Embed Size (px)
Citation preview

Naunyn-Schmiedeberg's Arch Pharmacol (1981) 317:214-218
Effects of Hypophyseetomy, Adrenalectomy and (--)-Propranolol on Ethanol-Induced Decrease in Plasma Amino Acids
Tomas Eriksson 1, Tor Magnusson ~, Arvid Carlsson ~, Mats Hagman 2, Rudolf Jagenburg 2, and Staffan Ed6n 3
1 Department of Pharmacology, University of G6teborg, P.O. Box 33031, S-400 33 G6teborg z Department of Clinical Chemistry, Sahlgren's Hospital, University of G6teborg, S-413 45 G6teborg 3 Department of Physiology, University of G6teborg, P.O. Box 33031, S-400 33 G6teborg, Sweden
Naunyn-Schmiedeberg's
Archives of Pharmacology �9 Springer-Verlag 1981
Summary. In previous studies we have demonstrated that an acute dose of ethanol cause an immediate decrease in most plasma amino acids in both man and rat. This effect of ethanol is partly inhibited by the /Ladrenergic antagonist (-)-propranolol, partly by adrenalectomy or hypophysectomy and almost completely by a combination of adrenalectomy with (-)-propranolol. This finding suggests an involvement of both/~-adrenergic mechanisms and steroids from the adrenal cortex in the ethanol-induced decrease in plasma amino acids.
Key words: Ethanol - Plasma amino acids - Adrenalectomy - Hypophysectomy - (-)-Propranolol - Rat
Introduction
The transportation of amino acids, and also of other sub- stances, from plasma into the brain is mediated via several different and specific carrier systems (Pardridge and Oldendorf 1977). One of these is common for the neutral amino acids. Thus threonine, valine, methionine, isoleucine, leucine, tyrosine, phenylalanine, histidine and tryptophan are assumed to be transported into the brain by the same system (Oldendorf 1975) and to compete with each other for the carrier. The relation between the levels of these plasma amino acids, rather than the actual concentration of a certain amino acid, seems to determine how much of each of the different amino acids that will be transported into the brain (Fernstrom and Wurtman 1972; Wurtman and Fernstrom 1976). Factors which influence the concentrations of plasma amino acids might thus be of importance for the availability of various amino acids in the brain.
We have recently found that acutely administered ethanol causes a rapid decrease in almost every plasma amino acid in rat (Eriksson et al. 1980) and in man (to be published). We have also reported that administration of ethanol increases the concentrations in rat brain of exogenously administered I.-dopa (Eriksson et al. 1979), tyrosine, tryptophan, e-methyldopa and 5-hydroxytryptophan (5-HTP) (Eriksson and Carlsson 1980). The cause of this increase in brain concentrations of administered amino acids is suggested to be, at least partly, the ethanol-induced decrease in plasma concentrations of endogenous neutral amino acids, thus favouring the transport of the administered amino acid into the brain.
Send offprint requests to T. Eriksson at the above address
These findings prompted us to investigate how ethanol exerts its plasma amino acid decreasing effect. In this study we have investigated the influence of hypophysectomy, adrenal- ectomy and/3-adrenergic blockade on the plasma amino acid decreasing effect of ethanol.
Methods
Animals. Male Sprague Dawley rats (about 220g) were purchased from Anticimex, Sollentuna, Sweden. Before use the animals were housed for at least one week in a room maintained on a 14/10 h light/dark cycle. They had free access to food and water.
(-)-Propranotot. In one experiment (-)-propranolol, 10mg kg- ~ (1 g 1- a), or an equivalent volume of saline was injected intraperitoneally (i.p.). Thirty minutes later ethanol, 2 g kg- ~ (20 % w/v), or an equivalent volume of saline was injected i.p. Sixty minutes after the last injection the animals were killed by decapitation.
Adrenalectomy and(-)-Propranolol. In a second experiment we investigated the effects of adrenalectomy and a combination of adrenalectomy with (-)-propranolol on the ethanol-induced decrease in plasma concentrations of tyrosine and tryp- tophan. One group of rats was adrenalectomized three days before the experiment. During the time between the operation and the experiment they had free access to food and water and also to a 0.9 % NaC1 solution. (-)-Propranolol, 10mg kg-1 (1 g 1-1), or an equivalent volume of saline was injected i.p. Twenty minutes later ethanol, 2 g kg-1 (20 % w/v), or an equivalent volume of saline was injected i.p. Fourty minutes after the last injection the animals were killed by decapitation.
