Embed Size (px)
Citation preview

lable at ScienceDirect
Food Microbiology 26 (2009) 497–503
Contents lists avai
Food Microbiology
journal homepage: www.elsevier .com/locate/ fm
Effects of organic acids on thermal inactivation of acid and cold stressedEnterococcus faecium
Ana Fernandez, Avelino Alvarez-Ordonez, Mercedes Lopez, Ana Bernardo*
Departamento de Higiene y Tecnologıa de los Alimentos, Facultad de Veterinaria, Universidad de Leon, Campus de Vegazana s/n, 24071 Leon, Spain
a r t i c l e i n f o
Article history:Received 19 December 2008Received in revised form5 March 2009Accepted 10 March 2009Available online 17 March 2009
Keywords:Enterococcus faeciumHeat inactivation kineticOrganic acidsAcid adaptationCold stress
* Corresponding author. Tel.: þ34 987 291182; fax:E-mail address: [email protected] (A. Bernardo).
0740-0020/$ – see front matter � 2009 Elsevier Ltd.doi:10.1016/j.fm.2009.03.002
a b s t r a c t
In this study the adaptative response to heat (70 �C) of Enterococcus faecium using fresh and refrigerated(at 4 �C for up to 1 month) stationary phase cells grown in Brain Heart Infusion (BHI) buffered at pH 7.4(non-acid-adapted cells) and acidified BHI at pH values of 6.4 and 5.4 with acetic, ascorbic, citric, lactic,malic and hydrochloric acids (acid-adapted cells) was evaluated. In all cases, the survival curves obtainedwere concave upward. A mathematical model based on the Weibull distribution accurately described theinactivation kinetic. The results indicate that previous adaptation to a low pH increased the bacterial heatresistance, whereas the subsequent cold storage of cells reduced E. faecium thermal tolerance. Fresh acid-adapted cells showed t2.5-values (time needed to obtain an inactivation level of 2.5 log10 cycles) rangingfrom 2.57 to 9.51 min, while non-acid-adapted cells showed t2.5-values of 1.92 min. The extent ofincreased heat tolerance varied with the acid examined, resulting in the following order: cit-ric� acetic>malic� lactic> hydrochloric� ascorbic. In contrast, cold storage progressively decreased E.faecium thermal resistance. The t2.5 values found at the end of the period studied were about 2–3-foldlower than those corresponding to non-refrigerated cells, although this decrease was more marked(about 5-fold) when cells were grown in buffered BHI and BHI acidified at pH 5.4 with hydrochloric acid.These findings highlight the need for a better understanding of microbial response to various preser-vation stresses in order to increase the efficiency of thermal processes and to indicate the convenience ofcounterbalancing the benefits of the hurdle concept.
� 2009 Elsevier Ltd. All rights reserved.
1. Introduction
Enterococcus faecium has long been recognised as havingimportant technological properties, such as the improvement offlavour of several fermented foods (Cintas et al., 1997; Centenoet al., 1999; Franz et al., 2003), although occasionally it has alsobeen associated with the spoilage of thermal processed foods(Houben, 1982; Magnus et al., 1986; Ghazala et al., 1995). Thismicroorganism is one of the most heat-resistant non-sporeformingbacterium and capable of growing at a wide range of temperatures(Murray, 1990; Devriese et al., 1993; Martınez et al., 2003) in thepresence of salt and at low pH values (Sanz Perez et al., 1982; Fla-haut et al., 1997; Giraffa et al., 1997). Its relative resistance toadverse conditions together with its ubiquitous nature makes E.faecium appropriate to evaluate the efficacy of pasteurizationprocesses (Smith et al., 1990; Ghazala et al., 1991) and disinfectionmethods using wet heat or water vapour (Spicher et al., 2002). Alarge variety of studies have been performed in order to know the
þ34 987 291284.
All rights reserved.
effects of environmental factors on heat inactivation of E. faecium.From some of these studies it can be concluded that the conditionswhich prevail before thermal treatment are specially relevant, suchas the physiological state of cells (Houben, 1982; Kearns et al., 1995;Martınez et al., 2003), the growth temperature (Ahmad et al., 2003;Martınez et al., 2003) and the culture medium composition (Ahmadet al., 2002). However, and despite the fact that the culture mediumpH and the acidulant identity are generally considered to stronglyaffect microbial heat resistance (Juneja et al., 1998; Duffy et al.,2000; Mazzotta, 2001; Tosun and Gonul, 2003; Alvarez-Ordonezet al., 2008), there does not appear to be literature available con-cerning their effects on the E. faecium heat resistance. Additionally,the microbial growth in acidic conditions may induce a cross-protection response which frequently results in an enhancedthermotolerance. While such stress adaptation has beendemonstrated in various foodborne pathogens, such as Listeriamonocytogenes (Farber and Pagotto, 1992; Lou and Yousef, 1996;Juneja et al., 1998), Escherichia coli (Buchanan and Edelson, 1999;Duffy et al., 2000; Evrendilek and Zhang, 2003), Salmonella typhi-murium (Bacon et al., 2003; Tosun and Gonul, 2003; Alvarez-Ordonez et al., 2008) and Salmonella enteritidis (Wilde et al., 2000;

A. Fernandez et al. / Food Microbiology 26 (2009) 497–503498
Mazzotta, 2001), the study of such phenomenon in E. faecium hasyet to be investigated. Furthermore, and taking into account the factthat the acidification is widely used together with cold storage insome processed foods to control the microbial growth, littleattention has been paid to the thermal inactivation of acid-adaptedmicroorganisms exposed to low temperatures. Recent studieswithin our research group (Alvarez-Ordonez et al., 2009) havedemonstrated that the combination of these sublethal stressesreduces Salmonella senftenberg heat tolerance.
Therefore, this investigation was performed to determine thethermal inactivation kinetic of E. faecium after its growth in acidi-fied media up to pH 5.4 using different organic acids (acetic,ascorbic, citric, lactic and malic) and hydrochloric acid. Moreover,the effect of cold storage on the subsequent heat resistance of acid-adapted cells was also researched into.
