Embed Size (px)
Citation preview

Eur J Appl Physiol (1984) 52:414-419 European Joumal of
Applied Physiology and Occupational Physiology �9 Springer-Verlag 1984
Effects of iron deficiency and exercise on myoglobin in rats
Roger McDonald r, Jack Hegenauer 2, Anthony Sucec 1, and Paul Saltman z
I Department of Physical Education, San Diego State University, San Diego, California 92182 2 Department of Biology, University of California, San Diego, La Jolla, California 92093, USA
Summary. The effects of iron deficiency and endur- ance training on muscle myoglobin (Mb), body weights, and blood lactic acid concentration were studied in rats. Fifty animals were divided into four groups: anemic trained (AT), normal trained (NT), anemic sedentary (AS), and normal sedentary (NS). Following 5 weeks of dietary control, the mean hemoglobin values for the AT and AS rats were 0.013 + 0.002 mmol�9 1-1 (8.7 4- 1.4 g �9 d1-1) and 0.014 + 0.003 mmol�9 1-1 (9.2 + 1.7 g �9 d1-1) respectively, and did not significantly change throughout the study. AT and NT rats were run on a motor driven treadmill 4 days/week for 6 weeks up to a pre-established time of 90 min. Following the training, body weights of the AT (157 + 13 g) and NT (153 + 13 g) rats were lower than their respective sedentary groups AS (172 _+ 9 g) and NS (176 + 15 g). Resting blood lactic acid concentration following training was lower in both trained groups, AT (3.3 + 2.0 raM) and NT (2.3 + 1.9 mM) compared to AS (8.2 + 2.6 mM) and NS (3.8 _+ 1.6 mM). Training increased Mb concentration in hearts of both the anemic and normal trained groups (AT, 0.66 _+ 0.13 mg �9 g-l; NT, 0.95 + 0.08 mg�9 g-l) compared to the sedentary groups (AS, 0.44 + 0.08 mg�9 g-l; NS, 0.70 + 0.13 mg �9 g-l). Only the AT rats showed an increase in skeletal muscle Mb. This study provides evidence that myoglobin may limit aerobic metabolism.
Key words: Iron deficiency - Myoglobin - Endur- ance training - Lactic acid - Hemoglobin
Offprint requests to R. McDonald's current address: Department of Biology, B-C 22 University of California, San Diego, LA Jolla CA, 92093, USA
Introduction
Iron-deficiency anemia has been shown to impair physical work performance (Anderson and Barkve 1970; Beutler et al. 1960; Vellar and Hermansen 1972; Wittenberg 1970). In these investigations, reduction of hemoglobin (Hb) as a result of dietary iron deficiency or phlebotomy has been viewed as the rate limiting factor in physical work performance. Although Hb accounts for 90% of total body iron content (Jacobs 1969), recent investigations have suggested that iron deficiency reduces the concen- tration of many, if not all, intracellular iron-contain- ing proteins (Dallman and Schwartz 1965; Ohira et al. 1982b). Decreased concentrations of heme iron or porphyrin complexes (Hagler et al. 1981; Hickson 1981), as well as mitochondrial iron-sulfur proteins (Finch et al. 1979; Gollnick et al. 1972; Holloszy and Oscai 1969), may reduce the oxidative potential during exercise of iron deficient animals.
The enhancement of 02 consumption in response to endurance training has been linked to the cell's ability to adapt biochemically to increased demands for energy (Saltin et al. 1976). Although changes in skeletal muscle iron-containing respiratory proteins in response to endurance training are well docu- mented (Gollnick et al. 1972; Saltin et al. 1976), few studies have investigated the role of myoglobin (Mb) in oxygen transfer. High concentrations of Mb in skeletal muscle found in diving sea mammals (Wit- tenberg 1970) suggests that Mb may serve as a temporary oxygen store. Ohira et al. (1982a) have also reported rapid changes in Mb concentration in rats subjected to short term endurance training and suggest that Mb may have a more direct role in work performance than is suggested by the oxygen store theory.

