Embed Size (px)
Citation preview

Plant and Soil 233: 261–268, 2001.© 2001 Kluwer Academic Publishers. Printed in the Netherlands.
261
Effects of NaCl and Na2SO4 on red-osier dogwood (Cornus stoloniferaMichx) seedlings
Sylvie Renault1,2, Clare Croser, Jennifer A. Franklin & Janusz J. ZwiazekDepartment of Renewable Resources, University of Alberta 4-42 Earth Sciences Bldg, Edmonton, AB, CanadaT6G 2E3. 1Department of Botany, University of Manitoba 505 Buller Building, Winnipeg, MB, Canada R3T 2N2.2Corresponding author∗
Received 4 July 2000. Accepted in revised form 4 April 2001
Key words: Cornus stolonifera, gas exchange, growth, ions, salinity, salt resistance
Abstract
Sodium chloride and sodium sulfate are commonly present in extraction tailings waters produced as a result ofsurface mining and affect plants on reclaimed areas. Red-osier dogwood (Cornus stolonifera Michx) seedlings weredemonstrated to be relatively resistant to these high salinity oil sands tailings waters. The objectives of this studywere to compare the effects of Na2SO4 and NaCl, on growth, tissue ion content, water relations and gas exchangein red-osier dogwood (Cornus stolonifera Michx) seedlings. In the present study, red-osier dogwood seedlingswere grown in aerated half-strength modified Hoagland’s mineral solution containing 0, 25, 50 or 100 mM ofNaCl or Na2SO4. After four weeks of treatment, plant dry weights decreased and the amount of Na+ in planttissues increased with increasing salt concentration. Na+ tissue content was higher in plants treated with NaCl thanNa2SO4 and it was greater in roots than shoots. However, Cl− concentration in the NaCl treated plants was higherin shoots than in roots. The decrease in stomatal conductance and photosynthetic rates observed in presence ofsalts is likely to contribute to the growth reduction. Our results suggest that red-osier dogwood is able to controlthe transport of Na+ from roots to shoots when external concentrations are 50 mM or less.
Introduction
Soils containing elevated levels of NaCl and Na2SO4are naturally present in many forested areas. Surfacemining has aggravated the salinity problems in someareas of the boreal forest in western Canada. Thedevelopment of a successful reclamation strategy forthese areas will depend on understanding the resist-ance mechanisms of boreal forest woody plants tosalinity. However, little is known about the response ofthese plants to an excess of Na+, Cl− and SO4
2− in thesoil solution. Our previous studies found that red-osierdogwood (Cornus stolonifera Michx) is relatively tol-erant to the high salinity tailings waters produced as aresult of oil sand mining (Renault et al., 1998, 1999).Red-osier dogwood is a rapidly growing woody plantwell adapted to disturbed sites. It is also a native boreal
∗ Fax No.: +204-474-7604. E-mail: [email protected]
species that is excellent at stabilizing soil (Hardy BBTLtd., 1989) and attracting wildlife (Robbins and Moen,1975), qualities that make this species suitable for landreclamation.
In our previous study, we demonstrated that red-osier dogwood can accumulate as much as 5 mg g−1
tissue dry weight of Na+ in the whole plant withoutexhibiting visible injury (Renault et al., 1999). How-ever, since Na+ is often accompanied by Cl− and(or) SO4
2− both in naturally saline areas and in theindustrial reclamation areas, there is a need to under-stand how these anions interact with Na+ to affectplant physiology and growth. Salinity can reduce plantgrowth by both osmotic and ionic effects (Green-way and Munns, 1980). An accumulation of ions inplant tissues can affect membrane selective permeab-ility thus altering the uptake of ions (Khan et al.,1997). The resulting ion imbalance may cause nu-trient deficiency or toxicity. In addition, salinity can

262
reduce photosynthesis due to an increase in stomatalresistance and disturbance of photosynthetic pigmentsbiosynthesis (Banuls and Primo Millo, 1992; Khanet al., 1997). Chloride ions have been reported to bemore toxic than Na+ in some species of woody plants(Greenway and Munns, 1980; Shannon et al., 1994).Eucalyptus and citrus species have shown extensiveleaf injury when grown in presence of Cl− (Marcarand Termaat, 1990; Maas, 1993). However, becauserelatively few studies have focused on the effects ofNa2SO4 on plant growth, the effects of the SO4 anionare relatively less known. The objectives of this studywere to examine the effects of both NaCl and Na2SO4on (1) growth and morphology (2) gas exchange andwater relations and (3) tissue ion content in red-osierdogwood seedlings.
