Embed Size (px)
Citation preview

Effects of Mannitol or Catalase on the Generation ofReactive Oxygen Species Leading to DNA Damage by
Chromium(VI) Reduction with Ascorbate
Tsui-Chun Tsou, Huey-Jen Lai, and Jia-Ling Yang*
Molecular Carcinogenesis Laboratory, Department of Life Sciences, National Tsing Hua University,Hsinchu 300, Taiwan, Republic of China
Received October 6, 1998
Interaction of Cr(VI) and ascorbate in vitro generates Cr(V), Cr(IV), Cr(III), carbon-basedalkyl radicals, COO•-, •OH, and ascorbate radicals and induces DNA interstrand cross-linksat guanines. To determine which specific Cr species and free radicals cause DNA damage, weinvestigated the effects of mannitol and catalase on the formation of reactive intermediates,Cr-DNA associations, DNA polymerase-stop sites, and 8-hydroxydeoxyguanosine (8-OHdG)adducts induced by Cr(VI)/ascorbate in a Hepes buffer. EPR spectra showed that mannitoltrapped reactive Cr(V), forming a stable Cr(V)-diol complex, and inhibited the radicals inducedby Cr(VI)/ascorbate, whereas catalase or heat-denatured catalase enhanced the levels of Cr(V)without altering the radical signals. Mannitol markedly inhibited the retarded gel electro-phoretic mobility of supercoiled plasmids and the formation of DNA polymerase-stop sitesinduced by Cr(VI)/ascorbate, but catalase did not. On the other hand, mannitol reduced only32% of the Cr-DNA adducts induced by Cr(VI)/ascorbate, suggesting that Cr monoadducts(possibly DNA-Cr-mannitol adducts) are the major lesions generated in the Cr(VI)/ascorbate/mannitol/DNA solution. Native catalase but not heat-denatured catalase protected ∼25% ofthe Cr-DNA adducts induced by Cr(VI)/ascorbate, suggesting that hydrogen peroxide may beinvolved. Mannitol could not completely inhibit the formation of 8-OHdG adducts induced byCr(VI)/ascorbate, indicating that this DNA damage may be generated before the action ofmannitol to trap Cr(V) and reactive oxygen species. Alternatively, Cr-peroxide intermediatesmay also lead to 8-OHdG formation to account for the incomplete prevention by mannitol.Catalase or heat-denatured catalase partially protected the formation of 8-OHdG adductsinduced by Cr(VI)/ascorbate, suggesting an effect of proteins. Together, the results from thisstudy suggest that the primary species generated during the reduction of Cr(VI) by ascorbateare hydroxyl radicals and Cr(V) species, responsible for the formation of 8-OHdG and DNAcross-links, respectively.
Introduction
Cr exists in many oxidation states, of which hexavalentCr is the most effective form for inducing genotoxicityand carcinogenicity (1, 2). Cr(VI) compounds significantlyincrease the risk of respiratory tract cancers (3, 4). Cr(VI)also induces chromosomal abnormalities, cell transfor-mations, apoptosis, signal transductions, and gene muta-tions in cultured mammalian cells (5-12). However,Cr(VI) compounds do not directly attack DNA or nucleiin vitro (13-15). The genotoxic and carcinogenic effectsof Cr(VI) compounds are associated with their ability toenter cells rapidly and to be activated through intercel-lular reduction (16, 17). The cellular components involvedin reducing Cr(VI) include ascorbate, glutathione, hy-drogen peroxide, cysteine, DT-diaphorase, cytochromeP450 reductases, and the mitochondrial electron trans-port chain (16-25). Upon activation of Cr(VI), severalreactive species are generated, e.g., Cr(V), Cr(IV), Cr(III),reactive oxygen species (ROS),1 and other free radicals(16-21). Those Cr intermediates, radicals, and ROS cansubsequently attack macromolecules and lead to DNA
damage, e.g., strand breaks, DNA-protein cross-links,DNA-DNA cross-links, Cr-DNA adducts, and basemodifications in cells (10, 21, 26-30).
Ascorbate is the major reductant of Cr(VI) in ratkidneys, liver, and lung ultrafiltrates (22). Pretreatmentof Chinese hamster V79 cells with ascorbate increasesthe number of Cr(VI)-induced DNA-protein cross-links,and decreases the number of alkali-labile sites (21).Ascorbate decreases the clastogenicity of lead chromatepossibly due to eliminated extracellular dissolution andCr(VI) uptake in Chinese hamster ovary cells (31).Interaction of Cr(VI) and ascorbate in vitro generatesCr(V), Cr(IV), Cr(III), carbon-based alkyl radicals, COO•-,•OH, and ascorbate radicals (32-34). The relative amountsof Cr species and radicals that are produced are depend-ent on the ratios of ascorbate to Cr(VI) and the reactionbuffer (32, 33). For example, Cr(V), COO•-, •OH, andcarbon-based alkyl radicals are induced by Cr(VI)/ascor-bate at roughly equal concentrations, whereas only
* To whom correspondence should be addressed. Phone: 886-3-5742756. Fax: 886-3-5721746, E-mail: [email protected].
1 Abbreviations: ROS, reactive oxygen species; Hepes, 4-(2-hydroxy-ethyl)-1-piperazineethanesulfonic acid; 8-OHdG, 8-hydroxydeoxygua-nosine; DMPO, 5,5-dimethyl-1-pyrroline 1-oxide; ICP-MS, inductivelycoupled plasma-mass spectrometer; dG, deoxyguanosine; ECD, elec-trochemical detection.
1002 Chem. Res. Toxicol. 1999, 12, 1002-1009
10.1021/tx9802264 CCC: $18.00 © 1999 American Chemical SocietyPublished on Web 09/18/1999

ascorbate radicals are observed when the amount ofascorbate exceeds that of Cr(VI) (32). Cr(VI)/ascorbatein a phosphate buffer enhances DNA strand breaks (35)and apurinic/apyrimidinic sites (36), whereas in a Hepesbuffer, Cr(VI)/ascorbate induces DNA interstrand cross-links at guanines (37). The levels of Cr(VI)/ascorbate-induced DNA strand breaks (35) and DNA interstrandcross-links (37) decreased when the amount of ascorbateincreased.
The reactive intermediates generated by Cr(VI)/ascor-bate are considered to be the etiology for the inductionof DNA lesions. However, what Cr species and freeradicals cause specific types of DNA damage remain tobe elucidated. In this study, we examined the ability ofCr(VI)/ascorbate to induce 8-OHdG in DNA, an importantoxidative DNA adduct leading to mutagenesis and car-cinogenesis. We also investigated the effects of mannitoland catalase on the generation of reactive intermediates,Cr-DNA adducts, DNA polymerase-stop sites, and 8-O-HdG adducts induced by Cr(VI)/ascorbate to gain moreinsight into the species responsible for the DNA damage.The results have suggested that ROS and Cr(V) may bethe primary species for inducing 8-OHdG adducts andDNA cross-links during the interaction of Cr(VI) andascorbate in a Hepes buffer.