Analysis of Tyrosine and To~ptophan. In the above mentioned two experiments about 5 ml of blood was collected in a tube containing 0.5 ml of a 1 ~ solution of EDTA. The plasma samples were purified and tyrosine and tryptophan were isolated employing a strong cation exchange column (Dowex 50W-X4) (Atack and Magnusson 1978; Kehr et al. 1972). Spectrophotofluorimetric analysis of tyrosine (Waalkes and Udenfriend 1957) and tryptophan (B6dard et al. 1972) were carried out.
Hypophysectomy. In a third experiment one group of rats was hypophysectomized according to a method previously de- scribed (Smith 1930). The operations were carried out about one week before the experiment. Ethanol, 2 g kg-1 (20~o
0028-1298/81/0317/0214/$01.00
120
IO0
8O
6O
4o
20
Tyrosine
H-
6
120
I- '- * ---I ~ I00
" 80
~ ~ 40
6
Tryptophan
-F
6
I **r - - I
T I---**'-1
I
I
6 5
Soline or Saline or (-)-Propranolol, Ethanol,
I0 mg-kg-' Lp. 2g.kg "1 i.p. Killed I I I 0 30 minutes 90
w/v), or an equivalent volume of saline was injected i.p. In a second control group the rats were not injected at all. Three Ioo L similar groups of rats with intact pituitaries were also investigated. Sixty minutes after the injection the animals w 80- were killed by decapitation and about 5 ml of blood was "~ 6o- collected in a tube containing 0.5 ml of a 1% EDTA solution.
�9 40
20 - Analysis of Amino Acids. In this experiment plasma amino acids were determined by ion exchange chromatography on a Kontron Liquimat III. The peak areas were integrated (Spectra Physics SP 4000). Before analysis 0.1 ml of sulphosa- licylic acid (50 ~ w/v), containing DL-norleucine as internal standard, was added to 1.0ml plasma. The mixture was centrifuged for 2 min at 9000 g and the clear supernatant was stored at room temperature until analysis.
Statistics. Statistical significances were assessed by one way analysis of variance followed by t-test.
Results
215
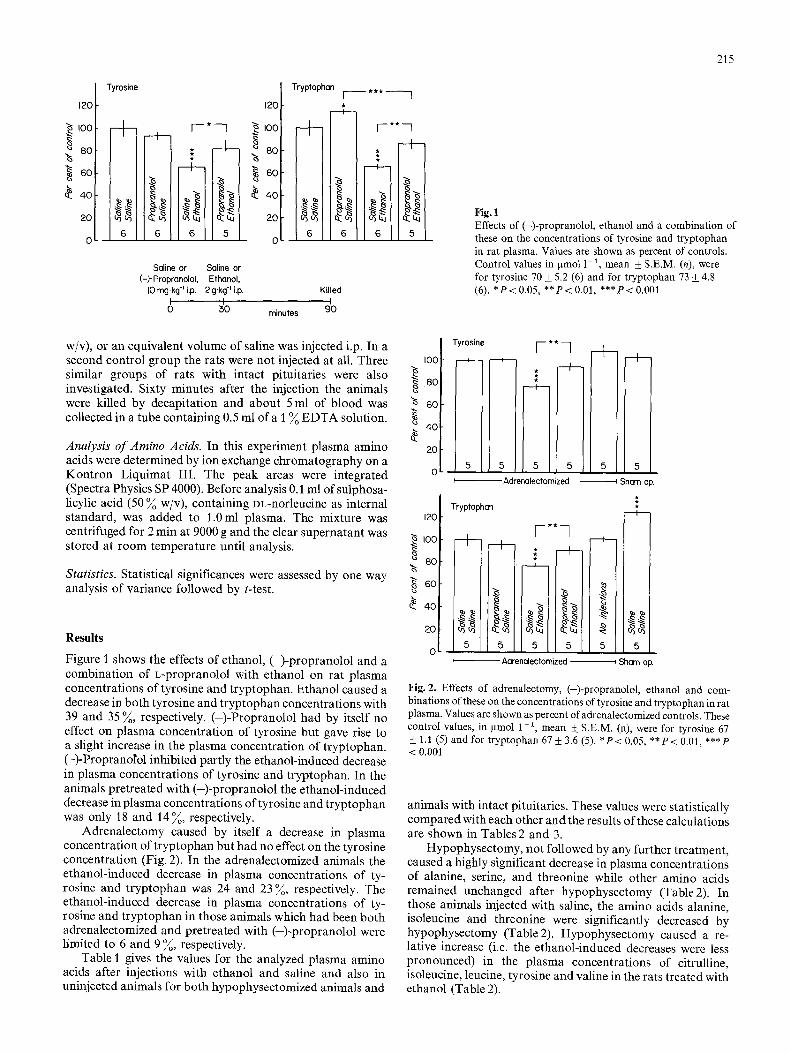