R. McDonald et al.: Muscle proteins and endurance training
T h e p r e s e n t s t u d y e x a m i n e s t h e e f fec t s o f i r o n - d e -
f i c i ency a n e m i a a n d e n d u r a n c e t r a i n i n g o n M b in
o r d e r to d e f i n e m o r e spec i f i ca l ly its ro l e in o x i d a t i v e
m e t a b o l i s m .
415
removed. The adhering blood was rinsed off, and connective tissue was cut away before weighing. Fresh muscle was homogenized in 2 ml of 0.175 M potassium chloride using a Polytron (Brinkmann Instruments, Westbury, NY, USA).
Methods
Animals and diet
Forty female weanling Sprague-Dawley rats (Simonson Labora- tories, Gilroy, CA, USA) were housed five per cage in plastic cages with stainless steel tops and raised plastic grid floors to prevent coprophagy and ingestion of adventitious iron. Animals were divided into four groups: training with normal diet (NT), training with low-iron diet (AT), sedentary with normal diet (NS), and sedentary with low-iron diet (AS). Iron deficient or normal diet and distilled water were given ad libitum. The semisynthetic iron-deficient diet contained less than 4 mg �9 kg -1 of iron (Ohira et al. 1982a); the normal diet was identical but contained 250 mg �9 kg -1 of iron in the form of ferrous sulphate. Some mortality was observed, with 32 animals finishing the experiment.
Training
After 5 weeks of dietary control, blood was drawn from the tail vein of each animal to determine Hb concentration for baseline iron status. On the following day the NT and AT groups began training. Rats were run on a five-lane motor driven treadmill (W. E. Collins, Brainbridge, MA, USA) 3 days per week for 6 weeks. Grade and speed were progressively increased from an initial grade of six degrees and a speed of 0.40 m �9 s -~ until the rats could run for 90 min at a grade of 10 degrees and a speed of 0.55 m - s -1 (Table 1). Daily mean training times were recorded for each experimental group. On the day following the final training session, a maximal work performance test was given to NT and AT groups. The end of the test was determined when an animal did not display the righting reflex when placed on its back.
T~sue preparagon
Within 24 h of the final training session, rats in all groups were killed by CO2 inhalation. Blood was drawn by ventricular heart puncture for lactic acid and Hb determination. CO2 inhalation does not effect lactic acid concentration. Heart and the lower leg complex of muscles (plantaris, soleus, and gastrocnemius) were
Biochemical analysis
Hemoglobin concentration was determined spectrophotometrical- ly as cyanmethemoglobin. Lactic acid concentration of blood was measured using an alpha-lactic acid diagnostic kit (+826a, Sigma Chemical Company St. Louis, MO, USA).
Myoglobin was determined by methods described by Akeson et al. (1960) and Paul et al. (1953). Snpernatant from muscle homogenate was fractionated in the cold by 50% saturation with ammonium sulphate and centrifugation at 12,000 g for 10 rain. The supernatant was dialyzed against two 40-volume changes of 0.02 M sodium phosphate buffer (pH 6.5) for 48 h in the dark. The sample was then chromatographed on a 1- x 10-cm column of carboxy- methylcellulose (CM-52, Whatman Inc., Clifton, NJ, USA) equilibrated with the same buffer. A 2-ml aliquot of effluent was mixed with 0.5 ml of distilled pyridine and 0.5 ml of 0.3 N sodium hydroxide. The sample was reduced with solid sodium dithionite, and tissue Mb concentration was determined immediately from the A~50nm-Ass7nm difference spectrum of the reduced pyridine hemochrome.
Unpaired two-tailed t-tests were used to evaluate differences between the experimental and control groups following the training period. All results are expressed as mean _+ SD Differences were considered significant at P < 0.05.
Results
Training and maximal work performance
D a i l y m e a n t r a i n i n g t i m e s a r e p r e s e n t e d in Fig . 1.