Experimental Procedures
Materials. Potassium dichromate (>99.95% pure), mannitol(D-form), and Hepes were purchased from Merck (Darmstadt,Germany). L-Ascorbate, 5,5-dimethyl-1-pyrroline 1-oxide (DMPO),and calf thymus DNA (activated type XV) were purchased fromSigma (St. Louis, MO). Plasmids pSP189 and pZ189 wereprovided by M. M. Seidman (Otsuka Pharmaceuticals, Rockville,MD). Plasmid pGEM3Zf(+) was purchased from Promega(Madison, WI). Chelex 100 resin (200-400 mesh, sodium form)was purchased from Bio-Rad Laboratories (Richmond, CA).Sephadex G-50 columns and catalase were purchased fromBoehringer Mannheim GmbH (Mannheim, Germany). All of thematerials were freshly prepared in Chelex 100-treated MilliQwater. Hepes was dissolved in MilliQ water at a concentrationof 1 M, and then passed through Chelex 100 resin twice.
EPR Spectroscopy. Mannitol (50 mM) or catalase (800units/mL) was delivered to an aqueous solution containing 100mM DMPO, 1 mM K2Cr2O7 [equivalent to 2 mM Cr(VI)], and 2mM ascorbate. The reaction mixtures were kept at roomtemperature for 4 min before the EPR spectra were recordedusing a Bruker ER (type EMX 10/12) spectrometer with a 100kHz field modulation, a 20 mW microwave power, and a 100 Gscan width. The g values were calculated from calibrationagainst 1,1-diphenyl-2-picrylhydrazyl (g ) 2.0037) (38). All ofthe materials for the EPR analysis were freshly prepared.
Agarose Gel Electrophoresis. Supercoiled plasmidspGEM3Zf(+) (0.4-2.8 µg) were mixed with K2Cr2O7 and/orascorbate in 10 mM Hepes (pH 7.0) in a total volume of 25 µL.The reaction mixture was incubated at 37 °C for 30 min. Inexperiments aimed at determining the effect of ROS scavengers,either mannitol or catalase was delivered to the DNA solutionbefore adding K2Cr2O7 and/or ascorbate. At the end of incuba-tion, DNA samples were analyzed using electrophoresis in 0.8%agarose gels containing ethidium bromide (0.5 µg/mL).
Polymerase-Stop Assay of Cr(VI)/Ascorbate-TreatedDNA. Plasmids pZ189 were treated with EcoRI and purifiedas described previously (39). The DNA sample (1.7 µg) was thentreated with Cr(VI)/ascorbate in the presence of either mannitol,catalase, or heat-denatured catalase in a total volume of 15 µLas described above. At the end of the reaction, the DNA sampleswere drop dialyzed against a 10 mM Hepes buffer for 4 h, andused as templates for the polymerase-stop assay. The in vitroDNA polymerase-stop assay was performed using the template,
[γ-32P]ATP (specific activity of 5000 Ci/mmol), end-labeledprimer 5′-ACGGGGTCTGACG, Sequenase, and four deoxyri-bonucleotides as described previously (39).
Determination of the Amount of Cr Bound to DNA.Immediately after treatment, plasmids pSP189 were filteredthrough Sephadex G-50 columns at 2700 rpm for 4 min. Theamount of Cr bound to the plasmid was determined using anICP-MS (SCIEX ELAN 5000, Perkin-Elmer, Norwalk, CT). TheICP-MS conditions were as follows: power of 5000 W, plasmaflow rate of 15 L/min, auxiliary flow rate of 0.8 L/min, andsample flow rate of 0.8 L/min. One set of control experimentswas conducted in which DNA samples were excluded. Thenumber of Cr molecules bound per 1000 nucleotides wascalculated as the data obtained in the reaction mixturescontaining DNA samples subtracted from those obtained inmixtures without DNA.
Determination of the Amounts of dG and 8-OHdG. Calfthymus DNA (100 µg) was treated with Cr(VI)/ascorbate in thepresence of either mannitol, catalase, or heat-denatured catalasein 10 mM Hepes (pH 7.0) in a total volume of 450 µL. Thereaction was carried out at 37 °C for 30 min and terminatedusing ethanol precipitation. The DNA sample was washed oncewith 70% ethanol, dried, dissolved in 10 mM Tris-HCl (pH 7.4),and heated at 100 °C for 3 min. DNA was digested to deoxyri-bonucleosides, and the amounts of dG and 8-OHdG wereanalyzed using HPLC with UV absorbance detection and ECD(BAS amperometric electrochemical detector, West Lafayette,IN) as previously described (40). The protection of Cr(VI)/ascorbate-induced 8-OHdG formation by mannitol, catalase, orheat-denatured catalase was determined with the equation 1- [(Xm+a - Xc)/(Xm - Xc)], where Xm is the amount of 8-OHdGadducts remaining after treatment with Cr(VI)/ascorbate, Xm+a
is the amount of 8-OHdG adducts remaining after treatmentwith Cr(VI)/ascorbate and a ROS modulator, and Xc is theamount of 8-OHdG adducts remaining in the untreated control.