Figure I shows the effects of ethanol, (-)-propranolol and a combination of L-propranolol with ethanol on rat plasma concentrations of tyrosine and tryptophan. Ethanol caused a decrease in both tyrosine and tryptophan concentrations with 39 and 35 ~, respectively. (-)-Propranolol had by itself no effect on plasma concentration of tyrosine but gave rise to a slight increase in the plasma concentration of tryptophan. (-)-Propranolol inhibited partly the ethanol-induced decrease in plasma concentrations of tyrosine and tryptophan. In the animals pretreated with (-)-propranolol the ethanol-induced decrease in plasma concentrations of tyrosine and tryptophan was only 18 and 14%, respectively.
Adrenalectomy caused by itself a decrease in plasma concentration of tryptophan but had no effect on the tyrosine concentration (Fig. 2). In the adrenalectomized animals the ethanol-induced decrease in plasma concentrations of ty- rosine and tryptophan was 24 and 23 ~, respectively. The ethanol-induced decrease in plasma concentrations of ty- rosine and tryptophan in those animals which had been both adrenalectomized and pretreated with (-)-propranolol were limited to 6 and 9 ~, respectively.
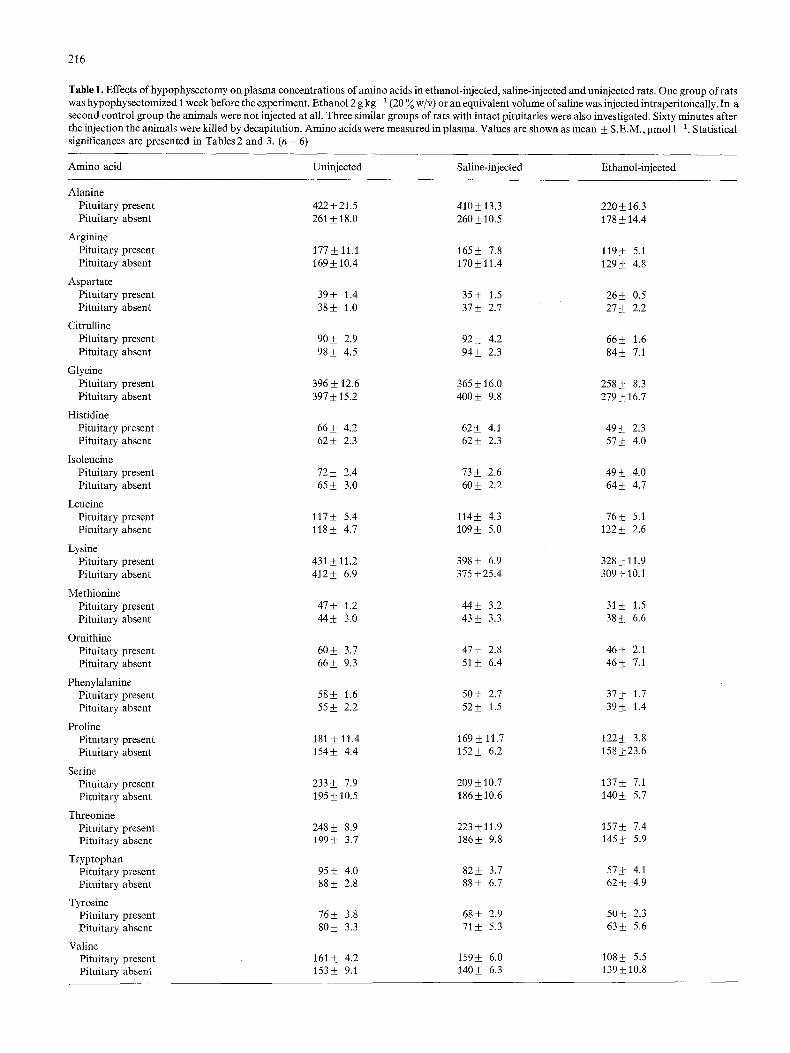
Table 1 gives the values for the analyzed plasma amino acids after injections with ethanol and saline and also in uninjected animals for both hypophysectomized animals and
Fig. 1 Effects of (-)-propranolol, ethanol and a combination of these on the concentrations of tyrosine and t ryptophan in rat plasma. Values are shown as percent of controls. Control values in gmol 1-1, mean + S.E.M. (n), were for tyrosine 70 + 5.2 (6) and for t ryptophan 73 _+ 4.8 (6). * P < 0.05, * * P < 0.01, * * * P < 0.001
0 -
,201 ,oo[
w ao 60
~. 40
2O
O
Tyrosine [-- * *
i I
I
I
5 5 5 5
Adrenelectomized
Tryptophan
I I - * ' 7
.... I
5 5 5 5
Adrenalectomized
I i
5
Sham op.
I
5
t Sham op.
Fig. 2. Effects of adrenalectomy, (-)-propranolol, ethanol and com- binations of these on the concentrations of tyrosine and tryptophan in rat plasma. Values are shown as percent of adrenalectomized controls. These control values, in pmol 1 - 2 mean • S.E.M. (n), were for tyrosine 67 + 1.1 (5) and for t ryptophan 67 + 3.6 (5). * P < 0.05, * * P < 0.01, ***P < 0.001