Materials and methods
Plant material and culture conditions
Dogwood (Cornus stolonifera Michx) seeds were col-lected at the Syncrude mining site, north of FortMcMurray, AB, Canada (57◦ 01.67′ N, 111◦ C30.6′W). The seeds were stratified for 6 weeks at 4 ◦Cand sown in a mixture of peat moss and sand (3:1,by volume) in a greenhouse maintained under thefollowing conditions: day/night temperature 25/18◦Cwith a 16 h photoperiod supplemented by 400 Whigh pressure sodium lamps (GTE Sylvania Lumalux,Drummondville, PQ, Canada). After 3 months, theseedlings were transferred to aerated solution culturecontaining half-strength modified Hoagland’s solution(Renault et al., 1998) for 4 days prior to the com-mencement of salt treatments for acclimatization tothe new conditions.
On the first day of treatment, seedlings were trans-ferred to solution culture containing the following con-centrations of salt dissolved in half-strength modifiedHoagland’s mineral solution: 25 mM NaCl (−0.16MPa), 50 mM NaCl (−0.29 MPa), 100 mM NaCl,(−0.56 MPa), 25 mM Na2SO4 (−0.23 MPa), 50 mMNa2SO4 (−0.43 MPa), or 100 mM Na2SO4 (−0.83MPa) or to a control solution containing only half-strength modified Hoagland’s solution. There were 30seedlings per treatment in two replicated experiments.The solutions were replaced once a week to avoid nu-trient deficiency. Shoot water potentials, transpirationrates and stomatal conductance were measured after1 day, 1 week and four weeks of treatment. At each
of these times, five plants from each treatment andreplicate were randomly selected for measurements.Plant survival, growth (shoot height, number of lateralshoots, root and shoot dry weights), photosyntheticrates, chlorophyll and ion content of dogwood seed-lings were determined at the end of the experiment.
Gas exchange
The stomatal conductance of attached leaves wasmeasured with a steady state porometer (Lambda In-struments Corp., Lincoln, NE, USA). The same leaveswere used to measure photosynthetic rates using theLCA-4 infrared gas analyzer (Analytical DevelopmentCompany Ltd., Hoddesdon, UK). The measurementswere performed between 10:00 and 16:00 on one leafof the uppermost pair of fully expanded leaves. Photo-synthetically active radiation was 450 µmol m−2 s−1,atmospheric CO2 252 µmol mol−1 and the relative hu-midity 50%. Leaf areas were measured with a LI 3000leaf area meter (Lambda Instruments Corp., Lincoln,NE, USA).
Shoot water potentials
Upon completion of gas exchange measurements oneach plant (between 10:00 and 16:00), the top 15-cmof shoot was excised and used to measure water po-tentials using a Scholander pressure chamber (PMSInstrument Co., Corvallis, OR, USA) operated at0.02 MPa s−1 pressurization rate, as described byScholander et al. (1965).
Ion and chlorophyll contents
Seedlings were harvested, washed in deionized wa-ter, and the shoots and roots separated and freeze-dried. Na+, Mg2+ and K+ were determined in eachplant sample by atomic absorption spectrophotometry(Perkins-Elmer 503) after digestion with H2SO4 andH2O2 at 360 ◦C (2 × 30 min). Chloride was determ-ined using a Cl−selective electrode (Accumet, FisherScientific, Edmonton, AB) after extracting ground tis-sues with 0.5 m HNO3 for 30 min at room temperature(Rieger and Litvin, 1998).
For the chlorophyll analyses, two fully expan-ded leaves were collected after 4 weeks of treatment.Eight leaf discs (0.5 cm2) were cut from the leavesand incubated in three changes (2 ml) of methanol.The chlorophyll concentration in the methanol extractswas determined spectrophotometrically and calculatedusing the MacKinney equations (Sestak et al., 1971).

263
Data analysis
The data were analyzed with a general linear model(GLM) procedure using a one way ANOVA. Themeans were compared using the Duncan’s multiplerange test.