2. Material and methods
2.1. Bacterial strain, media and culture conditions
The E. faecium strain (ATCC 49624) used in this study wasobtained from American Type Culture Collection (ATCC). Thelyophilized cultures were revived in Brain Heart Infusion (BHI,Oxoid) and incubated for 24 h at 37 �C. Pure cultures were main-tained on Agar BHI (BHIA, Oxoid) plates at 4 �C. Subcultures wereprepared by transferring an isolated colony from a plate into a testtube containing 10 mL of sterile BHI followed by incubation at 37 �Cfor 24 h. These fresh subcultures were used to produce acid-adapted and non-acid-adapted cells.
2.2. Preparation of acid and cold-adapted cells
Flasks containing 50 mL of sterile BHI (pH 7.4) and BHI acidifiedat pH values of 6.4 and 5.4 with several acids (acetic (Prolab),ascorbic (Merck), citric (Sigma), lactic (Merck), malic (Scharlau) andhydrochloric (Panreac)) were inoculated with the subculture toa final concentration of 103 cells/mL. Non-acid-adapted cells wereobtained by inoculating the same concentration of microorganismsinto buffered BHI adjusted to pH 7.4 by the addition of Sorensenbuffer 0.2 M (bisodium-monopotassium phosphate (Panreac)). Thecultures were then incubated at 37 �C for 36 h, resulting in all casescultures that were in a late stationary phase of growth. Threedifferent fresh grown cultures of E. faecium were used for eachexperimental run. Initial and final pH values of each culturecondition are shown in Table 2.
Cold adaptation consisted of maintaining cultures at 4 �C for 8 h,48 h, one week and one month. Preliminary studies confirmed thatin all cases cell viability remained constant throughout the coldstorage period.
2.3. Heat treatments
Heat treatments were carried out in a thermoresistometer TR-SC as described by Condon et al. (1993). Once the temperature ofthe heat treatment medium (350 mL of BHI) was stabilized(70� 0.05 �C), an inoculum of each culture was injected. Duringheating, samples of 0.3 mL were removed at predetermined timeintervals and were plated in BHIA. Plates were then incubated at37 �C for 72 h and survivors were counted with a modified ImageAnalyser Automatic Counter (Protos Analytical Measuring Systems,Cambridge, UK) as described elsewhere (Ibarz et al., 1991). Heattreatments were performed in triplicate on three different bacterialcultures.
2.4. Curve fitting
In order to fit the survival curves of E. faecium the followingmathematical function based on the Weibull distribution was used:
log Nt=N0 ¼ �ðt=dÞp (1)
where t is the treatment time, Nt and N0 are the populationdensities (cfu/mL) at time t and time zero, respectively, and d and pare the scale and shape parameters, respectively.
The d value represents the time needed to inactivate the firstlog10 cycle of the population. The p parameter accounts for upwardconcavity of a survival curve (p< 1), a linear survival curve (p¼ 1),and downward concavity (p> 1) (Mafart et al., 2002).
In order to determine the d and p values, the least-squarescriterion by the GraphPad PRISM (GraphPad Software, San Diego,CA) was used. Student’s t-test (Steel and Torrie, 1986) was used todetermine significant differences (p< 0.05) between d and p values(GraphPad Software, San Diego, CA).
3. Results
E. faecium thermal inactivation at 70 �C of acid and cold-adaptedcells was studied using stationary phase cells. Viable counts of E.faecium were monitored during its growth in buffered BHI at pH 7.4(non-acid-adapted cells), non-acidified BHI (non-acidified cells)and BHI acidified at pH values of 6.4 and 5.4 with several organicacids (acetic, ascorbic, citric, lactic and malic) and hydrochloric acid(acid-adapted cells). It is worth noting that this microorganismgrew well under all conditions assayed. Only significative differ-ences in the growth kinetic parameters were observed when aceticacid at pH 5.4 was used. This growth condition doubled thegeneration time (0.55 h) and the time needed to reach thestationary phase (16 h) and tripled the lag phase duration (2.75 h)and decreased about one log10 cycle the maximum populationdensity.
Following the growth in these media at 37 �C for 36 h, the heatresistance determinations at 70 �C in BHI were then performed forfresh cultures and after their later storage at 4 �C for 8 h, 48 h, oneweek and one month. In all cases, the populations of viable cellsremained static throughout the storage period, regardless of priorgrowth conditions (data not shown).
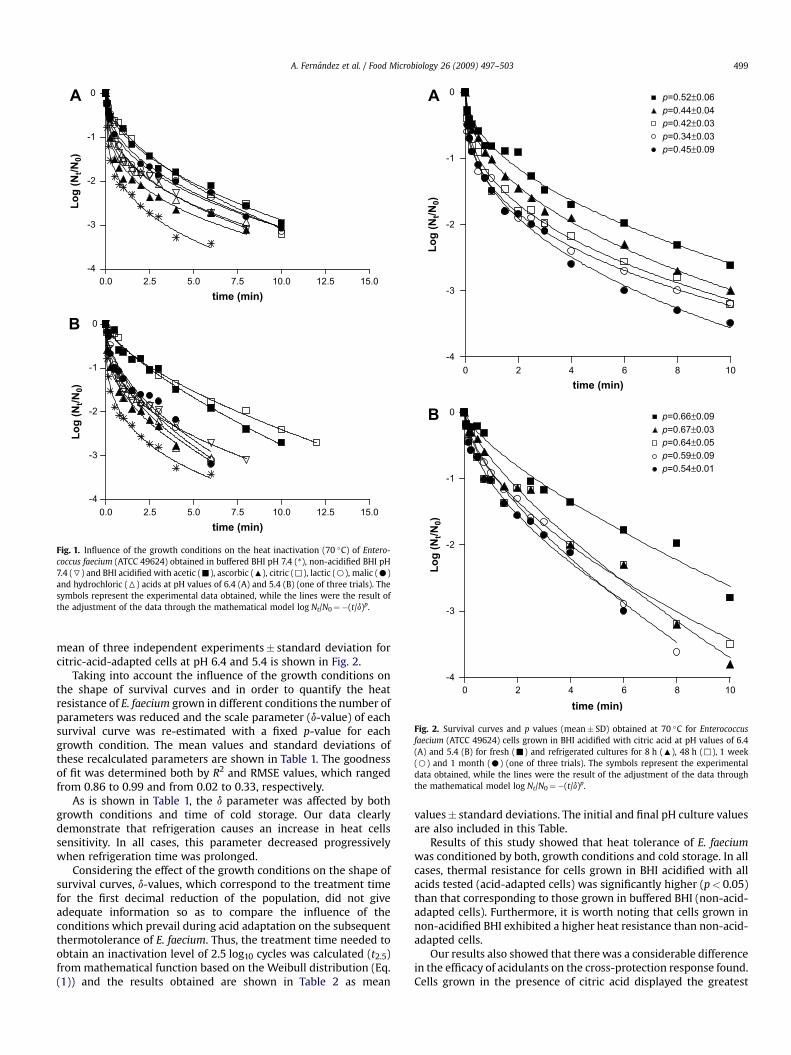
Survival curves of E. faecium obtained under the differentconditions assayed were concave upward, which indicates that itsinactivation was very fast during the first moments of the treat-ment and then the number of survivors slowly decreased as thetreatment time increased. In order to quantify and compare theheat inactivation, all survival curves obtained in this study werefitted to the mathematical model based on the Weibull distribution(Eq. (1)) and the corresponding shape parameter (p-value, whichaccounts for upward concavity of a survival curve (p< 1), a linearsurvival curve (p¼ 1), and downward concavity (p> 1)) and scaleparameter (d-value, which represents the time needed to inactivatethe first log10 cycle of the population) were estimated. An exampleof the survival curves obtained for fresh cultures under all condi-tions is shown in Fig. 1.