Results
Effects of Mannitol and Catalase on the Cr(VI)/Ascorbate-Induced EPR Spectrum. EPR spectroscopywas used to investigate reactive intermediates such asCr(V) and radicals generated in an aqueous solutioncontaining Cr(VI)/ascorbate in the presence of eithermannitol, catalase, or heat-denatured catalase in 10 mMHepes buffer (pH 7.0). The spin trap DMPO was addedat a concentration of 100 mM to each reaction mixture.The EPR spectrum obtained upon reaction of Cr(VI) andascorbate at a 1:1 ratio in a solution containing DMPOshowed the presence of Cr(V), DMPO-•OH, DMPO-COO•, DMPO-•C, and DMPO-R• radical adducts (Figure1A) which confirmed previous reports (32, 34). The signalintensities of Cr(V) and DMPO-radical adducts de-creased slowly when the incubation time increased; e.g.,the intensities observed 30 min after the reaction wereroughly 60% of those obtained at 4 min (data not shown).A similar spectrum for Figure 1A was obtained when theCr(VI):ascorbate ratio increased to 2:1, whereas only theascorbate radical anion (g ) 2.006, AH ) 1.82 G) wasobserved at a Cr(VI):ascorbate ratio of 1:2 (data notshown). DMPO did not affect the signal intensity of theCr(V) or the ascorbate radical anion (data not shown).The signal intensity of Cr(V) increased 3-fold, and theamounts of DMPO-radical adducts were not significantlyaltered by adding native or heat-denatured catalase (800units/mL) to the 2 mM Cr(VI)/2 mM ascorbate solution(Figure 1B,C). By contrast, mannitol (50 mM) markedlyincreased (∼200-fold) the magnitude of an altered Cr(V)signal, and inhibited the generation of DMPO-radicaladducts in the Cr(VI)/ascorbate solution (Figure 1D). This
Prevention of Cr(VI)/Ascorbate-Induced DNA Damage Chem. Res. Toxicol., Vol. 12, No. 10, 1999 1003
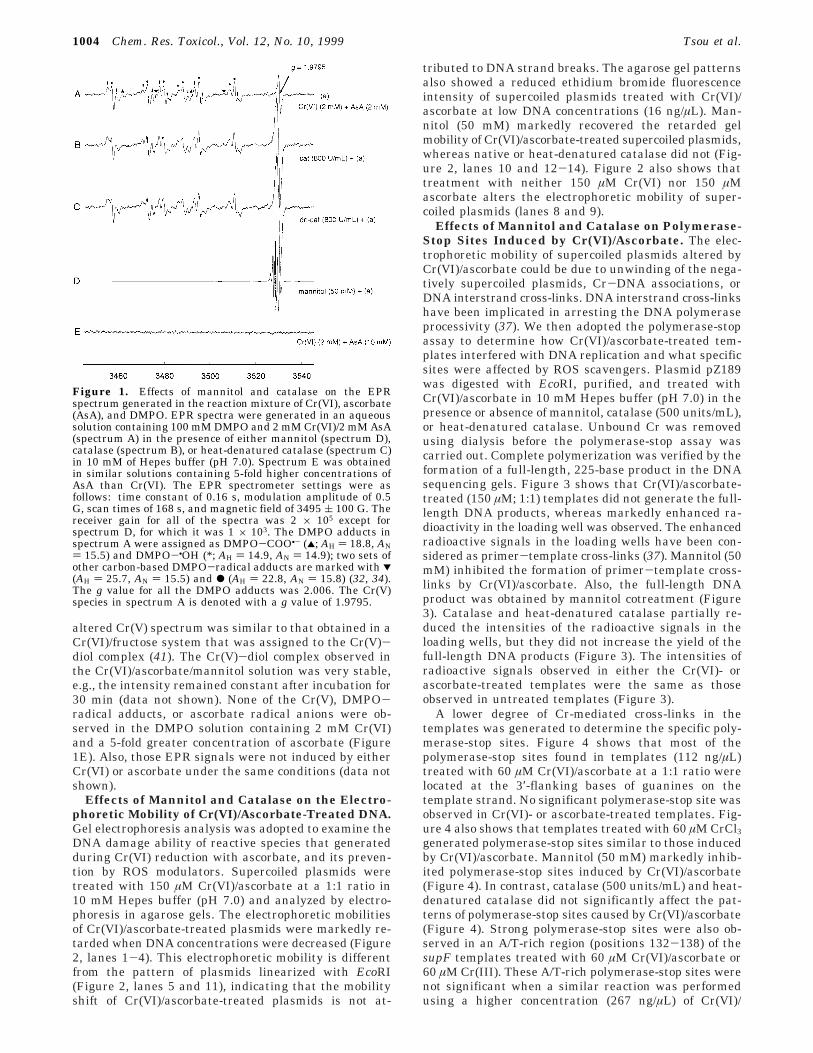
altered Cr(V) spectrum was similar to that obtained in aCr(VI)/fructose system that was assigned to the Cr(V)-diol complex (41). The Cr(V)-diol complex observed inthe Cr(VI)/ascorbate/mannitol solution was very stable,e.g., the intensity remained constant after incubation for30 min (data not shown). None of the Cr(V), DMPO-radical adducts, or ascorbate radical anions were ob-served in the DMPO solution containing 2 mM Cr(VI)and a 5-fold greater concentration of ascorbate (Figure1E). Also, those EPR signals were not induced by eitherCr(VI) or ascorbate under the same conditions (data notshown).
Effects of Mannitol and Catalase on the Electro-phoretic Mobility of Cr(VI)/Ascorbate-Treated DNA.Gel electrophoresis analysis was adopted to examine theDNA damage ability of reactive species that generatedduring Cr(VI) reduction with ascorbate, and its preven-tion by ROS modulators. Supercoiled plasmids weretreated with 150 µM Cr(VI)/ascorbate at a 1:1 ratio in10 mM Hepes buffer (pH 7.0) and analyzed by electro-phoresis in agarose gels. The electrophoretic mobilitiesof Cr(VI)/ascorbate-treated plasmids were markedly re-tarded when DNA concentrations were decreased (Figure2, lanes 1-4). This electrophoretic mobility is differentfrom the pattern of plasmids linearized with EcoRI(Figure 2, lanes 5 and 11), indicating that the mobilityshift of Cr(VI)/ascorbate-treated plasmids is not at-
tributed to DNA strand breaks. The agarose gel patternsalso showed a reduced ethidium bromide fluorescenceintensity of supercoiled plasmids treated with Cr(VI)/ascorbate at low DNA concentrations (16 ng/µL). Man-nitol (50 mM) markedly recovered the retarded gelmobility of Cr(VI)/ascorbate-treated supercoiled plasmids,whereas native or heat-denatured catalase did not (Fig-ure 2, lanes 10 and 12-14). Figure 2 also shows thattreatment with neither 150 µM Cr(VI) nor 150 µMascorbate alters the electrophoretic mobility of super-coiled plasmids (lanes 8 and 9).