D u r i n g t h e f irs t 8 days o f t r a in ing , t r a in ing d u r a t i o n
i n c r e a s e d in a g e n e r a l l y l i n e a r f a sh ion ; t h e A T g r o u p
p r o g r e s s e d at a s l o w e r ra te . O n t r a in ing days 1 1 - 1 2 ,
t h e A T g r o u p s h o w e d an a c c e l e r a t e d t r a in ing du ra -
t i on such tha t o n days 1 2 - 1 4 t h e r e was no d i f f e r e n c e
in t r a i n i n g d u r a t i o n b e t w e e n t h e t w o g roups . Th is
r e su l t was u n e x p e c t e d a n d has n o t b e e n r e p o r t e d
e l s e w h e r e . H o w e v e r , on t r a i n i n g d a y 15, t w o an ima l s
in t h e A T g r o u p d i e d w h i l e r u n n i n g , a n d t h e o t h e r
an ima l s w e r e u n a b l e to m a i n t a i n t h e t r a in ing l eve l s o f
Table 1. Endurance training protocol for anemic and normal rats performed on motor driven treadmill
Endurance training protocol
Day of training Speed (m - s -1) Grade (degrees)
1 - 3 0.40 6 4 - 6 0.43 6 7 - 9 0.46 8
10-12 0.50 8 13-15 0.55 10 16-18 0.55 10
loo
(Nr) "~ 75
~ / ANEMIC (AT)
r- 50 g
DAY OF TRAI~ING ~ 6 9 'i 115 118 WEEK ! 2 3 4 5 6
Fig. 1. Mean daily treadmill training times of anemic AT (n = 6) and normal NT (n = 7) rats
416
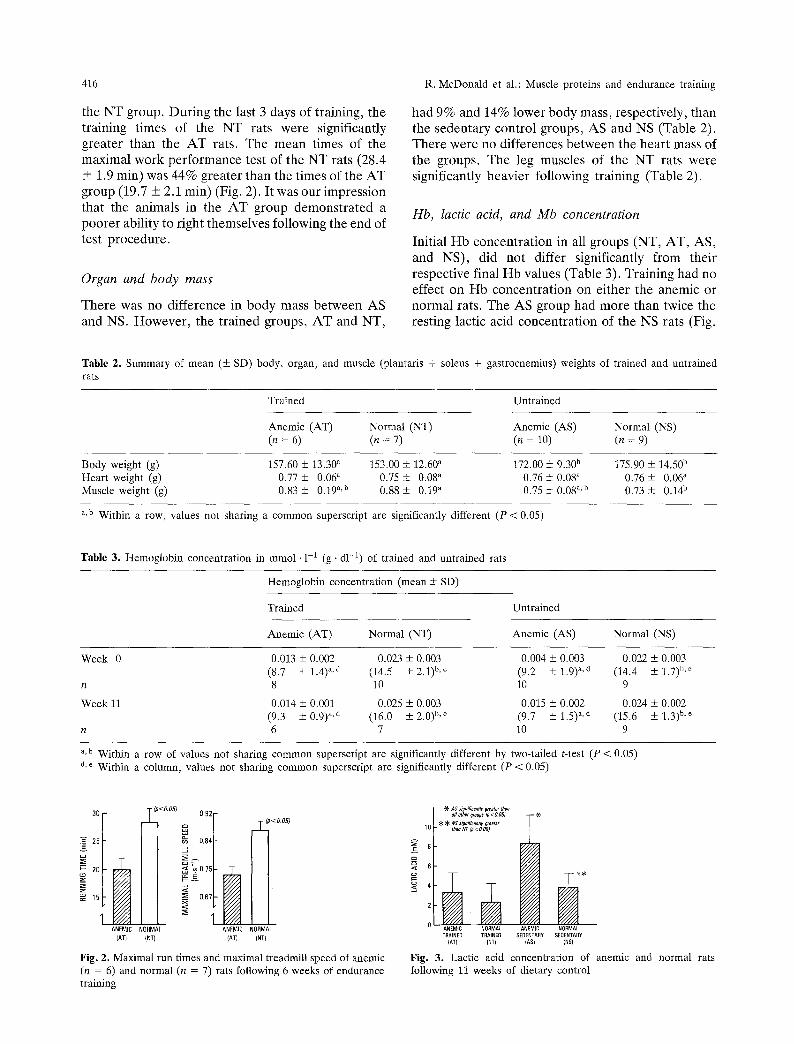
the NT group. During the last 3 days of training, the training times of the NT rats were significantly greater than the AT rats. The mean times of the maximal work performance test of the NT rats (28.4 + 1.9 min) was 44% greater than the times of the AT group (19.7 + 2.1 min) (Fig. 2). It was our impression that the animals in the AT group demonstrated a poorer ability to right themselves following the end of test procedure.