Results
Survival and growth
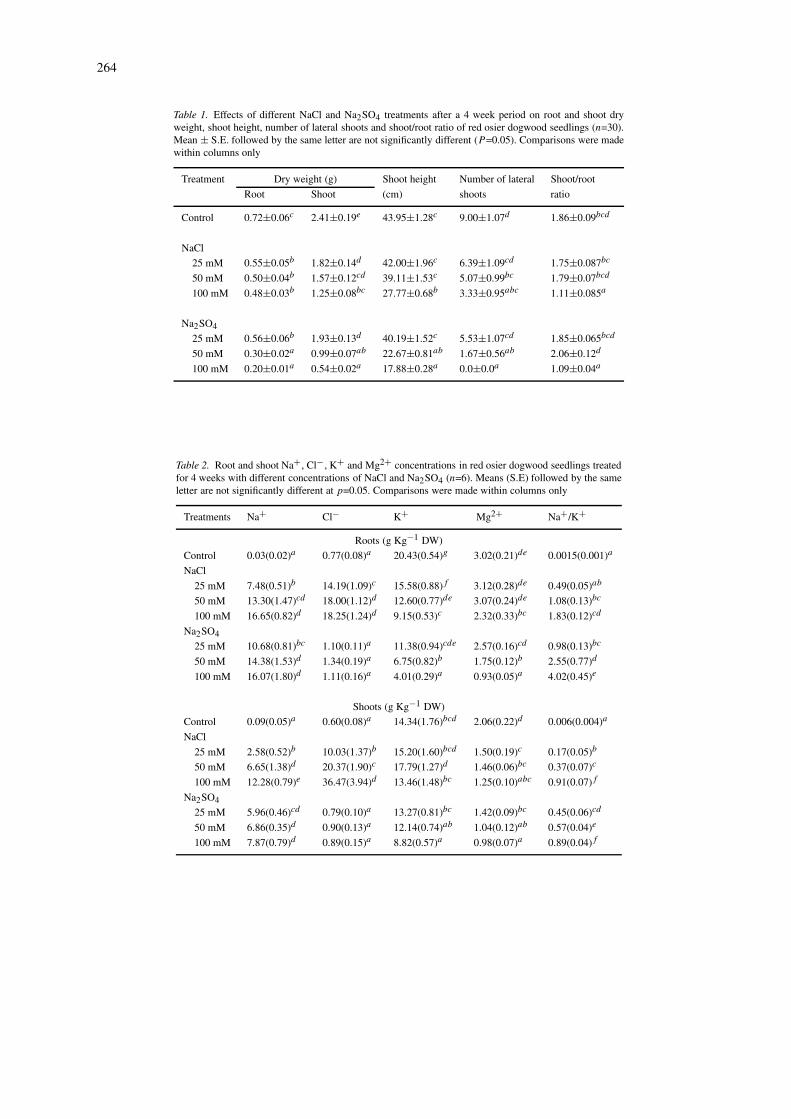
All plants survived treatment with control, 25 and50 mM salt solutions. However, after 4 weeks, therewas only 30% survival in 100 mM NaCl and 20%survival in 100 mM Na2SO4. Shoot and root dryweights of seedlings were significantly reduced by allsalt concentrations (Table 1). We observed significantdifferences between salts: 50 mM Na2SO4 reducedroot dry weight by 58% while 100 mM NaCl reducedit by only 33%. Similar trends were observed for theshoot dry weights. Root and shoot dry weights werereduced in 100 mM Na2SO4 treatment by 72% and78%, respectively (Table 1). However, shoot heightwas reduced only by the 100 mM NaCl treatment andby 50 mM and 100 mM Na2SO4 treatments (Table 1).The number of lateral shoots decreased in 50 mM and100 mM of both salts, NaCl and Na2SO4 (Table 1).Shoot/root ratio decreased from 1.86 in control seed-lings to 1.11 and 1.09 in plants treated with 100 mMNaCl and Na2SO4 respectively (Table 1).
Ion content
Na+ content of the roots and shoots increased with in-creasing concentration of NaCl and Na2SO4 (Table 2).Although no statistical comparison was made, theconcentrations were higher in roots than in shoots(Table 2). In the higher salt treatments, the concen-tration of Na+ in the shoots was higher in NaCl-treated plants than in Na2SO4-treated plants (based onequimolar Na+ concentrations). There was 12.28 gkg−1 DW Na+ present in the shoots of seedlingstreated with 100 mM NaCl, while in a compar-able Na+ concentration treatment of 50 mM Na2SO4,shoots accumulated only 6.86 g kg−1 DW (Table 2).Cl− accumulated in NaCl-treated plants to a greaterextent than Na+ (Table 2). In 25 mM NaCl, the con-centration of Cl− was higher in the roots than in theshoots, while in 100 mM NaCl the amount of Cl− wasgreater in the shoots than in the roots. K+ concentra-tion in roots decreased with increasing salt concentra-
tion (Table 2). The decrease was more pronounced in50 mM Na2SO4 than in 100 mM NaCl treatments. Inshoots, the only significant reduction of K+ observedwere in the plants treated with 100 mM Na2SO4.Mg2+ concentration in the root tissue was significantlyreduced in the 100mM NaCl treatment and 50 mM and100 mM Na2SO4 treatments. However, in the shoottissues, Mg2+ concentration was significantly reducedin all salt treatments. The Na+/K+ ratio increased sig-nificantly with increasing concentrations for both salttreatments in root and shoot tissues (Table 2).