There was a considerable difference in the effects of the growthconditions tested on the shape parameter, obtaining the lowestp-values for non-acid-adapted (p¼ 0.29� 0.04) and non-acidifiedcells (p¼ 0.37� 0.02) and the highest for cells grown at pH 5.4,which ranged from 0.44� 0.07 (lactic-acid-adapted cells) to0.78� 0.10 (acetic-acid-adapted cells). Our results also showed thatthe shape parameter was not significantly modified by cold storage.An example of the survival curves and the p-values estimated as the
A
0.0 2.5 5.0 7.5 10.0 12.5 15.0-4
-3
-2
-1
0
time (min)
0.0 2.5 5.0 7.5 10.0 12.5 15.0time (min)
Lo
g (N
t/N
0)
-4
-3
-2
-1
0
Lo
g (N
t/N
0)
B
Fig. 1. Influence of the growth conditions on the heat inactivation (70 �C) of Entero-coccus faecium (ATCC 49624) obtained in buffered BHI pH 7.4 (*), non-acidified BHI pH7.4 (7) and BHI acidified with acetic (-), ascorbic (:), citric (,), lactic (B), malic (C)and hydrochloric (6) acids at pH values of 6.4 (A) and 5.4 (B) (one of three trials). Thesymbols represent the experimental data obtained, while the lines were the result ofthe adjustment of the data through the mathematical model log Nt/N0¼�(t/d)p.
p=0.52±0.06p=0.44±0.04
p=0.34±0.03p=0.45±0.09
p=0.42±0.03
0 2 4 6 8 10
p=0.66±0.09
p=0.59±0.09p=0.54±0.01
p=0.67±0.03p=0.64±0.05
time (min)
0 2 4 6 8 10time (min)
-4
-3
-2
-1
0
Lo
g (N
t/N
0)
-4
-3
-2
-1
0
Lo
g (N
t/N
0)
A
B
Fig. 2. Survival curves and p values (mean� SD) obtained at 70 �C for Enterococcusfaecium (ATCC 49624) cells grown in BHI acidified with citric acid at pH values of 6.4(A) and 5.4 (B) for fresh (-) and refrigerated cultures for 8 h (:), 48 h (,), 1 week(B) and 1 month (C) (one of three trials). The symbols represent the experimentaldata obtained, while the lines were the result of the adjustment of the data throughthe mathematical model log Nt/N0¼�(t/d)p.
A. Fernandez et al. / Food Microbiology 26 (2009) 497–503 499
mean of three independent experiments� standard deviation forcitric-acid-adapted cells at pH 6.4 and 5.4 is shown in Fig. 2.
Taking into account the influence of the growth conditions onthe shape of survival curves and in order to quantify the heatresistance of E. faecium grown in different conditions the number ofparameters was reduced and the scale parameter (d-value) of eachsurvival curve was re-estimated with a fixed p-value for eachgrowth condition. The mean values and standard deviations ofthese recalculated parameters are shown in Table 1. The goodnessof fit was determined both by R2 and RMSE values, which rangedfrom 0.86 to 0.99 and from 0.02 to 0.33, respectively.
As is shown in Table 1, the d parameter was affected by bothgrowth conditions and time of cold storage. Our data clearlydemonstrate that refrigeration causes an increase in heat cellssensitivity. In all cases, this parameter decreased progressivelywhen refrigeration time was prolonged.
Considering the effect of the growth conditions on the shape ofsurvival curves, d-values, which correspond to the treatment timefor the first decimal reduction of the population, did not giveadequate information so as to compare the influence of theconditions which prevail during acid adaptation on the subsequentthermotolerance of E. faecium. Thus, the treatment time needed toobtain an inactivation level of 2.5 log10 cycles was calculated (t2.5)from mathematical function based on the Weibull distribution (Eq.(1)) and the results obtained are shown in Table 2 as mean
values� standard deviations. The initial and final pH culture valuesare also included in this Table.
Results of this study showed that heat tolerance of E. faeciumwas conditioned by both, growth conditions and cold storage. In allcases, thermal resistance for cells grown in BHI acidified with allacids tested (acid-adapted cells) was significantly higher (p< 0.05)than that corresponding to those grown in buffered BHI (non-acid-adapted cells). Furthermore, it is worth noting that cells grown innon-acidified BHI exhibited a higher heat resistance than non-acid-adapted cells.
Our results also showed that there was a considerable differencein the efficacy of acidulants on the cross-protection response found.Cells grown in the presence of citric acid displayed the greatest

Table 1Influence of cold storage on d-values estimated from the second fit of the mathematical model log Nt/N0¼�(t/d)p to experimental data of non-acid adapted and non-acidifiedcells of Enterococcus faecium (ATCC 49624) heat treated at 70 �C considering a constant p-value for each adaptation condition.