Organ and body mass
There was no difference in body mass between AS and NS. However, the trained groups, AT and NT,
R. McDonald et al.: Muscle proteins and endurance training
had 9% and 14% lower body mass, respectively, than the sedentary control groups, AS and NS (Table 2). There were no differences between the heart mass of the groups. The leg muscles of the NT rats were significantly heavier following training (Table 2).
Hb, lactic acid, and Mb concentration
Initial Hb concentration in all groups (NT, AT, AS, and NS), did not differ significantly from their respective final Hb values (Table 3). Training had no effect on Hb concentration on either the anemic or normal rats. The AS group had more than twice the resting lactic acid concentration of the NS rats (Fig.
Table 2. Summary of mean (• SD) body, organ, and muscle (plantaris + soleus + gastrocnemius) weights of trained and untrained rats
Trained Untrained
Anemic (AT) Normal (NT) Anemic (AS) Normal (NS) (n = 6) (n = 7) (n = 10) (n = 9)
Body weight (g) 157.60 _+ 13.30 a 153.00 • 12.60 a 172.00 • 9.30 b 175.90 _+ 14.50 b Heart weight (g) 0.77 + 0.06 ~ 0.75 + 0.08" 0.76 • 0.08 ~ 0.76 + 0.06 ~ Muscle weight (g) 0.83 + 0.19 ~,b 0.88 + 0.19" 0.75 • 0.08 ~,b 0.73 _+ 0.14 b
a,b Within a row, values not sharing a common superscript are significantly different (P < 0.05)
Table 3. Hemoglobin concentration in mmol �9 1-1 (g �9 d1-1) of trained and untrained rats
Hemoglobin concentration (mean • SD)
Trained Untrained
Anemic (AT) Normal (NT) Anemic (AS) Normal (NS)
Week 0 0.013 _+ 0.002 0.023 +_ 0.003 0.004 + 0.003 0.022 _+ 0.003 (8.7 + 1.4) a,d (14.5 + 2.1) b,e (9.2 +_ 1.9) a'd (14.4 +_ 1.7) b'e
n 8 10 10 9
Week 11 0.014 _+ 0.001 0.025 + 0.003 0.015 + 0.002 0.024 +_ 0.002 (9.3 + 0.9)a,d (16.0 -t- 2.0)b,e (9.7 +--- 1.5) "'d (15.6 _+_ ].3)b,e
n 6 7 i0 9
~,b Within a row of values not sharing common superscript are significantly different by two-tailed t-test (P < 0.05) d,r Within a column, values not sharing common superscript are significantly different (P < 0.05)
25:
N ~- 20
15 n
0.92
o.84
F-g
_~ 0.67
ANEMIC NORMAL ANEMIC NORMAL (AT) (NT) (ATI [NT}
Fig. 2. Maximal run times and maximal treadmill speed of anemic (n = 6) and normal (n = 7) rats following 6 weeks of endurance training
10
R
6
4
AS si#ni&ant/}, greater than a/l ether groups (g < ~ 05)
~4 NS significantly greater than NT (p<GOg)
ANEMIC NORMAL TRAINER TRAINER
(AT) INT)
ANEMIC NORMAL SEDENTARY SERENTARY
(AS) INS)
Fig. 3. Lactic acid concentration of anemic and normal rats following 11 weeks of dietary control

R. McDonald et al.: Muscle proteins and endurance training 417
Table 4. Mean (+ SD) myoglobin (Mb) concentration in heart and leg muscles (plantaris + soleus + gastrocnemius) of anemic and normal rats
Muscle Mb concentration (rag-g-1 tissue)
Trained Untrained
Anemic (AT) Normal (NT) Anemic (AS) Normal (NS) (n = 6) (n = 7) (n = 10) (n = 9)
Heart 0.66 + 0.13 ~ 0.95 _+ 0.17 b 0.44 + 0.08 c 0.70 _+ 0.13 a Leg muscle 0.34 +_ 0.10 ~ 0.26 + 0.05 u'd 0.14 + 0.04 c 0.24 + 0.05 d
a.b,c, d Within a row, values not sharing a common superscript are significantly different (P < 0.05)
3). Training significantly lowered resting lactic con- centration in the AT and the NT rats when compared with their sedentary controls.