Water relations
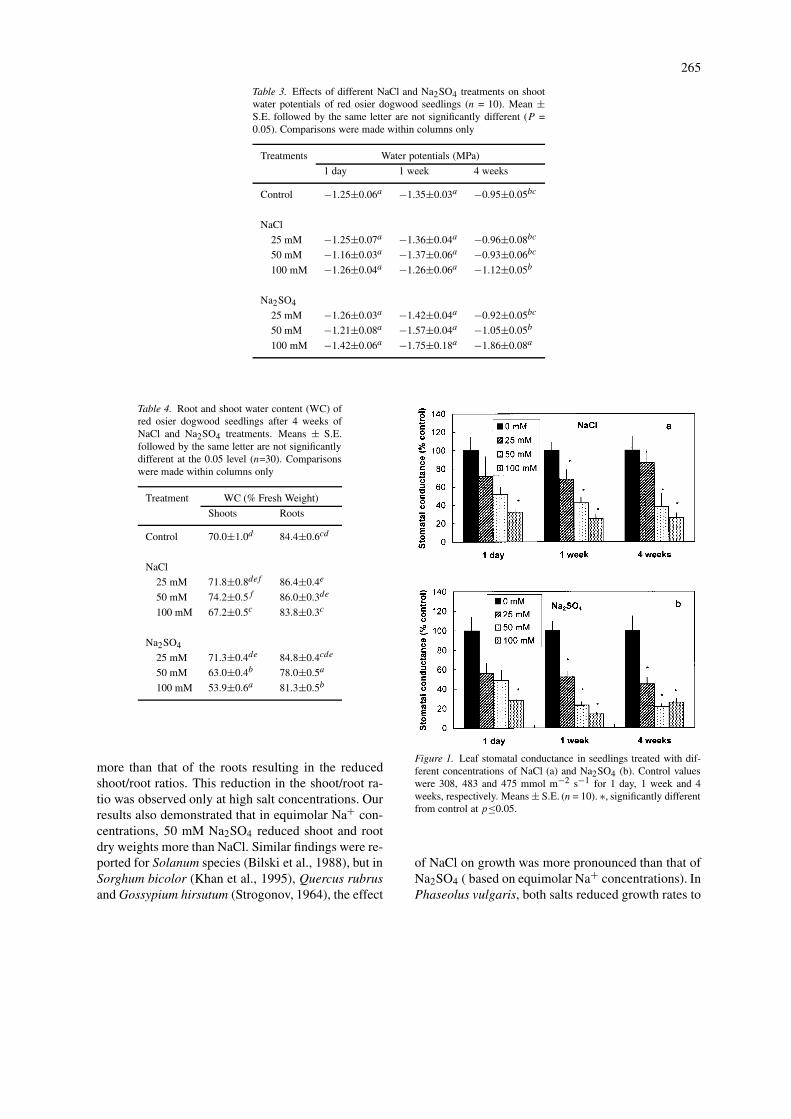
Salt treatments had little effect on shoot water po-tentials. The only significant decrease in shoot waterpotentials occurred in plants treated for 4 weeks with100 mM Na2SO4 (Table 3).
Shoot water content was significantly reduced in50 and 100 mM Na2SO4 and by 100 mM NaCl treat-ments, but increased in the 50 mM NaCl treatment(Table 4). A significant decrease in root water contentwas recorded in 50 and 100 mM Na2SO4 treatments,but root water content increased in the 25 mM NaCltreatment.
Chlorophyll content and gas exchange
Leaf chlorophyll a and chlorophyll b concentrationsin seedlings treated with 100 mM Na2SO4 were 66%and 56% respectively of the values found in controlplants (Table 5). All remaining treatments did notsignificantly affect chlorophyll a or chlorophyll b con-centration. In addition, chlorophyll a/b ratio was notaffected by the salt treatments (Table 5).
Concentrations of 50 and 100 mM Na2SO4 and100 mM NaCl decreased the photosynthetic rates to5%, 0.2% and 16%, respectively, of the rates measuredin the control seedlings (Table 5).
Stomatal conductance declined significantly afteronly one day of treatment in 100 mM NaCl and100 mM Na2SO4 (Figure 1). After 1 week, all treat-ments caused a reduction in stomatal conductance ofred-osier dogwood leaves (Figure 1).