Growthconditions
d-value (min) p-value
Time of cold storage
pH Media 0 h 8 h 48 h 1 week 1 month
7.4 NAA 0.085� 0.012a 0.046� 0.008b 0.039� 0.007b 0.025� 0.003c 0.018� 0.003d 0.29 � 0.04NA 0.55� 0.04a 0.39� 0.01b 0.34� 0.05bc 0.27� 0.08bc 0.24� 0.04c 0.42 � 0.08
6.4 Aace 0.87� 0.12a 0.47� 0.11b 0.41� 0.09bc 0.25� 0.05cd 0.24� 0.02d 0.38 � 0.06Aasc 0.16� 0.03a 0.13� 0.01a 0.12� 0.01ab 0.11� 0.02ab 0.09� 0.02b 0.31 � 0.03Acit 1.20� 0.19a 0.81� 0.08b 0.63� 0.08bc 0.53� 0.04c 0.47� 0.06c 0.43 � 0.08Alac 0.58� 0.01a 0.39� 0.03b 0.38� 0.04bc 0.37� 0.07bc 0.31� 0.03c 0.38 � 0.02Amal 0.64� 0.04a 0.46� 0.04b 0.44� 0.05b 0.42� 0.05b 0.39� 0.04b 0.40 � 0.06AHCl 0.34� 0.03a 0.20� 0.04b 0.19� 0.04b 0.17� 0.01b 0.17� 0.01b 0.36 � 0.06
5.4 Aace 2.17� 0.25a 1.38� 0.14b 1.22� 0.05b 1.13� 0.27b 0.98� 0.23b 0.70 � 0.09Aasc 0.25� 0.04a 0.19� 0.02ab 0.17� 0.01bc 0.15� 0.01c 0.09� 0.01d 0.42 � 0.04Acit 2.23� 0.25a 1.70� 0.12b 1.45� 0.50bc 1.10� 0.08c 1.08� 0.06c 0.62 � 0.09Alac 0.86� 0.08a 0.72� 0.03b 0.58� 0.04c 0.51� 0.05c 0.37� 0.03d 0.52� 0.09Amal 0.66� 0.06a 0.48� 0.05b 0.41� 0.04b 0.25� 0.03c 0.20� 0.02c 0.43 � 0.09AHCl 0.51� 0.03a 0.28� 0.04b 0.30� 0.04b 0.15� 0.01c 0.10� 0.02d 0.52 � 0.09
NAA: Non-acid-adapted cells, grown in buffered BHI pH 7.4.NA: Non-acidified cells, grown in BHI pH 7.4.AAce, AAsc, ACit, ALac, AMal, AHCl: Acid-adapted cells, grown in BHI acidified with: acetic, ascorbic, citric, lactic, malic and hydrochloric acids, respectively.a–dValues (mean of three experiments� standard deviation) with different superscript for each adaptation condition were statistically different (p< 0.05).
A. Fernandez et al. / Food Microbiology 26 (2009) 497–503500
thermotolerance. Non-refrigerated cells reached t2.5 values of about5-fold higher than those corresponding to non-acid-adapted cells(t2.5¼1.92� 0.15 min) whereas the lowest times of inactivationcorresponded to hydrochloric and ascorbic-acid-adapted cells,which only increased their themotolerance 1.3–2-fold.
Of particular interest in our data is the fact that in all cases theheat resistance decreased progressively during cold storage. Thus,the t2.5 values found at the end of the period studied were about2–3-fold lower than those corresponding to non-refrigerated cells,although this decrease was more marked (about 5-fold) when cellswere grown in buffered BHI (non-acid-adapted cells) and BHIacidified at pH 5.4 with hydrochloric acid.
In addition, it was also found that the acidification of growthmedium resulted in more heat sensitive cells, except for cellsadapted in presence of acetic and citric acids. This specific effect ofgrowth medium pH was more pronounced when cold storage wasprolonged. Thus, after one month of storage, the t2.5 values of cellsgrown at pH 6.4 with ascorbic (1.72� 0.44 min), lactic (3.56� 0.39min), malic (3.89� 0.60 min) and hydrochloric (2.49� 0.32 min)were in all cases significantly higher (p< 0.05) than those obtained
Table 2t2.5 values* (min) estimated from fitting of the mathematical model log Nt/N0¼�(t/d)p t
Growthmedium
InitialpH
Stationary-phase pH
Time of cold storage
0 h 8 h
NAA 7.4 7.4 1.92� 0.15a 1.13NA 6.2 5.18� 0.55b 3.46AAce 6.4 5.3 8.62� 1.15c 5.01AAsc 5.3 2.57� 0.31d 2.25ACit 5.5 9.51� 1.35c 6.37ALac 5.2 6.22� 0.41be 4.32AMal 5.4 6.40� 0.70be 4.34AHCl 5.3 3.93� 0.46f 2.78AAce 5.4 5.0 7.55� 1.30ce 5.04AAsc 4.9 2.56� 0.70ad 1.96ACit 5.1 9.16� 2.04ce 7.00ALac 4.9 5.30� 0.52b 4.41AMal 5.0 5.25� 0.53b 3.69AHCl 4.9 2.92� 0.36d 1.73
t2.5 values*: Time (min) needed at 70 �C to inactivate 2.5 log10 cycles of the population oNAA: Non-acid-adapted cells, grown in BHI buffered pH 7.4.NA: Non-acidified cells, grown in BHI pH 7.4.AAce, AAsc, ACit, ALac, AMal, AHCl: Acid-adapted cells, grown in BHI acidified at pH 6.4 and 5a–g Values (mean of three different experiments� standard deviation) with different sup
for cells grown at pH 5.4 (0.79� 0.09; 2.26� 0.48; 1.58� 0.18; and0.59� 0.09 min, respectively). To sum up, an overall view of our datashows that the order of acids in conferring E. faecium heat tolerancewas: citric� acetic>malic� lactic> hydrochloric� ascorbic.
4. Discussion
Organic acids are commonly used in the food industry tocontrol growth and survival of pathogenic and spoilage bacteria.However, microorganisms after their exposure under mild acidicconditions, which in several processed food conditions can beassociated with organic acids, could have enhanced their thermalresistance, and consequently, the heat treatment times needed toeliminate acid-adapted microorganisms from foods would beincreased, although on some occasions acid adaptation is indus-trially exploited to increase survival and activity of starter culturesduring and after preservation by drying and freezing (Parente andCogan, 2004).
In this study we have evaluated the heat tolerance of stationaryphase cells of E. faecium (ATCC 49624) with and without prior acid
o experimental data under the different conditions tested.