Mb concentration in heart was significantly great- er in the trained groups, than in their sedentary grotips (Table 4). Mb concentration of leg muscle was significantly greater only in the AT group as a result of training. Lower Mb concentrations were seen in the heart muscle in both anemic groups, AT and AS. Skeletal muscle Mb concentration was also depressed as a result of iron deficiency in the AS group when compared to the NS.
Discussion
The creation of severe iron-deficiency anemia through dietary restriction o1: phlebotomy has been associated with many abnormalities (Braunwald 1980; Cain 1965; Chapler et al. 1979; Vatner et al. 1972). These abnormalities are exacerbated by the severity of the anemia. It would therefore seem appropriate to establish an anemic state which would minimize the physiological stress placed on the animal yet allow for the hematologic effects of iron deficiency. The iron deficiency produced here was sufficiently pronounced to establish the desired effects of anemia (i.e., low concentrations of Hb and Mb), yet was not so severe as to be considered out of range of anemias associated with endurance perfor- mance in humans (Yoshimura 1970).
It was the intent of this investigation to provide an endurance training protocol that could be considered moderate , yet produce accepted training effects (Mathews and Fox 1976). In this regard previous investigations (Hagler et al. 1980; Holloszy and Oscai 1969) have used protocols that were approximately twice the intensity of ours and produced training effects more likely to be seen in highly trained human subjects. Considering the ability of our anemic rats to achieve a training state equM to that of the normal
rats, we consider our protocol to have been suffi- ciently intense.
The anemic animals' inability to perform at maximal work rates comparable to those of normal animals (Fig. 2) was expected and confirms results of previous investigations (Andersen and Barkve 1970; Davies et al. 1982; Ohira et al. 1982a; Wranne and Woodson 1973; Woodson et al. 1978). However , the ability of the trained anemic rats to achieve, if only temporarily, submaximal work similar levels to normal trained animals has not been previously described. This was a surprising result, which may be partly explained by the significant increase in heart and leg muscle Mb concentration of the A T rats over those of the AS rats. Training enabled the A T rats to compensate for a lower concentration of Hb and Mb associated with iron-deficiency anemia by increasing their Mb concentration as would be expected in normal trained rats. Investigations to date have failed to identifiy a specific iron-containing protein which is responsible for limiting or enhancing work perfor- mance (Davies et al. 1982; Finch et al. 1979). This would suggest an inter-relation of iron-containing proteins involved in aerobic metabolism. Although there are many factors affecting work performance, Mb concentration seems to have a particular impact on our animals' ability to perform at submaximal workloads.
The difference in work performance between normal and anemic animals suggests that submaximal and maximal work performance are limited by separate mechanisms. In our investigation, iron in both anemic and normal animals was preferentially distributed to increase Mb without significantly affecting Hb concentration following endurance training (Tables 3 and 4). These data suggest that the oxygen content of the blood, even in an anemic state of 0.014-0.015 mmol - 1-1 ( 9 - 1 0 g �9 dl-1), is suffi- cient to supply the respiratory needs of the cell during submaximal performance. However, when the ani- mal is worked beyond submaximal effort, as in our maximal work performance test, the oxygen content of the blood may not meet requirements.