Discussion
The results of our study showed that, similarly to otherwoody plants (Banuls and Primo-Millo, 1992; Funget al., 1998; Nabil and Coudret, 1995), the shootgrowth of red-osier dogwood was inhibited by salinity
264
Table 1. Effects of different NaCl and Na2SO4 treatments after a 4 week period on root and shoot dryweight, shoot height, number of lateral shoots and shoot/root ratio of red osier dogwood seedlings (n=30).Mean ± S.E. followed by the same letter are not significantly different (P =0.05). Comparisons were madewithin columns only
Treatment Dry weight (g) Shoot height Number of lateral Shoot/root
Root Shoot (cm) shoots ratio
Control 0.72±0.06c 2.41±0.19e 43.95±1.28c 9.00±1.07d 1.86±0.09bcd
NaCl
25 mM 0.55±0.05b 1.82±0.14d 42.00±1.96c 6.39±1.09cd 1.75±0.087bc
50 mM 0.50±0.04b 1.57±0.12cd 39.11±1.53c 5.07±0.99bc 1.79±0.07bcd
100 mM 0.48±0.03b 1.25±0.08bc 27.77±0.68b 3.33±0.95abc 1.11±0.085a
Na2SO4
25 mM 0.56±0.06b 1.93±0.13d 40.19±1.52c 5.53±1.07cd 1.85±0.065bcd
50 mM 0.30±0.02a 0.99±0.07ab 22.67±0.81ab 1.67±0.56ab 2.06±0.12d
100 mM 0.20±0.01a 0.54±0.02a 17.88±0.28a 0.0±0.0a 1.09±0.04a
Table 2. Root and shoot Na+, Cl−, K+ and Mg2+ concentrations in red osier dogwood seedlings treatedfor 4 weeks with different concentrations of NaCl and Na2SO4 (n=6). Means (S.E) followed by the sameletter are not significantly different at p=0.05. Comparisons were made within columns only
Treatments Na+ Cl− K+ Mg2+ Na+/K+
Roots (g Kg−1 DW)
Control 0.03(0.02)a 0.77(0.08)a 20.43(0.54)g 3.02(0.21)de 0.0015(0.001)a
NaCl
25 mM 7.48(0.51)b 14.19(1.09)c 15.58(0.88)f 3.12(0.28)de 0.49(0.05)ab
50 mM 13.30(1.47)cd 18.00(1.12)d 12.60(0.77)de 3.07(0.24)de 1.08(0.13)bc
100 mM 16.65(0.82)d 18.25(1.24)d 9.15(0.53)c 2.32(0.33)bc 1.83(0.12)cd
Na2SO4
25 mM 10.68(0.81)bc 1.10(0.11)a 11.38(0.94)cde 2.57(0.16)cd 0.98(0.13)bc
50 mM 14.38(1.53)d 1.34(0.19)a 6.75(0.82)b 1.75(0.12)b 2.55(0.77)d
100 mM 16.07(1.80)d 1.11(0.16)a 4.01(0.29)a 0.93(0.05)a 4.02(0.45)e
Shoots (g Kg−1 DW)
Control 0.09(0.05)a 0.60(0.08)a 14.34(1.76)bcd 2.06(0.22)d 0.006(0.004)a
NaCl
25 mM 2.58(0.52)b 10.03(1.37)b 15.20(1.60)bcd 1.50(0.19)c 0.17(0.05)b
50 mM 6.65(1.38)d 20.37(1.90)c 17.79(1.27)d 1.46(0.06)bc 0.37(0.07)c
100 mM 12.28(0.79)e 36.47(3.94)d 13.46(1.48)bc 1.25(0.10)abc 0.91(0.07)f
Na2SO4
25 mM 5.96(0.46)cd 0.79(0.10)a 13.27(0.81)bc 1.42(0.09)bc 0.45(0.06)cd
50 mM 6.86(0.35)d 0.90(0.13)a 12.14(0.74)ab 1.04(0.12)ab 0.57(0.04)e
100 mM 7.87(0.79)d 0.89(0.15)a 8.82(0.57)a 0.98(0.07)a 0.89(0.04)f
265
Table 3. Effects of different NaCl and Na2SO4 treatments on shootwater potentials of red osier dogwood seedlings (n = 10). Mean ±S.E. followed by the same letter are not significantly different (P =0.05). Comparisons were made within columns only
Treatments Water potentials (MPa)
1 day 1 week 4 weeks
Control −1.25±0.06a −1.35±0.03a −0.95±0.05bc
NaCl
25 mM −1.25±0.07a −1.36±0.04a −0.96±0.08bc
50 mM −1.16±0.03a −1.37±0.06a −0.93±0.06bc
100 mM −1.26±0.04a −1.26±0.06a −1.12±0.05b
Na2SO4
25 mM −1.26±0.03a −1.42±0.04a −0.92±0.05bc
50 mM −1.21±0.08a −1.57±0.04a −1.05±0.05b
100 mM −1.42±0.06a −1.75±0.18a −1.86±0.08a
Table 4. Root and shoot water content (WC) ofred osier dogwood seedlings after 4 weeks ofNaCl and Na2SO4 treatments. Means ± S.E.followed by the same letter are not significantlydifferent at the 0.05 level (n=30). Comparisonswere made within columns only
Treatment WC (% Fresh Weight)
Shoots Roots
Control 70.0±1.0d 84.4±0.6cd
NaCl
25 mM 71.8±0.8def 86.4±0.4e
50 mM 74.2±0.5f 86.0±0.3de
100 mM 67.2±0.5c 83.8±0.3c
Na2SO4
25 mM 71.3±0.4de 84.8±0.4cde
50 mM 63.0±0.4b 78.0±0.5a
100 mM 53.9±0.6a 81.3±0.5b
more than that of the roots resulting in the reducedshoot/root ratios. This reduction in the shoot/root ra-tio was observed only at high salt concentrations. Ourresults also demonstrated that in equimolar Na+ con-centrations, 50 mM Na2SO4 reduced shoot and rootdry weights more than NaCl. Similar findings were re-ported for Solanum species (Bilski et al., 1988), but inSorghum bicolor (Khan et al., 1995), Quercus rubrusand Gossypium hirsutum (Strogonov, 1964), the effect
Figure 1. Leaf stomatal conductance in seedlings treated with dif-ferent concentrations of NaCl (a) and Na2SO4 (b). Control valueswere 308, 483 and 475 mmol m−2 s−1 for 1 day, 1 week and 4weeks, respectively. Means ± S.E. (n = 10). ∗, significantly differentfrom control at p≤0.05.