48 h 1 week 1 month
� 0.17a 0.93� 0.15a 0.60� 0.17a 0.38� 0.06a
� 0.25b 3.08� 0.38bc 2.34� 0.69bc 2.04� 0.30bc
� 0.70cd 4.54� 0.60de 3.67� 0.43de 2.73� 0.54bde
� 0.23e 2.33� 0.25f 2.16� 0.22b 1.72� 0.44bc
� 0.68cf 5.47� 0.71d 4.65� 0.43f 3.91� 0.52d
� 0.42d 4.41� 0.50de 4.07� 0.81dfe 3.56� 0.39d
� 0.68bd 3.94� 0.40be 4.42� 0.50df 3.89� 0.60d
� 0.32e 2.60� 0.57cf 2.41� 0.09b 2.49� 0.32be
� 0.56cd 4.50� 0.35de 4.20� 0.88dfe 3.69� 0.70de
� 0.11g 1.58� 0.15g 0.99� 0.09g 0.79� 0.09f
� 0.81f 6.40� 1.21d 4.99� 0.20f 3.92� 0.39d
� 0.88bd 3.40� 0.35bc 3.00� 0.37ce 2.26� 0.48bc
� 0.40b 3.47� 0.29bc 2.02� 0.30b 1.58� 0.18c
� 0.2g 1.37� 0.25ag 0.81� 0.09ag 0.59� 0.09f
f Enterococcus faecium (ATCC 49624).
.4 with: acetic, ascorbic, citric, lactic, malic and hydrochloric acids, respectively.erscript in the same column were statistically different (p< 0.05).

A. Fernandez et al. / Food Microbiology 26 (2009) 497–503 501
adaptation. Cultures were grown in buffered BHI (pH 7.4), non-acidified BHI and acidified BHI at pH values of 6.4 and 5.4 withseveral organic acids (acetic, ascorbic, citric, lactic and malic) andhydrochloric acid. We have also examined the heat inactivation of E.faecium grown in the different above mentioned conditions after itsexposure to a cold storage at 4 �C for 8 h, 48 h, one week and onemonth. In all cases, survival curves obtained were concave upward,so the traditional first-order kinetic model did not accuratelydescribe them. Although several mathematical models are suitablefor fitting concave upward survival curves (Alvarez et al., 2003;Chen and Hoover, 2003), a mathematical model based on theWeibull distribution was used because of its simplicity and flexi-bility (Peleg and Cole, 1998; Mafart et al., 2002; van Boekel, 2002).This Weibull model is appropriate since it only uses two parametersto describe the survival curves whatever their shape is: d (timeneeded to inactivate the first log10 cycle of the population) and pwhich accounts for upward concavity of a survival curve (p< 1),a linear survival curve (p¼ 1), and downward concavity (p> 1)(Mafart et al., 2002). Our results showed that the shape of survivalcurves obtained depended on the growth medium pH and the typeof acidulant used to adapt cells. The survival curves for cells grownat pH 6.4 were markedly concave upward (p-values¼ 0.31–0.43)and for those grown at pH 5.4 were slightly upwardly concaved,with p-values ranging from 0.42� 0.05 for ascorbic and malic acids,to 0.70� 0.09 for acetic acid. For non-acid-adapted cells (grown inbuffered BHI, pH 7.4) a p-value of 0.29� 0.04 was obtained.Although in several studies corresponding to the thermal inacti-vation of E. faecium a classical first-order kinetic has been reported(Sanz Perez et al., 1982; Ahmad et al., 2002, 2003; Martınez et al.,2003; Houben and Tjeerdsma-van Bokhoven, 2004), the survivalcurves frequently obtained are those in which an exponentialkinetic is followed by a tailing where the number of survivorsremain constant or decrease slowly (Sanz Perez et al., 1982; Magnuset al., 1986; Silva et al., 2003). The exact nature of this tailing effectis not exactly known, but in practice it may be of great importance,specially in the particular case of a target microorganism. Thetraditional log-linear thermal processing calculations do not takeinto account the effect of deviations of the linearity of survivalcurves on microbial heat resistance. So, understanding the condi-tions that can affect this microbial behaviour could be important inestablishing accurate thermal processes.
Results of the present study show that the growth of E. faecium(ATCC 49624) in presence of all the acids tested enhanced itssubsequent heat resistance, although the extent of this increasevaried among acidulants. The times needed to inactivate 2.5 log10
cycles (1.92� 0.15 min) for cells grown in buffered BHI (non-acid-adapted cells) were slightly increased, for ascorbic-acid adaptedcells, which reached values of about 1.3-fold higher. This increasewas higher, around 2–3-fold, for the corresponding adapted cellswith lactic, malic and hydrochloric acids and even higher (around5-fold) for acetic and citric-adapted cells. It has also been found thatafter the growth of E. faecium in non-acidified BHI a cross-protec-tion against heat was developed. The degree of the toleranceacquired was similar to that caused by growing cells in lactic andmalic acids and even higher than that induced by hydrochloric andascorbic acids. It is important to note that BHI is a medium whichcontains 0.2% glucose and thus, the fermentation of sugar leads toa reduction of the extracellular pH value which fell to around 6.2after 8 h of incubation (Table 2), which probably causes thedevelopment of an acid adaptation response in E. faecium cells.These findings support previous researches indicating that theinclusion of a fermentable carbohydrate in a growth medium is aneffective means of inducing cross-protection against heat(Buchanan and Edelson, 1999; Wilde et al., 2000; Bacon et al., 2003;Tosun and Gonul, 2003; Alvarez-Ordonez et al., 2008).
The ability of microorganisms to adapt to a moderate acidicenvironment and, subsequently, become more heat resistant hasbeen extensively studied for several foodborne pathogenic bacteria(Farber and Pagotto, 1992; Lou and Yousef, 1996; Juneja et al., 1998;Buchanan and Edelson, 1999; Duffy et al., 2000; Wilde et al., 2000;Mazzotta, 2001; Bacon et al., 2003; Evrendilek and Zhang, 2003;Tosun and Gonul, 2003; Alvarez-Ordonez et al., 2008). However, asfar as we know, only two studies have been performed to examinethe effectiveness of organic acids in inducing this protectiveresponse. Juneja et al. (1998) showed that lactic acid was moreefficient than acetic acid in increasing the thermotolerance ofacid-adapted L. monocytogenes Scott A cells. In contrast, Alvarez-Ordonez et al. (2008) found for S. typhimurium grown in acidifiedmedia up to pH 4.5 with acetic, citric, lactic and hydrochloric thatthe cross-protection response was not affected by the identity ofacidulant.