418 R. McDonald et al.: Muscle proteins and endurance training
H igh res t ing lact ic acid concen t r a t i on in anemic animals have been o b s e r v e d by F inch et al. (1979) and Ca in (1965). I t wou ld a p p e a r tha t the increases in res t ing lac ta te , also seen in the A S rats , a re a resul t of i n c o m p l e t e ox ida t ion of lac ta te . B r o o k s and G a e s s e r (1980) d e m o n s t r a t e d t h rough the use of [14-C] lac ta te tha t a p p r o x i m a t e l y 45% of lac ta te fo l lowing r ecove ry f rom exerc ise is ox id ized to CO2. Prev ious invest i- ga t ions have r e p o r t e d d e c r e a s e d act ivi ty of the i ron-con ta in ing m i t o c h o n d r i a l enzymes , assoc ia ted with high lactic acid concen t r a t ions (Go l ln i ck et al. 1972; O h i r a et al. 1982a, b) dur ing i ron-def ic iency anemia . These r epo r t s e m p h a s i z e d tha t the t r icar- boxyl ic acid cycle mus t func t ion n o r m a l l y for eff icient c l ea rance of l ac ta te to occur . H o w e v e r , the high lac ta te levels a s soc ia t ed with d e c r e a s e d act ivi ty of a l p h a - g l y c e r o l p h o s p h a t e d e h y d r o g e n a s e , an i ron- con ta in ing e n z y m e invo lved in glycolysis , r e p o r t e d by F inch et al. (1979), w o u l d suggest tha t the increase in lac ta te a ssoc ia ted with i ron def ic iency is a funct ion of p r o d u c t i o n and not c l ea rance . W h e t h e r the high res t ing l ac ta te concen t r a t i on in i ron-def ic ien t an imals is due to inc reased p r o d u c t i o n and /o r dec rea sed c lea rance is uneso lved . H o w e v e r , in our invest iga- t ion, the A T rats were able to a d a p t by deve lop ing some m e c h a n i s m to o v e r c o m e the high l ac ta te concen t ra t ions assoc ia ted with i ron-def ic iency ane- mia (Fig. 3).
M a m m a l s show c o m p e n s a t o r y mechan i sms fol- lowing i ron-def ic iency a n e m i a tha t a re complex and remain to be fully unde r s tood . The resul ts p r e s e n t e d he re suggest tha t rats a d a p t the i r m e t a b o l i s m to low H b concen t r a t ions and the inc reased d e m a n d s of exerc ise by increas ing muscle Mb . The classical a p p r o a c h which descr ibes H b as the p r i m a r y l imit ing fac tor in w o r k p e r f o r m a n c e dur ing i ron-def ic iency a n e m i a needs to be m o d i f i e d to account for o the r i ron p ro t e in s of oxygen t r anspo r t and ox ida t ive resp i ra -
t ion.
Acknowledgements. The authors express their appreciation to Bruce Dyke, Ellen Chinn, and Markus Lauber for their technical assistance; to the Naval Health Research Center, San Diego for the use of the treadmill; and to Dr. Linda Strause for her advice and assistance.
This work was supported by USPHS NIH Research Grant AM 12386.
References
Akeson A, Ehrenstein G, Hevesy G, Theorell T (1960) Lifespan of myoglobin. Arch Biochem 91:310-318
Andersen HT, Barkve H (1970) Iron deficiency and muscular work performance; an evaluation of cardio-respiratory function of iron deficient subjects with and without anemia. Scand J Clin Lab Invest [Suppl 25] 144:1-39
Beutler E, Laush S, Tanzi F (1960) Iron enzymes in iron deficiency: VII. Oxygen consumption measurements in iron-deficient subjects. Am J Meal Sci 239:759-765
Braunwald E (1980) Heart Disease: A textbook of cardiovascular medicine, vol 1. WB Saunders Co, Philadelphia
Brooks GA, Gaesser GA (1980) End points of lactate and glucose metabolism after exhausting exercise. J Appl Physiol: Respir Environ Exercise Physiol 49:1057-1069
Cain SM (1965) Appearance of excess lactate in anesthetized dogs during anemic and hypoxic hypoxia. Am J Physiol 209:604-610
