Embed Size (px)
Citation preview

Abstract Muscle pain imposes significant changes onnatural motor tasks, but the consequences for stretch re-flexes are still disputed. The present study examined thejaw reflexes to fast (10 ms) stretches of the mandible inan experimental model with local pain in the massetermuscle and remote pain in the tibialis anterior muscle.The stretch reflexes were elicited in healthy volunteers(n=13) before, during, and after periods with constantlevels of experimental pain and while the subjectsclenched at 0%, 15%, 30%, and 45% of the maximalvoluntary contraction (MVC) levels. Surface electromy-ography (EMG) was used to record the reflex responses.Pain in the masseter muscle (mean ± SEM, 3.8±0.4 on a10-cm visual analogue scale), but not in the tibialis ante-rior muscle (3.4±0.3; paired t-test, P=0.318) was associ-ated with significant changes in both prestimulus EMGactivity (ANOVA, P=0.002) and in peak-to-peak ampli-tudes of the stretch reflex (ANOVA, P=0.022). However,when the changes in prestimulus EMG activity were tak-en into consideration a significant increase in the stretchreflex persisted in the painful muscle at 15% and 30%
MVC. Local circuits at the trigeminal level involving thefusimotor system are proposed to mediate a significantpart of this modulatory effect.
Keywords Muscle pain · Trigeminal physiology · Jaw reflexes · Human
Introduction
The jaw-stretch reflex is assumed to contribute to thestiffness of the mandible during movements and posture(Cooker et al. 1980; Luschei and Goldberg 1981). How-ever, the functional significance of the jaw-stretch reflexin craniofacial pain conditions is not known.
It has been reported that the amplitude of the short-latency stretch reflex evoked by a tap to the chin in pa-tients with painful temporomandibular disorders (TMD)is comparable with the amplitudes obtained in controlsubjects (Cruccu et al. 1992). However, the latency issignificantly longer and the amplitude significantlysmaller when the painful side is compared with theasymptomatic side in the TMD patients. In a subsequentstudy, these findings were confirmed and, in 6 out of 25subjects, even an absence of the stretch reflex was re-ported on the painful side (Cruccu et al. 1997). A signifi-cant asymmetry of the jaw-stretch reflex evoked with abite fork pulling the mandible down has also been de-scribed in myofascial TMD patients compared with con-trol subjects (Lobbezoo et al. 1993a). However, compari-son between the groups did not reveal any significantdifference, in particular when the reflex amplitudes werecorrected for background muscle activity (Lobbezoo etal. 1993a). Indeed, the amplitude of the jaw stretch re-flex is highly correlated with the background muscle ac-tivity (Murray and Klineberg 1984; Lobbezoo et al.1993b; Wang et al. 2000). The conflicting results ob-tained in patients, the interindividual variability, and therisk for false-positive diagnoses if stretch reflexes areused as a criterion for pathology (Mohl 1995) encour-aged us to study the phenomenon in the laboratory envi-
P. Svensson (✉ ) · K. WangOrofacial Pain Laboratory, Center for Sensory-Motor Interaction, Aalborg University, Fredrik Bajers Vej 7D-3, 9220 Aalborg E, Denmarke-mail: [email protected]: +45-9815-4008
P. SvenssonDepartment of Prosthetic Dentistry and Stomatognathic Physiology, Royal Dental College, University of Aarhus, Denmark
P. SvenssonDepartment of Oral and Maxillofacial Surgery, Aalborg Hospital, Denmark
G.M. MacalusoIstituto di Clinica Odontoiatrica, University of Parma, Italy
A. De LaatCluster of Oral Physiology, Department of Oral and Maxillofacial Surgery, Catholic University Leuven, Belgium
Exp Brain Res (2001) 139:495–502DOI 10.1007/s002210100806
R E S E A R C H A RT I C L E
Peter Svensson · Guido M. MacalusoAntoon De Laat · Kelun Wang
Effects of local and remote muscle pain on human jaw reflexes evokedby fast stretches at different clenching levels
Received: 19 June 2000 / Accepted: 5 April 2001 / Published online: 3 July 2001© Springer-Verlag 2001

ronment. Recently, an inhibition of the jaw-stretch reflexexcitation of low-threshold motor units was observedduring experimental pain (Svensson et al. 2000), where-as recordings of the reflex with surface and intramuscu-lar electromyography (EMG) showed an increase whenthe subjects clenched at 15% of the maximal voluntarycontraction (Wang et al. 2000). For stretch reflexes inother muscle groups, the results of experimental pain areequivocal. Matre et al. (1998, 1999) have demonstratedsignificant increases in the soleus stretch reflex, whereasZedka et al. (1999) have not find changes in erectorspinae stretch reflexes. It is likely that the modulation ofjaw-stretch reflex by painful inputs depends on the back-ground EMG activity. Thus, the first aim of the presentstudy was to investigate the effect of experimental jaw-muscle pain on the short-latency stretch reflex elicited atfour different levels of background EMG activity.