The mechanisms involved in the development of bacterial acidadaptation are still not completely understood, although evidencefor the involvement of internal pH maintenance was found. Thus,the different degree on microbial heat resistance conferred byorganic acids could be associated with their different ability to alterthe internal pH (pHi) of the cells (Cotter et al., 1999; Foster and Hall,1991). Certain organic acids may enter into the cell more easily thanothers and as a result the pHi is altered more quickly. Acetic acid,due to its low molecular weight and its great liposolubility, diffusesrapidly through the membrane, whereas lactic acid diffuses slowly.Citric acid, which is a tricarboxylic acid, with pKa1–3 values of 3.14;4.77; 6.39, once in the cell may be dissociated releasing threeprotons which cause more acidification than malic and ascorbicacids. On the other hand, hydrochloric acid fully dissociated in theextracellular medium is incapable of diffusing into the cell. In ourcase, after the determination of the pHi of stationary phase E. fae-cium cells grown in media acidified at pH 5.4 with all the differentorganic acids tested and hydrochloric acid non-significant differ-ences in pHi were found, obtaining a mean value of 7.5� 0.1 (datanot shown). These findings suggest that the pHi maintenance couldnot account for the different ability of acids in inducing the cross-protection response observed for E. faecium (ATCC 49624).However, the mechanisms of pHi maintenance are very complex.Various systems designed to alleviate acid stress throughconsumption of protons by aminoacid decarboxylases or itselimination by specific membrane transport systems present inmicroorganisms could have a different effectiveness (Kieboom andAbee, 2006; Lin et al., 1996). As an example, the HþATPase optimumpH value (pH0) varies even though within related microorganisms;whereas the HþATPase in Streptococcus thermophilus and Lacto-coccus lactis presents values of pH0 of 7.0–7.5, in Lactobacillus caseiand Lactobacillus plantarum is lower (5.0–5.5) (Siegumfeldt et al.,1999). On the other hand, it has been shown that microorganismsin response to a range of environmental stresses can developa series of constitutive and inducible strategies, which result inchanges in composition, density and fluidity of cellular membrane,production of several stress-induced proteins and chaperones,expression of transcriptional regulators, such as sigma factors,which alone or in combination can also be involved in the increasedstress tolerance which results in a cross-protection response(Annous et al., 1999; Foster, 2000; Dodd and Aldsworth, 2002;Sampathkumar et al., 2004; Alvarez-Ordonez et al., 2008, 2009).
A further finding from this study was that the cold storage underall conditions assayed progressively decreased the thermotoleranceof E. faecium (ATCC 49624), specially for non-acid-adapted cells.Our results also show that heat tolerance decreased as the growthmedium pH did, except when acetic and citric acid were used.Although little attention has been paid to the thermal inactivationof acid-adapted microorganisms exposed to low temperatures, an

A. Fernandez et al. / Food Microbiology 26 (2009) 497–503502
agreement appears to be in the fact that refrigeration increases thelethality of a subsequent heat treatment. Ingham and Uljas (1998)found that a prior storage at 4 �C for 24 h of E. coli in apple juiceacidified with malic acid at pH 3.4 significantly decreased its heatresistance. Alvarez-Ordonez et al. (2009) found that the refrigera-tion at 4 �C of hydrochloric acid-adapted S. senftenberg for one weekreduced its heat resistance, specially when the growth medium pHwas decreased. Our results confirm these previous studies and gofurther because in the present study we have demonstrated, fora wide range of growth conditions, that E. faecium (ATCC 49624)cold stressed cells turned out to be less heat resistant. These find-ings could have important consequences, as the common practiceof the previous use of refrigeration to a heat treatment minimizesthe risk that a sufficient portion of a microbial population couldpersist after the thermal process.
Microorganisms respond to cold stress in a variety of ways. It hasbeen pointed out that the systems that assist them to survive andadapt to environmental stresses may be decreased at low temper-atures or inhibited by factors expressed under these conditions,such as shock proteins or sigma factor (Goldstein et al., 1990; Linet al., 1996). Furthermore, cellular membrane has been implicatedas an important site in the microbial thermal destruction. A linkbetween the membrane fatty acid composition and the bacterialresistance has been shown in various studies (Annous et al., 1999;Sampathkumar et al., 2004; Alvarez-Ordonez et al., 2008). Ingeneral, these researchers have shown that cells with a decreasedconcentration of unsaturated fatty acids (UFA) or with an increasedcontent of saturated fatty acids (SFA) have a decreased membranefluidity which is linked to a higher heat resistance. Recently, it hasalso been demonstrated that storage under refrigeration conditionscaused an increase in the UFA/SFA ratio for acid-adapted S. senf-tenberg which were associated with a lower heat resistance(Alvarez-Ordonez et al., 2009).
To sum up, evidence provided in this study showed that theexposure of E. faecium (ATCC 49624) to simulated environmentalstresses, which are commonly used as food preservation methodsto control microbial growth, has significant effects on its subse-quent thermal inactivation. Whereas the acid adaptation resultedin a heat induced tolerance, a heat sensitization after cold storagewas found. As a result, differences in heat resistance in the moreextreme conditions up to 25-fold were observed. These findingshighlight the need for a better understanding of microbial responseto various preservation stresses in order to increase the efficiency ofthermal processes and to indicate the convenience of counter-balancing the benefits of the hurdle concept.
Acknowledgements
Fernandez A. and Alvarez-Ordonez A. gratefully acknowledgethe financial support from the University of Leon and SpanishMinistry of Education, respectively.
References
Ahmad, M., Smith, D.G., Mahboob, S., 2002. Effect of NaCl on heat tolerance ofEnterococcus faecium and Enterococcus faecalis. OnLine. J. Biol. Sci. 2 (7),483–484.
Ahmad, M., Smith, D.G., Mahboob, S., 2003. Determination of heat resistance ofexponential phase enterococcal cells. OnLine. J. Biol. Sci. 3, 440–446.
Alvarez, I., Virto, R., Raso, J., Condon, S., 2003. Comparing predicting models for theEscherichia coli inactivation by pulsed electric fields. Innovat. Food Sci. Emerg.Technol. 4, 195–202.