Chapler CK, Cain SM, Stainsby WN (1979) Blood flow and oxygen uptake in isolated canine skeletal muscle during acute anemia. J Appl Physiol: Respir Environ Exercise Physiol 46:1035-1038
Dallman PR, Schwartz HC (1965) Distribution of cytochrome c and myoglobin in rats with dietary iron deficiency. Pediatrics 35 : 677-686
Davies KJ, Maguire JJ, Dallman PR, Brooks GA, Packer L (1982) Exercise bioenergetics during dietary iron deficiency and repletion. In: Saltman P, Hegenauer J (eds) The biochemistry and physiology of iron. Elsevier/North Holland, New York Oxford Shannon, pp 591-594
Faulkner JA, Maxwell LC, Brook D, Lieberman DA (1971) Adaptation of guinea pig plantaris muscle fibers to endurance training. Am J Physiol 221:291-297
Finch CA, Gollinick PD, Hlastala MP, Miller LR, Dillmann E, Mackler B (1979) Lacticidosis as a result of iron deficiency. J Clin Invest 64:129-137
Gollnick PD, Armstrong RB, Saubert CW, Piehl K, Saltin B (1972) Enzyme activity and fiber composition in skeletal muscle of untrained and trained men. J Appl Physiol 33:312-319
Hagler L, Coppes RI, Askew EW, Hecker AL, Herman RH (1980) The influence of exercise and diet on myoglobin and metmyoglobin reductase in the rat. J Lab Clin Med 95 : 222-230
Hagler L, Askew EW, Neville JR, Mellick PW, Coppes RI, Lowder JF (1981) Influence of dietary iron deficiency on hemoglobin, myoglobin, their respective reductases, and skeletal muscle mitochondrial respiration. Am J Clin Nutr 34 : 2169-2177
Hickson RC (1981) Skeletal muscle cytochrome c and myoglobin, endurance, and frequency of training. J Appl Physiol: Respir Environ Exercise Physiol 51:746-749
Holloszy JO, Oscai LB (1979) Effect of exercise on alphaglyce- rophosphate dehydrogenase activity in skeletal muscle. Arch Biochem Biophysic 130:653-656
Jacobs A (1969) Tissue changes in iron deficiency. Br J Haemat 16:1-4
Mathews DK, Fox EL (1976) The physiological basis of physical education and athletics (2ed). WB Saunders Co., Philadel- phia
Ohira Y, Hegenauer J, Strause L, Chert C-S, Saltman P (1982a) Metabolic lesions in iron-deficient rat muscle. In: Saltman P, Hegenauer J (eds) The biochemistry and phsiology of iron. Elsevier/North Holland, New York Oxford Shannon, pp 535-542
Ohira Y, Hegenauer J, Saltman P, Edgerton VR (1982b) Distribution and metabolism of iron in muscles of iron-defi- cient rats. Biol Trace Element Res 4:45-56
Paul KG, Theorelt H, Akeson A (1953) The molar light absorption of pyridine ferroprotoporphyrin (pyridine haemochromogen). Acta Chem Scand 7:1284-1287
Saltin B, Nazar K, Costill DL, Stein E, Jansson E, Essen B, Gollnick PD (1976) The nature of training response; peripheral and central adaptations to one-legged exercise. Acta Physiol Scand 96 : 289-305
Stevenson J, Box B, Feleke V, Beaton J (1966) Bouts of exercise and food intake in the rat. J Appl Physiol 21 : 118-122

R. McDonald et al.: Muscle proteins and endurance training 419
Vatner SF, Higgins CB, Franklin D (1972) Regional circulatory adjustments to moderate and severe chronic anemia in conscious dogs at rest and during exercise. Circ Res 30 : 731-740
Vellar OD, Hermansen L (1972) Physical performance and hematological parameters, with special reference to hemoglo- bin and maximal oxygen uptake. Acta Med Scand Suppl 522 : 1 - 40
Wittenberg J (1970) Myoglobin-facilitated oxygen diffusion: role of myoglobin in oxygen entry into muscle. Physiol Rev 50 : 559-635
Wranne B, Woodson R (1973) A graded treadmill test for rats: maximal work performance in normal and anemic animals. J Appl Physiol 34 : 732-735
Woodson R, Wills R, Lenfant C (1978) Effect of acute and established anemia on 02 transport at rest, submaximal and maximal work. J Appl Physiol: Respir Environ Exercise Physiol 44:36-43
Yoshimura H (1970) Anemia during physical training (Sports anemia). Nutr Rev 28:252-253
Accepted February 28, 1984





