Another issue that deserves attention is whether pain-induced changes in the jaw-stretch reflex are mediatedvia local neural circuits in the trigeminal system orwhether the changes are part of a more generalized shiftin arousal and attention due to pain. There is some evi-dence that the short-latency component of the complexreflex responses evoked by a tap to the teeth can be mod-ulated by remote pain (Cadden et al. 1996a; van der Glaset al. 2000) as well as by increased attention or anticipa-tion (Cadden et al. 1996b; Scott and Cadden 1996).However, no studies have used the same type of condi-tioning stimulus both in a local and remote pain condi-tion. This is possible with infusion of hypertonic salineinto the masseter and tibialis muscle (Svensson et al.1998; Wang et al. 1999). Thus, the second aim of thepresent study was to compare the effect of local pain inthe masseter muscle to remote pain in the tibialis anteriormuscle on the short-latency jaw-stretch reflex.
Materials and methods
Subjects
A total of 13 subjects, 12 men and 1 woman, were recruited froma student population (mean age ± SEM, 27.0±1.2 years). None ofthe subjects had a history of or clinical signs of TMD according tostandard screening procedures (Okeson 1996). The study was con-ducted in accordance with the Helsinki Declaration, and informedconsent was obtained from all subjects. The local ethics commit-tees in the counties of Viborg and Nordjylland, Denmark, ap-proved the study.
Experimental protocol
The subjects participated in one session lasting about 2.5 h. Thejaw reflex was elicited before, during, and after two pain condi-tions, local and remote. The order was randomized to avoid se-quence effects, with at least 45 min between the two pain condi-tions. In all conditions the jaw-stretch reflexes were elicited at aclenching level of 0%, 15%, 30%, and 45% of the maximal volun-tary contraction (MVC). The order of the clenching levels wasalso randomized.
Recording of jaw-stretch reflexes
The jaw-stretch reflexes were evoked in the left and right masseterand temporalis anterior muscles with a muscle stretcher (Miles etal. 1993; Wang et al. 2000). A stainless steel bite bar was mountedon a frame attached to the floor. A powerful electromagnetic vi-brator (model 406, Ling Dynamic Systems, UK) imposed servo-controlled displacements of the lower jaw bar. A 200-N load cell(5039 A312; Kistler, Switzerland) in series with the moveableprobe of the vibrator measured forces on the lower jaw bar. Thedisplacement of the vibrator probe was measured with a linear po-tentiometer (type 20 FLP 30A-5 K; Sakae, Japan) mounted in par-allel with the probe. Acceleration in the vertical plane was mea-sured by an accelerometer (Delta Tron accelerometer type 4399;Bruel and Kjær, Denmark) mounted on the lower jaw bar. In allexperiments, the short-latency component of the jaw-stretch reflexwas evoked by a displacement of 1 mm with a ramp time of 10 ms(Poliakov and Miles 1994; Wang et al. 2000).
The surface EMG was recorded with use of bipolar disposableelectrodes (4×7 mm recording area; 720-01-k; Neuroline, Medico-test, Denmark) placed 10 mm apart along the central part of themasseter and the temporalis anterior muscles on both sides. Aground electrode soaked with saline was attached to the rightwrist. The EMG signals were amplified 2,000–5,000 times (MK2;Counterpoint, Denmark), filtered to the bandwidth 20 Hz to1 kHz, sampled at 4 kHz, and stored for later analysis.
Subjects were instructed to contract their muscle at a steadylevel of the preset clenching level (i.e., 0%, 15%, 30%, 45%MVC). To help the subjects achieve the correct clenching level,they were shown a screen display of the root-mean-square (rms)value in 200-ms intervals of their EMG activity in the left masse-ter muscle. The screen also showed the level of EMG activity cor-responding with ±10% of the clenching level. The display of theirEMG level changed from green to red when it crossed the upperand lower limits of the window (Svensson et al. 1998). The pro-gram automatically triggered the jaw-muscle stretcher when theEMG activity remained within the preset window for more than400 ms. A total of 300 ms EMG activity was recorded with100 ms prestimulus and 200 ms poststimulus. Sixteen trials withan interstimulus interval of about 10 s were recorded.
Induction of muscle pain
Sterile hypertonic (5.8%) saline was infused into the posterior partof the left masseter muscle about 2 cm away from the surfaceEMG electrode. A standardized infusion paradigm was used with0.2 ml saline infused over 20 s followed by a steady infusion rateof 6 ml/h for the next 440 s and, finally, 9 ml/h for the next 440 s(Svensson et al. 1998, 2000). In the remote pain condition, hyper-tonic saline was infused into the middle part of left tibialis anteriormuscle (15 cm distally from the patella). The infusion was startedwith a bolus of 0.5 ml over 20 s and followed by a steady infusionrate of 18 ml/h for the next 440 s and, finally, 36 ml/h for the last440 s (Wang et al. 1999). For safety purposes, individual changesin infusion rate could be made in steps of 3 ml/h and the infusionsystem could be halted by a keyboard command. The subjectscontinuously scored the pain intensity on a 10-cm electronic visualanalog scale (VAS), with the lower extreme marked “no pain” andthe upper extreme marked “most pain imaginable”. Stretch reflex-es were recorded when the VAS scores were constant 180 s afterthe start of infusion (Wang et al. 1999, 2000). The mean VASscore was calculated starting at the same time as the stretch-reflexrecordings. After the end of the infusion, subjects described thequality of pain on a Danish version of the McGill Pain Question-naire (MPQ) and drew the perceived area of pain on maps show-ing the face and leg, respectively.
Analysis
First, the rms value of the EMG activity in the prestimulus interval(–100 to 0 ms) of the averaged and rectified signal was calculated.