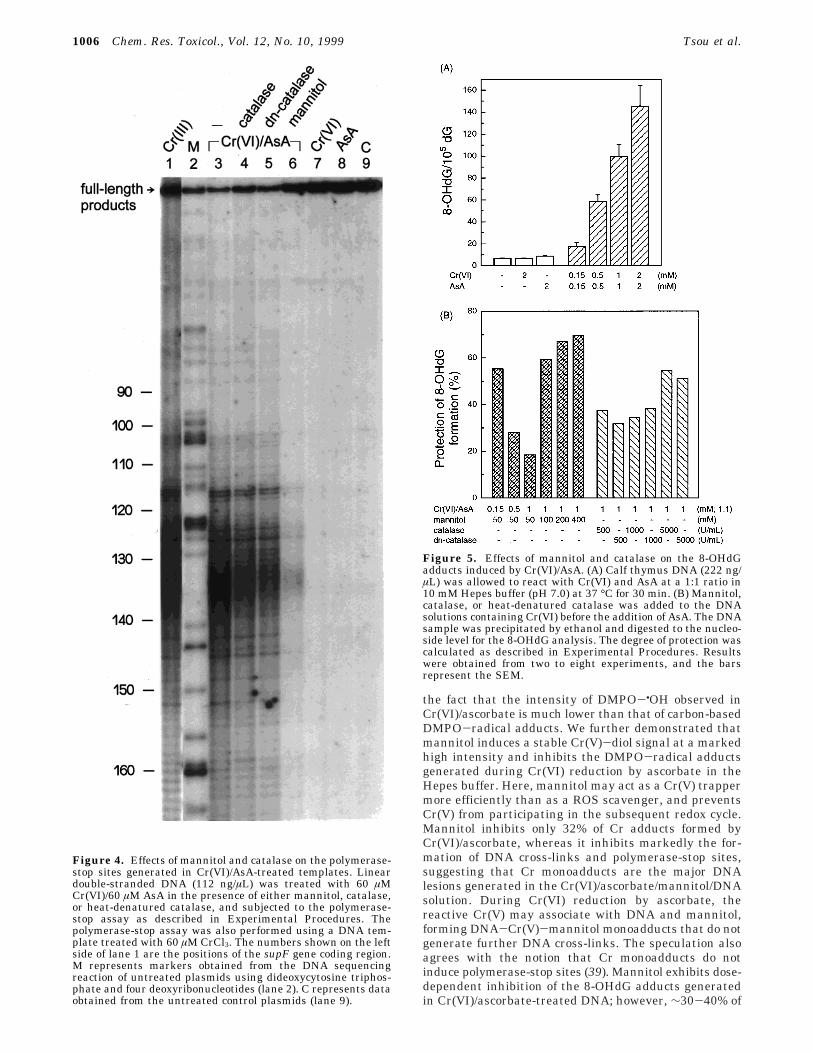
Effects of Mannitol and Catalase on Polymerase-Stop Sites Induced by Cr(VI)/Ascorbate. The elec-trophoretic mobility of supercoiled plasmids altered byCr(VI)/ascorbate could be due to unwinding of the nega-tively supercoiled plasmids, Cr-DNA associations, orDNA interstrand cross-links. DNA interstrand cross-linkshave been implicated in arresting the DNA polymeraseprocessivity (37). We then adopted the polymerase-stopassay to determine how Cr(VI)/ascorbate-treated tem-plates interfered with DNA replication and what specificsites were affected by ROS scavengers. Plasmid pZ189was digested with EcoRI, purified, and treated withCr(VI)/ascorbate in 10 mM Hepes buffer (pH 7.0) in thepresence or absence of mannitol, catalase (500 units/mL),or heat-denatured catalase. Unbound Cr was removedusing dialysis before the polymerase-stop assay wascarried out. Complete polymerization was verified by theformation of a full-length, 225-base product in the DNAsequencing gels. Figure 3 shows that Cr(VI)/ascorbate-treated (150 µM; 1:1) templates did not generate the full-length DNA products, whereas markedly enhanced ra-dioactivity in the loading well was observed. The enhancedradioactive signals in the loading wells have been con-sidered as primer-template cross-links (37). Mannitol (50mM) inhibited the formation of primer-template cross-links by Cr(VI)/ascorbate. Also, the full-length DNAproduct was obtained by mannitol cotreatment (Figure3). Catalase and heat-denatured catalase partially re-duced the intensities of the radioactive signals in theloading wells, but they did not increase the yield of thefull-length DNA products (Figure 3). The intensities ofradioactive signals observed in either the Cr(VI)- orascorbate-treated templates were the same as thoseobserved in untreated templates (Figure 3).
A lower degree of Cr-mediated cross-links in thetemplates was generated to determine the specific poly-merase-stop sites. Figure 4 shows that most of thepolymerase-stop sites found in templates (112 ng/µL)treated with 60 µM Cr(VI)/ascorbate at a 1:1 ratio werelocated at the 3′-flanking bases of guanines on thetemplate strand. No significant polymerase-stop site wasobserved in Cr(VI)- or ascorbate-treated templates. Fig-ure 4 also shows that templates treated with 60 µM CrCl3
generated polymerase-stop sites similar to those inducedby Cr(VI)/ascorbate. Mannitol (50 mM) markedly inhib-ited polymerase-stop sites induced by Cr(VI)/ascorbate(Figure 4). In contrast, catalase (500 units/mL) and heat-denatured catalase did not significantly affect the pat-terns of polymerase-stop sites caused by Cr(VI)/ascorbate(Figure 4). Strong polymerase-stop sites were also ob-served in an A/T-rich region (positions 132-138) of thesupF templates treated with 60 µM Cr(VI)/ascorbate or60 µM Cr(III). These A/T-rich polymerase-stop sites werenot significant when a similar reaction was performedusing a higher concentration (267 ng/µL) of Cr(VI)/
Figure 1. Effects of mannitol and catalase on the EPRspectrum generated in the reaction mixture of Cr(VI), ascorbate(AsA), and DMPO. EPR spectra were generated in an aqueoussolution containing 100 mM DMPO and 2 mM Cr(VI)/2 mM AsA(spectrum A) in the presence of either mannitol (spectrum D),catalase (spectrum B), or heat-denatured catalase (spectrum C)in 10 mM of Hepes buffer (pH 7.0). Spectrum E was obtainedin similar solutions containing 5-fold higher concentrations ofAsA than Cr(VI). The EPR spectrometer settings were asfollows: time constant of 0.16 s, modulation amplitude of 0.5G, scan times of 168 s, and magnetic field of 3495 ( 100 G. Thereceiver gain for all of the spectra was 2 × 105 except forspectrum D, for which it was 1 × 103. The DMPO adducts inspectrum A were assigned as DMPO-COO•- (2; AH ) 18.8, AN) 15.5) and DMPO-•OH (*; AH ) 14.9, AN ) 14.9); two sets ofother carbon-based DMPO-radical adducts are marked with 1(AH ) 25.7, AN ) 15.5) and b (AH ) 22.8, AN ) 15.8) (32, 34).The g value for all the DMPO adducts was 2.006. The Cr(V)species in spectrum A is denoted with a g value of 1.9795.
1004 Chem. Res. Toxicol., Vol. 12, No. 10, 1999 Tsou et al.

ascorbate-treated (data not shown) or Cr(III)-treatedtemplates (39), although the majority of polymerase-stopsites still occurred at guanine residues in DNA templates.