animals with intact pituitaries. These values were statistically compared with each other and the results of these calculations are shown in Tables 2 and 3.
Hypophysectomy, not followed by any further treatment, caused a highly significant decrease in plasma concentrations of alanine, serine, and threonine while other amino acids remained unchanged after hypophysectomy (Table2). In those animals injected with saline, the amino acids alanine, isoleucine and threonine were significantly decreased by hypophysectomy (Table2). Hypophysectomy caused a re- lative increase (i.e. the ethanol-induced decreases were less pronounced) in the plasma concentrations of citrulline, isoleucine, leucine, tyrosine and valine in the rats treated with ethanol (Table 2).
216
Table 1. Effects of hypophysectomy on plasma concentrations of amino acids in ethanol-injected, saline-injected and uninjected rats. One group of rats was hypophysectomized 1 week before the experiment. Ethanol 2 g kg- 1 (20 ~o w/v) or an equivalent volume of saline was injected intraperitoneally. In a second control group the animals were not injected at all. Three similar groups of rats with intact pituitaries were also investigated. Sixty minutes after the injection the animals were killed by decapitation. Amino acids were measured in plasma. Values are shown as mean • S.E.M., gmol l- 1. Statistical significances are presented in Tables 2 and 3. (n = 6)
Amino acid Uninjected Saline-injected Ethanol-injected
Alanine Pituitary present 422 + 21.5 410 + 13.3 220 _ 16.3 Pituitary absent 261 + 18.0 260 • 10.5 178 4-14.4
Arginine Pituitary present 177 + 11.1 165• 7.8 119_+ 5.1 Pituitary absent 169 • 10.4 170 • 11.4 129 + 4.8
Aspartate Pituitary present 39+ 1.4 35• 1.5 26• 0.5 Pituitary absent 38• 1.0 37• 2.7 27• 2.2
Citrulline Pituitary present 90• 2.9 92• 4.2 66• 1.6 Pituitary absent 98• 4.5 94__ 2.3 84+ 7.1
Glycine Pituitary present 396 • 12.6 365 • 16.0 258___ 8.3 Pituitary absent 397 • 15.2 400 _+ 9.8 279 • 16.7
Histidine Pituitary present 66• 4.2 62• 4.1 49• 2.3 Pituitary absent 62• 2.3 62• 2.3 57_ 4.0