496

In addition, the coefficient of variation (CV) defined as the stan-dard deviation divided by the mean in percentage was calculatedfrom the 16 single sweeps (Svensson et al. 1999). The onset laten-cy and peak-to-peak amplitude of the early reflex component,which appeared as a biphasic potential in the mean of the nonrecti-fied recordings, was measured in the different experimental condi-tions. The peak-to-peak amplitude was then normalized with re-spect to the prestimulus EMG activity. The CV of the peak-to-peak amplitude was also calculated for the 16 sweeps in orderto estimate the reflex variability during the different experimentalconditions.
Statistics
Three-way analysis of variance (ANOVA) was performed with theuse of repeated-measure models. The factors in the ANOVA mod-els were: muscle (4 levels), experimental condition (3 levels), andclenching level (4 levels). Post hoc Student-Newman-Keuls(SNK) tests were used to compensate for multiple comparisons.Student’s t-test was used to test for differences in mean painscores in the two pain conditions. The significance level was set atP<0.05.
Results
Experimental muscle pain
A total of 2.0 ml hypertonic saline was infused into theleft masseter muscle over 15 min. The subjects reporteda mean VAS score of 3.8±0.4 cm during the stretch-reflex recordings. The pain was always located in themuscle, but also irradiated toward the temple (5/13), theupper and lower teeth (5/13), and the temporomandibularjoint (2/13; Fig. 1A). None of the subjects reported painin the masseter 15 min after the infusion was stopped(VAS 0) but a slight tenderness to palpation and clench-ing could occasionally be detected.
A total of 7.1 ml hypertonic saline was infused intothe left tibialis anterior muscle. This caused a mean VASscore of 3.4±0.3 cm during the jaw-stretch recordings.The pain was frequently referred distally to the ankle(9/13), either completely separated from the local painarea (3/9) or in continuum with the local pain area (6/9;Fig. 1B). The mean VAS scores were not significantlydifferent between the local and the remote pain condi-tions (t-test, P=0.318).
Prestimulus EMG activity
The excitability of the motoneuron pool at the time ofthe arrival of the stimulus-evoked afferent barrage is acritical determinant of the amplitude of the stretch reflexand therefore it was important to quantify the prestimu-lus EMG activity. Three-way ANOVAs indicated a sig-nificant main effect of the infusion of hypertonic salineinto the left masseter on the prestimulus EMG activity ofthe other muscles (ANOVA, F=4.56; P=0.020). TheEMG activity of the left masseter muscle was used asfeedback and remained constant (Fig. 2A). The prestim-ulus EMG activity depended on clenching level
(F=25.08; P<0.001) and was different between muscles(F=10.32; P<0.001; Fig. 2A). Post hoc tests demonstrat-ed a significant increase in prestimulus EMG activity ofthe right masseter (nonpainful) at 15% and 45% MVCduring pain and after the pain in the left masseter (SNKP<0.05). In addition, significant increases in EMG activ-ity during and after pain were also noted in the tempora-lis anterior muscles at 45% MVC (SNK P<0.05).
Infusion of hypertonic saline into the tibialis anteriormuscle had no significant effect on the prestimulus EMGactivity in the masseter or temporalis anterior muscles(F=0.028; P=0.972; Fig. 2B). The prestimulus EMG ac-tivity was also dependent on the clenching level(F=25.64; P<0.001) and different between muscles(F=11.96; P<0.001; Fig. 2B) in this experiment.
There was no significant difference between the pre-stimulus EMG activity in the two prepain conditions (be-fore masseter pain versus before tibialis anterior pain) atany level of clenching (F<2.741; P>0.124) and no sig-nificant interactions between different muscles and con-ditions (F<0.926; P>0.438).
The coefficient of variation of the prestimulus EMGactivity (a measure of the variability of the motoneuronexcitability) showed no significant main effect of experi-mental condition either in the local pain experiment(F=1.53; P=0.236) or in the remote pain experiment(F=0.51; P=0.608). The CV values did not differ consis-tently between clenching levels (F<1.47; P>0.249).However, the CV values of the prestimulus EMG activi-ty (F>11.72; P<0.001) were consistently higher in thetemporalis anterior muscles (mean 30.4%) than in themasseter muscles (mean 17.8%; SNK, P<0.011).