of NaCl on growth was more pronounced than that ofNa2SO4 ( based on equimolar Na+ concentrations). InPhaseolus vulgaris, both salts reduced growth rates to

266
Table 5. Effects of different NaCl and Na2SO4 treatments after a 4 weekperiod on photosynthesis and chlorophyll a and b content of red osier dog-wood seedlings (n=10). Mean ± S.E. followed by the same letter are notsignificantly different (P = 0.05). Comparisons were made within columnsonly
Treatments Photosynthesis Chlorophyll a Chlorophyll b
(µmol CO2 m−2 s−1) (mg g−1 FW) (mg g−1 FW)
Control 1.89±0.37c 1.66±0.07bc 0.52±0.03bc
NaCl
25 mM 1.74±0.37c 1.50±0.11bc 0.50±0.06bc
50 mM 1.04±0.26bc 1.59±0.15bc 0.55±0.04c
100 mM 0.31±0.09ab 1.39±0.15abc 0.48±0.06bc
Na2SO4
25 mM 1.03±0.36bc 1.71±0.08c 0.54±0.04c
50 mM 0.10±0.03ab 1.33±0.06ab 0.36±0.07ab
100 mM 0.03±0.01a 1.11±0.13a 0.29±0.06a
the same extent (Meiri et al., 1971). Therefore, theeffects of salts on plant growth appear to be species-dependent. The reduction of plant growth during saltstress could be due to osmotic effects. Our resultsshowed that water content of plants treated with thehighest concentrations of salts was lower than that incontrol plants suggesting an osmotic effect. However,the differential effect of NaCl and Na2SO4 on growthin our experiment can not be explained by a differencein osmotic potential as the osmotic potential of 50 mMNa2SO4 solution was higher than the one of equimolarNa+ concentration of NaCl.
An excess of ions is known to cause ion toxicityand ion imbalance (Greenway and Munns, 1980). Inred-osier dogwood seedlings, an accumulation of Na+and Cl− in shoots increased with an increase in theexternal concentration of NaCl. Both Na+ and Cl− ac-cumulation has been reported for several other woodyplant species exposed to salinity (Banuls and Primo-Millo, 1992; Fung et al., 1998; Nabil and Coudret,1995). In many plants, salt resistance depends on theability of the root system to restrict Na+ and Cl−transport to the shoots (Grieve and Walker, 1983,Khan et al., 1995; Yeo et al., 1977). In our study,Na+ was present in higher concentrations in the rootscompared with the shoots, suggesting that Na+ waspreferentially stored in the roots and that this reducedthe potential for transport to the shoots, limiting theextent of shoot injury. In some plant species, Cl−uptake and transport appeared to be less controlled
than that of Na+ (Alaoui-Sasse et al., 1998). This hasbeen observed in Gleditsia tricanthos (Dirr, 1974) andSorghum bicolor (Khan et al., 1995), as well as inthe present study where Cl− concentration was higherthan Na+ concentration in shoot tissues. It has been re-ported that Cl− may be more injurious than Na+ (Dirr,1974; Marcar and Termaat, 1990, Shannon et al.,1994), and was found to cause leaf chlorosis in severalplant species (Alaoui-Sosse et al., 1998; Greenwayand Munns, 1980). However, to verify this hypo-thesis necrosis should be assessed at 100 mM salt andcorrelation with Cl− content should be investigated.
In the present study, Na+ content was higher inthe shoots treated with NaCl than those treated withNa2SO4 at isomolar levels of Na+. Similar resultswere reported in Gleditsia tricanthos (Dirr, 1974). Itis possible that chloride affects Na+ uptake by al-tering membrane lipid composition thus modifyingmembrane permeability. A change in membrane lipidcomposition in response to chloride was reported byKuiper (1968). In our study, an accumulation of Na+in red-osier dogwood seedling roots was accompan-ied by a decrease in K+ in the roots. Similar resultshave been reported for Quercus robur (Alaoui-Sasseet al., 1998) and Citrus reticulata (Ruiz et al., 1997).A high concentration of Na+ can interfere with K+uptake, resulting in deficiency and stunted growth. Ithas been suggested that in Vicia faba the two ions com-pete for uptake at the plasma membrane level (Ullah,1993). A high concentration of Na+ displaces Ca2+

267
from root membranes thus affecting Na+/K+ selectiv-ity (Cramer et al., 1985). In addition, SO4
2− has beenshown to affect Mg2+ and PO4
2− uptake (Drost et al.,1997) and disturb nitrogen metabolism (Strogonov,1973), while Cl− can compete with NO3
− and PO43−
for uptake (Curtin et al., 1993). In dogwood seedlings,Mg2+ levels were reduced more by Na2SO4 than byNaCl supporting the above hypothesis.