Isoleucine Pituitary present 72 • 2.4 73 • 2.6 49 + 4.0 Pituitary absent 65• 3.0 60• 2.2 64• 4.7
Leucine Pituitary present 117• 5.4 114• 4.3 76_+ 5.1 Pituitary absent 118• 4.7 109• 5.0 122• 2.6
Lysine Pituitary present 431 • 11.2 398 _+ 6.9 328 + 11.9 Pituitary absent 4l 2 • 6.9 375 • 25.4 309 • 10.1
Methionine Pituitary present 47• 1.2 44• 3.2 31• 1.5 Pituitary absent 44• 3.0 43• 3.3 38+ 6.6
Ornithine Pituitary present 60• 3.7 47• 2.8 46+ 2.1 Pituitary absent 66• 9.3 51• 6.4 46-+ 7.1
Phenylalanine Pituitary present 58• 1.6 50• 2.7 37• 1.7 Pituitary absent 55• 2.2 52-+ 1.5 39• 1.4
Proline Pituitary present 181 • 11.4 169 • 11.7 122• 3.8 Pituitary absent 154• 4.4 152• 6.2 158_+23.6
Serine Pituitary present 233 • 7.9 209 • 10.7 137 • 7.1 Pituitary absent 195 • 10.5 186• 10.6 140_+ 5.7
Threonine Pituitary present 248 • 8.9 223-+ 11.9 157• 7.4 Pituitary absent 199_+ 3.7 186+ 9.8 145• 5.9
Tryptophan Pituitary present 95+ 4.0 82_+ 3.7 57+ 4.1 Pituitary absent 88• 2.8 88• 6.7 62_+ 4.9
Tyrosine Pituitary present 76• 3.8 68• 2.9 50• 2.3 Pituitary absent 80• 3.3 71 • 5.3 63_+ 5.6
Valine Pituitary present 161• 4.2 159_+ 6.0 108_+ 5.5 Pituitary absent 153• 9.1 140• 6.3 139• 10.8
217
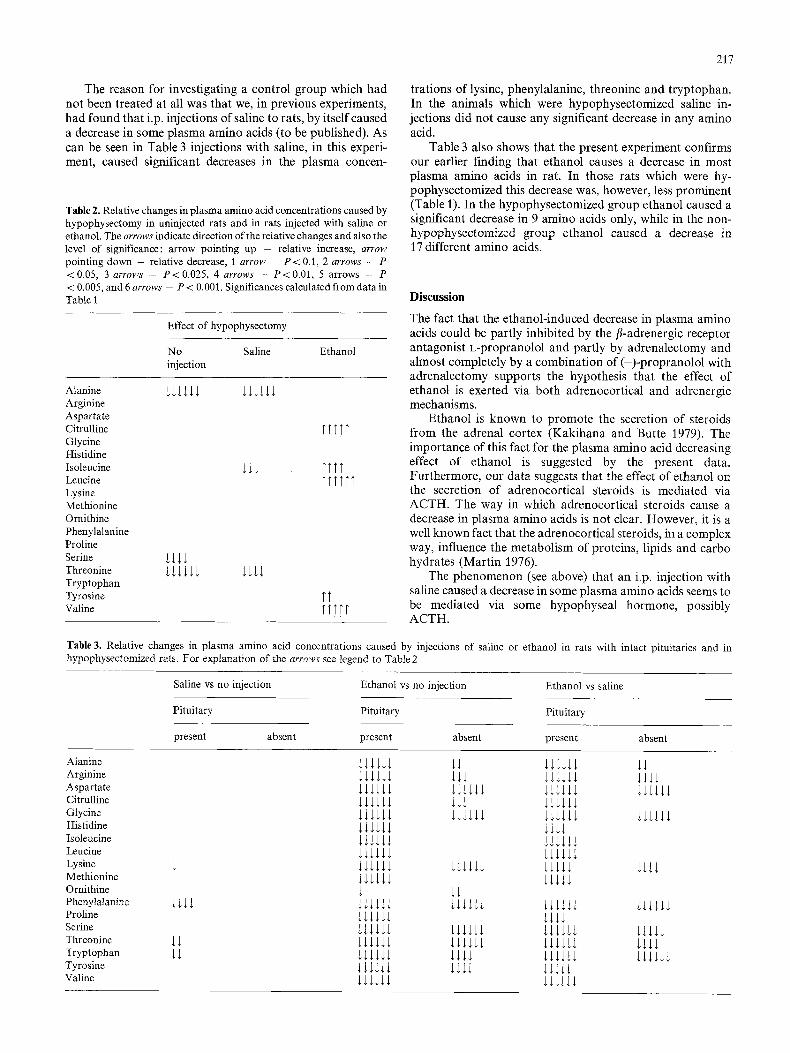
The reason for investigating a control group which had not been treated at all was that we, in previous experiments, had found that i.p. injections of saline to rats, by itself caused a decrease in some plasma amino acids (to be published). As can be seen in Table 3 injections with saline, in this experi- ment, caused significant decreases in the plasma concen-
Table 2. Relative changes in plasma amino acid concentrations caused by hypophysectomy in uninjected rats and in rats injected with saline or ethanol. The a r r o w s indicate direction of the relative changes and also the level of significance: arrow pointing up - relative increase, a r r o w
pointing down -- relative decrease, t a r r o w - P < 0.1, 2 a r r o w s - P
<0.05, 3 a r r o w s - P<0.025, 4 a r r o w s - P<0.01, 5 arrows - P < 0.005, and 6 a r r o w s - P < 0.001. Significances calculated from data in Table I
