285–1293.

480 ANNALS NEW YORK ACADEMY OF SCIENCES
20. HOO, R.L.C., D. ALEXANDRE, S.M. CHAN, et al. 2001. Structural and functionalidentification of the pituitary adenylate cyclase-activating polypeptide receptorVPAC2 receptor from the frog Rana tigrina rugulosa. J. Mol. Endocrinol. 27:229–238.
21. GRACIA-NAVARRO, F., M. LAMACZ, M.C. TONON, et al. 1992. Pituitary adenylatecyclase-activating polypeptide stimulates calcium mobilization in amphibian pi-tuitary cells. Endocrinology 131: 1069–1074.
22. SHERWOOD, N.M., L. SANDRA, L. KRUECKL, et al. 2000. The origin and functionof the pituitary adenylate cyclase-activating polypeptide (PACAP)/glucagon su-perfamily. Endocr. Rev. 21: 619–670.
23. JEANDEL, L., A. OKUNO, T. KOBAYASHI, et al. 1998. Effects of the two somatostatinvariants somatostatin-14 and [Pro2,Met13]somatostatin-14 on receptor binding,adenylyl cyclase activity and growth hormone release from the frog pituitary. J.Neuroendocrinol. 10: 187–192.
24. TOSTIVINT, H., D. VIEAU, N. CHARTREL, et al. 2002. Expression and processing ofthe [Pro2,Met13]somatostatin-14 precursor in the intermediate lobe of the frogpituitary. Endocrinology 143: 3472–3481.








![Regulation of the Hypothalamic Thyrotropin Releasing Hormone … · biosynthesis and secretion of thyroid stimulating hormone (TSH) from the pituitary [7; 8], which in turn, stimulates](https://img.pdfslide.us/doc/110x75/6045baff65e91974e54d0c71/regulation-of-the-hypothalamic-thyrotropin-releasing-hormone-biosynthesis-and-secretion.jpg)




