A decrease in stomatal conductance observed inthe present study in all salt concentrations suggeststhat the plants were responding to salt-induced wa-ter stress by closing the stomata to reduce water loss.A high correlation (r = 0.90) was observed betweenosmotic potential of treatment solutions and stomatalconductance measured after 1 week of treatment. Thisresponse would also reduce water and ion uptake.This could explain, at least in part, the lower levelsof Na+ in shoots of Na2SO4-treated plants comparedwith NaCl-treated plants. The decrease in transpira-tion rates likely contributed to the observed reductionin photosynthesis and growth. Under these conditions,further growth reduction could occur due to a redirec-tion of photosynthates from growth to osmoregulation(Greenway and Munns, 1980).
Changes in chlorophylls a and b during salinityvary depending on plant species, salt treatments andseedling age (Misra et al., 1997). In our study, chloro-phyll concentrations were reduced only by 100 mMNa2SO4 while growth and photosynthesis were re-duced by lower salt concentrations suggesting thatother factors also contributed to the decrease in pho-tosynthetic activity. Salinity likely affects photosyn-thesis by decreasing stomatal conductance. In addi-tion, enzyme activity can be altered by ion excess thuslimiting photosynthesis (Reddy et al., 1997). Growthreduction observed during salinity could lead to ac-cumulation of sugars in the mesophyll cells causingfeedback inhibition of photosynthesis (Munns, 1993;Reddy et al., 1997).
These results confirm our earlier finding (Renaultet al., 1998, 1999) that suggested that red-osier dog-wood tolerates the moderate level of salts present intailings waters. We showed that red-osier dogwoodabsorbed eight times less Na+ than other more sus-ceptible species such as white spruce (Picea glauca).This early–successional species could be planted inthe early stages of reclamation. Once established, theroot system would increase infiltration, and aid in theleaching of salts from the upper region of the soil. Sur-face salinity may also be reduced when the water levelis lowered by evapotranspiration (Tomar, 1997). How-
ever, to determine the suitability of red-osier dogwoodfor land reclamation of areas affected by these salinewaters, the interaction of salinity with other environ-mental variables must be investigated. In addition, tovalidate these results field studies are currently beingundertaken.
In summary, our results suggest that red-osier dog-wood avoided shoot injury by restricting the transportof Na+ to the leaves, when Na+ was present in nutrientsolution in the concentrations not exceeding 50 mM.This mechanism of resistance operates under bothNaCl and Na2SO4 salinity. We also demonstrated thatNa2SO4 and NaCl had different effects on seedlingssuggesting specific anion effects. To confirm the ex-istence of a Na+ regulatory mechanism that allowsmaintenance of low levels of Na+ in leaves, furtherresearch on Na+ localization and movement in rootswill be undertaken.
Acknowledgements
This study was funded by Syncrude Canada Ltd.,Suncor Energy Inc. and the Environmental Scienceand Technology Alliance Canada. The authors wouldlike to thank M. Fung and M. MacKinnon (Syn-crude Canada Ltd.), D. Sheeran and S. Tuttle (SuncorEnergy Inc.), J. Gulley and F. Bishay (Golder Asso-ciates) for their valuable advice. We also thank Dr.Piercey-Normore and the anonymous reviewers forcorrections and helpful comments on the manuscriptand M. Chomokovsky for technical assistance.
References
Alaoui-Sosse B L, Sehmer P, Barnola P and Dizengremel P 1998Effect of NaCl salinity on growth and mineral partitioning inQuercus robur L., a rhythmically growing species. Trees 12,424–430.
Banuls J and Primo-Millo E 1992 Effects of Cl and Na on gas ex-change parameters and water relations of Citrus plants. Physiol.Plant. 86, 115–123.
Bilski J J, Nelson D C and Conlon R L 1988 Response of six wildpotato species to chloride and sulfate salinity. Am. Potato. J. 65,605–612.
Cramer G R, Lauchli A and Polito V S 1985 Displacement of Ca2+from the plasmalemma of root cells. Plant Physiol. 79, 207–211.
Curtin D H, Stepphun and Selles F 1993 Plant responses to sulfateand chloride salinity: Growth and ionic relations. Soil Sci. Soc.Am. J. 57, 1304–1310.
Dirr M A 1974 Tolerance of honeylocust seedlings to soil-appliedsalts. Hortscience 9, 53–54.
Drost D T, MacAdam J W, Dudley L M and Soltani N 1997 Re-sponse of Bean and Broccoli to high-sulfate irrigation water.Hort. Technol. 7, 429–434.

268
Fung L E, Wang S S, Altman A and Huttermann A 1998 Effect ofNaCl on growth, photosynthesis, ion and water relations of fourpoplar genotypes. For. Ecol. Manage. 107, 135–146.