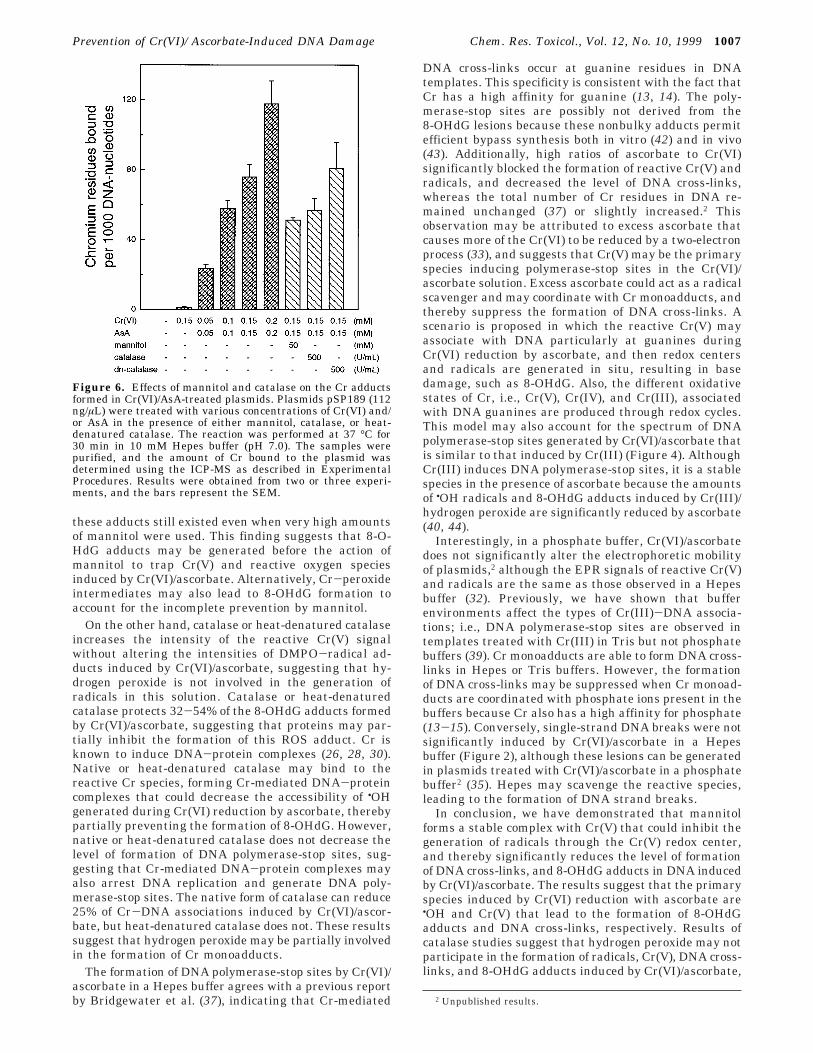
Effects of Mannitol and Catalase on the Forma-tion of 8-OHdG Adducts Induced by Cr(VI)/Ascor-bate. 8-OHdG is an important oxidative DNA adductleading to mutagenesis and carcinogenesis. The abilityof Cr(VI)/ascorbate to induce 8-OHdG formation in calfthymus DNA in 10 mM Hepes buffer (pH 7.0) wasdetermined using HPLC/UV/ECD. Figure 5A shows thatthe amount of 8-OHdG adducts in the DNA markedlyincreased when an equal ratio of the Cr(VI) and ascorbateconcentrations was increased (150 µM to 2 mM, in a1:1 ratio). The amounts of 8-OHdG adducts generatedin Cr(VI)/ascorbate-treated DNA at 150 µM and 2 mM(1:1 ratio) were 18/105 dG bases and 146/105 dG bases,respectively. Control experiments with 2 mM Cr(VI) orascorbate alone did not enhance 8-OHdG formation inthe DNA at the background level (6.5/105 dG bases).
Coadministrating 50 mM mannitol protected approxi-mately 55 and 28% of the 8-OHdG adducts induced by
150 µM Cr(VI) and 500 µM ascorbate (1:1 ratio), respec-tively (Figure 5B). Mannitol (50-200 mM) also exhibiteda dose-dependent protection on the 8-OHdG adductsinduced by 1 mM Cr(VI)/ascorbate (Figure 5B). Similarly,catalase (500-5000 units/mL) and heat-denatured cata-lase protected 32-54% of the 8-OHdG adducts inducedby 1 mM Cr(VI)/ascorbate (Figure 5B), suggesting thatthe prevention of 8-OHdG formation may be due to theproteins’ effect but not due to the catalase enzymaticactivity.
Effects of Mannitol and Catalase on the Forma-tion of Cr Adducts Induced by Cr(VI)/Ascorbate.The effects of mannitol and catalase on the formation ofCr adducts induced by Cr(VI)/ascorbate were determinedusing the ICP-MS. Plasmids pSP189 were treated withvarious concentrations of Cr(VI) and/or ascorbate in 10mM Hepes buffer (pH 7.0). Following removal of unboundCr ions by Sephadex G-50 gel filtration, the amounts ofCr bound to plasmids were assayed by the ICP-MS, andcalculated as described in Experimental Procedures. Asshown in Figure 6, the amount of Cr adducts in plasmidincreased linearly when Cr(VI)/ascorbate concentrationsat a 1:1 ratio were increased. Cr adducts formed by 150µM Cr(VI)/ascorbate were decreased approximately 32%by adding 50 mM mannitol. Catalase (500 units/ml)reduced 25% of the Cr adducts formed by 150 µM Cr(VI)/ascorbate, whereas heat-denatured catalase did not affectthe amounts of Cr(VI)/ascorbate-induced Cr adducts(Figure 6), suggesting that hydrogen peroxide may begenerated in the reaction mixture and contributes to theformation of Cr adducts.
Discussion
EPR spectroscopic results have confirmed that Cr(VI)reduction by ascorbate at stoichiometric ratios can inducereactive Cr(V) and radicals, including carbon-based,COO•-, •OH, and ascorbate radicals (32, 34). In the studypresented here, we also demonstrated that interactionof Cr(VI) with ascorbate generates ROS-related 8-OHdGadducts in DNA. The highly reactive •OH may be theprimary species produced in the Cr(VI)/ascorbate solutionand then attacks ascorbate to generate ascorbate-derivedCOO•- and carbon-based radicals. This could account for
Figure 2. Effects of mannitol and catalase on the electrophoretic mobility of Cr(VI)/AsA-treated supercoiled plasmids. Supercoiledplasmids pGEM3Zf(+) were allowed to react with 150 µM Cr(VI)/150 µM AsA and analyzed as described in Experimental Procedures(lanes 1-4 and 10). The concentration of plasmids in all of the reaction samples was 16 ng/µL, except for the concentrations of thesamples in lanes 2-4 which were 48, 80, and 112 ng/µL, respectively. The amount of plasmids per lane was 400 ng. Lanes 6 and 7contained untreated plasmids. Lanes 8 and 9 contained plasmids treated with Cr(VI) (150 µM) and AsA (150 µM), respectively.Lanes 5 and 11 contained plasmids linearized using EcoRI (one cut). Lanes 12-14 contained plasmids treated with 150 µM Cr(VI)/150 µM AsA in the presence of either mannitol (50 mM), catalase (40 units/mL), or heat-denatured catalase.
Figure 3. Effects of mannitol and catalase on the primer-template cross-link formation and inhibition of DNA synthesisby Cr(VI)/AsA. Linear double-stranded DNA (112 ng/µL) wastreated with 150 µM Cr(VI)/150 µM AsA in the presence of eithermannitol, catalase, or heat-denatured catalase, and subjectedto the polymerase-stop assay as described in ExperimentalProcedures. Complete DNA synthesis was verified by theobservation of a full-length, 225-base product in DNA sequenc-ing gels. The enhanced radioactive signals in the loading wellsobtained in Cr(VI)/AsA-treated DNA represent primer-templatecross-links. C represents data obtained from the untreatedcontrol plasmids.