Jaw-stretch reflexes
Statistical evaluation of the peak-to-peak amplitude ofthe jaw-stretch reflexes demonstrated a significant main
497
Fig. 1A, B Superimposed drawings of the distribution of per-ceived pain on maps of the face and the leg following infusion ofhypertonic saline into the masseter and tibialis anterior muscles bythe 13 subjects. Gray arrows indicate site of infusion. Note thediffuse and widespread nature of masseter (A) and tibialis anteriorpain (B)
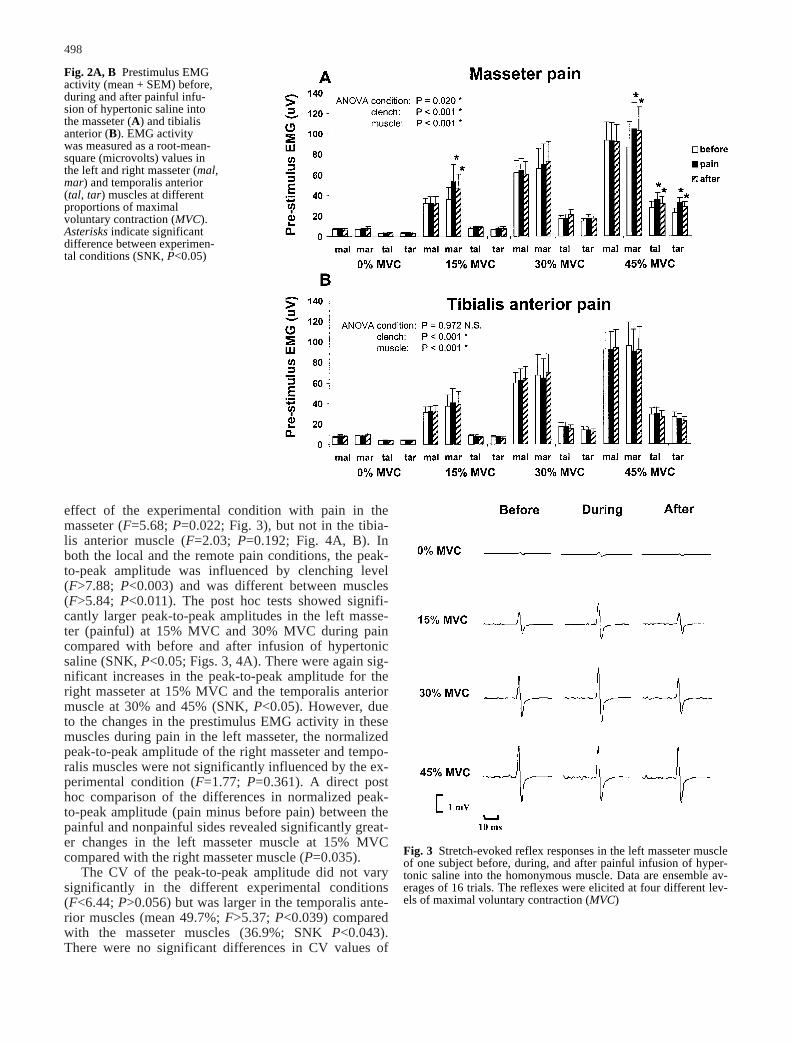
effect of the experimental condition with pain in themasseter (F=5.68; P=0.022; Fig. 3), but not in the tibia-lis anterior muscle (F=2.03; P=0.192; Fig. 4A, B). Inboth the local and the remote pain conditions, the peak-to-peak amplitude was influenced by clenching level(F>7.88; P<0.003) and was different between muscles(F>5.84; P<0.011). The post hoc tests showed signifi-cantly larger peak-to-peak amplitudes in the left masse-ter (painful) at 15% MVC and 30% MVC during paincompared with before and after infusion of hypertonicsaline (SNK, P<0.05; Figs. 3, 4A). There were again sig-nificant increases in the peak-to-peak amplitude for theright masseter at 15% MVC and the temporalis anteriormuscle at 30% and 45% (SNK, P<0.05). However, dueto the changes in the prestimulus EMG activity in thesemuscles during pain in the left masseter, the normalizedpeak-to-peak amplitude of the right masseter and tempo-ralis muscles were not significantly influenced by the ex-perimental condition (F=1.77; P=0.361). A direct posthoc comparison of the differences in normalized peak-to-peak amplitude (pain minus before pain) between thepainful and nonpainful sides revealed significantly great-er changes in the left masseter muscle at 15% MVCcompared with the right masseter muscle (P=0.035).
The CV of the peak-to-peak amplitude did not varysignificantly in the different experimental conditions(F<6.44; P>0.056) but was larger in the temporalis ante-rior muscles (mean 49.7%; F>5.37; P<0.039) comparedwith the masseter muscles (36.9%; SNK P<0.043).There were no significant differences in CV values of
498
Fig. 2A, B Prestimulus EMGactivity (mean + SEM) before,during and after painful infu-sion of hypertonic saline intothe masseter (A) and tibialisanterior (B). EMG activity was measured as a root-mean-square (microvolts) values inthe left and right masseter (mal,mar) and temporalis anterior(tal, tar) muscles at differentproportions of maximal voluntary contraction (MVC).Asterisks indicate significantdifference between experimen-tal conditions (SNK, P<0.05)
Fig. 3 Stretch-evoked reflex responses in the left masseter muscleof one subject before, during, and after painful infusion of hyper-tonic saline into the homonymous muscle. Data are ensemble av-erages of 16 trials. The reflexes were elicited at four different lev-els of maximal voluntary contraction (MVC)
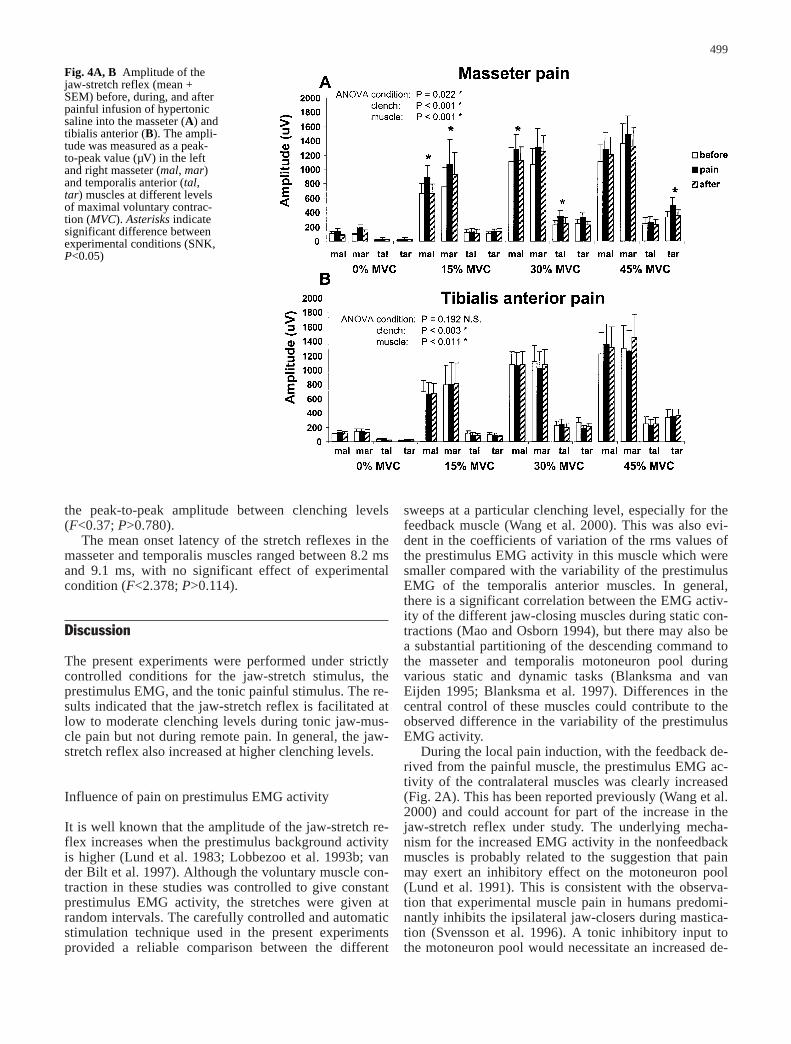
the peak-to-peak amplitude between clenching levels(F<0.37; P>0.780).
The mean onset latency of the stretch reflexes in themasseter and temporalis muscles ranged between 8.2 msand 9.1 ms, with no significant effect of experimentalcondition (F<2.378; P>0.114).
Discussion