Effect of hypophysectomy
No Saline Ethanol injection
Arginine Aspartate Citrulline Glycine Histidine Isoleucine J, ~ Leucine Lysine Methionine Ornithine Phenylalanine Proline Serine ~ ~ J, ; Threonine J, ~ ~, ], ~ ~ J, ~ ], Tryptophan Tyrosine Valine
tt~Tt
TTTT TTTTTT
t t tT t [ t
trations of lysine, phenylalanine, threonine and tryptophan. In the animals which were hypophysectomized saline in- jections did not cause any significant decrease in any amino acid.
Table 3 also shows that the present experiment confirms our earlier finding that ethanol causes a decrease in most plasma amino acids in rat. In those rats which were hy- pophysectomized this decrease was, however, less prominent (Table 1). In the hypophysectomized group ethanol caused a significant decrease in 9 amino acids only, while in the non- hypophysectomized group ethanol caused a decrease in 17 different amino acids.
Discussion
The fact that the ethanol-induced decrease in plasma amino acids could be part ly inhibited by the/3-adrenergic receptor antagonist L-propranolol and part ly by adrenalectomy and almost completely by a combinat ion of ( - ) -propranolo l with adrenalectomy supports the hypothesis that the effect of ethanol is exerted via both adrenocortical and adrenergic mechanisms.
Ethanol is known to promote the secretion of steroids from the adrenal cortex (Kakihana and Butte 1979). The importance of this fact for the plasma amino acid decreasing effect of ethanol is suggested by the present data. Furthermore, our data suggests that the effect of ethanol on the secretion of adrenocortical Steroids is mediated via ACTH. The way in which adrenocortical steroids cause a decrease in plasma amino acids is not clear. However, it is a well known fact that the adrenocortical steroids, in a complex way, influence the metabolism of proteins, lipids and carbo hydrates (Mart in 1976).
The phenomenon (see above) that an i.p. injection with saline caused a decrease in some plasma amino acids seems to be mediated via some hypophyseal hormone, possibly ACTH.