Prevention of Cr(VI)/Ascorbate-Induced DNA Damage Chem. Res. Toxicol., Vol. 12, No. 10, 1999 1005
the fact that the intensity of DMPO-•OH observed inCr(VI)/ascorbate is much lower than that of carbon-basedDMPO-radical adducts. We further demonstrated thatmannitol induces a stable Cr(V)-diol signal at a markedhigh intensity and inhibits the DMPO-radical adductsgenerated during Cr(VI) reduction by ascorbate in theHepes buffer. Here, mannitol may act as a Cr(V) trappermore efficiently than as a ROS scavenger, and preventsCr(V) from participating in the subsequent redox cycle.Mannitol inhibits only 32% of Cr adducts formed byCr(VI)/ascorbate, whereas it inhibits markedly the for-mation of DNA cross-links and polymerase-stop sites,suggesting that Cr monoadducts are the major DNAlesions generated in the Cr(VI)/ascorbate/mannitol/DNAsolution. During Cr(VI) reduction by ascorbate, thereactive Cr(V) may associate with DNA and mannitol,forming DNA-Cr(V)-mannitol monoadducts that do notgenerate further DNA cross-links. The speculation alsoagrees with the notion that Cr monoadducts do notinduce polymerase-stop sites (39). Mannitol exhibits dose-dependent inhibition of the 8-OHdG adducts generatedin Cr(VI)/ascorbate-treated DNA; however, ∼30-40% of
Figure 4. Effects of mannitol and catalase on the polymerase-stop sites generated in Cr(VI)/AsA-treated templates. Lineardouble-stranded DNA (112 ng/µL) was treated with 60 µMCr(VI)/60 µM AsA in the presence of either mannitol, catalase,or heat-denatured catalase, and subjected to the polymerase-stop assay as described in Experimental Procedures. Thepolymerase-stop assay was also performed using a DNA tem-plate treated with 60 µM CrCl3. The numbers shown on the leftside of lane 1 are the positions of the supF gene coding region.M represents markers obtained from the DNA sequencingreaction of untreated plasmids using dideoxycytosine triphos-phate and four deoxyribonucleotides (lane 2). C represents dataobtained from the untreated control plasmids (lane 9).
Figure 5. Effects of mannitol and catalase on the 8-OHdGadducts induced by Cr(VI)/AsA. (A) Calf thymus DNA (222 ng/µL) was allowed to react with Cr(VI) and AsA at a 1:1 ratio in10 mM Hepes buffer (pH 7.0) at 37 °C for 30 min. (B) Mannitol,catalase, or heat-denatured catalase was added to the DNAsolutions containing Cr(VI) before the addition of AsA. The DNAsample was precipitated by ethanol and digested to the nucleo-side level for the 8-OHdG analysis. The degree of protection wascalculated as described in Experimental Procedures. Resultswere obtained from two to eight experiments, and the barsrepresent the SEM.
1006 Chem. Res. Toxicol., Vol. 12, No. 10, 1999 Tsou et al.
these adducts still existed even when very high amountsof mannitol were used. This finding suggests that 8-O-HdG adducts may be generated before the action ofmannitol to trap Cr(V) and reactive oxygen speciesinduced by Cr(VI)/ascorbate. Alternatively, Cr-peroxideintermediates may also lead to 8-OHdG formation toaccount for the incomplete prevention by mannitol.
On the other hand, catalase or heat-denatured catalaseincreases the intensity of the reactive Cr(V) signalwithout altering the intensities of DMPO-radical ad-ducts induced by Cr(VI)/ascorbate, suggesting that hy-drogen peroxide is not involved in the generation ofradicals in this solution. Catalase or heat-denaturedcatalase protects 32-54% of the 8-OHdG adducts formedby Cr(VI)/ascorbate, suggesting that proteins may par-tially inhibit the formation of this ROS adduct. Cr isknown to induce DNA-protein complexes (26, 28, 30).Native or heat-denatured catalase may bind to thereactive Cr species, forming Cr-mediated DNA-proteincomplexes that could decrease the accessibility of •OHgenerated during Cr(VI) reduction by ascorbate, therebypartially preventing the formation of 8-OHdG. However,native or heat-denatured catalase does not decrease thelevel of formation of DNA polymerase-stop sites, sug-gesting that Cr-mediated DNA-protein complexes mayalso arrest DNA replication and generate DNA poly-merase-stop sites. The native form of catalase can reduce25% of Cr-DNA associations induced by Cr(VI)/ascor-bate, but heat-denatured catalase does not. These resultssuggest that hydrogen peroxide may be partially involvedin the formation of Cr monoadducts.
The formation of DNA polymerase-stop sites by Cr(VI)/ascorbate in a Hepes buffer agrees with a previous reportby Bridgewater et al. (37), indicating that Cr-mediated
DNA cross-links occur at guanine residues in DNAtemplates. This specificity is consistent with the fact thatCr has a high affinity for guanine (13, 14). The poly-merase-stop sites are possibly not derived from the8-OHdG lesions because these nonbulky adducts permitefficient bypass synthesis both in vitro (42) and in vivo(43). Additionally, high ratios of ascorbate to Cr(VI)significantly blocked the formation of reactive Cr(V) andradicals, and decreased the level of DNA cross-links,whereas the total number of Cr residues in DNA re-mained unchanged (37) or slightly increased.2 Thisobservation may be attributed to excess ascorbate thatcauses more of the Cr(VI) to be reduced by a two-electronprocess (33), and suggests that Cr(V) may be the primaryspecies inducing polymerase-stop sites in the Cr(VI)/ascorbate solution. Excess ascorbate could act as a radicalscavenger and may coordinate with Cr monoadducts, andthereby suppress the formation of DNA cross-links. Ascenario is proposed in which the reactive Cr(V) mayassociate with DNA particularly at guanines duringCr(VI) reduction by ascorbate, and then redox centersand radicals are generated in situ, resulting in basedamage, such as 8-OHdG. Also, the different oxidativestates of Cr, i.e., Cr(V), Cr(IV), and Cr(III), associatedwith DNA guanines are produced through redox cycles.This model may also account for the spectrum of DNApolymerase-stop sites generated by Cr(VI)/ascorbate thatis similar to that induced by Cr(III) (Figure 4). AlthoughCr(III) induces DNA polymerase-stop sites, it is a stablespecies in the presence of ascorbate because the amountsof •OH radicals and 8-OHdG adducts induced by Cr(III)/hydrogen peroxide are significantly reduced by ascorbate(40, 44).
Interestingly, in a phosphate buffer, Cr(VI)/ascorbatedoes not significantly alter the electrophoretic mobilityof plasmids,2 although the EPR signals of reactive Cr(V)and radicals are the same as those observed in a Hepesbuffer (32). Previously, we have shown that bufferenvironments affect the types of Cr(III)-DNA associa-tions; i.e., DNA polymerase-stop sites are observed intemplates treated with Cr(III) in Tris but not phosphatebuffers (39). Cr monoadducts are able to form DNA cross-links in Hepes or Tris buffers. However, the formationof DNA cross-links may be suppressed when Cr monoad-ducts are coordinated with phosphate ions present in thebuffers because Cr also has a high affinity for phosphate(13-15). Conversely, single-strand DNA breaks were notsignificantly induced by Cr(VI)/ascorbate in a Hepesbuffer (Figure 2), although these lesions can be generatedin plasmids treated with Cr(VI)/ascorbate in a phosphatebuffer2 (35). Hepes may scavenge the reactive species,leading to the formation of DNA strand breaks.