The present experiments were performed under strictlycontrolled conditions for the jaw-stretch stimulus, theprestimulus EMG, and the tonic painful stimulus. The re-sults indicated that the jaw-stretch reflex is facilitated atlow to moderate clenching levels during tonic jaw-mus-cle pain but not during remote pain. In general, the jaw-stretch reflex also increased at higher clenching levels.
Influence of pain on prestimulus EMG activity
It is well known that the amplitude of the jaw-stretch re-flex increases when the prestimulus background activityis higher (Lund et al. 1983; Lobbezoo et al. 1993b; vander Bilt et al. 1997). Although the voluntary muscle con-traction in these studies was controlled to give constantprestimulus EMG activity, the stretches were given atrandom intervals. The carefully controlled and automaticstimulation technique used in the present experimentsprovided a reliable comparison between the different
sweeps at a particular clenching level, especially for thefeedback muscle (Wang et al. 2000). This was also evi-dent in the coefficients of variation of the rms values ofthe prestimulus EMG activity in this muscle which weresmaller compared with the variability of the prestimulusEMG of the temporalis anterior muscles. In general,there is a significant correlation between the EMG activ-ity of the different jaw-closing muscles during static con-tractions (Mao and Osborn 1994), but there may also bea substantial partitioning of the descending command tothe masseter and temporalis motoneuron pool duringvarious static and dynamic tasks (Blanksma and van Eijden 1995; Blanksma et al. 1997). Differences in thecentral control of these muscles could contribute to theobserved difference in the variability of the prestimulusEMG activity.
During the local pain induction, with the feedback de-rived from the painful muscle, the prestimulus EMG ac-tivity of the contralateral muscles was clearly increased(Fig. 2A). This has been reported previously (Wang et al.2000) and could account for part of the increase in thejaw-stretch reflex under study. The underlying mecha-nism for the increased EMG activity in the nonfeedbackmuscles is probably related to the suggestion that painmay exert an inhibitory effect on the motoneuron pool(Lund et al. 1991). This is consistent with the observa-tion that experimental muscle pain in humans predomi-nantly inhibits the ipsilateral jaw-closers during mastica-tion (Svensson et al. 1996). A tonic inhibitory input tothe motoneuron pool would necessitate an increased de-
499
Fig. 4A, B Amplitude of thejaw-stretch reflex (mean +SEM) before, during, and afterpainful infusion of hypertonicsaline into the masseter (A) andtibialis anterior (B). The ampli-tude was measured as a peak-to-peak value (µV) in the leftand right masseter (mal, mar)and temporalis anterior (tal,tar) muscles at different levelsof maximal voluntary contrac-tion (MVC). Asterisks indicatesignificant difference betweenexperimental conditions (SNK,P<0.05)

scending drive to get the feedback muscle to the desiredclenching level. Since the corticotrigeminal projectionsare bilateral (Cruccu et al. 1989), with a stronger contra-lateral projection (Butler et al. 2001), the nonfeedbackmuscles may exhibit a net increase in background activi-ty during pain (Svensson et al. 2000).