Alvarez-Ordonez, A., Fernandez, A., Lopez, M., Arenas, R., Bernardo, A., 2008.Modifications in membrane fatty acid composition of Salmonella typhimuriumin response to growth conditions and their effect on heat resistance. Int. J. FoodMicrobiol. 123, 212–219.
Alvarez-Ordonez, A., Fernandez, A., Lopez, M., Bernardo, A., 2009. Relationshipbetween membrane fatty acid composition and heat resistance of acid and cold
stressed Salmonella senftenberg cells CECT 4384. Food Microbiol.,doi:10.1016/j.fm.2008.11.002.
Annous, B.A., Kozempel, M.F., Kurantz, M.J., 1999. Changes in membrane fatty acidcomposition of Pediococcus sp. strain NRRL B-2354 in response to growth condi-tions and its effect on thermal resistance. Appl. Environ. Microbiol. 65, 2857–2862.
Bacon, R.T., Ransom, J.R., Sofos, J.N., Kendall, P.A., Belk, K.E., Smith, G.C., 2003.Thermal inactivation of susceptible and multiantimicrobial-resistant Salmonellastrains grown in the absence or presence of glucose. Appl. Environ. Microbiol.69, 4123–4128.
Buchanan, R.L., Edelson, S.G., 1999. Effect of pH-dependent, stationary phase acidresistance on the thermal tolerance of E. coli O157:H7. Food Microbiol.16, 447–458.
van Boekel, M.A.J.S., 2002. On the use of the Weibull model to describe thermalinactivation of microbial vegetative cells. Int. J. Food Microbiol. 74, 139–159.
Centeno, J.A., Menendez, S., Hermida, M.A., Rodrıguez-Otero, J.L., 1999. Effects of theaddition of Enterococcus faecalis in Cebreiro cheese manufacture. Int. J. FoodMicrobiol. 48, 97–111.
Chen, H., Hoover, D.G., 2003. Pressure inactivation kinetics of Yersinia enterocoliticaATCC 35669. Int. J. Food Microbiol. 87, 161–171.
Cintas, L.M., Casaus, P., Havarstein, L.S., Hernandez, P.E., Nes, I.F., 1997. Biochemicaland genetic characterization of enterocin P, a novel sec-dependent bacteriocinfrom Enterococcus faecium P13 with a broad antimicrobial spectrum. Appl.Environ. Microbiol. 63, 4321–4330.
Condon, S., Arrizubieta, M.J., Sala, F.J., 1993. Microbial heat resistance determina-tions by the multipoint system with the thermoresistometer TR-SC.Improvement of this methodology. J. Microbiol. Methods 18, 357–366.
Cotter, P.D., Emerson, N., Gahan, C.G., Hill, C., 1999. Identification and disruption oflisRK, a genetic locus encoding a two-component signal transduction systeminvolved in stress tolerance and virulence in Listeria monocytogenes. J. Bacteriol.181, 6840–6843.
Devriese, L.A., Pot, B., Collins, M.D., 1993. Phenotypic identification of the genusEnterococcus and differentiation of phylogenetically distinct enterococcalspecies and species groups. J. Appl. Bacteriol. 75, 399–408.
Dodd, C.E.R., Aldsworth, T.G., 2002. The importance of RpoS in the survival ofbacteria through food processing. Int. J. Food Microbiol. 74, 189–194.
Duffy, G., Riordan, D.C.R., Sheridan, J.J., Call, J.E., Whiting, R.C., Blair, I.S.,McDowell, D.A., 2000. Effect of pH on survival, thermotolerance and verotoxinproduction of E. coli O157:H7 during simulated fermentation and storage. J.Food Prot. 63, 12–18.
Evrendilek, G.A., Zhang, Q.H., 2003. Effects of pH, temperature, and pre-pulsedelectric field treatment on pulsed electric field and heat inactivation ofEscherichia coli O157:H7. J. Food Prot. 66, 755–759.
Farber, J.M., Pagotto, F., 1992. The effect of acid shock on the heat resistance ofListeria monocytogenes. Lett. Appl. Microbiol. 15, 197–201.
Flahaut, S., Frere, J., Boutibonnes, P., Auffray, Y., 1997. Relationship between thethermotolerance and the increase of DnaK and GroEL synthesis in Enterococcusfaecalis ATCC 19433. J. Basic Microbiol. 37, 251–258.
Foster, J.W., Hall, H.K., 1991. Inducible pH homeostasis and the acid toleranceresponse of Salmonella typhimurium. J. Bacteriol. 173, 5129–5135.
Foster, J.W., 2000. Microbial responses to acid stress. In: Storz, G., Hengge-Aronis, R.(Eds.), Bacterial Stress Responses. ASM Press, Washington DC, pp. 99–115.
Franz, C.M.A.P., Stiles, M.E., Schleifer, K.H., Holzapfel, W.H., 2003. Enterococci infoods – a conundrum for food safety. Int. J. Food Microbiol. 88, 105–122.
Ghazala, S., Ramaswamy, S., Smith, J.P., Sipmpson, B.K., Simpson, M.V., 1991.Establishing sous-vide processing schedules for spaghetti meat sauce and ricesalmon based on thermal destruction of Streptococcus faecium. In: Proceedingsof XVIIIth International Congress of Refrigeration, Montreal.
Ghazala, S., Coxworthy, D., Alkanani, T., 1995. Thermal kinetics of Streptococcusfaecium in nutrient broth/sous vide products under pasteurization conditions. J.Food Process. Preserv. 19, 243–257.
Giraffa, G., Carminati, D., Neviani, E., 1997. Enterococci isolated from dairy products:a review of risks and potential technological use. J. Food Prot. 60, 732–738.
Goldstein, J.N., Pollitt, S., Inouye, M., 1990. Major cold shock protein of Escherichiacoli. Proc. Natl. Acad. Sci. U.S.A. 87, 283–287.
Houben, J.H., 1982. Heat resistance of Streptococcus faecium in pasteurized ham.Fleischwirtschaft 62, 490–493.
Houben, J.H., Tjeerdsma-van Bokhoven, J.L.M., 2004. Growth inhibition of heat-injured Enterococcus faecium by oligophosphates in a cured meat model. Int. J.Food Microbiol. 97, 85–91.