In conclusion, we have demonstrated that mannitolforms a stable complex with Cr(V) that could inhibit thegeneration of radicals through the Cr(V) redox center,and thereby significantly reduces the level of formationof DNA cross-links, and 8-OHdG adducts in DNA inducedby Cr(VI)/ascorbate. The results suggest that the primaryspecies induced by Cr(VI) reduction with ascorbate are•OH and Cr(V) that lead to the formation of 8-OHdGadducts and DNA cross-links, respectively. Results ofcatalase studies suggest that hydrogen peroxide may notparticipate in the formation of radicals, Cr(V), DNA cross-links, and 8-OHdG adducts induced by Cr(VI)/ascorbate,
2 Unpublished results.
Figure 6. Effects of mannitol and catalase on the Cr adductsformed in Cr(VI)/AsA-treated plasmids. Plasmids pSP189 (112ng/µL) were treated with various concentrations of Cr(VI) and/or AsA in the presence of either mannitol, catalase, or heat-denatured catalase. The reaction was performed at 37 °C for30 min in 10 mM Hepes buffer (pH 7.0). The samples werepurified, and the amount of Cr bound to the plasmid wasdetermined using the ICP-MS as described in ExperimentalProcedures. Results were obtained from two or three experi-ments, and the bars represent the SEM.
Prevention of Cr(VI)/Ascorbate-Induced DNA Damage Chem. Res. Toxicol., Vol. 12, No. 10, 1999 1007

whereas it may be involved in the formation of Crmonoadducts. Native or heat-denatured catalase partiallyprevents the formation of 8-OHdG adducts, but they donot decrease the levels of radicals generated by Cr(VI)/ascorbate, suggesting that Cr-mediated DNA-proteincomplexes are less accessible for ROS. However, theseCr-mediated DNA-protein complexes may generate poly-merase-stop sites. Together, this study suggests that8-OHdG adducts and DNA cross-links could be the majoradducts induced by Cr(VI)/ascorbate, leading to mu-tagenesis and carcinogenesis.
Acknowledgment. This work was supported byGrant NSC86-2113-M007-006 from the National ScienceCouncil, Republic of China.
References
(1) De Flora, S., Bagnasco, M., Serra, D., and Zanacchi, P. (1990)Genotoxicity of chromium compounds. A review. Mutat. Res. 238,99-172.
(2) Snow, E. T. (1992) Metal carcinogenesis: mechanistic implica-tions. Pharmacol. Ther. 53, 31-65.
(3) Langard, S., and Norseth, T. (1986) Chromium. In Handbook onthe Toxicology of Metals (Friberg, L., Nordberg, G. F., and Vouk,V., Eds.) 2nd ed., pp 185-210, Elsevier Science, Amsterdam.
(4) International Agency for Research on Cancer (1990) Chromium,nickel, and wedding. In IARC Monographs on the evaluation ofthe carcinogenic risks of chemicals to humans, Vol. 19, Interna-tional Agency for Cancer Research, Lyon, France.
(5) Sen, P., and Costa, M. (1986) Incidence and localization of sisterchromatid exchanges induced by nickel and chromium com-pounds. Carcinogenesis 7, 1527-1533.
(6) Sen, P., Conway, K., and Costa, M. (1987) Comparison of thelocalization of chromosome damage induced by calcium chromateand nickel compounds. Cancer Res. 47, 2142-2147.
(7) Biedermann, K. A., and Landolph, J. R. (1990) Role of valencestate and solubility of chromium compounds on induction ofcytotoxicity, mutagenesis, and anchorage independence in diploidhuman fibroblasts. Cancer Res. 50, 7835-7842.
(8) Yang, J.-L., Hsieh, Y.-C., Wu, C.-W., and Lee, T.-C. (1992)Mutational specificity of chromium(VI) compounds in the hprtlocus of Chinese hamster ovary-K1 cells. Carcinogenesis 13,2053-2057.
(9) Chen, J., and Thilly, W. G. (1994) Mutational spectrum ofchromium(VI) in human cells. Mutat. Res. 323, 21-27.
(10) Manning, F. C. R., Blankenship, L. J., Wise, J. P., Xu, J.,Bridgewater, L. C., and Patierno, S. R. (1994) Induction ofinternucleosomal DNA fragmentation by carcinogenic chromate:relationship to DNA damage, genotoxicity, and inhibition ofmacromolecular synthesis. Environ. Health Perspect. 102 (Suppl.3), 159-167.
(11) Ye, J., Zhang, X., Young, H. A., Mao, Y., and Shi, X. (1995)Chromium(VI)-induced nuclear factor-κB activation in intact cellsvia free radical reactions. Carcinogenesis 16, 2401-2405.
(12) Kim, G., and Yurkow, E. J. (1996) Chromium induces a persistentactivation of mitogen-activated protein kinases by a redox-sensitive mechanism in H4 rat hepatoma cells. Cancer Res. 56,2045-2051.
(13) Tsapakos, M. J., and Wetterhahn, K. E. (1983) The interactionof chromium with nucleic acids. Chem.-Biol. Interact. 46, 265-277.
(14) Koster, A., and Beyersmann, D. (1985) Chromium binding by calfthymus nuclei and effects on chromatin. Toxicol. Environ. Chem.10, 307-313.
(15) Hneihen, A. S., Standeven, A. M., and Wetterhahn, K. E. (1993)Differential binding of chromium(VI) and chromium(III) com-plexes to salmon sperm nuclei and nuclear DNA and isolated calfthymus DNA. Carcinogenesis 14, 1795-1803.
(16) Wetterhahn, K. E., Hamilton, J. W., Aiyar, J., Borges, K. M., andFloyd, R. (1989) Mechanism of chromium(VI) carcinogenesis:reactive intermediates and effect on gene expression. Biol. TraceElem. Res. 21, 405-411.
(17) Aiyar, J., Berkovits, H. J., Floyd, R. A., and Wetterhahn, K. E.(1991) Reaction of chromium(VI) with glutathione or with hydro-gen peroxide: identification of reactive intermediates and theirrole in chromium(VI)-induced DNA damage. Environ. HealthPerspect. 92, 53-62.
(18) Kawanishi, S., Inoue, S., and Sano, S. (1986) Mechanism of DNAcleavage induced by sodium chromium(VI) in the presence ofhydrogen peroxide. J. Biol. Chem. 261, 5952-5958.
(19) Kortenkamp, A., Oetken, G., and Beyersmann, D. (1990) The DNAcleavage induced by a chromium(V) complex and by chromate andglutathione is mediated by activated oxygen species. Mutat. Res.232, 155-161.