Importantly for this experiment, there were no signifi-cant differences in the prestimulus EMG activity be-tween the two prepain conditions, which were separatedby at least 45 min. This suggests that there were no car-ry-over effects at this time interval from one pain condi-tion to the next. However, the prestimulus EMG activitywas still increased in the postpain recording after masse-ter pain had disappeared, which confirms the findings ofour previous studies (Wang et al. 2000; Romaniello et al.2000). These results indicate that nociceptive inputshave a longer-lasting (~15 min) effect on trigeminal cir-cuits involved in the control of muscle activity. Ro andCapra (1999) recorded from neurons in nucleus interpo-laris before and after injection of hypertonic saline intothe cat masseter muscle. They found that this nociceptivestimulation caused an initial increase in the cutaneousmechanosensitivity of wide-dynamic-range neurons fol-lowed by a prolonged (~12 min) reversible depression. Itwas suggested that this could represent a central modula-tory response to an acute nociceptive input, which couldalso modulate oral motor behavior. Although the neuralcircuits underlying the observed changes in prestimulusEMG activity still remain speculative, wide-dynamic-range neurons and high-threshold neurons in nucleus in-terpolaris are known to receive muscle afferent input andproject to more rostral nuclei related to trigeminal pre-motor neurons which are associated with the generationof cyclical bursts in the jaw-muscle motoneuron pool(see Ro and Capra 1999).
Another interesting finding of the present study wasthat the modulation in the postpain period and the in-crease in prestimulus EMG activity were absent whentonic pain was induced in a remote muscle. This meansthat the observed changes were mediated at the trigemi-nal level and did not result from a generalized increasein central drive based upon arousal or change of atten-tion (Cadden et al. 1996a, 1996b; Scott and Cadden1996; van der Glas et al. 2000).
The coefficients of variation of the prestimulus EMGactivity were not influenced by the painful conditions.There was more variability in the temporalis muscles,which appeared to be driven in variable ways accordingto the individual biting strategy of the subject. The largervariability in the prestimulus EMG activity in the tempo-ralis anterior muscles was also reflected in higher vari-ability in the peak-to-peak amplitude in these muscles, aswas observed by Wang et al. (2000), and might havemasked relatively small pain-related changes. The lackof significant differences in variability of the peak-to-peak amplitude at different clenching levels is inagreement with the finding of stable, heteronymous H-reflex recordings in the temporalis anterior muscle(Svensson et al. 1999).
Influence of pain on stretch reflexes
The design of the present study allowed discriminationbetween the effects of tonic pain induced locally or in aremote muscle. The level of the pain experienced as wellas the nature and distribution of the pain (Fig. 1A, B)were similar to those described earlier (Graven-Nielsenet al. 1997; Stohler and Kowalski 1999; Svensson et al.1998, 2000; Wang et al. 1999, 2000). It has been arguedthat the quality and intensity of saline-evoked musclepain resembles pain in clinical patients (Stohler and Kowalski 1999) and therefore is a valid model to studythe basic effects of muscle pain on motor performance.
The saline-evoked pain in the present study was notassociated with any change in the onset latency of thejaw-stretch reflex. Previous studies have described onsetlatencies of the jaw-stretch reflex in the range of 8–9 ms(Cooker et al. 1980; Poliakov and Miles 1994; Wang etal. 2000; Wang and Svensson 2001), which is in closeaccordance with the present findings. However, the lackof pain-related latency changes differs from the signifi-cant latency asymmetry reported in patients with chronicTMD pain (Cruccu et al. 1992, 1997). The reason forthis discrepancy is not known. Chronic pain may differin its effects from acute pain, and factors other than painitself, such as structural changes in the muscle tissue,may also contribute to the reflex abnormalities in thechronic condition.
At all clenching levels used in the present study, therewas an increase in the peak-to-peak amplitude of thestretch reflex during pain in the homonymous musclewhen the background EMG activity remained constant;however, this was significant only at the 15% and 30%MVC level (Fig. 3, 4 A). The peak-to-peak amplitude ofthe stretch reflex recorded in the nonpainful masseterand temporalis muscles also tended to increase duringpain, but this was associated with an increase in prestim-ulus EMG activity. A direct comparison between the dif-ference in normalized peak-to-peak amplitude (pain mi-nus prepain) confirmed a significantly greater change inthe painful masseter compared with the nonpainful mas-seter at 15% MVC, but not at 30% MVC, probably be-cause the prestimulus EMG activity in the nonpainfulmasseter not was significantly different between the pre-pain and pain condition (Fig. 2A). The predominant fa-cilitation of the jaw-stretch reflex at 15% and 30% MVCoccurred within the functional range of EMG activity formost of the normal masticatory motor tasks such asspeech, swallowing, and mastication (for a review, seeDubner et al. 1978; Bradley 1995). The functional signif-icance of the short-latency jaw-stretch reflex as opposedto the longer-latency reflex is still under discussion(Cooker et al. 1980; Poliakov and Miles 1994) and needsfurther study under normal and painful conditions.