Ibarz, P., Palop, A., Condon, S., Sala, F.J., 1991. Adaptador de un contador automaticode U.F.C. al recuento de placas densamente pobladas. In: Proceedings of the XIIICongreso Nacional de Microbiologıa, Salamanca, Spain.
Ingham, S.C., Uljas, H.E., 1998. Prior storage conditions influence the destruction ofE. coli O157:H7 during heating of apple cider and juice. J. Food Prot. 61, 390–394.
Juneja, V.K., Foglia, T.A., Marmer, B.S., 1998. Heat resistance and fatty acid compo-sition of Listeria monocytogenes: effect of pH, acidulant, and growthtemperature. J. Food Prot. 61, 683–687.
Kearns, A.M., Freeman, R., Lightfoot, N.F., 1995. Nosocomial enterococci: resistanceto heat and sodium hypochlorite. J. Hosp. Infect. 30, 193–199.
Kieboom, J., Abee, T., 2006. Arginine-dependent acid resistance in Salmonellaenterica serovar Typhimurium. J. Bacteriol. 188, 5650–5653.
Lin, J., Smith, M.P., Chapin, K.C., Baik, H.S., Bennet, G.N., Foster, J.W., 1996. Mecha-nisms of acid resistance in enterohemorrhagic Escherichia coli. Appl. Environ.Microbiol. 62, 3094–3100.
Lou, Y., Yousef, A.E., 1996. Resistance of Listeria monocytogenes to heat after adap-tation to environmental stresses. J. Food Prot. 59, 465–471.

A. Fernandez et al. / Food Microbiology 26 (2009) 497–503 503
Magnus, C.A., Ingledew, W.M., McCurdy, A.R., 1986. Thermal resistance ofstreptococci isolated from pasteurized ham. Can. Inst. Food Sci. Technol. J. 19(2), 62–67.
Mafart, P., Couvert, O., Gaillard, S., Leguerinel, I., 2002. On calculating sterility inthermal preservation methods: application of the Weibull frequency distribu-tion model. Int. J. Food Microbiol. 72, 107–113.
Martınez, S., Lopez, M., Bernardo, A., 2003. Thermal inactivation of Enterococcusfaecium: effect of growth temperature and physiological state of microbial cells.Lett. Appl. Microbiol. 37, 475–481.
Mazzotta, A.S., 2001. Thermal inactivation of stationary-phase and acid-adapted E.coli O157:H7, Salmonella and Listeria monocytogenes in fruit juices. J. Food Prot.64, 315–320.
Murray, B.E.,1990. The life and times of the Enterococcus. Clin. Microbiol. Rev. 3, 46–65.Parente, E., Cogan, T.M., 2004. Starter cultures: general aspects. In: Fox, P.F.,
McSweeney, P.L.H., Cogan, T.M., Guinee, T.P. (Eds.), Cheese, Chemistry, Physicsand Microbiology, third ed. Elsevier Academic Press, London, pp. 123–148.
Peleg, M., Cole, M.B., 1998. Reinterpretation of microbial survival curves. Crit. Rev.Food Sci. Nutr. 38, 353–380.
Sampathkumar, B., Khachatourians, G.G., Korber, D.R., 2004. Treatment of Salmo-nella enterica serovar Enteritidis with a sublethal concentration of trisodiumphosphate or alkaline pH induces thermotolerance. Appl. Environ. Microbiol.70, 4613–4620.
Sanz Perez, B., Lopez Lorenzo, P., Garcıa, M.L., Hernandez, P.E., Ordonez, J.A., 1982.Heat resistance of enterococci. Milchwissenschaft 37, 724–726.
Siegumfeldt, H., Rechinger, K.B., Jakobsen, M., 1999. Use of fluorescence ratioimaging for intracellular pH determination of individual bacterial cells in mixedcultures. Microbiology 145, 1703–1709.
Silva, M., Ramos, M., Costa, C., do Carmo, M., Giambiagi-deMarval, M., 2003. Heat-resistance and heat-shock response in the nosocomial pathogen Enterococcusfaecium. Curr. Microbiol. 46, 313–317.
Smith, J.P., Toupin, C., Gagnon, B., Voyer, R., Fiset, P.P., Simpson, M.V., 1990. A hazardanalysis critical control point approach (HACCP) to ensure the microbiologicalsafety of sous vide processed meat/pasta product. Food Microbiol. 7, 177–198.
Spicher, G., Peters, J., Nurnberg, M., Schwebke, I., 2002. Microbicidal efficacy ofsuperheated steam. II. Studies involving E. faecium and spores of B. xero-thermodurans and B. coagulans. Int. J. Hyg. Environ. Health 204, 309–316.
Steel, R.G.D., Torrie, J.H., 1986. Comparacion entre dos medias muestrales. In:Bioestadıstica: Principios y Procedimientos. McGraw Hill, Madrid, pp. 83–116.
Tosun, H., Gonul, S.A., 2003. Acid adaptation protects Salmonella typhimurium fromenvironmental stress. Turk. J. Biol. 27, 31–36.
Wilde, S., Jorgensen, F., Campbell, A., Rowbury, R., Humphrey, T., 2000. Growth ofSalmonella enterica serovar Enteritidis PT4 in media containing glucose resultsin enhanced RpoS-independent heat and acid tolerance but does not affect theability to survive air-drying on surfaces. Food Microbiol. 17, 679–686.








![Research Article Antibacterial Derivatives of ...Journal of Pharmaceutics (%), and Enterococcus faecium (%) [ ]. E. coli was the most common bacteria associated with necrotizing fasciitis](https://img.pdfslide.us/doc/110x75/611eb892c14ac75a1342aa16/research-article-antibacterial-derivatives-of-journal-of-pharmaceutics-.jpg)







![EasyGenotyping ITS-PCR - DNA GDAŃSK · PDF file˜’ rrs rrl rrf A D BC ... Staphylococcus aureus, Enterococcus faecium). Reagent Volume [µl] MMix for tests Sum [µl] To be portioned](https://img.pdfslide.us/doc/110x75/5aa1fe417f8b9aa0108c99cc/easygenotyping-its-pcr-dna-gdansk-rrs-rrl-rrf-a-d-bc-staphylococcus.jpg)