Greenway H and Munns R 1980 Mechanisms of salt tolerance innonhalophytes. Ann. Rev. Plant. Physiol. 31, 149–190.
Grieve A M and Walker R R 1983 Uptake and distribution of chlor-ide, sodium and potassium ions in salt-treated Citrus plants.Aust. J. Agricul. Res. 34, 133–143.
Hardy B B T Limited 1989 Manual of plant species suitability forreclamation in Alberta. 2nd edn. Rep No. RRTAC 89-4 AlbertaLand Conservation and Reclamation Council, Edmonton, Alta.
Khan A H, Ashraf M Y, Naqvi S S M, Khanzada B and Ali M 1995Growth, ion and solute contents of sorghum grown under NaCland Na2SO4 salinity stress. Acta Physiol. Plant. 17, 261–268.
Khan M S A, Hamid A, Salahuddin A B M, Quasem A and Karim MA 1997 Effect of NaCl on growth, photosynthesis and mineralions accumulation of different types of rice (Oryza sativa L.). J.Agron. Crop Sci. 179, 149–161.
Maas E V 1993 Salinity and citriculture. Tree Physiol. 12, 196–216.Marcar N E and Termaat A 1990 Effects of root-zone solutes on
Eucalyptus camaldulensis and Eucalyptus bicostata seedlings:Responses to Na+, Mg2+ and Cl−. Plant Soil 125, 245–254.
Martin P K and Koebner R M D 1995 Na and Cl ions contributesynergistically to salt toxicity in wheat. Biol. Plant. 37, 265–271.
Meiri A, Kamburoff J and Poljakoff-Mayber A 1971 Response ofbean plants to sodium chloride and sodium sulfate salinization.Ann. Bot. 35, 837.
Misra A N, Sahu S M, Misra M, Singh P, Meera I, Das N, Kar M andSahu P 1997 Sodium chloride induced changes in leaf growthand pigment and protein contents in two rice cultivars. Biol.Plant. 39, 257–262.
Munns R 1993 Physiological process limiting plant growth in salinesoils. Some dogma and hypothesis. Plant Cell Environ. 16. 15–24.
Nabil M and Coudret A 1995 Effects of sodium chloride on growth,tissue elasticity and solute adjustment in two Acacia niloticasubspecies. Physiol. Plant. 93, 217–224.
Reddy M P, Rao U S and Iyengar E R R 1997 Carbon metabolismunder salt stress. In Strategies for Improving Salt Tolerance in
Higher Plants. Eds. P K Jaiwal, R P Singh and Gulati, pp 159-181. Sci. Pub, Inc. New Hampshire.
Renault S, Lait C, Zwiazek J J and MacKinnon M D 1998 Effect ofhigh salinity tailings waters produced from gypsum treatment ofoil sands tailings on plants of the boreal forest. Environ. Pollut.102, 177–184.
Renault S, Paton E, Nilsson G, Zwiazek J J and MacKinnon MD 1999 Responses of boreal plants to high salinity oil sandstailings. J. Env. Quality 28, 1957–1962.
Rieger M and P Litvin 1998 Ion selective electrodes for measure-ment of Na and Cl in salinity experiments. J. Plant Nutr. 21,205–215.
Robbins C T and Moen A N 1975 Composition and digestability ofseveral deciduous browses in the northeast. J. Wild. Manag. 39,337–341.
Ruiz D, Martinez V and Cerda A 1997 Citrus response to salinity:Growth and nutrient uptake. Tree Physiol. 17, 141–150.
Scholander P F, Hammel H T, Bradstreef E D and Hemmingsen EA 1965 Sap pressure in vascular plants. Science 148, 339–346.
Sestak K, Catsky J and Jarvis P G 1971 Plant photosyntheticproduction. W Junk . The Hague.
Shannon M C, Grieve C M and Francois L C 1994 Whole-plantresponse to salinity. In Plant Environment Interactions. Ed. R EWilkinson, pp 199–244. M Dekker Inc. New York.
Strogonov B S 1964 Physiological basis of salt tolerance of plants.In Eds. A Poljakoff-Mayber and AM Mayer, Monson S., Jerus-alem.
Strogonov B P 1973 Structure and function of plant cells in salinehabitats. In Ed. B Gollek, John Wiley and Sons, Inc., New York.
Tomar O S 1997 Technologies of afforestation of salt-affected soils.Int. Tree Crops J. 9: 131–158.
Ullah S M, Soja G and Gerzabek M H 1993 Ion uptake, osmore-gulation and plant water relations in faba beans (Vicia faba L.)under salt stress. Bodenkultur 44, 291–301.
Yeo A R, Kramer D, Lauchli A and Gullasch J 1977 Ion distribitionin salt stressed mature Zea mays roots in relation to ultrastructureand retention of sodium. J. Exp. Bot. 28, 17–29.
Section editor: J.M. Cheeseman





