Several factors may influence the amplitude of thestretch reflex and, in contrast to previous studies, thepresent experiments two important modulating factors tobe evaluated within the same subject and in a single ses-sion. The present findings are consistent with previous
500

reports that local pain facilitates the stretch reflex (Matreet al. 1998, 1999; Wang et al. 2000). A similar facili-tation was not found for the H-reflex in either the limb muscles (Matre et al. 1998) or the jaw muscles(Svensson et al. 1998). This suggests that the reason forthe increase is not to be found in an increased excitabili-ty of the motoneuron pool or changes in tonic presynap-tic inhibition of primary muscle spindles (Svensson et al.2000; Wang et al. 2000). Capra and Ro (2000) have re-cently reported that injection of hypertonic saline intothe cat masseter muscle is associated with significantchanges in the mean firing rates of neurons in the medialedge of subnucleus interpolaris and the adjacent parvi-cellular reticular formation. These neurons were subdi-vided into dynamic-static or static cells depending ontheir responses to different speeds and amplitudes of jawmovements. A clear inhibitory effect of hypertonic salinewas noted on the static neurons, whereas a majority ofthe responsive dynamic-static neurons increased theirmean firing rates during pain. Capra and Ro (2000) aswell as Wang et al. (2000) ruled out the possibility ofvolume effects, since injection of isotonic saline was notassociated with changes in reflex excitability. Further-more, we have demonstrated that the facilitation of thejaw-stretch reflex cannot be explained by cross-talk fromfacial expression activity during pain, since intramuscu-lar wire electrodes inserted into the masseter musclesdemonstrated facilitatory effects similar to those seen insurface EMG recordings (Wang et al. 2000). Capra andRo (2000) have suggested that the fusimotor drive,through interneurons, could be altered by the nociceptivebarrage from the muscle and that this subsequentlywould have an effect on the muscle spindle primary andsecondary endings. Hellström et al. (2000) have recentlyshown that injection of bradykinin into the massetermuscle of cats increases the responses of single musclespindle afferents from the trapezius and splenius mus-cles, which suggests increased fusimotor drive. Thiswould correspond to an increase in reflex-mediated mus-cle stiffness which, however, would not necessarilycause an increase in the resting activity of the alpha-motoneurons to a level that is manifest as an increase inEMG activity of the resting muscle (Hellström et al.2000).
Finally, the present observation of an absence of re-flex modulation during remote pain strengthens the sug-gestion that the nociceptive-induced changes in trigemi-nal reflexes occur at the trigeminal level, and that gener-al factors related to arousal or attention do not accountfor the facilitation of the reflex after local pain.
In conclusion, the present study demonstrates that theamplitude of the jaw-stretch reflex is facilitated by local,but not remote muscle pain when the background EMGactivity is taken into consideration. These pain-inducedchanges are most pronounced at 15% and 30% MVC,suggesting a possible functional significance of the jaw-stretch reflex during natural motor functions such asspeech, swallowing, and mastication.
Acknowledgements The Danish National Research Foundationand the Danish Dental Association supported the study. A.D.L.and G.M.M. were visiting professors at SMI supported by a grantfrom the Danish Rectors’ Conference and by grants from the Ital-ian and Flemish Governments, respectively. The valuable com-ments from Dr. Timothy S. Miles are greatly appreciated.
References
Bilt A van der, Ottenhoff FAM, Glas HW van der, Bosman F, Abbink JH (1997) Modulation of the mandibular stretch reflexsensitivity during various phases of rhythmic open-closemovements in humans. J Dent Res 76:839–847
Blanksma NG, Eijden TM van (1995) Electromyographic hetero-geneity in the human temporalis and masseter muscles duringstatic biting, open/close excursions, and chewing. J Dent Res74:1318–1327
Blanksma NG, Eijden TM van, Ruijven LJ van, Weijs WA (1997)Electromyographic heterogeneity in the human temporalis andmasseter muscles during dynamic tasks guided by visual feed-back. J Dent Res 76:542–551
Bradley RM (1995) Essentials of oral physiology. Mosby, St.Louis
Butler SL, Miles TS, Thompson PD, Nordstrom MA (2001) Taskdependent control of human masseter muscles from ipsilateraland contralateral motor cortex. Exp Brain Res 137:65–70
Cadden SW, Glas HW van der, Lobbezoo F, Bilt A van der(1996a) Effects of remote noxious stimulation on the extero-ceptive reflexes in human jaw-closing muscles. Brain Res 726:189–197
Cadden SW, Glas HW van der, Lobbezoo F, Bilt A van der(1996b) The influence of attentional factors on short- andlong-latency jaw reflexes in man. Arch Oral Biol 41:995–998
Capra NF, Ro JY (2000) Experimental muscle pain produces cen-tral modulation of proprioceptive signals arising from jawmuscle spindles. Pain 86:151–162
Cooker HS, Larson CR, Luschei ES (1980) Evidence that the hu-man jaw stretch reflex increases the resistance of the mandibleto small displacements. J Physiol (Lond) 308:61–78