Table 3. Relative changes in plasma amino acid concentrations caused by injections of saline or ethanol in rats with intact pituitaries and in hypophysectomized rats. For explanation of the a r r o w s see legend to Table 2
Saline vs no injection Ethanol vs no injection Ethanol vs saline
Pituitary Pituitary Pituitary
present absent present absent present absent
Alanine Arginine Aspartate Citrulline Glycine Histidine Isoleucine Leucine Lysine Methionine Ornithine Phenylalanine Proline Serine Threonine Tryptophan Tyrosine Valine

218
High doses o f e thanol have been repor ted to increase the secret ion o f adrenal ine and noradrena l ine f rom the adrenal medul la (Kak ihana and Butte 1979). I t is possible that such an increase in circulat ing ca techolamines is involved in the e thanol - induced decrease in p lasma amino acids, thus ex- plaining the inhibi tory effect o f ( - ) -p rop rano lo l on the amino acid decreasing effect o f ethanol . The way in which this adrenergic effect is exer ted is howeve r more obscure. One possibil i ty could be that the increased b lood f low th rough the liver, caused by s t imula t ion of f l -adrenergic receptors (Greenway and Stark 1971) gives rise to an increase in amino acid me tabo l i sm in the liver and secondari ly to a decrease in p lasma amino acids
Acknowledgements. This work was supported by the Swedish Medical Research Council (project No. 155), the Faculty of Medicine at the University of G6teborg and the G6teborg Medical Society.
We want to express our sincere gratitude to Mrs Birgitta Holmgren for her expert technical assistance.
References
Atack A, Maguusson T (1978) A procedure for the isolation of nor- adrenaline (together with adrenaline), dopamine, 5-hydroxytrypt- amine and histamine from the same tissue sample using a single column of strongly acidic cation exchange resin. Acta Pharmacol Toxicol 42: 35 - 57
B6dard P, Carlsson A, Lindqvist M (1972) Effect of transverse cerebral hemisection on 5-hydroxytryptamine metabolism in the rat brain. Naunyn-Schmiedeberg's Arch Pharmacol 272:1-15
Eriksson T, Carlsson A (1980) Ethanol-induced increase in brain concentrations of administered neutral amino acids. Naunyn- Schmiedeberg's Arch Pharmacol 314: 4 7 - 50
Eriksson T, Liljequist S, Carlsson A (1979) Ethanol-induced increase in the penetration of exogenously administered L-dopa through the blood-brain barrier. J Pharm Pharmacol 31:636- 637
Eriksson T, Carlsson A, Liljequist S, Hagman M, Jagenburg R (1980) Decrease in plasma amino acids in rat after acute administration of ethanol. J Pharm Pharmacol 32: 512 - 513
Fernstrom JD, Wurtman RJ (1972) Brain serotonin content: physiologi- cal regulation by plasma neutral amino acids. Science 178 : 414- 416
Greenway CV, Stark RD (1971) Hepatic vascular bed. Physiol Rev 51:23-65
Kakihana R, Butte JC (1979) Ethanol and endocrine function. In: Majchrowicz E, Noble EP (eds) Biochemistry and pharmacology of ethanol, vo12. Plenum Press, New York, pp 147-164
Kehr W, Carlsson A, Lindqvist M (1972) A method for the determination of 3,4-dihydroxyphenylalanine (DOPA) in brain. Naunyn- Schmiedeberg's Arch Pharmacol 274:273 - 280
Martin CR (1976) Textbook of endocrine physiology. Williams and Wilkins Co., Baltimore
OldendorfWH (1975) Permeability of the blood-brain barrier. In: Tower DB (ed) The Nervous system, vol 1 : the basic neurosciences. Raven Press, New York, pp 279-289
Pardridge WM, Oldendorf WH (1977) Transport of metabolic substrates through the blood-brain barrier. J Neurochem 28:5-12
Smith PE (1930) Hypophysectomy and a replacement therapy in the rat Am J Anat 45 : 205- 273
Waalkes TP, Udenfriend S (1957) A fluorimetric method for the estimation of tyrosine in plasma and tissues. J Lab Clin Med 50:733-736
Wurtman RJ, Fernstrom JD (1976) Control of brain neurotransmitter synthesis by precursor availability and nutritional state. Biochem Pharmacol 25 : 1691 - 1696
Received March 26/Accepted June 16, 1981


























![Characterization of propranolol-resistant (-)-[125I]-cyanopindolol](https://img.pdfslide.us/doc/110x75/58668a461a28ab2c408b6e44/characterization-of-propranolol-resistant-125i-cyanopindolol-.jpg)