(20) Shi, X., and Dalal, N. S. (1994) Generation of hydroxyl radical bychromate in biologically relevant systems: role of Cr(V) complexesversus tetraperoxochromate(V). Environ. Health Perspect. 102(Suppl. 3), 231-236.
(21) Sugiyama, M., Tsuzuki, K., and Ogura, R. (1991) Effect of ascorbicacid on DNA damage, cytotoxicity, glutathione reductase, andformation of paramagnetic chromium in Chinese hamster V-79cells treated with sodium chromate(VI). J. Biol. Chem. 266, 3383-3386.
(22) Standeven, A. M., and Wetterhahn, K. E. (1992) Ascorbate is theprincipal reductant of chromium(VI) in rat lung ultrafiltrates andcytosols, and mediates chromium-DNA binding in vitro. Carcino-genesis 13, 1319-1324.
(23) De Flora, S., Morelli, A., Basso, C., Romano, M., Serra, D., andDe Flora, A. (1985) Prominent role of DT-diaphorase as a cellularmechanism reducing chromium(VI) and reverting its mutagenic-ity. Cancer Res. 45, 3188-3196.
(24) Ryberg, D., and Alexander, J. (1990) Mechanisms of chromiumtoxicity in mitochondria. Chem.-Biol. Interact. 75, 141-151.
(25) Mikalsen, A., Alexander, J., Wallin, H., Ingelman-Sundberg, M.,and Andersen, R. A. (1991) Reductive metabolism and proteinbinding of chromium(VI) by P450 protein enzymes. Carcinogenesis12, 825-831.
(26) Wedrychowski, A., Ward, W. S., Schmidt, W. N., and Hnilica, L.S. (1985) Chromium-induced cross-linking of nuclear proteins andDNA. J. Biol. Chem. 260, 7150-7155.
(27) Sugiyama, M., Wang, X. W., and Costa, M. (1986) Comparison ofDNA lesions and cytotoxicity induced by calcium chromate inhuman, mouse, and hamster cell lines. Cancer Res. 46, 4547-4551.
(28) Salnikow, K., Zhitkovich, A., and Costa, M. (1992) Analysis ofthe binding sites of chromium to DNA and protein in vitro andin intact cells. Carcinogenesis 13, 2341-2346.
(29) Misra, M., Alcedo, J. A., and Wetterhahn, K. E. (1994) Twopathways for chromium(VI)-induced DNA damage in 14 day chickembryos: Cr-DNA binding in liver and 8-oxo-2′-deoxyguanosinein red blood cells. Carcinogenesis 15, 2911-2917.
(30) Mattagajasingh, S. N., and Misra, H. P. (1996) Mechanisms ofthe carcinogenic chromium(VI)-induced DNA-protein cross-linkingand their characterization in cultured intact human cells. J. Biol.Chem. 271, 33550-33560.
(31) Wise, J. P., Orenstein, J. M., and Patierno, S. R. (1993) Inhibitionof lead chromate clastogenesis by ascorbate: relationship toparticle dissolution and uptake. Carcinogenesis 14, 429-434.
(32) Stearns, D. M., and Wetterhahn, K. E. (1994) Reaction ofchromium(VI) with ascorbate produces chromium(V), chromium-(IV), and carbon-based radicals. Chem. Res. Toxicol. 7, 219-230.
(33) Stearns, D. M., Kennedy, L. J., Courtney, K. D., Giangrande, R.H., Phieffer, L. S., and Wetterhahn, K. E. (1995) Reduction ofchromium(VI) by ascorbate leads to chromium-DNA binding andDNA strand breaks in vitro. Biochemistry 34, 910-919.
(34) Shi, X., Mao, Y., Knapton, A. D., Ding, M., Rojanasakul, Y.,Gannett, P. M., Dalal, N., and Liu, K. (1994) Reaction of Cr(VI)with ascorbate and hydrogen peroxide generates hydroxyl radicalsand causes DNA damage: role of a Cr(IV)-mediated Fenton-likereaction. Carcinogenesis 15, 2475-2478.
(35) da Cruz Fresco, P., and Kortenkamp, A. (1994) The formation ofDNA cleaving species during the reduction of chromate byascorbate. Carcinogenesis 15, 1773-1778.
(36) da Cruz Fresco, P., Shacker, F., and Kortenkamp, A. (1995) Thereductive conversion of chromium(VI) by ascorbate gives rise toapurinic/apyrimidinic sites in isolated DNA. Chem. Res. Toxicol.8, 884-890.
(37) Bridgewater, L. C., Manning, F. C. R., and Patierno, S. R. (1994)Base-specific arrest of in vitro DNA replication by carcinogenicchromium: relationship to DNA interstrand crosslinking. Car-cinogenesis 15, 2421-2427.
(38) Wertz, J. E., and Bolton, J. R. (1986) Experimental methods;spectrometer performance. In Electron Spin Resonance: Elemen-tary Theory and Practical Applications, pp 464-465, Chapmanand Hall, New York.
(39) Tsou, T.-C., Lin, R.-J., and Yang, J.-L. (1997) Mutational spectruminduced by chromium(III) in shuttle vectors replicated in humancells: relationship to Cr(III)-DNA interactions. Chem. Res. Toxi-col. 10, 962-970.
1008 Chem. Res. Toxicol., Vol. 12, No. 10, 1999 Tsou et al.

(40) Tsou, T.-C., Chen, C.-L., Liu, T.-Y., and Yang, J.-L. (1996)Induction of 8-hydroxydeoxyguanosine in DNA by chromium(III)plus hydrogen peroxide and its prevention by scavengers. Car-cinogenesis 17, 103-108.
(41) Shi, X., and Dalal, N. S. (1990) On the hydroxyl radical formationin the reaction between hydrogen peroxide and biologicallygenerated chromium(V) species. Arch. Biochem. Biophys. 277,342-350.
(42) Shibutani, S., Takeshita, M., and Grollman, A. P. (1991) Insertionof specific bases during DNA synthesis past the oxidation-damaged base 8-oxodG. Nature 349, 431-434.
(43) Wood, M. L., Dizdaroglu, M., Gajewski, E., and Essigmann, J.M. (1990) Mechanistic studies of ionizing radiation and oxidativemutagenesis: genetic effects of a single 8-hydroxyguanine (7-hydro-8-oxoguanine) residue inserted at a unique site in a viralgenome. Biochemistry 29, 7024-7032.
(44) Tsou, T.-C., and Yang, J.-L. (1996) Formation of reactive oxygenspecies and DNA strand breakage during interaction of chro-mium(III) and hydrogen peroxide in vitro: evidence for a chro-mium(III)-mediated Fenton-like reaction. Chem.-Biol. Interact.102, 133-153.
TX9802264
Prevention of Cr(VI)/Ascorbate-Induced DNA Damage Chem. Res. Toxicol., Vol. 12, No. 10, 1999 1009





