Cruccu G, Berardelli A, Inghilleri, Manfredi M (1989) Functionalorganization of the trigeminal motor system in man. A neuro-physiological study. Brain 112:1333–1350
Cruccu G, Frisardi G, Steenberghe D van (1992) Side asymmetryof the jaw jerk in human craniomandibular dysfunction. ArchsOral Biol 37:257–262
Cruccu G, Frisardi G, Pauletti G, Romaniello A, Manfredi M (1997)Excitability of the central masticatory pathways in patients withpainful temporomandibular disorders. Pain 73:447–454
Dubner R, Sessle BJ, Storey AT (1978) The neural basis of oraland facial function. Plenum Press, New York
Glas HW van der, Cadden SW, Bilt A van der (2000) Mechanismsunderlying the effects of remote noxious stimulation and men-tal activities on exteroceptive jaw reflexes in man. Pain 84:193–202
Graven-Nielsen T, Arendt-Nielsen L, Svensson P, Jensen TS(1997) Stimulus-response functions in areas with experimen-tally induced referred muscle pain – a psychophysical study.Brain Res 744:121–128
Hellström F, Thunberg J, Bergenheim M, Sjölander P, Pedersen J,Johansson H (2000) Elevated intramuscular concentration ofbradykinin in jaw muscles increases the fusimotor drive toneck muscles in the cat. J Dent Res 79:1815–1822
Lobbezoo F, Glas HW van der, Buchner R, Bosman F (1993a) Bi-lateral asymmetries in the jaw-jerk reflex activity in man.Archs Oral Biol 38:689–698
Lobbezoo F, Glas HW van der, Buchner R, Bilt A van der,Bosman F (1993b) Jaw-jerk reflex activity in relation to vari-ous clenching tasks in man. Exp Brain Res 93:139–147
Lund JP, Lamarre Y, Lavigne G, Duquet G (1983) Human jaw re-flexes. Adv Neurol 39:739–755
501

Lund JP, Donga R, Widmer CG, Stohler CS (1991) The pain-adap-tation model: a discussion of the relationship between chronicmusculoskeletal pain and motor activity. Cand J Physiol Phar-macol 69:683–694
Luschei ES, Goldberg LJ (1981) Neural mechanisms of mandibu-lar control: mastication and voluntary biting. In: Brooks VB(ed) Handbook of physiology. Motor Control, vol 2. AmericanPhysiological Society, Bethesda, pp 1237–1274
Mao J, Osborn JW (1994) Direction of a bite force determines thepattern of activity in jaw-closing muscles. J Dent Res 73:1112–1120
Matre DA, Sinkjaer T, Svensson P, Arendt-Nielsen L (1998) Ex-perimental muscle pain increases the human stretch reflex.Pain 75:331–339
Matre DA, Sinkjaer T, Knardahl S, Andersen JB, Arendt-NielsenL (1999) The influence of experimental muscle pain on the hu-man soleus stretch reflex during sitting and walking. ClinNeurophysiol 110:2033–2043
Miles TS, Poliakov AV, Flavel SC (1993) An apparatus for con-trolled stretch of human jaw-closing muscles. J Neurosci Meth46:197–202
Mohl ND (1995) Standards for testing new diagnostic strategiesfor temporomandibular disorders. In: Fricton JR, Dubner RB(eds) Orofacial pain and temporomandibular disorders. RavenPress, New York pp 265–275
Murray GM, Klineberg IJ (1984) Electromyographic recordings ofhuman jaw-jerk reflex characteristics evoked under standardi-sed conditions. Archs Oral Biol 29:537–549
Okeson JP (1996) Orofacial pain. Guidelines for assessment, diag-nosis, and management. Quintessence, Chicago
Poliakov AV, Miles TS (1994) Stretch reflexes in human masseter.J Physiol (Lond) 476:323–331
Ro JY, Capra NF (1999) Evidence for subnucleus interpolaris incraniofacial muscle pain mechanisms demonstrated by intra-muscular injections with hypertonic saline. Brain Res 842:166–183
Romaniello A, Cruccu G, McMillan AS, Arendt-Nielsen L, Svensson P (2000) Effect of experimental pain from trigemi-nal muscle and skin on motor cortex excitability in humans.Brain Res 882:120–127
Scott AJ, Cadden SW (1996) Suppression of an inhibitory jaw re-flex by the anticipation of pain in man. Pain 66:125–131
Stohler CS, Kowalski CJ (1999) Spatial and temporal summationof sensory and affective dimensions of deep somatic pain. Pain79:165–173
Svensson P, Arendt-Nielsen L, Houe L (1996) Sensory-motor in-teractions of human experimental unilateral jaw muscle pain: aquantitative analysis. Pain 64:241–249
Svensson P, De Laat A, Graven-Nielsen T, Arendt-Nielsen L(1998) Experimental jaw-muscle pain does not change heter-onymous H-reflexes in the human temporalis muscle. ExpBrain Res 121:311–318
Svensson P, De Laat A, Graven-Nielsen T, Arendt-Nielsen L,Macaluso GM (1999) Effect of clenching levels on heterony-mous H-reflex in human temporalis muscle. Exp Brain Res126:467–472
Svensson P, Miles TS, Graven-Nielsen T, Arendt-Nielsen L (2000)Modulation of stretch-evoked reflexes in single motor units inhuman masseter muscle by experimental pain. Exp Brain Res132:65–171
Wang K, Svensson P (2001) Influence of methodological parame-ters on human jaw-stretch reflexes. Eur J Oral Sci 109:1–9
Wang K, Svensson P, Arendt-Nielsen L (1999) Modulation of ex-teroceptive suppression periods in human jaw-closing musclesby local and remote experimental muscle pain. Pain 82:253–262
Wang K, Svensson P, Arendt-Nielsen L (2000) Effect of tonicmuscle pain on short-latency jaw stretch reflexes in humans.Pain 88:189–197
Zedka M, Prochazka A, Knight B, Gillard D, Gauthier M (1999)Voluntary and reflex control of human back muscles during in-duced pain. J Physiol (Lond) 520:591–604
502